
Microbial Biopesticides: Diversity, Scope, and Mechanisms Involved in Plant Disease Control

 , , ,
, , ,
Abstract
1. Introduction
2. Direct Interaction between Pathogens and Biocontrol Agents through Physical Contact
2.1. Lysis of Phytopathogenic Bacteria by Bacteriophages
2.2. Destruction of Bacterial Pathogens by Predatory Bacteria
2.3. Mycophagy
2.3.1. Mycophagous Bacteria
2.3.2. Mycophagous Yeasts
2.3.3. Mycoparasitism
Biotrophic Mycoparasitism
Necrotrophic Mycoparasitism
2.4. Hypovirulence of Fungal Pathogens Caused by Mycoviruses
2.5. RNAi-Based-Biocontrol Strategies
3. Interaction between Pathogens and Biocontrol Agents without Physical Contact
3.1. Production of Antimicrobial Compounds
3.1.1. Non-Volatile Antimicrobial Compounds
Antibiotics
Bacteriocins
Killer Toxins
3.1.2. Volatile Organic Compounds with Antimicrobial Activity
3.2. Competition for Nutrients or Space So They Should Be Clearly Identified before
3.3. Inhibition of Virulence Factors of Pathogenic Bacteria by the Elimination of Quorum Sensing Signals
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations (FAO). The State of Food and Agriculture. Moving Forward on Food Loss and Waste Reduction; FAO: Rome, Italy, 2019. [Google Scholar]
- Peng, W.; Berry, E.M. Global nutrition 1990–2015: A shrinking hungry and expanding fat world. PLoS ONE 2018, 13, e0194821. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations (FAO). The State of Food Security and Nutrition in the World 2022. Repurposing Food and Agricultural Policies to Make Healthy Diets More Affordable; FAO: Rome, Italy, 2022. [Google Scholar]
- Beausang, C.; Hall, C.; Toma, L. Food waste and losses in primary production: Qualitative insights from horticulture. Resour. Conserv. Recycl. 2017, 126, 177–185. [Google Scholar] [CrossRef]
- Liu, W.; Liu, X.; Yang, H.; Ciais, P.; Wada, Y. Global water scarcity assessment incorporating green water in crop production. Water Resour. Res. 2022, 58, e2020WR028570. [Google Scholar] [CrossRef]
- Magalhães, V.S.M.; Ferreira, L.M.D.F.; Silva, C. Using a methodological approach to model causes of food loss and waste in fruit and vegetable supply chains. J. Clean. Prod. 2021, 283, 124574. [Google Scholar] [CrossRef]
- US EPA. What are Biopesticides? Available online: https://www.epa.gov/ingredients-used-pesticide-products/what-are-biopesticides (accessed on 24 January 2023).
- Glare, T.R.; O’Callaghan, M. Microbial biopesticides for control of invertebrates: Progress from New Zealand. J. Invertebr. Pathol. 2019, 165, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, S.J.; Reeves, P.T.; Hoang, B.T.; Mitter, N. A perspective on RNAi-based biopesticides. Front. Plant Sci. 2020, 11, 51. [Google Scholar] [CrossRef]
- Zaczek-Moczydłowska, M.A.; Young, G.K.; Trudgett, J.; Plahe, C.; Fleming, C.C.; Campbell, K.; Hanlon, R.O. Phage cocktail containing Podoviridae and Myoviridae bacteriophages inhibits the growth of Pectobacterium spp. under in vitro and in vivo conditions. PLoS ONE 2020, 15, e0230842. [Google Scholar] [CrossRef]
- Czajkowski, R.; Ozymko, Z.; de Jager, V.; Siwinska, J.; Smolarska, A.; Ossowicki, A.; Narajczyk, M.; Lojkowska, E. Genomic, proteomic and morphological characterization of two novel broad host lytic bacteriophages ΦPD10.3 and ΦPD23.1. PLoS ONE 2015, 10, e0119812. [Google Scholar] [CrossRef]
- Pinheiro, L.A.M.; Pereira, C.; Barreal, M.E.; Gallego, P.P.; Balcão, V.M.; Almeida, A. Use of phage ϕ6 to inactivate Pseudomonas syringae pv. actinidiae in kiwifruit plants: In vitro and ex vivo experiments. Appl. Microbiol. Biotechnol. 2020, 104, 1319–1330. [Google Scholar] [CrossRef]
- Rabiey, M.; Roy, S.R.; Holtappels, D.; Franceschetti, L.; Quilty, B.J.; Creeth, R.; Sundin, G.; Wagemans, J.; Lavigne, R.; Jackson, R.W. Phage biocontrol to combat Pseudomonas syringae pathogens causing disease in cherry. Microbiol. Biotechnol. 2020, 13, 1428–1445. [Google Scholar] [CrossRef]
- Akbaba, M.; Ozaktan, H. Evaluation of bacteriophages in the biocontrol of Pseudomonas syringae pv. syringae isolated from cankers on sweet cherry (Prunus avium L.) in Turkey. Egypt. J. Biol. Pest. Control. 2021, 31, 35. [Google Scholar] [CrossRef]
- Hernandez, C.A.; Salazar, A.J.; Koskella, B. Bacteriophage-mediated reduction of bacterial speck on tomato seedlings. Phage 2020, 1, 205–212. [Google Scholar] [CrossRef]
- Goyer, C. Isolation and characterization of phages Stsc1 and Stsc3 infecting Streptomyces scabiei and their potential as biocontrol agents. Can. J. Plant. Pathol. 2005, 27, 210–216. [Google Scholar] [CrossRef]
- Kimmelshue, C.; Goggi, A.S.; Cademartiri, R. The use of biological seed coatings based on bacteriophages and polymers against Clavibacter michiganensis subsp. nebraskensis in maize seeds. Sci. Rep. 2019, 9, 17950. [Google Scholar] [CrossRef]
- Soo, R.M.; Woodcroft, B.J.; Parks, D.H.; Tyson, G.W.; Hugenholtz, P. Back from the dead; the curious tale of the predatory cyanobacterium Vampirovibrio chlorellavorus. Peer. J. 2015, 21, e968. [Google Scholar] [CrossRef]
- Odooli, S.; Roghanian, R.; Emtiazi, G.; Mohkam, M.; Ghasemi, Y. Characterization of the first highly predatory Bdellovibrio bacteriovorus from Iran and its potential lytic activity against principal pathogenic Enterobacteriaceae. Iran. J. Basic. Med. Sci. 2020, 23, 1275–1285. [Google Scholar]
- Youdkes, D.; Helman, Y.; Burdman, S.; Matan, O.; Jurkevitch, E. Potential control of potato soft rot disease by the obligate predators Bdellovibrio and like organisms. Appl. Environ. Microbiol. 2020, 86, e02543-19. [Google Scholar]
- Dong, H.; Xu, X.; Gao, R.; Li, Y.; Li, A.; Yao, Q.; Zhu, H. Myxococcus xanthus R31 suppresses tomato bacterial wilt by inhibiting the pathogen Ralstonia solanacearum with secreted proteins. Front. Microbiol. 2022, 12, 4032. [Google Scholar] [CrossRef]
- Li, Z.; Wang, T.; Luo, X.; Li, X.; Xia, C.; Zhao, Y.; Ye, Z.; Huang, Y.; Gu, X.; Cao, H.; et al. Biocontrol potential of Myxococcus sp. strain BS against bacterial soft rot of calla lily caused by Pectobacterium carotovorum. Biol. Control. 2018, 126, 36–44. [Google Scholar] [CrossRef]
- Pimenta, R.S.; Silva, F.L.; Silva, J.F.; Morais, P.B.; Braga, D.T.; Rosa, C.A.; Corrêa, A. Biological control of Penicillium italicum, P. digitatum and P. expansum by the predacious yeast Saccharomycopsis schoenii on oranges. Braz. J. Microbiol. 2008, 39, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Ferraz, P.; Lopes Brandão, R.; Cássio, F.; Cândida, L. Moniliophthora perniciosa, the causal agent of cacao witches’ broom disease is killed in vitro by Saccharomyces cerevisiae and Wickerhamomyces anomalus yeasts. Front. Microbiol. 2021, 12, 706675. [Google Scholar] [CrossRef]
- Németh, M.Z.; Mizuno, Y.; Kobayashi, H.; Seress, D.; Shishido, N.; Kimura, Y.; Takamatsu, S.; Suzuki, T.; Takikawa, Y.; Kakutani, K.; et al. Ampelomyces strains isolated from diverse powdery mildew hosts in Japan: Their phylogeny and mycoparasitic activity, including timing and quantifying mycoparasitism of Pseudoidium neolycopersici on tomato. PLoS ONE 2021, 16, e0251444. [Google Scholar] [CrossRef] [PubMed]
- Druzhinina, I.S.; Seidl-Seiboth, V.; Herrera-Estrella, A.; Horwitz, B.A.; Kenerley, C.M.; Monte, E.; Mukherjee, P.K.; Zeilinger, S.; Grigoriev, I.V.; Kubicek, C.P. Trichoderma: The genomics of opportunistic success. Nat. Rev. Microbiol. 2011, 9, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Xiao, X.; Fu, Y.; Liu, H.; Cheng, J.; Ghabrial, S.A.; Li, G.; Jiang, D. A novel mycovirus closely related to hypoviruses that infects the plant pathogenic fungus Sclerotinia sclerotiorum. Virology 2011, 418, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Wu, S.; Cheng, J.; Fu, Y.; Jiang, D.; Xie, J. Molecular characterization of two positive-strand RNA viruses co-infecting a hypovirulent strain of Sclerotinia sclerotiorum. Virology 2014, 464, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Niño-Sánchez, J.; Sambasivam, P.T.; Sawyer, A.; Hamby, R.; Chen, A.; Czislowski, E.; Li, P.; Manzie, N.; Gndiner, D.M.; Ford, R.; et al. BioClay™ prolongs RNA interference-mediated crop protection against Botrytis cinerea. J. Integr. Plant. Biol. 2022, 64, 2187–2198. [Google Scholar] [CrossRef]
- Clokie, M.R.; Millard, A.D.; Letarov, A.V.; Heaphy, S. Phages in nature. Bacteriophage 2011, 1, 31–45. [Google Scholar] [CrossRef]
- Ackermann, H.W.; Prangishvili, D. Prokaryote viruses studied by electron microscopy. Arch. Virol. 2012, 157, 1843–1849. [Google Scholar] [CrossRef]
- Guttman, B.; Raya, R.; Kutter, E. Basic phage biology. In Bacteriophages: Biology and Application; Kutter, E., Sulakvelidze, A., Eds.; CRC Press: Boca Raton, FL, USA, 2005; pp. 29–66. [Google Scholar]
- Kazi, M.; Annapure, U.S. Bacteriophage biocontrol of foodborne pathogens. J. Food. Sci. Technol. 2016, 53, 1355–1362. [Google Scholar] [CrossRef]
- Korniienko, N.; Kharina, A.; Budzanivska, I.; Burketová, L.; Kalachova, T. Phages of phytopathogenic bacteria: High potential, but challenging application. Plant Prot. Sci. 2022, 582, 81–91. [Google Scholar] [CrossRef]
- Loc-Carrillo, C.; Abedon, S.T. Pros and cons of phage therapy. Bacteriophage 2011, 1, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Buttimer, C.; McAuliffe, O.; Ross, R.P.; Hill, C.; O’Mahony, J.; Coffey, A. Bacteriophages and bacterial plant diseases. Front. Microbiol. 2017, 8, 34. [Google Scholar] [CrossRef]
- Ramírez, M.; Neuman, B.W.; Ramírez, C.A. Bacteriophages as promising agents for the biological control of Moko disease (Ralstonia solanacearum) of banana. Biol. Control 2020, 149, 104238. [Google Scholar] [CrossRef]
- Umrao, P.D.; Kumar, V.; Kaistha, S.D. Biocontrol potential of bacteriophage ɸsp1 against bacterial wilt-causing Ralstonia solanacearum in Solanaceae crops. Egypt. J. Biol. Pest. Control 2021, 31, 61. [Google Scholar] [CrossRef]
- Obradovic, A.; Mavridis, A.; Rudolph, K. Characterization and PCR-based Typing of Xanthomonas campestris pv. vesicatoria from peppers and tomatoes in Serbia. Eur. J. Plant. Pathol. 2004, 110, 285–292. [Google Scholar] [CrossRef]
- Ranjani, P.; Gowthami, Y.; Gnanamanickam, S.S.; Palani, P. Bacteriophages: A new weapon for the control of bacterial blight disease in rice caused by Xanthomonas oryzae. Microbiol. Biotechnol. Lett. 2018, 46, 346–359. [Google Scholar] [CrossRef]
- Nagy, J.K.; Király, L.; Schwarczinger, I. Phage therapy for plant disease control with a focus on fire blight. Cent. Eur. J. Biol. 2012, 7, 1–12. [Google Scholar] [CrossRef]
- Park, Y.S.; Dutta, S.; Ann, M.; Raaijmakers, J.M.; Park, K. Promotion of plant growth by Pseudomonas fluorescens strain SS101 via novel volatile organic compounds. Biochem. Biophys. Res. Commun. 2015, 461, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Das, M.; Bhowmick, T.S.; Ahern, S.J.; Young, R.; Gonzalez, C.F. Control of Pierce’s disease by phage. PLoS ONE 2015, 10, e0128902. [Google Scholar] [CrossRef] [PubMed]
- Nair, R.R.; Velicer, G.J. Predatory bacteria select for sustained prey diversity. Microorganisms 2021, 9, 2079. [Google Scholar] [CrossRef]
- Madigan, M.Y.; Bender, K.S.; Buckley, D.H.; Sattley, W.M.; Stahl, D.A. Brock Biology of Microorganisms, 16th ed.; Pearson: New York, NY, USA, 2021. [Google Scholar]
- Song, W.Y. Identification and characterization of Bdellovibrio bacteriovorus, a predator of Burkholderia glumae. J. Microbiol. Biotechnol. 2004, 14, 48–55. [Google Scholar]
- Pérez, J.; Moraleda-Muñoz, A.; Marcos-Torres, F.J.; Muñoz-Dorado, J. Bacterial predation. Environ. Microbiol. 2016, 18, 766–779. [Google Scholar] [CrossRef]
- Li, Z.; Ye, X.; Chen, P.; Ji, K.; Zhou, J.; Wang, F.; Weiliang, D.; Yan, H.; Zhengguang, Z.; Cui, Z. Antifungal potential of Corallococcus sp. strain EGB against plant pathogenic fungi. Biol. Control. 2017, 110, 10–17. [Google Scholar] [CrossRef]
- De Boer, W.; Leveau, J.H.J.; Kowalchuk, G.A.; Klein Gunnewiek, P.J.A.; Abeln, E.C.A.; Figge, M.J.; Sjollema, K.; Janse, J.D.; Van Veen, J.A. Collimonas fungivorans gen. nov., sp nov., a chitinolytic soil bacterium with the ability to grow on living fungal hyphae. Int. J. Syst. Evol. Microbiol. 2004, 54, 857–864. [Google Scholar] [CrossRef]
- Kamilova, F.; Leveau, J.H.; Lugtenberg, B. Collimonas fungivorans, an unpredicted in vitro but efficient in vivo biocontrol agent for the suppression of tomato foot and root rot. Environ. Microbiol. 2007, 9, 1597–1603. [Google Scholar] [CrossRef] [PubMed]
- Leveau, J.H.J.; Uroz, S.; De Boer, W. The bacterial genus Collimonas: Mycophagy, weathering and other adaptive solutions to life in oligotrophic soil environments. Environ. Microbiol. 2010, 12, 281–292. [Google Scholar] [CrossRef]
- Lachance, M.A.; Pang, W.M. Predacious Yeasts. Yeast 1997, 13, 225–232. [Google Scholar] [CrossRef]
- Leveau, J.H.J.; Preston, G.M. Bacterial mycophagy: Definition and diagnosis of a unique bacterial-fungal interaction. New Phytol. 2008, 177, 859–876. [Google Scholar] [CrossRef] [PubMed]
- Jeffries, P. Biology and ecology of mycoparasitism. Can. J. Bot. 1995, 73, 1284–1290. [Google Scholar] [CrossRef]
- Jeffries, P.; Young, T.W.K. Interfungal Parasitic Relationships; CAB International: Wallingford, UK, 1994. [Google Scholar]
- Kiss, L.; Russell, J.C.; Szentivanyi, O.; Xu, X.; Jeffries, P. Biology and biocontrol potential of Ampelomyces mycoparasites, natural antagonists of powdery mildew fungi. Biocontrol. Sci. Technol. 2004, 14, 635–651. [Google Scholar] [CrossRef]
- Manjunatha, L.; Singh, S.; Ravikumara, B.M.; Reddy, G.N.; Senthilkumar, M. Ampelomyces. In Beneficial Microbes in Agro-Ecology; Amaresan, N., Senthil Kumar, M., Annapurna, K., Krishna Kumar, A., Eds.; Academic Press: Amsterdam, The Netherlands, 2020; pp. 833–860. [Google Scholar]
- Palmieri, D.; Ianiri, G.; Del Grosso, C.; Barone, G.; De Curtis, F.; Castoria, R.; Lima, G. Advances and perspectives in the use of biocontrol agents against fungal plant diseases. Horticulturae 2022, 8, 577. [Google Scholar] [CrossRef]
- Silva, L.G.; Camargo, R.C.; Mascarin, G.M.; Nunes, P.D.O.; Dunlap, C.; Bettiol, W. Dual functionality of Trichoderma: Biocontrol of Sclerotinia sclerotiorum and biostimulant of cotton plants. Front. Plant Sci. 2022, 13, 983127. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, J.P.; Mead, A.; Payne, T.; Whipps, J.M. Effect of environmental factors and Sclerotium cepivorum isolate on sclerotial degradation and biological control of white rot by Trichoderma. Plant. Pathol. 2004, 53, 353–362. [Google Scholar] [CrossRef]
- Cabrera, M.; Garmendia, G.; Rufo, C.; Pereyra, S.; Vero, S. Trichoderma atroviride as a biocontrol agent of Fusarium head blight by reducing the inoculum of the pathogen in wheat straw. Terra. Latinoamericana 2020, 38, 629–651. [Google Scholar] [CrossRef]
- Whipps, J.M.; Sreenivasaprasad, S.; Muthumeenakshi, S.; Rogers, C.; Challen, M. Use of Coniothyrium minitans as a biocontrol agent and some molecular aspects of sclerotial mycoparasitism. Eur. J. Plant. Pathol. 2008, 121, 323–330. [Google Scholar] [CrossRef]
- Broberg, M.; Dubey, M.; Iqbal, M.; Gudmundssson, M.; Ihrmark, K.; Schroers, H.J.; Jenssen, D.F.; Brandström Durling, M.; Karlsson, M. Comparative genomics highlights the importance of drug efflux transporters during evolution of mycoparasitism in Clonostachys subgenus Bionectria (Fungi, Ascomycota, Hypocreales). Evol. Appl. 2021, 14, 476–497. [Google Scholar] [CrossRef] [PubMed]
- Chet, I.; Harman, G.E.; Baker, R. Trichoderma hamatum: Its hyphal interactions with Rhizoctonia solani and Pythium spp. Microb. Ecol. 1981, 7, 29–38. [Google Scholar] [CrossRef]
- Lu, Z.; Tombolini, R.; Woo, S.; Zeilinger, S.; Lorito, M.; Jansson, J.K. In vivo study of Trichoderma-pathogen-plant interactions, using constitutive and inducible green fluorescent protein reporter systems. Appl. Environ. Microbiol. 2004, 70, 3073–3081. [Google Scholar] [CrossRef]
- Karlsson, M.; Durling, M.B.; Choi, J.; Kosawang, C.; Lackner, G.; Tzelepis, G.D.; Nygren, K.; Dubey, M.K.; Kamou, N.; Levasseur, A.; et al. Insights on the evolution of mycoparasitism from the genome of Clonostachys rosea. Genome Biol. Evol. 2015, 7, 465–480. [Google Scholar] [CrossRef]
- Li, G.Q.H.; Huang, C.; Kokko, E.G.; Acharya, S.N. Ultrastructural study of mycoparasitism of Gliocladium roseum on Botrytis cinerea. Bot. Bull. Acad. Sin. 2002, 43, 211–218. [Google Scholar]
- Lübeck, M.; Knudsen, I.M.B.; Jensen, B.; Thrane, U.; Janvier, C.; Jensen, D.F. GUS and GFP transformation of the biocontrol strain Clonostachys rosea IK726 and the use of these marker genes in ecological studies. Mycol. Res. 2002, 106, 815–826. [Google Scholar] [CrossRef]
- Viterbo, A.; Horwitz, B.A. Mycoparasitism. In Cellular and Molecular Biology of Filamentous Fungi; Borkovich, K.A., Ebbole, D.J., Eds.; ASM Press: Washington DC, USA, 2010; pp. 676–693. [Google Scholar]
- Inbar, J.; Chet, I. Biomimics of fungal cell-cell recognition by use of lectin-coated nylon fibers. J. Bacteriol. 1992, 174, 1055–1059. [Google Scholar] [CrossRef]
- Zeilinger, S.; Galhaup, C.; Payer, K.; Woo, S.L.; Mach, R.L.; Fekete, C.; Lorito, M.; Kubicek, C.P. Chitinase gene expression during mycoparasitic interaction of Trichoderma harzianum with its host. Fungal Genet. Biol. 1999, 26, 131–140. [Google Scholar] [CrossRef]
- Karlsson, M.; Atanasova, L.; Jensen, D.F.; Zeilinger, S. Necrotrophic mycoparasites and their genomes. Microbiol. Spectr. 2017, 5, 10.1128. [Google Scholar] [CrossRef] [PubMed]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Barbetti, M.J.; Li, H.; Woo, S.L.; Lorito, M. A novel role for Trichoderma secondary metabolites in the interactions with plants. Mol. Plant Pathol. 2008, 72, 80–86. [Google Scholar] [CrossRef]
- Utermark, J.; Karlovsky, P. Role of zearalenone lactonase in protection of Gliocladium roseum from fungitoxic effects of the mycotoxin zearalenone. Appl. Environ. Microbiol. 2007, 73, 637–642. [Google Scholar] [CrossRef]
- Rosado, I.V.; Rey, M.; Codón, A.C.; Govantes, J.; Moreno-Mateos, M.A.; Benítez, T. QID74 Cell wall protein of Trichoderma harzianum is involved in cell protection and adherence to hydrophobic surfaces. Fungal. Genet. Biol. 2007, 44, 950–964. [Google Scholar] [CrossRef]
- Son, M.; Yu, J.; Kim, K.H. Five questions about mycoviruses. PLoS Pathogens 2015, 11, e1005172. [Google Scholar] [CrossRef] [PubMed]
- Anagnostakis, S.L. Biological control of chestnut blight. Science 1982, 215, 466–471. [Google Scholar] [CrossRef] [PubMed]
- García-Pedrajas, M.D.; Cañizares, M.C.; Sarmiento-Villamil, J.L.; Jacquat, A.G.; Dambolena, J.S. Mycoviruses in biological control: From basic research to field implementation. Phytopathology 2019, 109, 1828–1839. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Bhattacharjee, P.; Wang, S.; Zhang, L.; Ahmed, I.; Guo, L. Mycoviruses in Fusarium species: An update. Front. Cell. Infect. Microbiol. 2019, 9, 257. [Google Scholar] [CrossRef]
- Aihara, M.; Urayama, S.I.; Le, M.T. Infection by Magnaporthe oryzae chrysovirus 1 strain A triggers reduced virulence and pathogenic race conversion of its host fungus, Magnaporthe oryzae. J. Gen. Plant. Pathol. 2018, 84, 92–103. [Google Scholar] [CrossRef]
- Chiba, S.; Salaipeth, L.; Lin, Y.H.; Sasaki, A.; Kanematsu, S.; Suzuki, N. A novel bipartite double-stranded RNA Mycovirus from the white root rot Fungus Rosellinia necatrix: Molecular and biological characterization, taxonomic considerations, and potential for biological control. J. Virol. 2009, 83, 12801–12812. [Google Scholar] [CrossRef]
- Niu, Y.; Yuan, Y.; Mao, J.; Yang, Z.; Cao, Q.; Zhang, T.; Wang, S.; Liu, D. Characterization of two novel mycoviruses from Penicillium digitatum and the related fungicide resistance analysis. Sci. Rep. 2018, 8, 5513. [Google Scholar] [CrossRef]
- Zhai, L.; Yang, M.; Zhang, M.; Hong, N.; Wang, G. Characterization of a botybirnavirus conferring hypovirulence in the phytopathogenic fungus Botryosphaeria dothidea. Viruses 2019, 11, 266. [Google Scholar] [CrossRef]
- Wang, Q.; Zou, Q.; Dai, Z.; Hong, N.; Wang, G.; Wang, L. Four novel mycoviruses from the hypovirulent Botrytis cinerea SZ-2-3y isolate from Paris polyphylla: Molecular characterization and mitoviral sequence transboundary entry into plants. Viruses 2022, 14, 151. [Google Scholar] [CrossRef]
- Ran, H.; Liu, L.; Li, B.; Cheng, J.; Fu, Y.; Jiang, D.; Xie, J. Co-infection of a hypovirulent isolate of Sclerotinia sclerotiorum with a new botybirnavirus and a strain of a mitovirus. Virol. J. 2016, 6, 92. [Google Scholar] [CrossRef]
- Xiao, X.; Cheng, J.; Tang, J.; Fu, Y.; Jiang, D.; Baker, T.S.; Xie, J. A novel partitivirus that confers hypovirulence on plant pathogenic fungi. J. Virol. 2014, 88, 10120–10133. [Google Scholar] [CrossRef]
- Kanematsu, S.; Shimizu, T.; Salaipeth, L.; Yaegashi, H.; Sasaki, A.; Ito, T.; Suzuki, N. Genome rearrangement of a mycovirus Rosellinia necatrix megabirnavirus 1 affecting its ability to attenuate virulence of the host fungus. Virology 2014, 450, 308–315. [Google Scholar] [CrossRef]
- Lee, K.M.; Yu, J.; Son, M.; Lee, Y.W.; Kim, K.H. Transmission of Fusarium boothii mycovirus via protoplast fusion causes hypovirulence in other phytopathogenic fungi. PLoS ONE 2011, 6, e21629. [Google Scholar] [CrossRef]
- van Diepeningen, A.D.; de Vos, O.J.; Korthals, G.W.; van Bruggen, A.H.C. Effects of organic versus conventional management on chemical and biological parameters in agricultural soils. Appl. Soil. Ecol. 2006, 31, 120–135. [Google Scholar] [CrossRef]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent Andspecific Genetic Interferenceby Double-StrandedRNA in Caenorhabditis eelegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Wang, M.; Weiberg, A.; Lin, F.M. Bidirectional cross-kingdom RNAi and fungal uptake of external RNAs confer plant protection. Nat. Plants 2016, 2, 16151. [Google Scholar] [CrossRef]
- Cai, Q.; He, B.; Kogel, K.H.; Jin, H. Cross-kingdom RNA trafficking and environmental RNAi—Nature’s blueprint for modern crop protection strategies. Curr. Opin. Microbiol. 2018, 46, 58–64. [Google Scholar] [CrossRef]
- Nowara, D.; Gay, A.; Lacomme, C.; Shaw, J.; Ridout, C.; Douchkov, D.; Hensel, G.; Kumlehn, J.; Schweizer, P. HIGS: Host-induced gene silencing in the obligate biotrophic fungal pathogen Blumeria graminis. Plant Cell 2010, 22, 3130–3141. [Google Scholar] [CrossRef]
- Koch, A.; Kumar, N.; Weber, L.; Keller, H.; Imani, J.; Kogel, K.H. host-induced gene silencing of cytochrome P450 lanosterol C14α demethylase-encoding genes confers strong resistance to Fusarium species. Proc. Natl. Acad. Sci. USA 2013, 110, 19324–19329. [Google Scholar] [CrossRef]
- Yin, C.; Jurgenson, J.E.; Hulbert, S.H. Development of a host-induced RNAi system in the wheat stripe rust fungus Puccinia striiformis f. sp. tritici. Mol. Plant-Microbe Interact. 2010, 24, 554–561. [Google Scholar] [CrossRef]
- Jahan, S.N.; Åsman, A.K.M.; Corcoran, P.; Fogelqvist, J.; Vetukuri, R.R.; Dixelius, C. Plant-mediated gene silencing restricts growth of the potato late blight pathogen Phytophthora infestans. J. Exp. Bot. 2015, 66, 2785–2794. [Google Scholar] [CrossRef]
- Liu, Q.; Li, Y.; Xu, K.; Li, D.; Hu, H.; Zhou, F. Clay nanosheet-mediated delivery of recombinant plasmids expressing artificial miRNAs via leaf spray to prevent infection by plant DNA viruses. Hortic. Res. 2020, 7, 179. [Google Scholar] [CrossRef]
- Mitter, N.; Worrall, E.A.; Robinson, K.E.; Li, P.; Jain, R.G.; Taochy, C. Clay nanosheets for topical delivery of RNAi for sustained protection against plant viruses. Nat. Plants 2017, 3, 1607. [Google Scholar] [CrossRef]
- Mehlhorn, S.G.; Geibel, S.; Bucher, G.; Nauen, R. Profiling of RNAi sensitivity after foliar dsRNA exposure in different European populations of Colorado potato beetle reveals a robust response with minor variability. Pest. Biochem. Physiol. 2020, 166, 104569. [Google Scholar] [CrossRef]
- Kolge, H.; Kadam, K.; Galande, S.; Lanjekar, V.; Ghormade, V. New Frontiers in Pest Control: Chitosan nanoparticles-shielded dsRNA as an effective topical RNAi spray for Gram podborer biocontrol. ACS Appl. Bio. Mater. 2021, 4, 5145–5157. [Google Scholar] [CrossRef]
- Gebremichael, D.E.; Haile, Z.M.; Negrini, F.; Sabbadini, S.; Capriotti, L.; Mezzetti, B.; Baraldi, E. RNA Interference Strategies for Future Management of Plant Pathogenic Fungi: Prospects and Challenges. Plants 2021, 10, 650. [Google Scholar] [CrossRef]
- Koch, A.; Höfle, L.; Werner, B.T.; Imani, J.; Schmidt, A.; Jelonek, L. SIGS vs HIGS: A study on the efficacy of two dsRNA delivery strategies to silence Fusarium FgCYP51 genes in infected host and non-host plants. Mol. Plant Pathol. 2019, 20, 1636–1644. [Google Scholar] [CrossRef]
- Sundaresha, S.; Sharma, S.; Bairwa, A.; Tomar, M.; Kumar, R.; Bhardwaj, V. Spraying of dsRNA molecules derived from Phytophthora infestans, as an effective plant protection strategies for the management of potato late blight. Pest. Manag. Sci. 2022, 78, 3183–3192. [Google Scholar]
- Castellanos, N.L.; Smagghe, G.; Sharma, R.; Oliveira, E.E.; Christiaens, O. Liposome encapsulation and EDTA formulation of dsRNA targeting essential genes increase oral RNAi-caused mortality in the Neotropical stink bug Euschistus heros. Pest. Manag. Sci. 2019, 75, 537–548. [Google Scholar] [CrossRef]
- Schwartz, S.; Hendrix, B.; Hoffer, P.; Sanders, R.; Zheng, W. Carbon dots for efficient small interfering RNA delivery and gene silencing in plants. Plant Physiol. 2020, 184, 647–657. [Google Scholar] [CrossRef]
- Ahsan, T.; Yuanhua, W. Plant virus disease management by two modern applications (dsRNA nano-clay sheet and CRISPR/Cas). Arch. Phytopathol. Plant Prot. 2021, 54, 1292–1304. [Google Scholar] [CrossRef]
- Jain, R.G.; Fletcher, S.J.; Manzie, N.; Robinson, K.E.; Li, P.; Lu, E. Foliar application of clay-delivered RNA interference for whitefly control. Nat. Plants 2022, 8, 535–548. [Google Scholar] [CrossRef]
- Daranas, N.; Roselló, G.; Cabrefiga, J.; Donati, I.; Francés, J.; Badosa, E.; Bonaterra, A. Biological control of bacterial plant diseases with Lactobacillus plantarum strains selected for their broad-spectrum activity. Ann. Appl. Biol. 2019, 174, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Balthazar, C.; St-Onge, R.; Léger, G.; Lamarre, S.G.; Joly, D.L.; Filion, M. Pyoluteorin and 2, 4-diacetylphloroglucinol are major contributors to Pseudomonas protegens Pf-5 biocontrol against Botrytis cinerea in cannabis. Front. Microbiol. 2022, 13, 945498. [Google Scholar] [CrossRef]
- Arseneault, T.; Goyer, C.; Filion, M. Pseudomonas fluorescens LBUM223 increases potato yield and reduces common scab symptoms in the field. Phytopathology 2015, 105, 1311–1317. [Google Scholar] [CrossRef]
- Mouloud, G.; Daoud, H.; Bassem, J.; Laribi Atef, I.; Hani, B. New bacteriocin from Bacillus clausii strainGM17: Purification, characterization, and biological activity. Appl. Biochem. Biotechnol. 2013, 171, 2186–2200. [Google Scholar] [CrossRef]
- Ugras, S.; Sezen, K.; Kati, H.; Demirbag, Z. Purification and characterization of the bacteriocin thuricin Bn1 produced by Bacillus thuringiensis subsp. kurstaki Bn1 isolated from a hazelnut pest. J. Microbiol. Biotechnol. 2013, 23, 167–176. [Google Scholar] [CrossRef]
- Scholz, R.; Vater, J.; Budiharjo, A.; Wang, Z.; He, Y.; Dietel, K.; Borriss, R. Amylocyclicin, a novel circular bacteriocin produced by Bacillus amyloliquefaciens FZB42. J. Bacteriol. 2014, 196, 1842–1852. [Google Scholar] [CrossRef]
- Balciunas, E.M.; Martinez, F.A.C.; Todorov, S.D.; de Melo Franco, B.D.G.; Converti, A.; de Souza Oliveira, R.P. Novel biotechnological applications of bacteriocins: A review. Food Control 2013, 32, 134–142. [Google Scholar] [CrossRef]
- Grzegorczyk, M.; Żarowska, B.; Restuccia, C.; Cirvilleri, G. Postharvest biocontrol ability of killer yeasts against Monilinia fructigena and Monilinia fructicola on stone fruit. Food Microbiol. 2017, 61, 93–101. [Google Scholar] [CrossRef]
- Czarnecka, M.; Zarowska, B.; Połomska, X.; Restuccia, C.; Cirvilleri, G. Role of biocontrol yeasts Debaryomyces hansenii and Wickerhamomyces anomalus in plants’ defence mechanisms against Monilinia fructicola in apple fruits. Food Microbiol. 2019, 83, 1–8. [Google Scholar] [CrossRef]
- Madbouly, A.K.; Elyousr, K.A.A.; Ismail, I.M. Biocontrol of Monilinia fructigena, causal agent of brown rot of apple fruit, by using endophytic yeasts. Biol. Control 2020, 144, 104239. [Google Scholar] [CrossRef]
- Bleve, G.; Grieco, F.; Cozzi, G.; Logrieco, A.; Visconti, A. Isolation of epiphytic yeasts with potential for biocontrol of Aspergillus carbonarius and A. niger on grape. Int. Food Microbiol 2006, 108, 204–209. [Google Scholar] [CrossRef]
- Chung, J.H.; Song, G.C.; Ryu, C.M. Sweet scents from good bacteria: Case studies on bacterial volatile compounds for plant growth and immunity. Plant Mol. Biol. 2016, 90, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Vlassi, A.; Nesler, A.; Perazzolli, M.; Lazazzara, V.; Büschl, C.; Parich, A.; Puopolo, G.; Schuhmacher, R. Volatile organic compounds from Lysobacter capsici AZ78 as potential candidates for biological control of soilborne plant pathogens. Front. Microbiol. 2020, 11, 1748. [Google Scholar] [CrossRef]
- Arrarte, E.; Garmendia, G.; Rossini, C.; Wisniewski, M.; Vero, S. Volatile organic compounds produced by Antarctic strains of Candida sake play a role in the control of postharvest pathogens of apples. Biol. Control 2017, 109, 14–20. [Google Scholar] [CrossRef]
- Sepúlveda, X.; Silva, D.; Ceballos, R.; Vero, S.; López, M.D.; Vargas, M. Endophytic yeasts for the biocontrol of Phlyctema vagabunda in apples. Horticulturae 2022, 8, 535. [Google Scholar] [CrossRef]
- Vero, S.; Mondino, P.; Burgueño, J.; Soubes, M.; Wisniewski, M. Characterization of biocontrol activity of two yeast strains from Uruguay against blue mold of apple. Postharvest. Biol. Technol. 2002, 26, 91–98. [Google Scholar] [CrossRef]
- Calvente, V.; De Orellano, M.E.; Sansone, G.; Benuzzi, D.; Sanz de Tosetti, M.I. Effect of nitrogen source and pH on siderophore production by Rhodotorula strains and their application to biocontrol of phytopathogenic moulds. J. Indl. Microbiol. Biotechnol. 2001, 26, 226–229. [Google Scholar] [CrossRef]
- Sansone, G.; Rezza, I.; Calvente, V.; Benuzzi, D.; Sanz de Tosetti, M.I. Control of Botrytis cinerea strains resistant to iprodione in apple with rhodotorulic acid and yeasts. Postharvest. Biol. Technol. 2005, 35, 245–251. [Google Scholar] [CrossRef]
- Vero, S.; Garmendia, G.; González, M.B.; Bentancur, O.; Wisniewski, M. Evaluation of yeasts obtained from Antarctic soil samples as biocontrol agents for the management of postharvest diseases of apple (Malus domestica). FEMS Yeast Res. 2013, 13, 189–199. [Google Scholar] [CrossRef]
- Mahmoudi, E.; Tabatabaei, B.E.S.; Venturi, V. Virulence attenuation of Pectobacterium carotovorum using N-Acyl-homoserine lactone degrading bacteria isolated from potato rhizosphere. Plant Pathol. J. 2011, 27, 242–248. [Google Scholar] [CrossRef]
- Garge, S.S.; Nerurkar, A.S. Attenuation of quorum sensing regulated virulence of Pectobacterium carotovorum subsp. carotovorum through an AHL lactonase produced by Lysinibacillus sp Gs50. PLoS ONE 2016, 11, e0167344. [Google Scholar] [CrossRef]
- Dogi, C.A.; Fochesato, A.; Armando, R.; Pribull, B.; de Souza, M.M.S.; da Silva Coelho, I.; Cavaglieri, L. Selection of lactic acid bacteria to promote efficient silage fermentation capable of inhibiting the activity of Aspergillus parasiticus and Fusarium gramineraum and mycotoxin production. J. Appl. Microbiol. 2013, 114, 1650–1660. [Google Scholar] [CrossRef]
- Thomashow, L.S.; Bonsall, R.E.; Weller, D.M. Antibiotic production by soil and rhizosphere microbes in situ. In Manual of Environmental Microbiology; Hurst, C.J., Knudsen, G.R., McInerney, M.J., Stetzenbach, L.D., Walter, M.V., Eds.; ASM Press: Washington, DC, USA, 1997; pp. 493–499. [Google Scholar]
- Demain, A.L. Pharmaceutically active secondary metabolites of microorganisms. Appl. Microbiol. Biotechnol 1999, 52, 455–463. [Google Scholar] [CrossRef]
- Schaffrath, R.; Meinhardt, F.; Klassen, R. Yeast killer toxins: Fundamentals and applications. In Physiology and Genetics. The Mycota; Anke, T., Schüffler, A., Eds.; Springer: Cham, Switzerland, 2017; Volume 15, pp. 87–118. [Google Scholar]
- Cesa Luna, C.; Baez, A.; Quintero Hernández, V.; de la Cruz Enríquez, J.; Castañeda, A.D.; Muñoz Rojas, J. The importance of antimicrobial compounds produced by beneficial bacteria on the biocontrol of phytopathogens. Acta Biolo. Colom. 2020, 25, 140–154. [Google Scholar] [CrossRef]
- Grady, E.N.; MacDonald, J.; Liu, L.; Richman, A.; Yuan, Z.C. Current knowledge and perspectives of Paenibacillus: A review. Microb. Cell Factories 2016, 15, 203. [Google Scholar] [CrossRef] [PubMed]
- LaGier, M.J.; McDaniel, M.; Ragner, A.; Castillo, A. Identification and characterization of a potential antibiotic producing strain of Pantoea ananatis. J. Genom. 2022, 10, 26–32. [Google Scholar] [CrossRef]
- Christiansen, L.; Alanin, K.S.; Phippen, C.B.; Olsson, S.; Stougaard, P.; Hennessy, R.C. Fungal-associated molecules induce key genes involved in the biosynthesis of the antifungal secondary metabolites nunamycin and nunapeptin in the biocontrol strain Pseudomonas fluorescens In5. Appl. Environ. Microbiol. 2020, 86, e01284-20. [Google Scholar] [CrossRef] [PubMed]
- Michelsen, C.F.; Watrous, J.; Glaring, M.A.; Kersten, R.; Koyama, N.; Dorrestein, P.C.; Stougaard, P. Nonribosomal peptides, key biocontrol components for Pseudomonas fluorescens In5, isolated from a Greenlandic suppressive soil. mBio 2015, 6, e00079-15. [Google Scholar] [CrossRef]
- Malviya, D.; Sahu, P.K.; Singh, U.B.; Paul, S.; Gupta, A.; Gupta, A.R.; Singh, S.; Kumar, M.; Paul, D.; Rai, J.P.; et al. Lesson from ecotoxicity: Revisiting the microbial lipopeptides for the management of emerging diseases for crop protection. Int. J. Environ. Res. Public Health 2020, 23, 1434. [Google Scholar] [CrossRef]
- Kinkel, L.L.; Schlatter, D.C.; Bakker, M.G.; Arenz, B.E. Streptomyces competition and co-evolution in relation to plant disease suppression. Res. Microbiol. 2012, 163, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Moreno, Z.R.; Vinchira-Villarraga, D.M.; Vergara-Morales, D.I.; Castellanos, L.; Ramos, F.A.; Guarnaccia, C.; Moreno-Sarmiento, N. Plant-growth promotion and biocontrol properties of three Streptomyces spp. isolates to control bacterial rice pathogens. Front. Microbiol. 2019, 10, 290. [Google Scholar] [CrossRef]
- Ramachander Turaga, V.N. Peptaibols: Antimicrobial peptides from fungi. In Bioactive Natural products in Drug Discovery; Singh, J., Meshram, V., Gupta, M., Eds.; Springer: Singapore, 2020; pp. 713–730. [Google Scholar]
- Han, P.; Zhang, X.; Xu, D.; Zhang, B.; Lai, D.; Zhou, L. Metabolites from Clonostachys fungi and their biological activities. J. Fungi 2020, 6, 229. [Google Scholar] [CrossRef] [PubMed]
- Moreira Saraiva, R.; Borges, A.V.; Borel, F.C.; Maffia, L.A. Compounds produced by Clonostachys rosea deleterious to Botrytis cinerea. Braz. J. Agric. 2020, 95, 34–47. [Google Scholar]
- Raaijmakers, J.M.; Mazzola, M. Diversity and natural functions of antibiotics produced by beneficial and plant pathogenic bacteria. Annu. Rev. Phytopathol. 2012, 50, 403–424. [Google Scholar] [CrossRef]
- Vaz Jauri, P.; Bakker, M.G.; Salomon, C.E.; Kinkel, L.L. Subinhibitory antibiotic concentrations mediate nutrient use and competition among soil Streptomyces. PLoS ONE 2013, 8, e81064. [Google Scholar] [CrossRef]
- Arseneault, T.; Filion, M. Biocontrol through antibiosis: Exploring the role played by subinhibitory concentrations of antibiotics in soil and their impact on plant pathogens. Can. J. Plant Pathol. 2017, 39, 267–274. [Google Scholar] [CrossRef]
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of action of microbial biological control agents against plant diseases: Relevance beyond efficacy. Front. Plant Sci. 2019, 10, 845. [Google Scholar] [CrossRef]
- Schoenborn, A.A.; Yannarell, S.M.; Wallace, E.D.; Clapper, H.; Weinstein, I.C.; Shank, E.A. Defining the expression, production, and signaling roles of specialized metabolites during Bacillus subtilis differentiation. J. Bacteriol. 2021, 203, e00337-21. [Google Scholar] [CrossRef]
- López, D.; Vlamakis, H.; Kolter, R. Biofilms. Cold Spring Harb. Perspect. Biol. 2010, 2, a000398. [Google Scholar] [CrossRef]
- Huang, R.; Li, Q.; Wang, D.; Feng, H.; Zhang, N.; Shao, J.; Zhang, R. A unique genomic island-governed cannibalism in Bacillus enhanced biofilm formation through a novel regulation mechanism. bioRxiv 2021, 10. [Google Scholar]
- Arseneault, T.; Goyer, C.; Filion, M. Phenazine production by Pseudomonas sp. LBUM223 contributes to the biological control of potato common scab. Phytopathology 2013, 103, 995–1000. [Google Scholar] [CrossRef]
- Bakker, P.A.; Pieterse, C.M.; van Loon, L.C. Induced systemic resistance by fluorescent Pseudomonas spp. Phytopathology 2007, 97, 239–243. [Google Scholar] [CrossRef]
- Phillips, D.A.; Fox, T.C.; King, M.D.; Bhuvaneswari, T.V.; Teuber, L.R. Microbial products trigger amino acid exudation from plant roots. Plant Physiol. 2004, 136, 2887–2894. [Google Scholar] [CrossRef]
- De Leij, F.A.; Dixon-Hardy, J.E.; Lynch, J.M. Effect of 2, 4-diacetylphloroglucinol-producing and non-producing strains of Pseudomonas fluorescens on root development of pea seedlings in three different soil types and its effect on nodulation by Rhizobium. Biol. Fertil. Soils 2002, 35, 114–121. [Google Scholar] [CrossRef]
- Brazelton, J.N.; Pfeufer, E.E.; Sweat, T.A.; Gardener, B.B.; Coenen, C. 2,4-diacetylphloroglucinol alters plant root development. Mol. Plant Microbe Interact. 2008, 21, 1349–1358. [Google Scholar] [CrossRef]
- Combes-Meynet, E.; Pothier, J.F.; Moënne-Loccoz, Y.; Prigent-Combaret, C. The Pseudomonas secondary metabolite 2, 4-diacetylphloroglucinol is a signal inducing rhizoplane expression of Azospirillum genes involved in plant-growth promotion. Mol. Plant-Microbe Int. 2011, 24, 271–284. [Google Scholar] [CrossRef]
- Bernier, S.P.; Surette, M.G. Concentration-dependent activity of antibiotics in natural environments. Front. Microbiol. 2013, 4, 20. [Google Scholar] [CrossRef]
- Penyalver, R.; Vicedo, B.; López, M.M. Use of the genetically engineered Agrobacterium strain K1026 for biological control of crown gall. Eur. J. Plant Pathol. 2000, 106, 801–810. [Google Scholar] [CrossRef]
- Collinge, D.B.; Jensen, D.F.; Rabiey, M.; Sarrocco, S.; Shaw, M.W.; Shaw, R.H. Biological control of plant diseases–What has been achieved and what is the direction? Plant Pathol. 2022, 71, 1024–1047. [Google Scholar] [CrossRef]
- Ajouz, S.; Nicot, P.C.; Bardin, M. Adaptation to pyrrolnitrin in Botrytis cinerea and cost of resistance. Plant Pathol. 2010, 59, 556–566. [Google Scholar] [CrossRef]
- Benítez-Chao, D.F.; León-Buitimea, A.; Lerma-Escalera, J.A.; Morones-Ramírez, J.R. Bacteriocins: An overview of antimicrobial, toxicity, and biosafety assessment by in vivo models. Front. Microbiol. 2021, 12, 630695. [Google Scholar] [CrossRef]
- Martínez, B.; Rodríguez, A.; Suárez, E. Antimicrobial Peptides Produced by Bacteria: The Bacteriocins. In New Weapons to Control Bacterial Growth; Villa, T., Vinas, M., Eds.; Springer: Cham, Switzerland, 2006; pp. 15–38. [Google Scholar]
- Torres, N.I.; Noll, K.S.; Xu, S.; Li, J.; Huang, Q.; Sinko, P.J.; Chikindas, M.L. Safety, formulation and in vitro antiviral activity of the antimicrobial peptide subtilosin against herpes simplex virus type 1. Probiotics Antimicrob. Proteins 2013, 5, 26–35. [Google Scholar] [CrossRef]
- Akerey, B.; Le-Lay, C.; Fliss, I.; Subirade, M.; Rouabhia, M. In vitro efficacy of nisin Z against Candida albicans adhesion and transition following contact with normal human gingival cells. J. Appl. Microbiol. 2019, 107, 1298–1307. [Google Scholar] [CrossRef]
- Hernández-González, J.C.; Martínez-Tapia, A.; Lazcano-Hernández, G.; García-Pérez, B.E.; Castrejón-Jiménez, N.S. Bacteriocins from lactic acid bacteria. A powerful alternative as antimicrobials, probiotics, and immunomodulators in veterinary medicine. Animals 2021, 11, 979. [Google Scholar] [CrossRef]
- Simons, A.; Alhanout, K.; Duval, R.E. Bacteriocins, antimicrobial peptides from bacterial origin: Overview of their biology and their impact against multidrug-resistant bacteria. Microorganisms 2020, 27, 639. [Google Scholar] [CrossRef]
- Lavermicocca, P.; Lonigro, S.L.; Valerio, F.; Evidente, A.; Visconti, A. Reduction of olive knot disease by a bacteriocin from Pseudomonas syringae pv. ciccaronei. Appl. Environ. Microbiol. 2002, 68, 1403–1407. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.Y.; Echandi, E. Effects of avirulent bacteriocin-producing strains of Pseudomonas solanacearum on the control of bacterial wilt of tobacco. Plant Pathol. 1984, 33, 245–253. [Google Scholar] [CrossRef]
- Sakthivel, N.; Mew, T.W. Efficacy of bacteriocinogenic strains of Xanthomonas oryzae pv. oryzae on the incidence of bacterial blight disease of rice (Oryza sativa L.). Can. J. Microbiol. 1991, 37, 764–768. [Google Scholar] [CrossRef]
- Liu, G.L.; Chi, Z.; Wang, G.Y.; Wang, Z.P.; Li, Y.; Chi, Z.M. Yeast killer toxins, molecular mechanisms of their action and their applications. Crit. Rev. Biotechnol. 2015, 35, 222–234. [Google Scholar] [CrossRef]
- Bevan, E.A.; Makower, M. The physiological basis of the killer character in yeast. Proc. XIth Int. Congr. Genet. 1963, 1, 202–203. [Google Scholar]
- Schmitt, M.; Breinig, F. Yeast viral killer toxins: Lethality and self-protection. Nat. Rev. Microbiol. 2006, 4, 212–221. [Google Scholar] [CrossRef]
- Mannazzu, I.; Domizio, P.; Carboni, G.; Zara, S.; Zara, G.; Comitini, F.; Ciani, M. Yeast killer toxins: From ecological significance to application. Crit. Rev. Biotechnol. 2019, 39, 603–617. [Google Scholar] [CrossRef]
- Muccilli, V.; Cunsolo, V.; Saletti, R.; Foti, S.; Margiotta, B.; Scossa, F.; Masci, S.; Lafiandra, D. Characterization of a specific class of typical low molecular weight glutenin subunits of durum wheat by a proteomic approach. J. Cereal. Sci. 2010, 51, 134–139. [Google Scholar] [CrossRef][Green Version]
- Muccilli, S.; Wemhoff, S.; Restuccia, C.; Meinhardt, F. Exoglucanase-encoding genes from three Wickerhamomyces anomalus killer strains isolated from olive brine. Yeast 2013, 30, 33–43. [Google Scholar] [CrossRef]
- Platania, C.; Restuccia, C.; Muccilli, S.; Cirvilleri, G. Efficacy of killer yeasts in the biological control of Penicillium digitatum on Tarocco orange fruits (Citrus sinensis). Food Microbiol. 2012, 30, 219–225. [Google Scholar] [CrossRef]
- Perez, M.F.; Contreras, L.; Garnica, N.M.; Fernández-Zenoff, M.V.; Farías, M.E.; Sepulveda, M.; Dib, J.R. Native killer yeasts as biocontrol agents of postharvest fungal diseases in lemons. PLoS ONE 2016, 11, e0165590. [Google Scholar] [CrossRef]
- Marquina, D.; Santos, A.; Peinado, J. Biology of killer yeasts. Int. Microbiol. 2002, 5, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.; Marquina, D. Killer toxin of Pichia membranifaciens and its possible use as a biocontrol agent against grey mould disease of grapevine. Microbiology 2004, 150, 2527–2534. [Google Scholar] [CrossRef] [PubMed]
- Díaz, M.A.; Pereyra, M.M.; Picón-Montenegro, E.; Meinhardt, F.; Dib, J.R. Killer yeasts for the biological control of postharvest fungal crop diseases. Microorganisms 2020, 8, 1680. [Google Scholar] [CrossRef]
- Magliani, W.; Conti, S.; Salati, A.; Vaccari, S.; Ravanetti, L.; Maffei, D.L.; Polonelli, L. Therapeutic potential of yeast killer toxin-like antibodies and mimotopes. FEMS Yeast Res. 2004, 5, 11–18. [Google Scholar] [CrossRef]
- Liu, Z.; Du, S.; Ren, Y.; Liu, Y. Biocontrol ability of killer yeasts (Saccharomyces cerevisiae) isolated from wine against Colletotrichum gloeosporioides on grape. J. Basic Microbiol. 2018, 58, 60–67. [Google Scholar] [CrossRef]
- Lopes, M.R.; Klein, M.N.; Ferraz, L.P.; da Silva, A.C.; Kupper, K.C. Saccharomyces cerevisiae: A novel and e_cient biological control agent for Colletotrichum acutatum during pre-harvest. Microbiol. Res. 2015, 175, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Perez, M.F.; Ibarreche, J.P.; Isas, A.S.; Sepulveda, M.; Ramallo, J.; Dib, J.R. Antagonistic yeasts for the biological control of Penicillium digitatum on lemons stored under export conditions. Biol. Control 2017, 115, 135–140. [Google Scholar] [CrossRef]
- Perez, M.F.; Díaz, M.A.; Pereyra, M.M.; Córdoba, J.M.; Isas, A.S.; Sepúlveda, M.; Dib, J.R. Biocontrol features of Clavispora lusitaniae against Penicillium digitatum on lemons. Postharvest. Biol. Technol. 2019, 155, 57–64. [Google Scholar] [CrossRef]
- Da Cunha, T.; Ferraz, L.P.; Wehr, P.P.; Kupper, K.C. Antifungal activity and action mechanisms of yeasts isolates from citrus against Penicillium italicum. Int. J. Food Microbiol. 2018, 276, 20–27. [Google Scholar] [CrossRef]
- Vespermann, A.; Kai, M.; Piechulla, B. Rhizobacterial volatiles affect the growth of fungi and Arabidopsis thaliana. Appl. Environ. Microbiol. 2007, 17, 5639–5641. [Google Scholar] [CrossRef]
- Bennett, J.W.; Hung, R.; Lee, S.; Padhi, S. Fungal and bacterial volatile organic compounds: An overview and their role as ecological signaling agents. In Fungal Associations. The Mycota; Hock, B., Ed.; Springer: Berlin Heidelberg, Germany, 2012; Volume 9, pp. 373–393. [Google Scholar]
- Werner, S.; Polle, A.; Brinkmann, N. Belowground communication: Impacts of volatile organic compounds (VOCs) from soil fungi on other soil-inhabiting organisms. Appl. Microbiol. Biotechnol. 2016, 100, 8651–8665. [Google Scholar] [CrossRef] [PubMed]
- Morath, S.U.; Hung, R.; Bennett, J.W. Fungal volatile organic compounds: A review with emphasis on their biotechnological potential. Fungal Biol. Rev. 2012, 26, 73–83. [Google Scholar] [CrossRef]
- Zhao, X.; Zhou, J.; Tian, R.; Liu, Y. Microbial volatile organic compounds: Antifungal mechanisms, applications, and challenges. Front. Microbiol. 2022, 13, 922450. [Google Scholar] [CrossRef]
- Korpi, A.; Jill Järnberg, J.; Pasanen, A.L. Microbial volatile organic compounds. Cri. Rev. Toxicol. 2009, 39, 139–193. [Google Scholar] [CrossRef]
- Parafati, L.; Vitale, A.; Restuccia, C.; Cirvilleri, G. Biocontrol ability and action mechanism of food-isolated yeast strains against Botrytis cinerea causing post-harvest bunch rot of table grape. Food Microbiol. 2015, 47, 85–92. [Google Scholar] [CrossRef]
- Arrebola, E.; Sivakumar, D.; Korsten, L. Effect of volatile compounds produced by Bacillus strains on postharvest decay in citrus. Biol. Control 2010, 53, 122–128. [Google Scholar] [CrossRef]
- Chen, H.; Xiao, X.; Wang, J.; Wu, L.J.; Zheng, Z.M.; Yu, Z.L. Antagonistic effects of volatiles generated by Bacillus subtilis on spore germination and hyphal growth of the plant pathogen Botrytis cinerea. Biotechnol. Lett. 2008, 30, 919–923. [Google Scholar] [CrossRef]
- Yuan, J.; Raza, W.; Shen, Q.R.; Huang, Q.W. Antifungal activity of Bacillus amyloliquefaciens NJN-6 volatile compounds against Fusarium oxysporum f. sp cubense. Appl. Environ. Microbiol. 2012, 78, 5942–5944. [Google Scholar] [CrossRef]
- Gotor-Vila, A.; Teixido, N.; Di Francesco, A.; Usall, J.; Ugolini, L.; Torres, R.; Mari, M. Antifungal effect of volatile organic compounds produced by Bacillus amyloliquefaciens CPA-8 against fruit pathogen decays of cherry. Food Microbiol. 2017, 64, 219–225. [Google Scholar] [CrossRef]
- Farbo, M.G.; Urgeghe, P.P.; Fiori, S.; Marcello, A.; Oggiano, S.; Balmas, V. Effect of yeast volatile organic compounds on ochratoxin A-producing Aspergillus carbonarius and A. ochraceus. Int. J. Food Microbiol. 2018, 284, 1–10. [Google Scholar] [CrossRef]
- Tilocca, B.; Balmas, V.; Hassan, Z.U.; Jaoua, S.; Migheli, Q. A proteomic investigation of Aspergillus carbonarius exposed to yeast volatilome or to its major component 2-phenylethanol reveals major shifts in fungal metabolism. Int. J. Food Microbiol. 2019, 306, 108265. [Google Scholar] [CrossRef]
- Rania, A.; Rana, A.; Kumar Dhakac, R.; Pratap Singhd, A.; Chahare, M.; Singhf, S.; Naing, L.; Pal Singhh, K.; Minzj, D. Bacterial volatile organic compounds as biopesticides, growth promoters and plant-defense elicitors: Current understanding and future scope. Biotechnol. Adv. 2023, 63, 108078. [Google Scholar] [CrossRef]
- Groenhagen, U.; Baumgartner, R.; Bailly, A.; Gardiner, A.; Eberl, L.; Schulz, S.; Weisskopf, L. Production of bioactive volatiles by different Burkholderia ambifaria strains. J. Chem. Ecol. 2013, 39, 892–906. [Google Scholar] [CrossRef]
- Popova, A.A.; Koksharova, O.A.; Lipasova, V.A.; Zaitseva, J.V.; Katkova-Zhukotskaya, O.A.; Eremina, S.I. Inhibitory and toxic effects of volatiles emitted by strains of Pseudomonas and Serratia on growth and survival of selected microorganisms, Caenorhabditis elegans, and Drosophila melanogaster. Biomed. Res. 2014, 2014, 125704. [Google Scholar]
- Jing Li, A.; Kumar Pal, V.; Kurunthachalam Kannan, K. A review of environmental occurrence, toxicity, biotransformation and biomonitoring of volatile organic compounds. Environ. Chem. Ecotoxicol. 2021, 3, 91–116. [Google Scholar]
- Rendueles, O.; Ghigo, J.M. Mechanisms of competition in biofilm communities. Microbiol. Spectr. 2015, 3, 319–342. [Google Scholar] [CrossRef] [PubMed]
- Spadaro, D.; Droby, S. Development of biocontrol products for postharvest diseases of fruit: The importance of elucidating the mechanisms of action of yeast antagonists. Trends Food Sci. Technol. 2016, 47, 39–49. [Google Scholar] [CrossRef]
- Janisiewicz, W.J.; Korsten, L. Biological control of postharvest diseases of fruits. Annu. Rev. Phytopathol. 2002, 40, 411–441. [Google Scholar] [CrossRef] [PubMed]
- Bencheqroun, S.K.; Bajji, M.; Massart, S.; Labhilili, M.; El Jaafari, S.; Jijakli, H. In vitro and in situ study of postharvest apple blue mold biocontrol by Aureobasidium pullulans: Evidence for the involvement of competition for nutrients. Postharvest. Biol. Technol. 2007, 46, 128–135. [Google Scholar] [CrossRef]
- Lahlali, R.; Ezrari, S.; Radouane, N.; Kenfaoui, J.; Esmaeel, Q.; El Hamss, H.; Belabess, Z.; Barka, E.A. Biological control of plant pathogens: A Global Perspective. Microorganisms 2022, 10, 596. [Google Scholar] [CrossRef]
- van Loon, L.C. Helping plants to defend themselves: Biocontrol by disease-suppressing rhizobacteria. In Developments in Plant Genetics and Breeding; de Vries, G.E., Metzlaff, K., Eds.; Elsevier: Amsterdam, The Netherlands, 2000; pp. 203–213. [Google Scholar]
- Hider, R.C.; Kong, X. Chemistry and biology of siderophores. Nat. Prod. Rep. 2010, 27, 637–657. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Perez, L.J.; Ng, W.L.; Semmelhack, M.F.; Bassler, B.L. Mechanism of Vibrio cholerae autoinducer-1 biosynthesis. ACS Chem. Biol. 2011, 6, 356–365. [Google Scholar] [CrossRef]
- Alam, A.; Kumar, A.; Tripathi, P.; Ehtesham, N.Z.; Hasnain, S.E. Biofilms: A phenotypic mechanism of bacteria conferring tolerance against stress and antibiotics. In Mycobacterium Tuberculosis: Molecular Infection Biology, Pathogenesis, Diagnostics and New Interventions; Hasnain, S., Ehtesham, N., Grover, S., Eds.; Springer: Singapore, 2019; pp. 315–333. [Google Scholar]
- Kröber, M.; Verwaaijen, B.; Wibberg, D.; Winkler, A.; Pühler, A.; Schlüter, A. Comparative transcriptome analysis of the biocontrol strain Bacillus amyloliquefaciens FZB42 as response to biofilm formation analyzed by RNA sequencing. J. Biotechnol. 2016, 231, 212–223. [Google Scholar] [CrossRef]
- Arrarte, E.; Garmendia, G.; Wisniewski, M.; Vero, S. Biocontrol activity of Debaryomyces hansenii against blue mold on apple and pear during cold storage. Agrociencia Urug. 2021, 25, e839. [Google Scholar] [CrossRef]
- Li, Y.H.; Tian, X. Quorum Sensing and Bacterial Social Interactions in Biofilms. Sensors 2012, 12, 2519–2538. [Google Scholar] [CrossRef]
- Prazdnova, E.V.; Gorovtsov, A.V.; Vasilchenko, N.G.; Kulikov, M.P.; Statsenko, V.N.; Bogdanova, A.A.; Refeld, A.G.; Brislavskiy, Y.A.; Chistyakov, V.A.; Chikindas, M.L. Quorum-sensing inhibition by Gram-positive bacteria. Microorganisms 2022, 10, 350. [Google Scholar] [CrossRef]
- Jayaraman, A.; Wood, T.K. Bacterial quorum sensing: Signals, circuits, and implications for biofilms and disease. Annu. Rev. Biomed. Eng. 2008, 10, 145–167. [Google Scholar] [CrossRef]
- Deng, Y.; Wu, J.; Tao, F.; Zhang, L.H. Listening to a new language: DSF based quorum sensing in Gram-negative bacteria. Chem. Rev. 2011, 111, 160–173. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, S.T.; Bassler, B.L. Bacterial quorum sensing: Its role in virulence and possibilities for its control. Cold Spring Harb. Perspect. Med. 2012, 2, a012427. [Google Scholar] [CrossRef]
- Novick, R.P.; Geisinger, E. Quorum sensing in staphylococci. Annu. Rev. Genet. 2008, 42, 541–564. [Google Scholar] [CrossRef] [PubMed]
- Ng, W.L.; Bassler, B.L. Bacterial quorum-sensing network architectures. Annu. Rev. Genet. 2009, 43, 19. [Google Scholar] [CrossRef]
- Fan, X.; Ye, T.; Li, Q.; Bhatt, P.; Zhang, L.; Chen, S. Potential of a quorum quenching bacteria isolate Ochrobactrum intermedium D-2 against soft rot pathogen Pectobacterium carotovorum subsp. carotovorum. Front. Microbiol. 2020, 11, 898. [Google Scholar] [CrossRef]
- LaSarre, B.; Federle, M.J. Exploiting quorum sensing to confuse bacterial pathogens. Microbiol. Mol. Biol. Rev. 2013, 77, 73–111. [Google Scholar] [CrossRef] [PubMed]
- Grandclement, C.; Tannieres, M.; Morera, S.; Dessaux, Y.; Faure, D. Quorum quenching: Role in nature and applied developments. FEMS Microbiol. Rev. 2016, 40, 86–116. [Google Scholar] [CrossRef]
- Dong, Y.H.; Gusti, A.R.; Zhang, Q.; Xu, J.L.; Zhang, L.H. Identification of quorum-quenching N-acyl homoserine lactonases from Bacillus species. Appl. Environ. Microbiol. 2002, 68, 1754–1759. [Google Scholar] [CrossRef]
- Haque, S.; Yadav, D.K.; Bisht, S.C.; Yadav, N.; Singh, V.; Dubey, K.K.; Jawed, A.; Wahid, M.; Dar, S.A. Quorum sensing pathways in Gram-positive and-negative bacteria: Potential of their interruption in abating drug resistance. J. Chemother. 2019, 31, 161–187. [Google Scholar] [CrossRef] [PubMed]
- Paluch, E.; Rewak-Soroczynska, J.; Jedrusik, I.; Mazurkiewicz, E.; Jermakow, K. Prevention of biofilm formation by quorum quenching. Appl. Microbiol. Biotechnol. 2020, 104, 1871–1881. [Google Scholar] [CrossRef]
- Lim, J.; Jee, S.; Lee, D.H.; Roh, E.; Jung, K.; Oh, C. Biocontrol of Pectobacterium carotovorum subsp. carotovorum using bacteriophage PP1. J. Microbiol. Biotechn. 2013, 23, 1147–1153. [Google Scholar] [CrossRef]
- Joshi, J.R.; Burdman, S.; Lipsky, A.; Yariv, S.; Yedidia, I. Plant phenolic acids affect the virulence of Pectobacterium aroidearum and P. carotovorum ssp brasiliense via quorum sensing regulation. Mol. Plant Pathol. 2016, 17, 487–500. [Google Scholar] [CrossRef]
- Pollumaa, L.; Alamaee, T.; Maee, A. Quorum sensing and expression of virulence in Pectobacteria. Sensors 2012, 12, 3327–3349. [Google Scholar] [CrossRef]
- Mehmood, A.; Liu, G.; Wang, X.; Meng, G.; Wang, C.; Liu, Y. Fungal quorum-sensing molecules and inhibitors with potential antifungal activity: A review. Molecules 2019, 24, 1950. [Google Scholar] [CrossRef]
- Wongsuk, T.; Pumeesat, P.; Luplertlop, N. Fungal quorum sensing molecules: Role in fungal morphogenesis and pathogenicity. J. Basic Microbiol. 2016, 56, 440–447. [Google Scholar] [CrossRef]
- Williams, H.E.; Steele, J.C.; Clements, M.O. γ-Heptalactone is an endogenously produced quorum sensing molecule regulating growth and secondary metabolite production by Aspergillus nidulans. Appl. Microbiol. Biotech. 2012, 96, 773–781. [Google Scholar] [CrossRef]
- Rasmussen, T.B.; Skindersoe, M.E.; Bjarnsholt, T.; Phipps, R.K.; Christensen, K.B.; Jensen, P.O.; Andersen, J.B.; Koch, B.; Larsen, T.O.; Hentzer, M.; et al. Identity and effects of quorum-sensing inhibitors produced by Penicillium species. Microbiology 2005, 151, 1325–1340. [Google Scholar] [CrossRef]
- Dor, S.; Prusky, D.; Afriat-Jurnou, L. Bacterial quorum-quenching lactonase hydrolyzes fungal mycotoxin and reduces pathogenicity of Penicillium expansum—Suggesting a mechanism of bacterial antagonism. J. Fungi 2021, 7, 826. [Google Scholar] [CrossRef]
- Köhl, J.; Postma, J.; Nicot, P.; Ruocco, M.; Blum, B. Stepwise screening of microorganisms for commercial use in biological control of plant-pathogenic fungi and bacteria. Biol. Control 2011, 57, 1–12. [Google Scholar] [CrossRef]


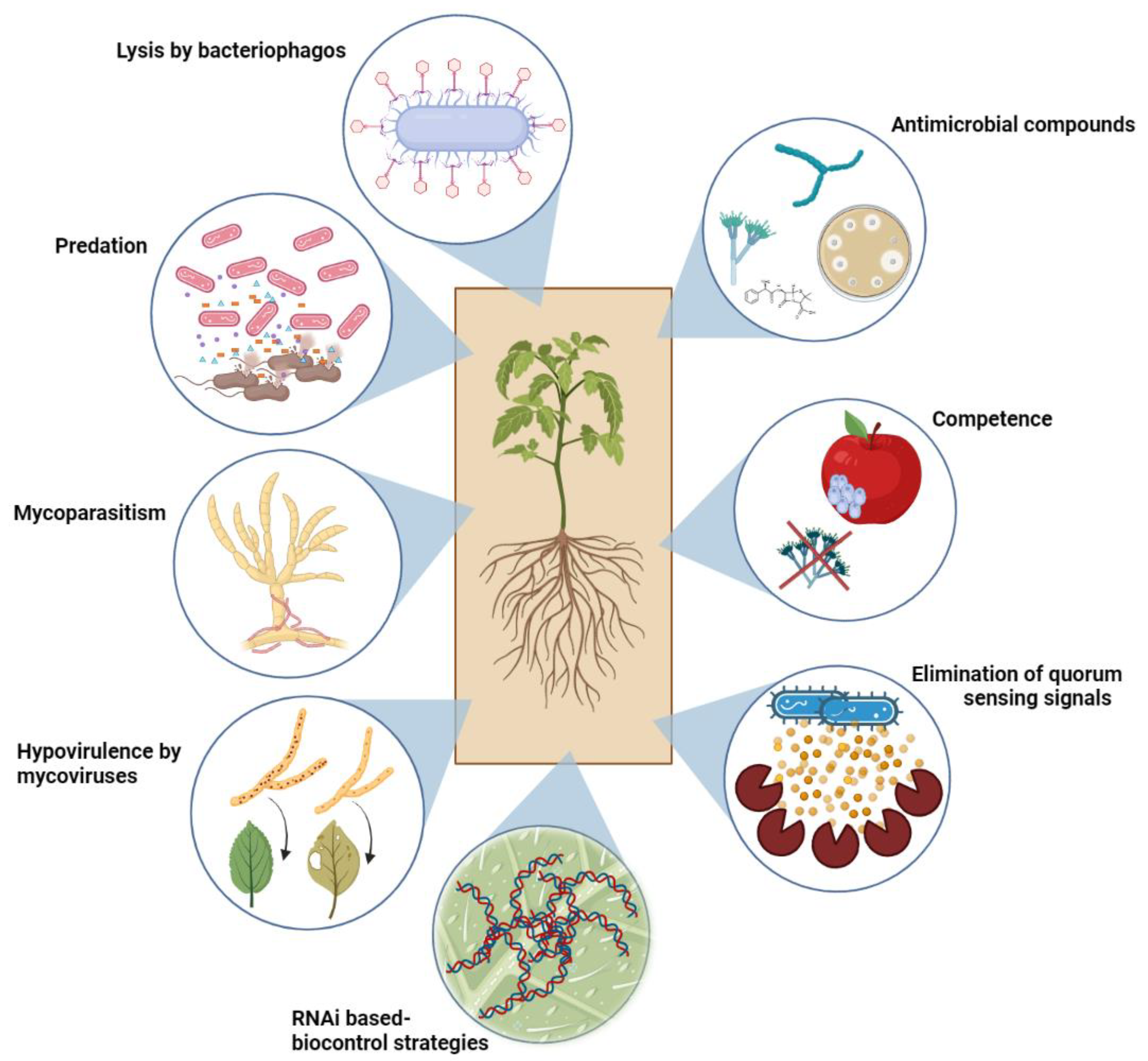{kind=link}
| Mechanism of Action | Biocontrol Agent | Pathogen | Crop | Reference |
|---|---|---|---|---|
| Lysis of phytopathogenic bacteria | Bacteriophages | Pectobacterium spp. | Potato | Zaczek-Moczydłowska et al., 2020 [10] |
| Pectobacterium spp. and Dickeya spp. | Potato | Czajkowski et al., 2015; Zaczek-Moczydłowska et al., 2020 [10,11] | ||
| Pseudomonas syringae pv. actinidiae | Leaves of kiwifruit | Pinheiro et al., 2020 [12] | ||
| Pseudomonas syringae pv. syringae | Cherry leaves Sweet cherry plantlets | Rabiey et al., 2020; Akbaba and Ozaktan, 2021 [13,14] | ||
| Pseudomonas syringae pv. tomato | Tomato seedlings | Hernandez et al., 2020 [15] | ||
| Streptomyces scabies | Potato | Goyer, 2005 [16] | ||
| Clavibacter michiganensis | Maize seeds | Kimmelshue et al., 2019 [17] | ||
| Destruction of bacterial pathogens by predatory bacteria | Vampirovibrio chlorellavorus (Epibiotic strategy) | Chlorella vulgaris | - | Soo et al., 2015 [18] |
| Bdellovibrio bacteriovorus strain SOIR-1 (Endobiotic strategy) | Xanthomonas campestris Pantoea sp. Pectobacterium carotovorum subsp. brasilense | Potato slices Onion bulbs Potato slices | Odooli et al. (2020) [19] Youdkes et al. (2020) [20] | |
| Myxococcus xanthus R31 Myxococcus sp. strain BS (Group attack) | Ralstonia solanacearum Pectobacterium carotovorum | Tomato Calla lily | Dong et al., 2022 [21] Li et al., 2018 [22] | |
| Mycophagy | Saccharomycopsis schoenii Wickerhamomyces anomalus LBCM1105 | Penicillium digitatum Penicillium expansum Moniliophthora perniciosa | Oranges Apples Cacao | Pimenta et al., 2018 [23] Ferraz et al., 2021 [24] |
| Mycoparasitism | Ampelomyces quisqualis (Biotrophic mycoparasitism) Trichoderma spp. (Necrotrophic mycoparasitism) | Pseudoidium neolycopersici Rhizoctonia solani, Botrytis cinerea, Sclerotinia sclerotiorum, Alternaria alternata, Fusarium spp., and oomycetes such as Pythium ultimum | Tomato/powdery mildew Many crops | Németh et al., 2021 [25] Druzhinina et al., 2011 [26] |
| Hypovirulence of fungal pathogens | Mycoviruses: SsHV1 and SsHV2 | Sclerotinia sclerotiorum | - | Xie et al., 2014 [27,28] |
| Silencing target genes in plant pathogens by RNA interference (RNAi) | Specific dsRNA | Botrytis cinerea | Tomato leaves | Niño-Sánchez, J. et al., 2022 [29] |
| Mechanism of Action | Biocontrol Agent | Pathogen | Crop | Reference |
|---|---|---|---|---|
| Pathogen inhibition by primary metabolites | Lactic acid produced by Lactobacillus plantarum | Pseudomonas syringae pv. actinidiae Xanthomonas arboricola pv. pruni Xanthomonas fragariae in strawberry | Kiwifruit Prunus Strawberries | Daranas et al., 2019 [108] |
| Pathogen inhibition by antibiotics | Pyoluteorin and 2,4-diacetylphloroglucinol produced by Pseudomonas protegens | Botrytis cinerea | Cannabis | Balthazar et al., 2022 [109] |
| Phenazine-1-carboxylic acid (PCA) produced by Pseudomonas fluorescens LBUM223 | Streptomyces scabies | Potato | Arseneault et al., 2015 [110] | |
| Pathogen inhibition by bacteriocins | BacGM17 produced by Bacillus clausii GM17 | Agrobacterium tumefaciens | - | Mouloud et al., 2013 [111] |
| Thuricin Bn1 secreted by Bacillus thuringiensis subsp. kurstaki Bn1 | Pseudomonas savastanoi and Pseudomonas syringae | - | Ugras et al., 2014 [112] | |
| Amylocyclicin produced by Bacillus amyloliquefaciens subsp. plantarum FZB42 | Clavibacter michiganensis | - | Scholz et al., 2014 [113] | |
| Nisin produced by Lactic acid bacteria | Clostridium botulinum, Bacillus cereus, Listeria monocytogenes, and Staphylococcus aureus | - | Balciunas et al., 2013 [114] | |
| Pathogen inhibition by killer toxins | Debaryomyces hansenii MI1a, D. hansenii K12a and Wickerhamomyces anomalus BS91 | Monilinia fructigena and Monilinia fructicola | Stone fruit | Grzergorczyk et al., 2017 [115] |
| Schwanniomyces sp., Galactomyces sp., and Rhodotorula sp. | Monilinia fructigena, Monilinia fructicola, and Aspergillus niger | Apples | Madbouly et al., 2020 and Czarnecka et al., 2019 [116,117] | |
| Issatchenkia orientalis strains 17C2 and 16C2 | Aspegillus carbonarius and Aspergillus niger | Grapes | Bleve et al., 2006 [118] | |
| Pathogen inhibition by volatile organic compounds with antimicrobial activity | Bacillus sp. and Enterobacter sp. | Botrytis cinerea, Colletotrichum heterostrophus, and Setosphaeria turcica | Tobacco and maize plants | Chung et al., 2016; Vlassi et al., 2020 [119,120] |
| Candida sake | Penicillium expansum, Botrytis cinerea, Alternaria alternata, Alternaria tenuissima, and Alternaria arborescens | Apples | Arrarte et al., 2017 [121] | |
| Vishniacozyma victoriae | Phlyctema vagabunda | Apples | Sepúlveda et al., 2022 [122] | |
| Competition for resources | Competition for nitrogen: Cryptococcus laurentii 317 and Candida ciferrii 283 | Penicillium expansum | Apples | Vero, et al., 2002 [123] |
| Competition for iron: rhodotorulic acid produced by Rhodotorula glutinis | Penicillium expansum Botrytis cinerea | Apples | Calvente et al., 2001 Sansone et al., 2005 [124,125] | |
| Competition for space: Leucosporidium scottii | Penicillium expansum Botrytis cinerea | Apple | Vero et al. 2013 [126] | |
| Inhibition of virulence factors by elimination of quorum sensing signals | AHL lactonases produced by Mesorhizobium sp. and Lysinibacillus sp. | Pectobacterium carotovorum subsp. carotovorum | Potato, carrot, and cucumber | Mahmoudi et al.,2011; Garge and Nerurkar, 2016 [127,128] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vero, S.; Garmendia, G.; Allori, E.; Sanz, J.M.; Gonda, M.; Alconada, T.; Cavello, I.; Dib, J.R.; Diaz, M.A.; Nally, C.; et al. Microbial Biopesticides: Diversity, Scope, and Mechanisms Involved in Plant Disease Control. Diversity 2023, 15, 457. https://doi.org/10.3390/d15030457
Vero S, Garmendia G, Allori E, Sanz JM, Gonda M, Alconada T, Cavello I, Dib JR, Diaz MA, Nally C, et al. Microbial Biopesticides: Diversity, Scope, and Mechanisms Involved in Plant Disease Control. Diversity. 2023; 15(3):457. https://doi.org/10.3390/d15030457
Chicago/Turabian StyleVero, Silvana, Gabriela Garmendia, Enzo Allori, José María Sanz, Mariana Gonda, Teresa Alconada, Ivana Cavello, Julián Rafael Dib, Mariana Andrea Diaz, Cristina Nally, and et al. 2023. "Microbial Biopesticides: Diversity, Scope, and Mechanisms Involved in Plant Disease Control" Diversity 15, no. 3: 457. https://doi.org/10.3390/d15030457
APA StyleVero, S., Garmendia, G., Allori, E., Sanz, J. M., Gonda, M., Alconada, T., Cavello, I., Dib, J. R., Diaz, M. A., Nally, C., Pimenta, R. S., da Silva, J. F. M., Vargas, M., Zaccari, F., & Wisniewski, M. (2023). Microbial Biopesticides: Diversity, Scope, and Mechanisms Involved in Plant Disease Control. Diversity, 15(3), 457. https://doi.org/10.3390/d15030457