
Macrobenthos of the Tortolì Lagoon: A Peculiar Case of High Benthic Biodiversity among Mediterranean Lagoons


Abstract
1. Introduction
2. Materials and Methods
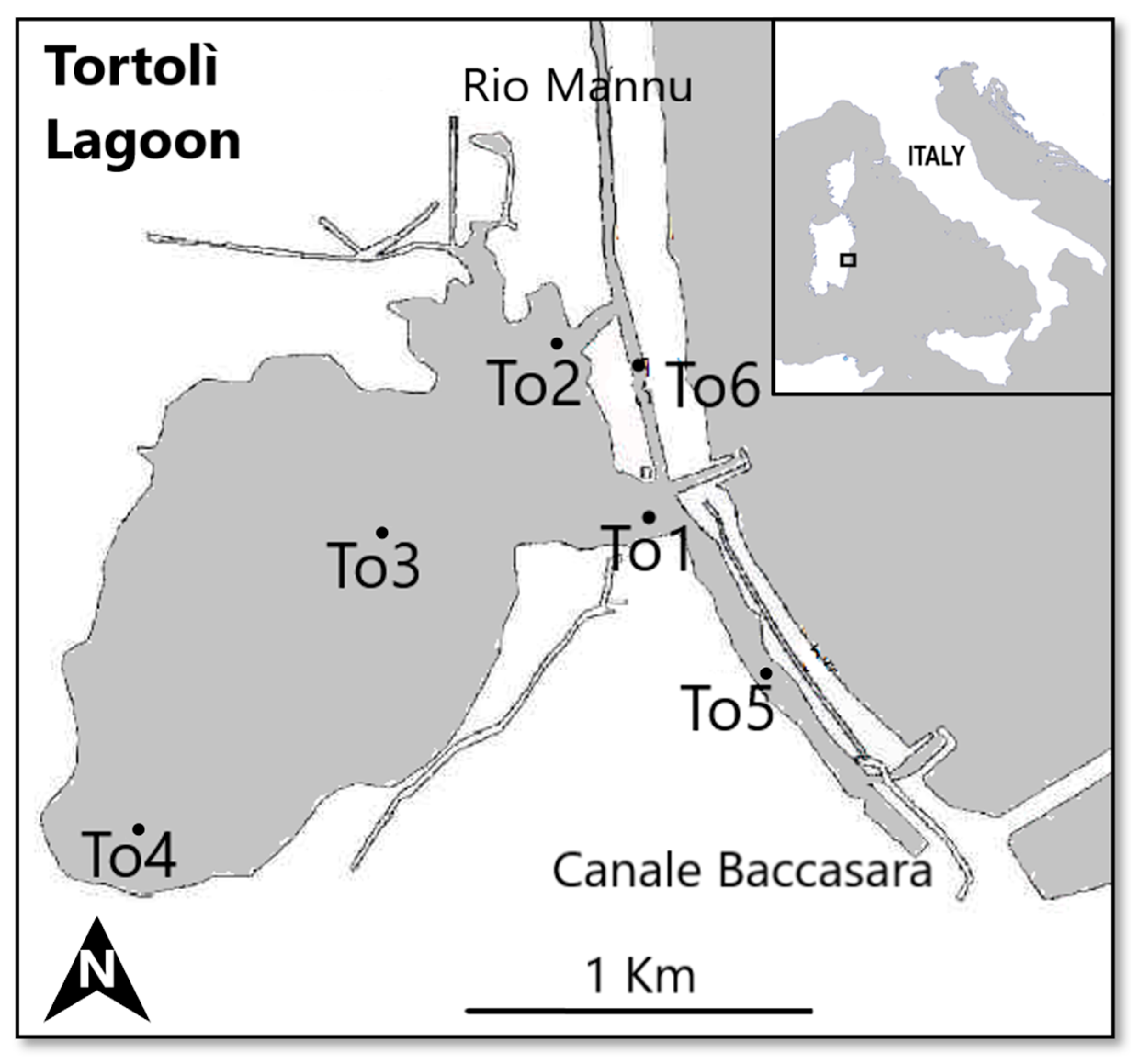
2.1. Study Area
2.2. Sampling Methods and Taxonomical Analysis
2.3. Data Analysis
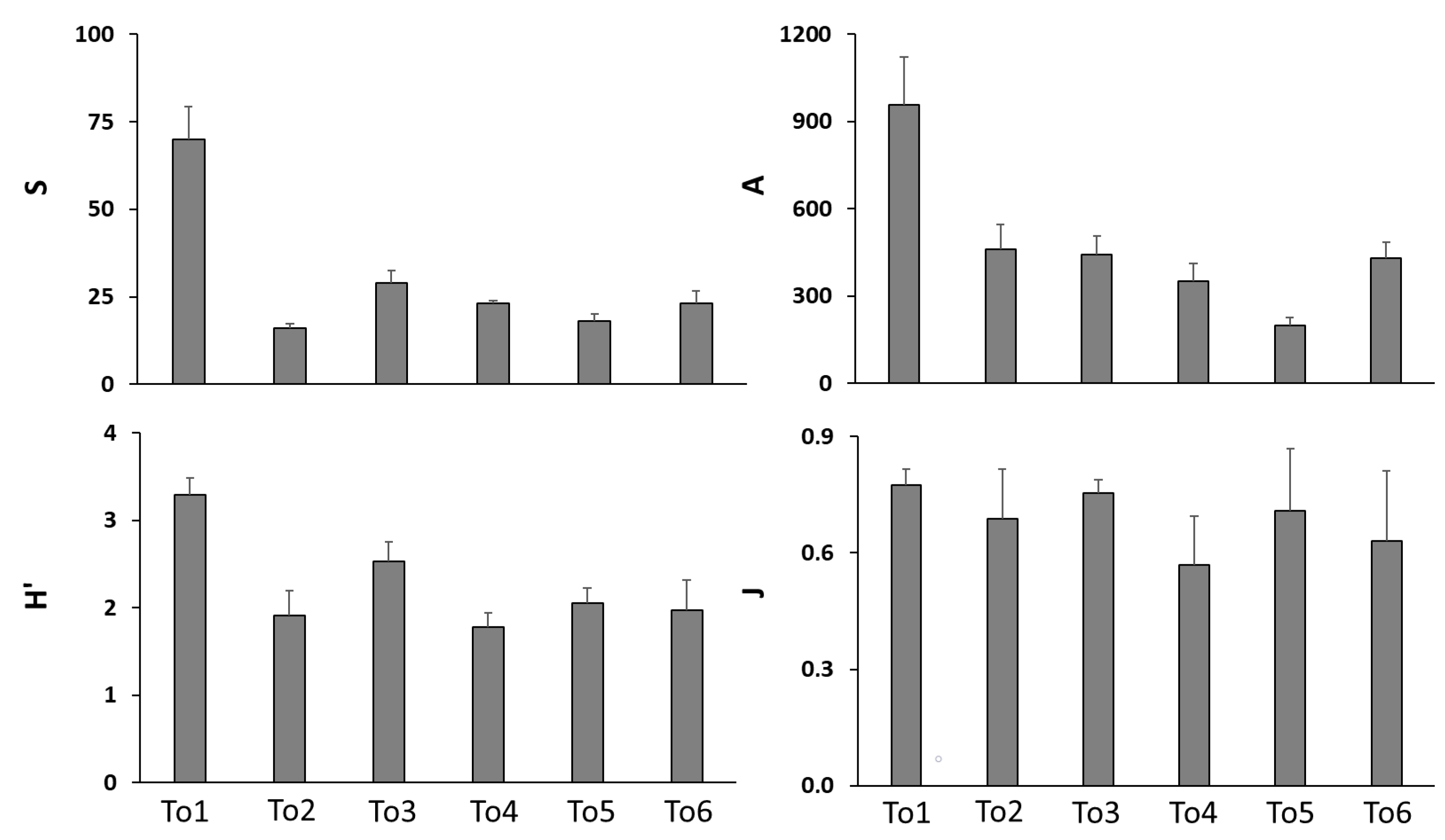
3. Results
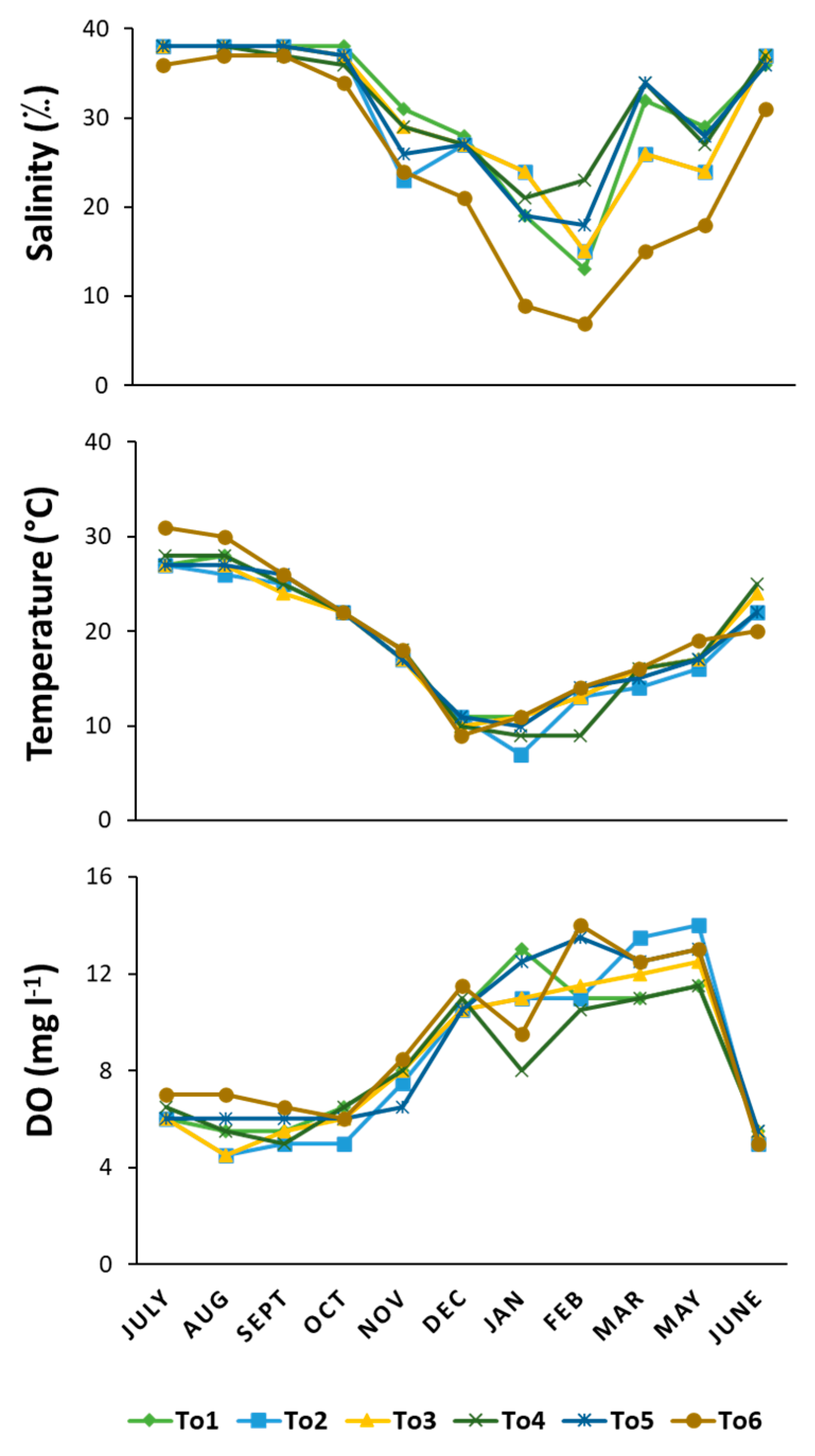
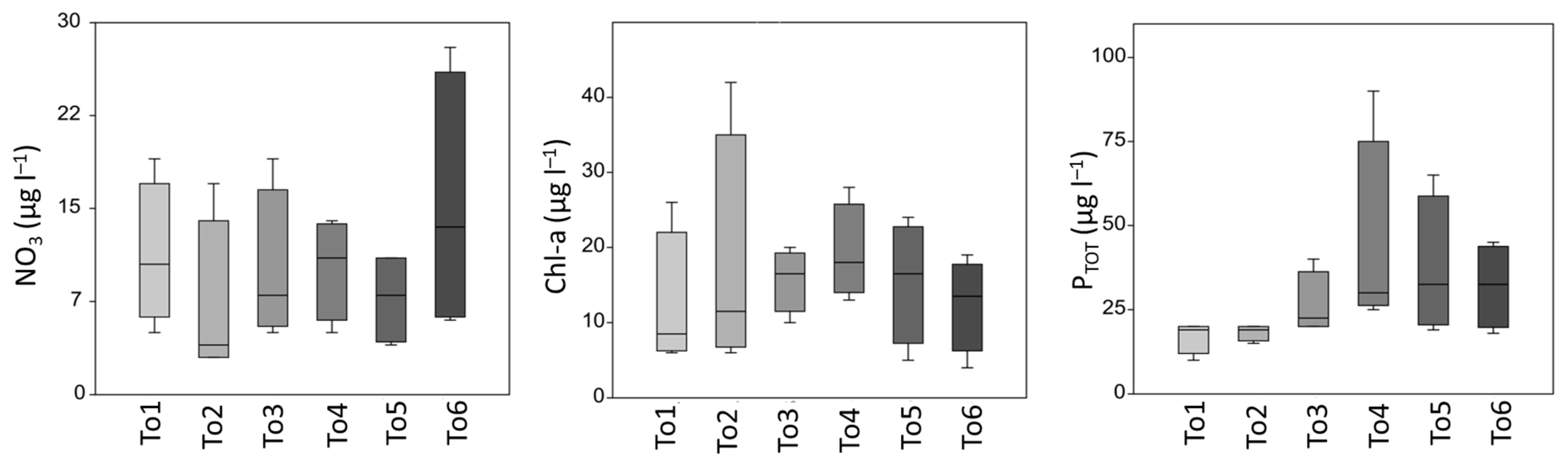
3.1. Water Variables
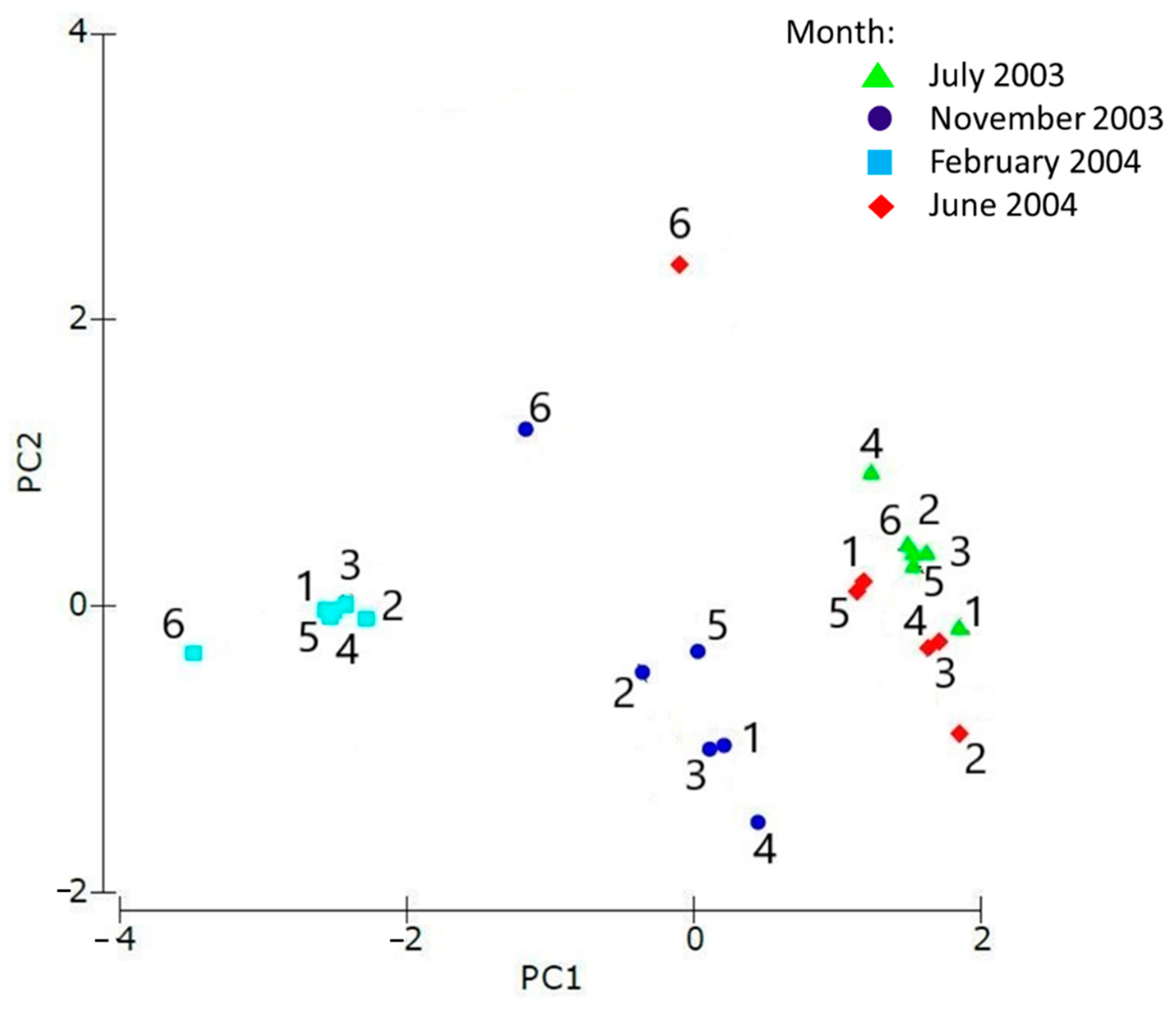
3.2. Macrozoobenthic Community
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Graham, P.; Falconer, L.; Telfer, T.; Mossone, P.; Viale, I.; Carboni, S. A modelling approach to classify the suitability of shallow Mediterranean lagoons for pacific oyster, Crassostrea gigas (Thunberg, 1793) farming. Ocean Coast. Manag. 2020, 192, 105234. [Google Scholar] [CrossRef]
- Vallainc, D.; Concu, D.; Papiol, G.G.; Loi, B.; Leggieri, F.; Brundu, G.; Chindris, A.; Sanna, G.; Fois, N.; Antognarelli, F.; et al. Producing flat-head grey mullet Mugil cephalus (Linnaeus, 1758) fries in captivity from sexually mature adults collected in Sardinian lagoons. Aquac. Rep. 2021, 21, 100844. [Google Scholar] [CrossRef]
- Magni, P.; Gravina, M.F. Macrobenthos of lagoon ecosystems: A comparison in vegetated and bare sediments. Adv. Oceanogr. Limnol. 2023, 14. [Google Scholar] [CrossRef]
- Pérez-Ruzafa, A.; Marcos, C.; Pérez-Ruzafa, I.M.; Pérez-Marcos, M. Coastal lagoons: “transitional ecosystems” between transitional and coastal waters. J. Coast. Conserv. 2011, 15, 369–392. [Google Scholar] [CrossRef]
- Colombo, G. Lagoons. In The Coastline; Barnes, R.S.K., Ed.; Wiley: London, UK; New York, NY, USA, 1977; pp. 63–81. [Google Scholar]
- Barnes, R.S.K. Coastal Lagoons; University Press Cambridge: Cambridge, UK, 1980; pp. 1–106. [Google Scholar]
- Sacchi, C. Les formes biologiques animals comme indicatrices du milieu: Considerations générales et exemples pratique. Rend. Sem. Fac. Sci. Univ. Cagliari 1984, 54, 303–318. [Google Scholar]
- Cognetti, G.; Maltagliati, F. Perspectives on the ecological assessment of transitional waters. Mar. Pollut. Bull. 2008, 56, 607–608. [Google Scholar] [CrossRef]
- Magni, P.; Tagliapietra, D.; Lardicci, C.; Balthis, L.; Castelli, A.; Como, S.; Frangipane, G.; Giordani, G.; Hyland, J.; Maltagliati, F.; et al. Animal-sediment relationships: Evaluating the “Pearson–Rosenberg paradigm” in Mediterranean coastal lagoons. Mar. Pollut. Bull. 2009, 58, 478–486. [Google Scholar] [CrossRef]
- Magni, P.; Como, S.; Kamijo, A.; Montani, S. Effects of Zostera marina on the patterns of spatial distribution of sediments and macrozoobenthos in the boreal lagoon of Furen (Hokkaido, Japan). Mar. Environ. Res. 2017, 131, 90–102. [Google Scholar] [CrossRef]
- Tagliapietra, D.; Pessa, G.; Cornello, M.; Zitelli, A.; Magni, P. Temporal distribution of intertidal macrozoobenthic assemblages in a Nanozostera noltii dominated area (Lagoon of Venice). Mar. Environ. Res. 2016, 114, 31–39. [Google Scholar] [CrossRef]
- Gravina, M.F.; Cabiddu, S.; Como, S.; Floris, A.; Padedda, B.M.; Pusceddu, A.; Magni, P. Disentangling heterogeneity and commonalities in nanotidal Mediterranean lagoons through environmental features and macrozoobenthic assemblages. Estuar. Coast. Shelf. Sci. 2020, 237, 106688. [Google Scholar] [CrossRef]
- Brundu, G.; Magni, P. Context-dependent effect of serpulid reefs on the variability of soft-bottom macrobenthic assemblages in three Mediterranean lagoons (Sardinia, Italy). Estuarine. Coast. Shelf Sci. 2021, 262, 107589. [Google Scholar] [CrossRef]
- De Falco, G.; Magni, P.; Teräsvuori, L.M.H.; Matteucci, G. Sediment grain size and organic carbon distribution in the Cabras lagoon (Sardinia, western Mediterranean). Chem. Ecol. 2004, 20, 367–377. [Google Scholar] [CrossRef]
- Como, S.; Magni, P.; Casu, D.; Floris, A.; Giordani, G.; Natale, S.; Fenzi, G.A.; Signa, G.; De Falco, G. Sediment characteristics and macrofauna distribution along a human-modified inlet in the Gulf of Oristano (Sardinia, Italy). Mar. Pollut. Bull. 2007, 54, 733–744. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Gosselink, J.G. The value of wetlands: Importance of scale and landscape setting. Ecol. Econ. 2000, 35, 25–33. [Google Scholar] [CrossRef]
- Levin, L.A.; Boesch, D.F.; Covich, A.; Dahm, C.; Erseus, C.; Ewel, K.C.; Kneib, R.T.; Moldenke, A.; Palmer, M.A.; Snelgrove, P.; et al. The function of marine critical transition zones and the importance of sediment biodiversity. Ecosystems 2001, 4, 430–451. [Google Scholar] [CrossRef]
- Pérez-Ruzafa, A.; Marcos, C.; Pérez-Ruzafa, I.M. Mediterranean coastal lagoons in an ecosystem and aquatic resources management context. Phys. Chem. Earth Parts A/B/C 2011, 36, 160–166. [Google Scholar] [CrossRef]
- Obiefuna, J.N.; Nwilo, P.C.; Atagbaza, A.O.; Okolie, C.J. Land cover dynamics associated with the spatial changes in the wetlands of Lagos/Lekki Lagoon system of Lagos, Nigeria. J. Coast. Res. 2013, 29, 671–679. [Google Scholar]
- Newton, A.; Icely, J.; Cristina, S.; Brito, A.; Cardoso, A.C.; Colijn, F.; Dalla Riva, S.; Gertz, F.; Hansen, J.W.; Holmer, M. An overview of ecological status, vulnerability and future perspectives of European large shallow, semi-enclosed coastal systems, lagoons and transitional waters. Estuar. Coast. Shelf Sci. 2014, 140, 95–122. [Google Scholar] [CrossRef]
- Crooks, S.; Turner, R.K. Integrated coastal management: Sustaining estuarine natural resources. Adv. Ecol. Res. 1999, 29, 241–289. [Google Scholar]
- Magni, P.; Micheletti, S.; Casu, D.; Floris, A.; Giordani, G.; Petrov, A.N.; De Falco, G.; Castelli, A. Relationship between chemical characteristics of sedimentsband macrofaunal communities in the Cabras lagoon (Western Mediterranean, Italy). Hydrobiologia 2005, 550, 105.e119. [Google Scholar] [CrossRef]
- Panigrahi, S.; Acharya, B.C.; Panigrahy, R.C.; Nayak, B.K.; Banarjee, K.; Sarkar, S.K. Anthropogenic impact on water quality of Chilika lagoon RAMSAR site: A statistical approach. Wetl. Ecol. Manag. 2007, 15, 113–126. [Google Scholar] [CrossRef]
- Kolios, S.; Stylios, C.D. Identification of land cover/land use changes in the greater area of the Preveza peninsula in Greece using Landsat satellite data. Appl. Geogr. 2013, 40, 150–160. [Google Scholar] [CrossRef]
- Khedhri, I.; Afli, A.; Aleya, L. Structuring factors of the spatio-temporal variability of macrozoobenthos assemblages in a southern Mediterranean lagoon: How useful for bioindication is a multi-biotic indices approach? Mar. Pollut. Bull. 2017, 114, 515–527. [Google Scholar] [CrossRef]
- Semprucci, F.; Gravina, M.F.; Magni, P. Meiofaunal Dynamics and Heterogeneity along Salinity and Trophic Gradients in a Mediterranean Transitional System. Water 2019, 11, 1488. [Google Scholar] [CrossRef]
- Gravina, M.F.; Bonifazi, A.; Del Pasqua, M.; Giampaoletti, J.; Lezzi, M.; Ventura, D.; Giangrande, A. Perception of changes in marine benthic habitats: The relevance of taxonomic and ecological memory. Diversity 2020, 12, 480. [Google Scholar] [CrossRef]
- Boero, F. Episodic events: Their relevance to ecology and evolution. Mar. Ecol. 1996, 17, 237–250. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Boero, F.; Fonda Umani, S.; Morri, C.; Vacchi, M. Successione e cambiamento negli ecosistemi marini. Biol. Mar. Medit. 1998, 5, 117–135. [Google Scholar]
- Muxika, I.; Borja, A.; Bald, J. Using historical data, expert judgement and multivariate analysis in assessing reference conditions and benthic ecological status, according to the European Water Framework Directive. Mar. Pollut. Bull. 2007, 55, 16–29. [Google Scholar] [CrossRef] [PubMed]
- Pinnegar, J.K.; Engelhard, G.H. The “shifting baseline” phenomenon: A global perspective. Rev. Fish Biol. 2008, 18, 1–16. [Google Scholar] [CrossRef]
- Papworth, S.K.; Rist, J.; Coad, L.; Milner-Gulland, E.J. Evidence for shifting baseline syndrome in conservation. Conserv. Lett. 2009, 2, 93–100. [Google Scholar] [CrossRef]
- Balaguer, L.; Escudero, A.; Martín-Duque, J.F.; Mola, I.; Aronson, J. The historical reference in restoration ecology: Re-defining a cornerstone concept. Biol. Conserv. 2014, 176, 12–20. [Google Scholar] [CrossRef]
- Soga, M.; Gaston, K.J. Shifting baseline syndrome: Causes, consequences, and implications. Front. Ecol. Environ. 2018, 16, 222–230. [Google Scholar] [CrossRef]
- Magni, P. Biological benthic tools as indicators of coastal marine ecosystems health. Chem. Ecol. 2003, 19, 363–372. [Google Scholar] [CrossRef]
- Nonnis Marzano, C.; Scalera Liaci, L.; Fianchini, A.; Gravina, M.F.; Mercurio, M.; Corriero, G. Distribution, persistence and change in the macrobenthos of the lagoon of Lesina (Apulia, southern Adriatic Sea). Oceanol. Acta 2003, 26, 57–66. [Google Scholar] [CrossRef]
- Basset, A.; Sangiorgio, F.; Sabetta, L. (Curatori) Nuovi approcci metodologici per la classificazione dello stato di qualità degli ecosistemi acquatici di transizione—Metodologie per la determinazione della struttura dimensionale di fitoplancton e macroinvertebrati bentonici. In Manuali e Linee Guida ISPRA; ISPRA: Rome, Italy; Università del Salento: Lecce, Italy, 2009. [Google Scholar]
- Giangrande, A.; Gravina, M.F. Brackish-water polychaetes, good descriptors of environmental changes in space and time. Transit. Waters Bull. 2015, 9, 42–55. [Google Scholar]
- Salati, F.; Meloni, A.; Marongiu, E.; Kusuda, R. Microbiological study on the flora of mussel, Mytilus galloprovincialis, cultured in Tortoli Lagoon, South-East Sardinia, Italy. Fish. Sci. 1999, 65, 657–658. [Google Scholar] [CrossRef]
- Parson, T.; Maita, T.; Lalli, M. (Eds.) A Manual of Chemical and Biological Methods for Seawater Analysis; Pergamon Press: Oxford, UK, 1984; p. 173. [Google Scholar]
- Clarke, K.R.; Warwick, R.M. (Eds.) Changes in Marine Communities: An Approach to Statistical Analysis and Interpretation; Primer-E Ltd.: Plymouth, UK, 2001; p. 172. [Google Scholar]
- Clarke, K.R. Non-parametric multivariate analyses of change of community structure. Austral Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Tagliapietra, D.; Sigovini, M.; Magni, P. Saprobity: A unified view of benthic succession models for coastal lagoons. Hydrobiologia 2012, 686, 15–28. [Google Scholar] [CrossRef]
- Basset, A.; Sabetta, L.; Galuppo, N. Environmental heterogeneity and benthic macroinvertebrate guilds in Italian lagoons. Transit. Waters Bull. 2007, 1, 48–63. [Google Scholar]
- Basset, A.; Pinna, M.; Sabetta, L.; Barbone, E.; Galuppo, N. Hierarchical scaling of biodiversity in lagoon ecosystems. Transit. Waters Bull. 2008, 3, 75–86. [Google Scholar]
- Magni, P.; Draredja, B.; Melouah, K.; Como, S. Patterns of seasonal variation in lagoonal macrozoobenthic assemblages (Mellah lagoon, Algeria). Mar. Environ. Res. 2015, 109, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Cognetti Varriale, A.M. Su due policheti Owenidae di sabbie infralitorali del Golfo di Follonica. Atti Soc. Tosc. Sci. Nat. Mem. Ser. B. 1979, 86, 263–267. [Google Scholar]
- Bhaud, M.R. Identification of adults and larvae in Spiochaetopterus (Polychaeta, Chaetopteridae): Consequences for larval transport and recruitment. Hydrobiologia 2003, 496, 279–287. [Google Scholar] [CrossRef]
- Magni, P.; Como, S.; Cucco, A.; De Falco, G.; Domenici, P.; Ghezzo, M.; Lefrançois, C.; Simeone, S.; Perilli, A. A multidisciplinary and ecosystemic approach in the Oristano Lagoon-Gulf system (Sardinia, Italy) as a tool in management plans. Transit. Waters Bull. 2008, 2, 41–62. [Google Scholar]
- Como, S.; Floris, A.; Pais, A.; Rumolo, P.; Saba, S.; Sprovieri, M.; Magni, P. Assessing the impact of the Asian mussel Arcuatula senhousia in the recently invaded Oristano Lagoon-Gulf system (W Sardinia, Italy). Estuar. Coast. Shelf Sci. 2018, 201, 123–131. [Google Scholar] [CrossRef]
- Cardone, F.; Corriero, G.; Fianchini, A.; Gravina, M.F.; Nonnis Marzano, C. Biodiversity of transitional waters: Species composition and comparative analysis of hard bottom communities from the south-eastern Italian coast. J. Mar. Biol. Assoc. U. K. 2014, 94, 25–34. [Google Scholar] [CrossRef]
- Rossi, R.; Cannas, A. Gli stagni sardi dalla pesca tradizionale all’uso delle nuove tecnologie. Oebalia 1992, 17, 1–10. [Google Scholar]
- Cannas, A.; Trisolini, R.; Rossi, R. La pesca nello stagno di Tortolì (Sardegna). Oebalia 1992, 17, 127–128. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Factors | Total Sum of Squares | df | Mean Square | Pseudo-F | p |
|---|---|---|---|---|---|---|
| Species | Sites | 1.31 | 5 | 0.26 | 8.96 | 0.0001 |
| Months | 0.49 | 3 | 0.16 | 5.66 | 0.0006 | |
| Sites × Months | 1.9 | 15 | 0.13 | 4.34 | 0.0001 | |
| Residual | 0.7 | 24 | 0.03 | |||
| Total | 4.4 | 47 | ||||
| Abundance | Sites | 0.78 | 5 | 0.16 | 3.86 | 0.0008 |
| Months | 3.45 | 3 | 1.15 | 28.49 | 0.0001 | |
| Sites × Months | 4.29 | 15 | 0.29 | 7.07 | 0.0001 | |
| Residual | 0.97 | 24 | 0.04 | |||
| Total | 9.49 | 47 | ||||
| Diversity | Sites | 1.1 | 5 | 0.22 | 3.57 | 0.0011 |
| Months | 0.42 | 3 | 0.14 | 2.27 | 0.0249 | |
| Sites × Months | 1.83 | 15 | 0.12 | 1.98 | 0.02 | |
| Residual | 1.48 | 24 | 0.06 | |||
| Total | 4.83 | 47 | ||||
| Evenness | Sites | 0.38 | 5 | 0.08 | 7.28 | 0.001 |
| Months | 0.53 | 3 | 0.18 | 17.15 | 0.0001 | |
| Sites × Months | 0.75 | 15 | 0.05 | 4.81 | 0.0024 | |
| Residual | 0.25 | 24 | 0.01 | |||
| Total | 1.9 | 47 |
| Species | Higher Taxon | Ecological Affinity | Species Occur. | Contrib. % | To1 | To2 | To3 | To4 | To5 | To6 |
|---|---|---|---|---|---|---|---|---|---|---|
| Abra segmentum (Récluz, 1843) | B | Cs | 6 | 5.19 |  | | | | | |
| Apocorophium acutum (Chevreux, 1908) | C | Mr | 6 | 4.22 | | | | | | |
| Cerastoderma glaucum (Bruguière, 1789) | B | Br | 6 | 2.63 | | | | | | |
| Heteromastus filiformis (Claparède, 1864) | P | Op | 6 | 2.21 | | | | | | |
| Capitella capitata (Fabricius, 1780) | P | Op | 5 | 7.69 | | | | | | |
| Microdeutopus gryllotalpa Costa, 1853 | C | Cs | 5 | 3.60 | | | | | | |
| Monocorophium insidiosum (Crawford, 1937) | C | Br | 5 | 13.27 | | | | | | |
| Nephtys hombergii Savigny in Lamarck, 1818 | P | Mr | 5 | 4.51 | | | | | | |
| Phoronis psammophila Cori, 1889 | Ph | Mr | 5 | 0.86 | | | | | | |
| Prionospio cirrifera Wirén, 1883 | P | Op | 5 | 0.63 | | | | | | |
| Iphinoe trispinosa (Goodsir, 1843) | C | Mr | 4 | 0.30 | | | | | ||
| Melita palmata (Montagu, 1804) | C | Cs | 4 | 0.57 | | | | | ||
| Chironomidae | Ch | Op | 3 | 4.59 | | | | |||
| Venerupis corrugata (Gmelin, 1791) | B | Mr | 3 | 1.68 | | | | |||
| Aphelochaeta marioni (Saint-Joseph, 1894) | P | Mr | 2 | 1.05 | | | ||||
| Chaetozone setosa Malmgren, 1867 | P | Mr | 2 | 0.26 | | | ||||
| Diopatra neapolitana Delle Chiaje, 1841 | P | Mr | 2 | 0.26 | | | ||||
| Elasmopus pocillimanus (Spence Bate, 1862) | C | Mr | 2 | 4.30 | | | ||||
| Ericthonius brasiliensis (Dana, 1853) | C | Mr | 2 | 0.25 | | | ||||
| Gammarus insensibilis Stock, 1966 | C | Cs | 2 | 0.39 | | | ||||
| Loripes orbiculatus Poli, 1795 | B | Cs | 2 | 0.45 | | | ||||
| Malacoceros fuliginosus (Claparède, 1868) | P | Op | 2 | 1.62 | | | ||||
| Microdeutopus algicola Della Valle, 1893 | C | Mr | 2 | 2.14 | | | ||||
| Neanthes acuminata (Ehlers, 1868) | P | Cs | 2 | 1.12 | | | ||||
| Oligochaeta | O | 2 | 0.41 | | | |||||
| Paranemonia cinerea (Contarini, 1844) | Cn | Cs | 2 | 0.46 | | | ||||
| Parvicardium exiguum (Gmelin, 1791) | B | Mr | 2 | 0.38 | | | ||||
| Polydora ciliata (Johnston, 1838) | P | Op | 2 | 8.64 | | | ||||
| Protodorvillea kefersteini (McIntosh, 1869) | P | Mr | 2 | 0.58 | | | ||||
| Styela plicata (Lesueur, 1823) | A | Cs | 2 | 1.53 | | | ||||
| Tanais dulongii (Audouin, 1826) | C | Op | 2 | 0.29 | | | ||||
| Amphibalanus amphitrite (Darwin, 1854) | C | Cs | 1 | 1.60 | | |||||
| Amphibalanus eburneus (Gould, 1841) | C | Br | 1 | 2.71 | | |||||
| Amphipholis squamata (Delle Chiaje, 1828) | E | Mr | 1 | 0.82 | | |||||
| Caecum clarkii P. P. Carpenter, 1859 | G | Mr | 1 | 0.47 | | |||||
| Hilbigneris gracilis (Ehlers, 1868) | P | Mr | 1 | 0.25 | | |||||
| Lagis koreni Malmgren, 1866 | P | Mr | 1 | 0.50 | | |||||
| Lumbrineris latreilli Audouin & Milne Edwards, 1833 | P | Cs | 1 | 0.50 | | |||||
| Myriochele heeri Malmgren, 1867 | P | Mr | 1 | 4.50 | | |||||
| Pseudolirius kroyeri (Haller, 1879) | C | Mr | 1 | 0.66 | | |||||
| Sphaerosyllis taylori Perkins, 1981 | P | Mr | 1 | 0.28 | | |||||
| Spiochaetopterus costarum (Claparède, 1869) | P | Mr | 1 | 4.75 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giampaoletti, J.; Sbrana, A.; Magni, P.; Gravina, M.F. Macrobenthos of the Tortolì Lagoon: A Peculiar Case of High Benthic Biodiversity among Mediterranean Lagoons. Diversity 2023, 15, 783. https://doi.org/10.3390/d15060783
Giampaoletti J, Sbrana A, Magni P, Gravina MF. Macrobenthos of the Tortolì Lagoon: A Peculiar Case of High Benthic Biodiversity among Mediterranean Lagoons. Diversity. 2023; 15(6):783. https://doi.org/10.3390/d15060783
Chicago/Turabian StyleGiampaoletti, Jacopo, Alice Sbrana, Paolo Magni, and Maria Flavia Gravina. 2023. "Macrobenthos of the Tortolì Lagoon: A Peculiar Case of High Benthic Biodiversity among Mediterranean Lagoons" Diversity 15, no. 6: 783. https://doi.org/10.3390/d15060783
APA StyleGiampaoletti, J., Sbrana, A., Magni, P., & Gravina, M. F. (2023). Macrobenthos of the Tortolì Lagoon: A Peculiar Case of High Benthic Biodiversity among Mediterranean Lagoons. Diversity, 15(6), 783. https://doi.org/10.3390/d15060783