
Microeukaryotic Communities of the Long-Term Ice-Covered Freshwater Lakes in the Subarctic Region of Yakutia, Russia

 , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Environmental Parameters
2.2. Amplicon Library Preparation, Sequencing, Raw Data Processing and Quality Control
2.3. Bioinformatics, Statistical Analyses, and Data Visualization
3. Results
3.1. Environmental Parameters at Sampling Sites
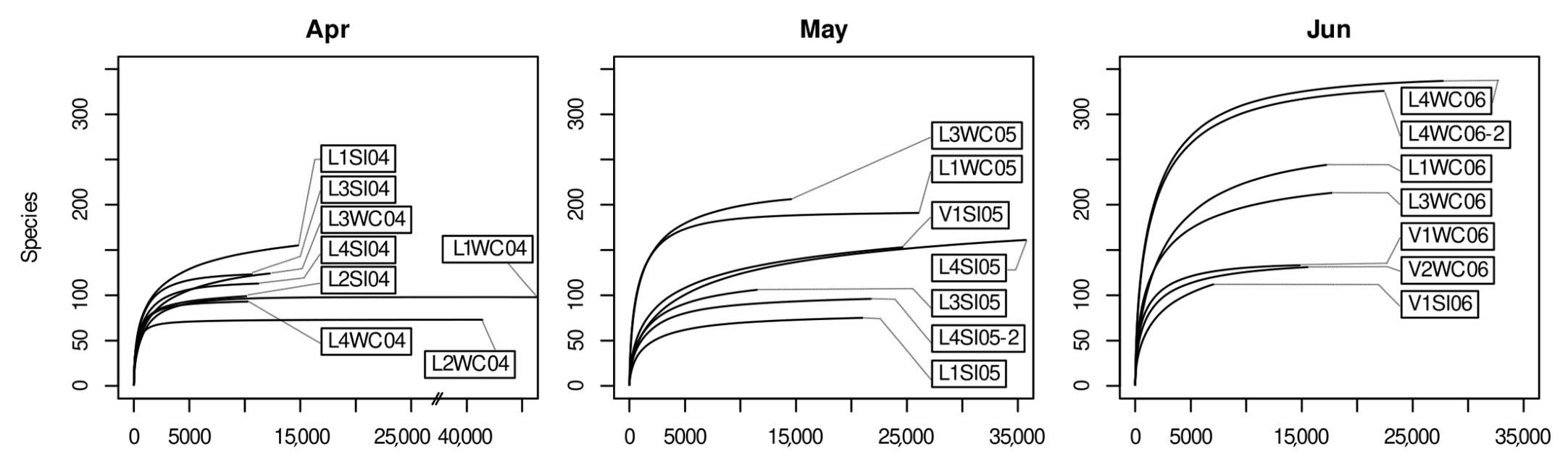
3.2. Quality Control of the Amplicon Clustering Results and Rarefaction Analysis
3.3. Factors Influencing the Hydrophysical and, Hydrochemical Characteristics of Samples and the Alpha-Diversity Metrics of Microeukaryotic Communities
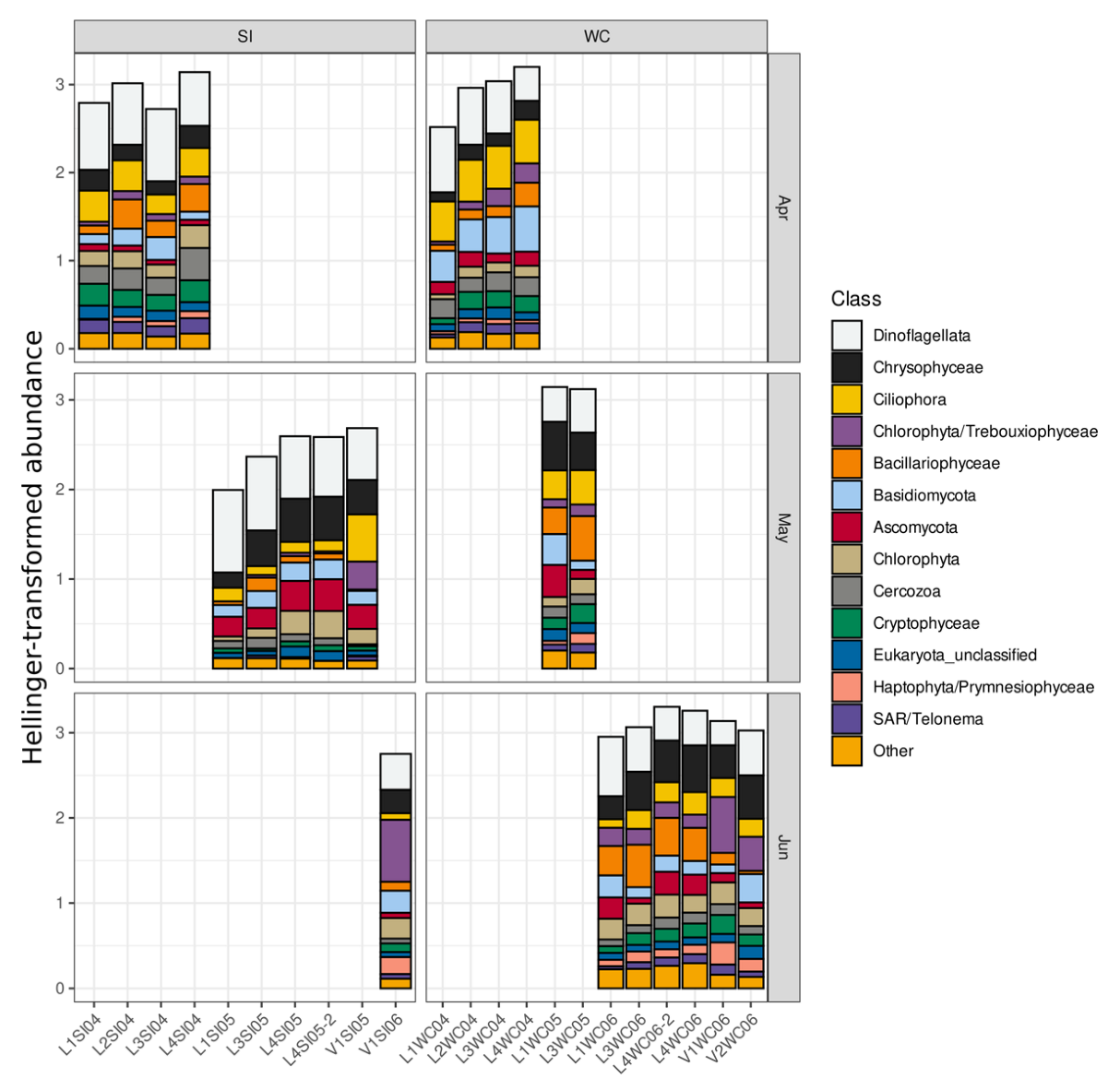
3.4. Overall Community Composition and Comparison of Microeukaryotic Profiles
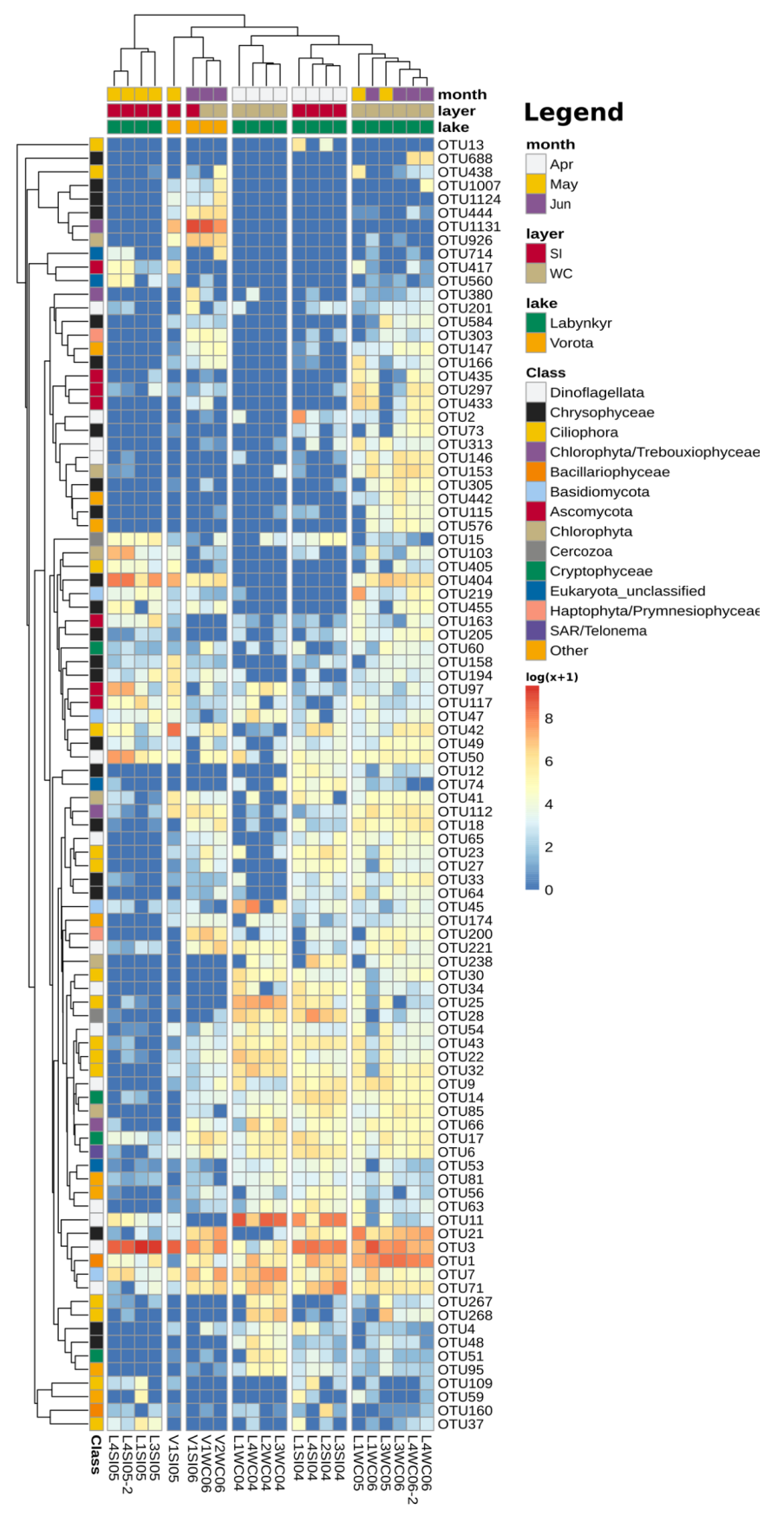
3.5. Differentially Abundant OTUs of LL Communities in April
3.6. Differentially Abundant OTUs of LL Communities in May–June
3.7. Microeukaryotic Communities of LV and Their Comparison with LL Communities
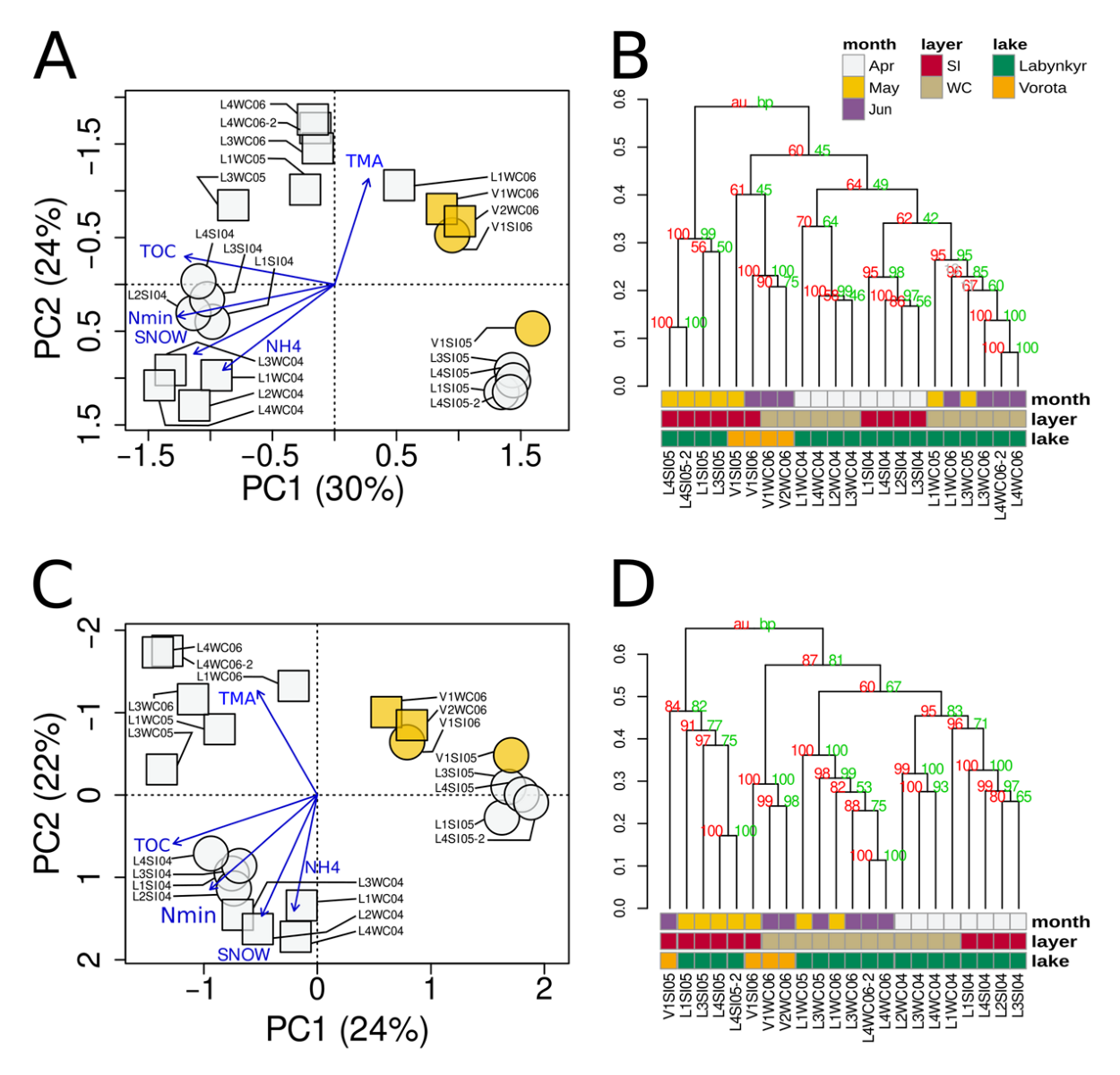
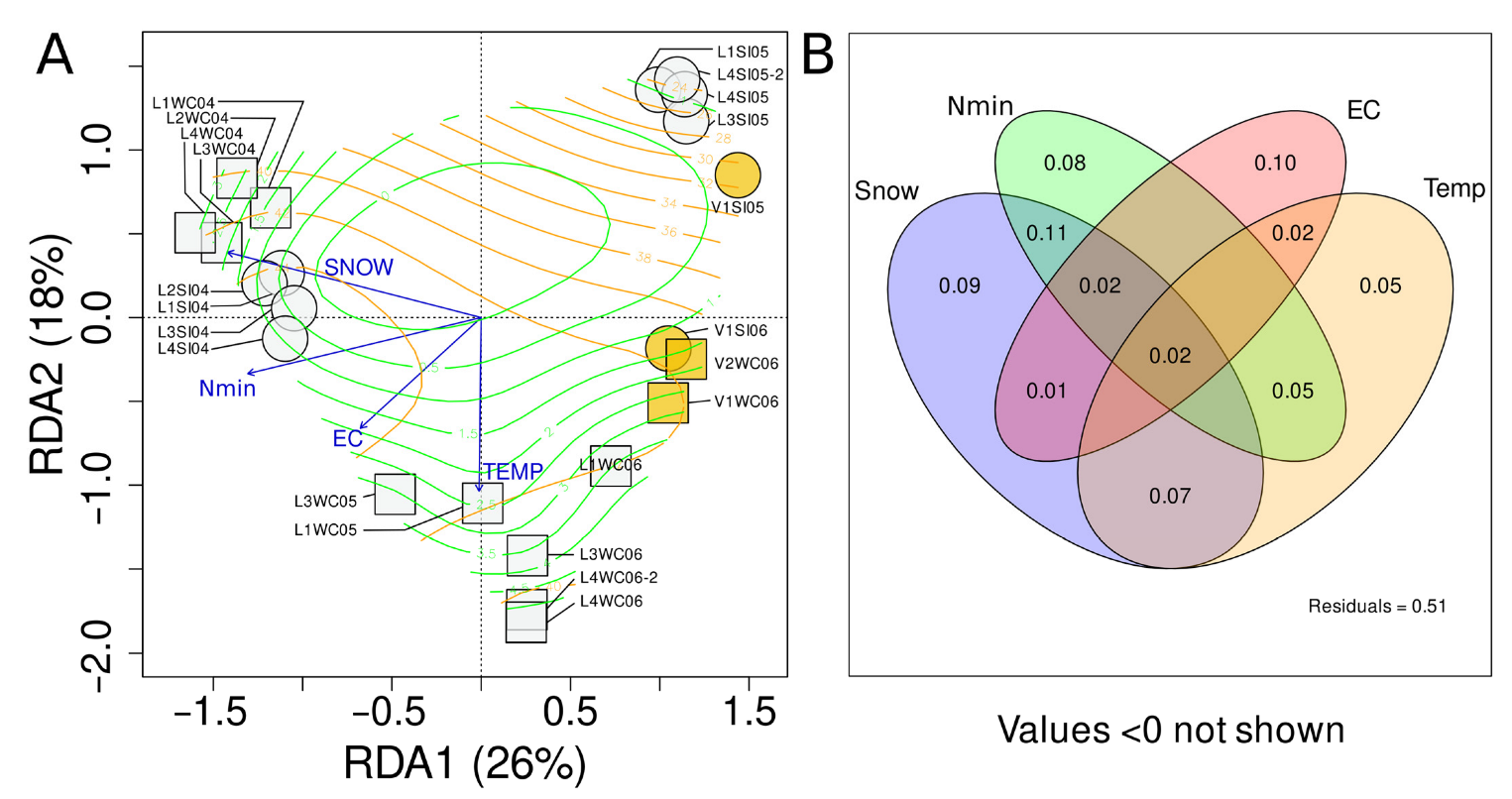
3.8. Environmental Factors as Quantitative Explanatory Variables of Microeukaryotic Community Structure
3.9. Consistency of Results from the Point of View of Technical Replicates
4. Discussion
4.1. Environmental Factors Influencing the Spatial Differences and Temporal Changes in LL Microeukaryotic Community Composition
4.2. Seasonal Dynamics of Microeukaryotic Community Structure
4.3. Peculiarities of the Specific Microeukaryotic High-Rank Phylotypes in LL and LV
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Debroas, D.; Domaizon, I.; Humbert, J.-F.; Jardillier, L.; Lepère, C.; Oudart, A.; Taïb, N. Overview of freshwater microbial eukaryotes diversity: A first analysis of publicly available metabarcoding data. FEMS Microbiol. Ecol. 2017, 93, fix023. [Google Scholar] [CrossRef]
- Schartau, A.K.; Mariash, H.L.; Christoffersen, K.S.; Bogan, D.; Dubovskaya, O.P.; Fefilova, E.B.; Hayden, B.; Ingvason, H.R.; Ivanova, E.A.; Kononova, O.N.; et al. First circumpolar assessment of Arctic freshwater phytoplankton and zooplankton diversity: Spatial patterns and environmental factors. Freshw. Biol. 2022, 67, 141–158. [Google Scholar] [CrossRef]
- Larsen, A.S.; O’Donnell, J.A.; Schmidt, J.H.; Kristenson, H.J.; Swanson, D.K. Physical and chemical characteristics of lakes across heterogeneous landscapes in arctic and subarctic Alaska. J. Geophys. Res. Biogeosci. 2017, 122, 989–1008. [Google Scholar] [CrossRef]
- Arzhakova, S.K.; Zhirkov, I.I.; Kusatov, K.I.; Androsov, I.M. A Quick-Reference Book on Rivers and Lakes of Yakutia; Bichik: Yakutsk, Russia, 2007; pp. 1–136. (In Russian) [Google Scholar]
- Bessudova, A.U.; Tomberg, I.V.; Firsova, A.D.; Kopyrina, L.I.; Likhoshway, Y.V. Silica-scaled chrysophytes in lakes Labynkyr and Vorota, of the Sakha (Yakutia) Republic, Russia. Nova Hedwigua 2019, 148, 35–48. [Google Scholar] [CrossRef]
- Kopyrina, L.; Firsova, A.; Rodionova, E.; Zakharova, Y.; Bashenkhaeva, M.; Usoltseva, M.; Likhoshway, Y. The insight into diatom diversity, ecology, and biogeography of an extreme cold ultraoligotrophic Lake Labynkyr at the Pole of Cold in the Northern Hemisphere. Extremophiles 2020, 24, 603–623. [Google Scholar] [CrossRef]
- Usoltseva, M.; Kopyrina, L.; Titova, L.; Morozov, A.; Firsova, A.; Zakharova, Y.; Bashenkhaeva, M.; Maslennikova, M.; Likhoshway, Y. Finding of a putative Lake Baikal endemic, Lindavia minuta, in distant lakes near the Arctic pole in Yakutia (Russia). Diatom Res. 2020, 35, 141–153. [Google Scholar] [CrossRef]
- Zakharova, Y.R.; Bedoshvili, Y.D.; Petrova, D.P.; Marchenkov, A.M.; Volokitina, N.A.; Bashenkhaeva, M.V.; Kopyrina, L.I.; Grachev, M.A.; Likhoshway, Y.V. Morphological description and molecular phylogeny of two diatom clones from the genus Ulnaria (Kützing) Compère isolated from an ultraoligotrophic lake at the Pole of Cold in the Northern Hemisphere, Republic of Sakha (Yakutia), Russia. Cryptogam. Algol. 2020, 41, 37–45. [Google Scholar] [CrossRef]
- Bessudova, A.; Firsova, A.; Bukin, Y.; Kopyrina, L.; Zakharova, Y.; Likhoshway, Y. Under-ice development of scaled chrysophytes with different trophic mode in two ultraoligotrophic lakes of Yakutia. Diversity 2023, 15, 326. [Google Scholar] [CrossRef]
- Zakharova, Y.; Bashenkhaeva, M.; Galachyants, Y.; Petrova, D.; Tomberg, I.; Marchenkov, A.; Kopyrina, L.; Likhoshway, Y. Variability of microbial communities in two long-term ice-covered freshwater lakes in the subarctic region of Yakutia, Russia. Microb. Ecol. 2022, 84, 958–973. [Google Scholar] [CrossRef]
- Hampton, S.E.; Galloway, A.W.E.; Powers, S.M.; Ozersky, T.; Woo, K.H.; Batt, R.D.; Labou, S.G.; O’Reilly, C.M.; Sharma, S.; Lottig, N.R.; et al. Ecology under lake ice. Ecol. Lett. 2017, 20, 98–101. [Google Scholar] [CrossRef]
- Obertegger, U.; Flaim, G.; Corradini, S.; Cerasino, L.; Zohary, T. Multi-annual comparisons of summer and under-ice phytoplankton communities of a mountain lake. Hydrobiologia 2022, 849, 4613–4635. [Google Scholar] [CrossRef]
- Jansen, J.; MacIntyre, S.; Barrett, D.C.; Chin, Y.-P.; Cortés, A.; Forrest, A.L.; Hrycik, A.R.; Martin, R.; McMeans, B.C.; Rautio, M.; et al. Winter limnology: How do hydrodynamics and biogeochemistry shape ecosystems under ice? J. Geophys. Res. Biogeosci. 2021, 126, e2020JG006237. [Google Scholar] [CrossRef]
- Bashenkhaeva, M.V.; Zakharova, Y.R.; Petrova, D.P.; Khanaev, I.V.; Galachyants, Y.P.; Likhoshway, Y.V. Sub-ice microalgal and bacterial communities in freshwater Lake Baikal, Russia. Microb. Ecol. 2015, 70, 751–765. [Google Scholar] [CrossRef]
- Bashenkhaeva, M.V.; Zakharova, Y.R.; Galachyants, Y.P.; Khanaev, I.V.; Likhoshway, Y.V. Bacterial communities during the period of massive under-ice dinoflagellate development in Lake Baikal. Microbiology 2017, 86, 524–532. [Google Scholar] [CrossRef]
- Bondarenko, N.A.; Timoshkin, O.A.; Ropstorf, P.; Melnik, N.G. The under-ice and bottom periods in the life cycle of Aulacoseira baicalensis (K. Meyer) Simonsen, a principal Lake Baikal alga. Hydrobiologia 2006, 568, 107–109. [Google Scholar] [CrossRef]
- Bondarenko, N.A.; Belykh, O.I.; Golobokova, L.P.; Artemyeva, O.V.; Logacheva, N.F.; Tikhonova, I.V.; Lipko, I.A.; Kostornova, T.Y.; Parfenova, V.V.; Khodzher, T.V.; et al. Stratified distribution of nutrients and extremophile biota within freshwater ice covering the surface of Lake Baikal. J. Microbiol. 2012, 50, 8–16. [Google Scholar] [CrossRef]
- Bashenkhaeva, M.V.; Galachyants, Y.P.; Khanaev, I.V.; Sakirko, M.V.; Petrova, D.P.; Likhoshway, Y.V.; Zakharova, Y.R. Comparative analysis of free-living and particle-associated bacterial communities of Lake Baikal during the ice-covered period. J. Great Lakes Res. 2020, 46, 508–518. [Google Scholar] [CrossRef]
- Rusch, D.B.; Halpern, A.L.; Sutton, G.; Heidelberg, K.B.; Williamson, S.; Yooseph, S.; Wu, D.; Eisen, J.A.; Hoffman, J.M.; Remington, K.; et al. The Sorcerer II Global Ocean Sampling Expedition: Northwest Atlantic through Eastern Tropical Pacific. PLoS Biol. 2007, 5, e77. [Google Scholar] [CrossRef] [PubMed]
- Bradley, I.M.; Pinto, A.J.; Guest, J.S. Design and evaluation of Illumina MiSeq-compatible, 18S rRNA gene-specific primers for improved characterization of mixed phototrophic communities. Appl. Environ. Microbiol. 2016, 82, 5878–5891. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahe, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Pruesse, E.; Peplies, J.; Glöckner, F.O. SINA: Accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 2012, 28, 1823–1829. [Google Scholar] [CrossRef]
- Oksanen, J.; Simpson, G.L.; Blanchet, G.F.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package. R Package Version 2.5-6. 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 24 November 2022).
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Suzuki, R.; Terada, Y.; Shimodaira, H. Pvclust: Hierarchical Clustering with p-Values via Multiscale Bootstrap Resampling. R Package Version 2.2-0. 2019. Available online: https://CRAN.R-project.org/package=pvclust (accessed on 25 January 2023).
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Logares, R.; Daugbjerg, N.; Boltovskoy, A.; Kremp, A.; Laybourn-Parry, J.; Rengefors, K. Recent evolutionary diversification of a protist lineage. Environ. Microbiol. 2008, 10, 1231–1243. [Google Scholar] [CrossRef]
- Lefèvre, E.; Bardot, C.; Noël, C.; Carrias, J.-F.; Viscogliosi, E.; Amblard, C.; Sime-Ngando, T. Unveiling fungal zooflagellates as members of freshwater picoeukaryotes: Evidence from a molecular diversity study in a deep meromictic lake. Environ. Microbiol. 2007, 9, 61–71. [Google Scholar] [CrossRef]
- Andersen, R.A.; de Peer, Y.V.; Potter, D.; Sexton, J.P.; Kawachi, M.; LaJeunesse, T. Phylogenetic Analysis of the SSU rRNA from Members of the Chrysophyceae. Protist 1999, 150, 71–84. [Google Scholar] [CrossRef]
- Boenigk, J.; Pfandl, K.; Stadler, P.; Chatzinotas, A. High diversity of the “Spumella-like” flagellates: An investigation based on the SSU rRNA gene sequences of isolates from habitats located in six different geographic regions. Environ. Microbiol. 2005, 7, 685–697. [Google Scholar] [CrossRef]
- Nakamura, K.; Sudo, M. Analysis of bacterial predatory protozoa inhabiting natural environments. J. Jpn. Soc. Civ. Eng. 2012, 68, III_31–III_40. [Google Scholar] [CrossRef]
- Monchy, S.; Sanciu, G.; Jobard, M.; Rasconi, S.; Gerphagnon, M.; Chabé, M.; Cian, A.; Meloni, D.; Niquil, N.; Christaki, U.; et al. Exploring and quantifying fungal diversity in freshwater lake ecosystems using rDNA cloning/sequencing and SSU tag pyrosequencing. Environ. Microbiol. 2011, 13, 1433–1453. [Google Scholar] [CrossRef]
- Tsai, S.-F.; Chen, W.-T.; Chiang, K.-P. Phylogenetic position of the genus Cyrtostrombidium, with a description of Cyrtostrombidium paralongisomum nov. spec. and a redescription of Cyrtostrombidium longisomum Lynn & Gilron, 1993 (Protozoa, Ciliophora) based on live observation, protargol impregnation, and 18S r DNA sequences. J. Eukaryot. Microbiol. 2014, 62, 239–248. [Google Scholar] [CrossRef]
- Gao, F.; Li, J.; Song, W.; Xu, D.; Warren, A.; Yi, Z.; Gao, S. Multi-gene-based phylogenetic analysis of oligotrich ciliates with emphasis on two dominant groups: Cyrtostrombidiids and strombidiids (Protozoa, Ciliophora). Mol. Phylogenet. Evol. 2016, 105, 241–250. [Google Scholar] [CrossRef]
- Nakai, R.; Abe, T.; Baba, T.; Imura, S.; Kagoshima, H.; Kanda, H.; Kohara, Y.; Koi, A.; Niki, H.; Yanagihara, K.; et al. Eukaryotic phylotypes in aquatic moss pillars inhabiting a freshwater lake in East Antarctica, based on 18S rRNA gene analysis. Polar Biol. 2012, 35, 1495–1504. [Google Scholar] [CrossRef]
- Motohashi, K.; Inaba, S.; Anzai, K.; Takamatsu, S.; Nakashima, C. Phylogenetic analyses of japanese species of Phyllosticta sensu stricto. Mycoscience 2009, 50, 291–302. [Google Scholar] [CrossRef]
- Charvet, S.; Vincent, W.F.; Lovejoy, C. Effects of light and prey availability on Arctic freshwater protist communities examined by high-throughput DNA and RNA sequencing. FEMS Microbiol. Ecol. 2014, 88, 550–564. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, K.H. Introduction to the biology and ecology of the freshwater cryophilic dinoflagellate Woloszynskia pascheri causing red ice. Hydrobiologia 2017, 784, 305–319. [Google Scholar] [CrossRef]
- Wolf, M.; Buchheim, M.; Hegewald, E.; Krienitz, L.; Hepperle, D. Phylogenetic position of the Sphaeropleaceae (Chlorophyta). Plant Syst. Evol. 2002, 230, 161–171. [Google Scholar] [CrossRef]
- Pröschold, T.; Darienko, T. Choricystis and Lewiniosphaera gen. nov. (Trebouxiophyceae, Chlorophyta), two different green algal endosymbionts in freshwater sponges. Symbiosis 2020, 82, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Kulakova, N.V.; Kaskin, S.A.; Bukin, Y.S. The genetic diversity and phylogeny of green microalgae in the genus Choricystis (Trebouxiophyceae, Chlorophyta) in Lake Baikal. Limnology 2020, 21, 15–24. [Google Scholar] [CrossRef]
- Votintsev, K.K. Hydrochemistry of Lake Baikal. Proc. Baikal Limnol. St. 1961, 20, 1–312. [Google Scholar]
- Jewson, D.H.; Granin, N.G.; Zhdanov, A.A.; Gnatovsky, R.Y. Effect of snow depth on under-ice irradiance and growth of Aulacoseira baicalensis in Lake Baikal. Aquat. Ecol. 2009, 43, 673–679. [Google Scholar] [CrossRef]
- Granin, N.G.; Jewson, D.H.; Gnatovsky, R.Y.; Levin, L.A.; Zhdanov, A.A.; Gorbunova, L.A.; Tsekhanovsky, V.V.; Doroschenko, L.M.; Mogilev, N.Y. Turbulent mixing under ice and the growth of diatoms in Lake Baikal. SIL Proc. 1922–2010 2000, 275, 2812–2814. [Google Scholar] [CrossRef]
- Dagenais-Bellefeuille, S.; Morse, D. Putting the N in dinoflagellates. Front. Microbiol. 2013, 4, 369. [Google Scholar] [CrossRef] [PubMed]
- Terrado, R.; Pasulka, A.; Lie, A.Y.; Orphan, V.J.; Heidelberg, K.B.; Caron, D.A. Autotrophic and heterotrophic acquisition of carbon and nitrogen by a mixotrophic chrysophyte established through stable isotope analysis. ISME J. 2017, 11, 2022–2034. [Google Scholar] [CrossRef] [PubMed]
- Bessudova, A.; Bukin, Y.; Likhoshway, Y. Dispersal of Silica-Scaled Chrysophytes in Northern Water Bodies. Diversity 2021, 13, 284. [Google Scholar] [CrossRef]
- Charvet, S.; Vincent, W.F.; Comeau, A.; Lovejoy, C. Pyrosequencing analysis of the protist communities in a High Arctic meromictic lake: DNA preservation and change. Front. Microbiol. 2012, 3, 422. [Google Scholar] [CrossRef]
- Spilling, K. Dense sub-ice bloom of dinoflagellates in the Baltic Sea, potentially limited by high pH. J. Plankton Res. 2007, 29, 895–901. [Google Scholar] [CrossRef]
- Phillips, K.A.; Fawley, M.W. Winter phytoplankton blooms under ice associated with elevated oxygen levels. J. Phycol. 2002, 38, 1068–1073. [Google Scholar] [CrossRef]
- Różańska, M.; Gosselin, M.; Poulin, M.; Wiktor, J.M.; Michel, C. Influence of environmental factors on the development of bottom ice protist communities during the winter–spring transition. Mar. Ecol. Prog. Ser. 2009, 386, 43–59. [Google Scholar] [CrossRef]
- Kirchner, M.; Sahling, G.; Uhlig, G.; Gunkel, W.; Klings, K.-W. Does the red tide-forming dinoflagellate Noctiluca scintillans feed on bacteria? Sarsia 1996, 81, 45–55. [Google Scholar] [CrossRef]
- Obolkina, L.A. Planktonic ciliates of Lake Baikal. Hydrobiologia 2006, 568, 193–199. [Google Scholar] [CrossRef]
- Mikhailov, I.S.; Galachyants, Y.P.; Bukin, Y.S.; Petrova, D.P.; Bashenkhaeva, M.V.; Sakirko, M.V.; Blinov, V.V.; Titova, L.A.; Zakharova, Y.R.; Likhoshway, Y.V. Seasonal succession and coherence among bacteria and microeukaryotes in Lake Baikal. Microb. Ecol. 2022, 84, 404–422. [Google Scholar] [CrossRef]
- Posch, T.; Eugster, B.; Pomati, F.; Pernthaler, J.; Pitsch, G.; Eckert, E.M. Network of interactions between ciliates and phytoplankton during spring. Front. Microbiol. 2015, 6, 1289. [Google Scholar] [CrossRef] [PubMed]
- Tirok, K.; Gaedke, U. Regulation of planktonic ciliate dynamics and functional composition during spring in Lake Constance. Aquat. Microb. Ecol. 2007, 49, 87–100. [Google Scholar] [CrossRef]
- Twiss, M.R.; McKay, R.M.L.; Bourbonniere, R.A.; Bullerjahn, G.S.; Carrick, H.J.; Smith, R.E.H.; Winter, J.G.; D’souza, N.A.; Furey., P.C.; Lashaway, A.R.; et al. Diatoms abound in ice-covered Lake Erie: An investigation of offshore winter limnology in Lake Erie over the period 2007 to 2010. J. Great Lakes Res. 2012, 38, 18–30. [Google Scholar] [CrossRef]
- Bertilsson, S.; Burgin, A.; Carey, C.C.; Fey, S.B.; Grossart, H.P.; Grubisic, L.M.; Jones, I.D.; Kirillin, G.; Lennon, J.T.; Shade, A.; et al. The under-ice microbiome of seasonally frozen lakes. Limnol. Oceanogr. 2013, 58, 1998–2012. [Google Scholar] [CrossRef]
- D’souza, N.A.; Kawarasaki, Y.; Ganyz, J.D., Jr.; Lee, R.E.; Beall, B.F.N.; Shtarkman, Y.M.; Koçer, Z.A.; Rogers, S.O.; Wildschutte, H.; Bullerjahn, G.S.; et al. Diatom assemblages promote ice formation in large lakes. ISME J. 2013, 7, 1632–1640. [Google Scholar] [CrossRef]
- Dumack, K.; Pundt, J.; Bonkowski, M. Food choice experiments indicate selective fungivorous predation in fisculla terrestris (Thecofilosea, Cercozoa). J. Eukaryot. Microbiol. 2019, 66, 525–527. [Google Scholar] [CrossRef]
- Tsuji, M.; Tsujimoto, M.; Imura, S. Cystobasidium tubakii and Cystobasidium ongulense, new basidiomycetous yeast species isolated from East Ongul Island, East Antarctica. Mycoscience 2017, 58, 103–110. [Google Scholar] [CrossRef]
- Batista, T.M.; Hilário, H.O.; de Brito, G.A.M.; Moreira, R.G.; Furtado, C.; de Menezes, G.C.A.; Rosa, C.A.; Rosa, L.H.; Franco, G.R. Whole-genome sequencing of the endemic Antarctic fungus Antarctomyces pellizariae reveals an ice-binding protein, a scarce set of secondary metabolites gene clusters and provides insights on Thelebolales phylogeny. Genomics 2020, 112, 2915–2921. [Google Scholar] [CrossRef] [PubMed]
- Grujcic, V.; Nuy, J.K.; Salcher, M.M.; Shabarova, T.; Kasalicky, V.; Boenigk, J.; Jensen, M.; Simek, K. Cryptophyta as major bacterivores in freshwater summer plankton. ISME J. 2018, 12, 1668–1681. [Google Scholar] [CrossRef] [PubMed]
- Henshaw, T.; Laybourn-Parry, J. The annual patterns of photosynthesis in two large, freshwater, ultra-oligotrophic Antarctic lakes. Polar Biol. 2002, 25, 744–752. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | Sample | Temp | pH | EC | DO | PO43− | NH4+ | NO2− | NO3− | Nmin | TDS | TOC | TMA | TMB | Snow | Ice |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| April | L1SI04 | 0.4 | 8.28 | 49 | 10.6 | 0.001 | 0.021 | 0.002 | 0.35 | 0.1 | 33.19 | 3.26 | 19.57 | 0.07 | 30 | 91 |
| L1WC04 | 1.2 | 7.84 | 43.22 | 10.6 | 0.003 | 0.059 | 0.002 | 0.3 | 0.12 | 29.29 | 2.58 | 13.7 | 0.03 | 30 | 91 | |
| L2SI04 | 1.2 | 7.71 | 45.98 | 7.5 | 0.001 | 0.062 | 0.002 | 0.34 | 0.13 | 31.82 | 2.88 | 24.1 | 0.06 | 32 | 86 | |
| L2WC04 | 2.6 | 7.93 | 39.49 | 7.5 | 0.002 | 0.058 | 0.001 | 0.29 | 0.11 | 26.67 | 2.21 | 35.95 | 0.01 | 32 | 86 | |
| L3SI04 | 0.4 | 7.55 | 44.92 | 8.9 | 0.001 | 0.014 | 0.002 | 0.33 | 0.08 | 30.72 | 3.45 | 20.95 | 0.02 | 30 | 109 | |
| L3WC04 | 1.9 | 7.93 | 39.49 | 8.9 | 0.002 | 0.058 | 0.001 | 0.29 | 0.11 | 26.67 | 2.21 | 28.85 | 0.03 | 30 | 109 | |
| L4SI04 | 0.4 | 7.48 | 50.1 | 8.2 | 0.004 | 0.019 | 0.001 | 0.42 | 0.11 | 33.96 | 3.04 | 40.4 | 0.04 | 35 | 86 | |
| L4WC04 | 3.7 | 7.7 | 39.85 | 8.2 | 0.001 | 0.069 | 0.002 | 0.28 | 0.12 | 26.54 | 3.04 | 39.55 | 0.13 | 35 | 86 | |
| May | L1SI05 | 0.5 | 8.78 | 38.41 | 8.5 | 0.009 | 0.013 | 0.002 | 0.33 | 0.08 | 25.33 | 1.45 | 15.75 | 0.01 | 5 | 111 |
| L1WC05 | 2.6 | 9.34 | 45.72 | 8.5 | 0.003 | 0.012 | 0.002 | 0.34 | 0.09 | 30.3 | 2.14 | 222.23 | 0.08 | 5 | 111 | |
| L3SI05 | 1.2 | 7.81 | 23.34 | 9.8 | 0.006 | 0.007 | 0.003 | 0.19 | 0.05 | 17.16 | 1.99 | 34.4 | 0.03 | 0.5 | 110 | |
| L3WC05 | 3.4 | 9.26 | 40.76 | 9.8 | 0.002 | 0.016 | 0.001 | 0.2 | 0.09 | 27.71 | 2.66 | 313.67 | 0.03 | 0.5 | 110 | |
| L4SI05 | 1.3 | 6.97 | 8.517 | 9.6 | 0 | 0.009 | 0.002 | 0.17 | 0.05 | 6.56 | 1.5 | 52.56 | 0.06 | 0.5 | 86 | |
| V1SI05 | 0.4 | 8.94 | 41.48 | 8.5 | 0.008 | 0.001 | 0.003 | 0.1 | 0.02 | 31.74 | 1.24 | 192.7 | 0.03 | 1 | 100 | |
| V1WC05 | 3.1 | 8.82 | 51.21 | 8.5 | 0.012 | 0.004 | 0 | 0.08 | 0.02 | 42.47 | 0.86 | 79.55 | 0.08 | 1 | 100 | |
| June | L1WC06 | 3.6 | 6.8 | 30.89 | 8.3 | 0.003 | 0.017 | 0.003 | 0.33 | 0.09 | - | 1.05 | 173.3 | 0.02 | 5 | 110 |
| L3WC06 | 2.5 | 6.97 | 37.71 | 7.3 | 0.003 | 0.012 | 0.003 | 0.38 | 0.09 | - | 0.86 | 127.1 | 0.03 | 1 | 80 | |
| L4WC06 | 5.6 | 6.98 | 41.07 | 8.4 | 0.016 | 0.012 | 0.005 | 0.45 | 0.11 | - | 0.86 | 121.3 | 0.05 | 0.5 | 86 | |
| V1SI06 | 0.4 | 7.15 | 49.55 | 6.7 | 0.014 | 0.015 | 0.003 | 0.05 | 0.02 | - | 0.83 | 55.67 | 0.02 | 5 | 110 | |
| V1WC06 | 3.2 | 7.21 | 54.75 | 6.7 | 0.023 | 0.016 | 0.006 | 0.06 | 0.02 | - | 1.84 | 95.13 | 0.04 | 5 | 110 | |
| V2WC06 | 2.2 | 7.1 | 33.8 | 8.3 | 0.014 | 0.016 | 0.003 | 0.04 | 0.03 | - | 0.75 | 84.91 | 0.02 | 5 | 83 |
| Sample | Reads | Richness | ACE | Shannon | Simpson | Inverse Simpson |
|---|---|---|---|---|---|---|
| L1SI04 | 14,781 | 155 | 168.37 | 3.18 | 0.90 | 10.11 |
| L1WC04 | 51,984 | 98 | 98.00 | 2.54 | 0.80 | 5.05 |
| L2SI04 | 10,123 | 99 | 102.69 | 3.21 | 0.92 | 11.95 |
| L2WC04 | 45,914 | 73 | 73.00 | 2.74 | 0.85 | 6.67 |
| L3SI04 | 12,274 | 124 | 133.40 | 2.69 | 0.86 | 7.34 |
| L3WC04 | 11,225 | 113 | 114.46 | 3.05 | 0.89 | 9.26 |
| L4SI04 | 10,601 | 123 | 124.47 | 3.10 | 0.90 | 9.53 |
| L4WC04 | 10,258 | 93 | 93.70 | 3.30 | 0.94 | 15.55 |
| L1SI05 | 20,979 | 75 | 77.40 | 0.99 | 0.31 | 1.45 |
| L1WC05 | 26,057 | 191 | 191.91 | 3.64 | 0.93 | 14.74 |
| L3SI05 | 11,460 | 106 | 114.76 | 1.62 | 0.54 | 2.19 |
| L3WC05 | 14,548 | 206 | 212.88 | 3.48 | 0.91 | 11.04 |
| L4SI05 * | 35,709 | 161 | 177.54 | 2.25 | 0.81 | 5.15 |
| L4SI05-2 * | 21,734 | 96 | 98.73 | 2.23 | 0.82 | 5.41 |
| V1SI05 | 17,205 | 244 | 257.79 | 2.90 | 0.82 | 5.64 |
| V1WC05 | 531 | - | - | - | - | - |
| L1WC06 | 17,685 | 213 | 224.49 | 3.31 | 0.90 | 9.74 |
| L3WC06 | 27,683 | 337 | 340.35 | 4.16 | 0.95 | 21.76 |
| L4WC06 * | 22,401 | 326 | 331.59 | 4.05 | 0.94 | 17.57 |
| L4WC06-2 * | 24,580 | 153 | 171.89 | 2.42 | 0.81 | 5.35 |
| V1SI06 | 7032 | 112 | 134.98 | 2.27 | 0.75 | 3.99 |
| V1WC06 | 14,788 | 133 | 134.42 | 2.91 | 0.82 | 5.58 |
| V2WC06 | 15,535 | 131 | 135.60 | 3.09 | 0.92 | 11.78 |
| Indep * Var #1 | Indep Var #2 | Dep Var | ANOVA | Kruskal–Wallis | ||||
|---|---|---|---|---|---|---|---|---|
| p | padj | padj Sign | p | padj | padj Sign | |||
| Environmental variables | ||||||||
| Lake | - | PO4 | 2.64 × 10−4 | 1.23 × 10−3 | ** | 7.56 × 10−3 | 3.53 × 10−2 | * |
| Lake | - | NO2 | 1.81 × 10−2 | 6.33 × 10−2 | . | 1.48 × 10−2 | 5.19 × 10−2 | . |
| Lake | - | NO3 | 6.52 × 10−6 | 5.49 × 10−5 | *** | 2.44 × 10−3 | 1.71 × 10−2 | * |
| Lake | - | totN | 7.84 × 10−6 | 5.49 × 10−5 | *** | 2.25 × 10−3 | 1.71 × 10−2 | * |
| Lake | - | TOC | 3.39 × 10−2 | 9.49 × 10−2 | . | 2.32 × 10−2 | 6.49 × 10−2 | . |
| Month | - | pH | 7.36 × 10−4 | 2.49 × 10−3 | ** | 5.73 × 10−3 | 1.61 × 10−2 | * |
| Month | - | DO | 3.76 × 10−2 | 5.84 × 10−2 | . | 2.71 × 10−2 | 4.21 × 10−2 | * |
| Month | - | PO4 | 2.92 × 10−3 | 6.81 × 10−3 | ** | 1.22 × 10−2 | 2.13 × 10−2 | * |
| Month | - | NH4 | 6.06 × 10−4 | 2.49 × 10−3 | ** | 1.86 × 10−3 | 7.58 × 10−3 | ** |
| Month | - | NO2 | 8.88 × 10−4 | 2.49 × 10−3 | ** | 2.16 × 10−3 | 7.58 × 10−3 | ** |
| Month | - | Nmin | 6.54 × 10−3 | 1.31 × 10−2 | * | 7.66 × 10−3 | 1.79 × 10−2 | * |
| Month | - | TOC | 7.00 × 10−6 | 4.90 × 10−5 | *** | 6.49 × 10−4 | 4.54 × 10−3 | ** |
| Month | - | TMA | 2.20 × 10−2 | 3.86 × 10−2 | * | 1.21 × 10−2 | 2.13 × 10−2 | * |
| Month | - | Snow | 4.15 × 10−15 | 5.82 × 10−14 | *** | 6.03 × 10−4 | 4.54 × 10−3 | ** |
| Layer | - | Temp | 2.81 × 10−5 | 3.93 × 10−4 | *** | 2.69 × 10−4 | 3.77 × 10−3 | ** |
| WC | Month | pH | 8.98 × 10−8 | 2.52 × 10−6 | *** | 1.34 × 10−2 | 9.87 × 10−2 | . |
| SI | Month | PO4 | 2.89 × 10−2 | 7.35 × 10−2 | . | 1.66 × 10−1 | 2.74 × 10−1 | . |
| WC | Month | NH4 | 1.87 × 10−7 | 2.53 × 10−6 | *** | 2.52 × 10−2 | 9.87 × 10−2 | . |
| WC | Month | NO2 | 1.64 × 10−2 | 4.72 × 10−2 | * | 1.89 × 10−2 | 9.87 × 10−2 | . |
| SI | Month | NO3 | 1.54 × 10−2 | 4.72 × 10−2 | * | 4.50 × 10−2 | 9.87 × 10−2 | . |
| SI | Month | Nmin | 1.69 × 10−2 | 4.72 × 10−2 | * | 5.00 × 10−2 | 1.00 × 10−1 | . |
| SI | Month | TOC | 2.52 × 10−4 | 1.17 × 10−3 | ** | 3.57 × 10−2 | 9.87 × 10−2 | . |
| WC | Month | TOC | 1.76 × 10−3 | 7.06 × 10−3 | ** | 2.20 × 10−2 | 9.87 × 10−2 | . |
| WC | Month | TMA | 1.41 × 10−4 | 7.90 × 10−4 | *** | 1.36 × 10−2 | 9.87 × 10−2 | . |
| SI | Month | Snow | 3.99 × 10−6 | 2.80 × 10−5 | *** | 3.80 × 10−2 | 9.87 × 10−2 | . |
| WC | Month | Snow | 2.72 × 10−7 | 2.53 × 10−6 | *** | 2.39 × 10−2 | 9.87 × 10−2 | . |
| Alpha-diversity metrics ** | ||||||||
| Month | - | Richness | 3.95 × 10−2 | 9.87 × 10−2 | . | 1.54 × 10−2 | 3.84 × 10−2 | * |
| Month | - | ACE | 2.98 × 10−2 | 9.87 × 10−2 | . | 1.16 × 10−2 | 3.84 × 10−2 | * |
| Layer | - | Shannon | 3.16 × 10−3 | 1.58 × 10−2 | * | 6.86 × 10−3 | 3.43 × 10−2 | * |
| Layer | - | Simpson | 3.74 × 10−2 | 6.24 × 10−2 | . | 2.50 × 10−2 | 4.16 × 10−2 | * |
| Layer | - | Inverse Simpson | 2.00 × 10−2 | 4.99 × 10−2 | * | 2.50 × 10−2 | 4.16 × 10−2 | * |
| WC | Month | Richness | 2.71 × 10−2 | 6.78 × 10−2 | . | 6.58 × 10−3 | 2.10 × 10−2 | * |
| WC | Month | ACE | 2.30 × 10−2 | 6.78 × 10−2 | . | 6.58 × 10−3 | 2.10 × 10−2 | * |
| SI | Month | Shannon | 6.84 × 10−3 | 3.42 × 10−2 | * | 1.05 × 10−2 | 2.10 × 10−2 | * |
| SI | Month | Inverse Simpson | 1.03 × 10−3 | 1.03 × 10−2 | * | 1.05 × 10−2 | 2.10 × 10−2 | * |
| Factor * | Dep Var | p | padj | padj Sign |
|---|---|---|---|---|
| Environmental variables | ||||
| Layer | Temp | 1.83 × 10−2 | 9.61 × 10−2 | . |
| Month | EC | 1.47 × 10−2 | 8.84 × 10−2 | . |
| Layer | NH4 | 9.94 × 10−4 | 1.39 × 10−2 | * |
| Month | Nmin | 1.01 × 10−2 | 7.05 × 10−2 | . |
| Month | TOC | 1.53 × 10−5 | 3.22 × 10−4 | *** |
| Month:Layer | TOC | 8.00 × 10−3 | 6.72 × 10−2 | . |
| Month:Layer | TMA | 7.64 × 10−3 | 6.72 × 10−2 | . |
| Month | Snow | 1.58 × 10−10 | 6.63 × 10−9 | *** |
| Alpha-diversity metrics ** | ||||
| Month:Layer | Richness | 3.61 × 10−2 | 8.30 × 10−2 | . |
| Month:Layer | ACE | 3.87 × 10−2 | 8.30 × 10−2 | . |
| Month | Shannon | 2.16 × 10−3 | 1.62 × 10−2 | * |
| Month:Layer | Shannon | 8.96 × 10−4 | 1.34 × 10−2 | * |
| Month | Simpson | 8.41 × 10−3 | 4.20 × 10−2 | * |
| Month:Layer | Simpson | 2.57 × 10−2 | 8.19 × 10−2 | . |
| Month | Inverse Simpson | 4.54 × 10−2 | 8.51 × 10−2 | . |
| Month:Layer | Inverse Simpson | 2.73 × 10−2 | 8.19 × 10−2 | . |
| OUT Number | Nearest Match | GenBank Accession | Sequence Identity, % | Source |
|---|---|---|---|---|
| OTU3 | Scrippsiella hangoei strain SHTV6 Peridinium aciculiferum strain PAER-2 | EF417316 EF417314 | 99 | The Gulf of Finland, Baltic Sea Lake Erken, Sweden [29] |
| OTU65 | Uncultured alveolate | DQ244019 | 99 | Lake Pavin, France [30] |
| OTU12 | Chrysolepidomonas dendrolepidota CCMP293Spumella-like isolate JBM19 | AF123297 FR865768 | 98 | Lake Superior, Keeweenaw Country, MI, USA [31] Lake Hallstatt, Austria [32] |
| OTU158 | Bacterivorous protozoa | AB749149 | 97 | Hirose River, Japan [33] |
| OTU13 | Free-living ciliate of class Oligohymenophorea | LR025746 HQ219368 | 98 | Lake Zurich, Switzerland Lake Aydat, France [34] |
| OTU27 | Cyrtostrombidium longisomum | KJ534582 | 97 | Coastal waters of northeastern Taiwan [35] |
| OTU42 | Uncultured ciliate Uncultured ciliate Limnostrombidium viride SW2012122001 | GU067975 LC165025 KU525754 | 99 | Esch-sur-Sure, Luxembourg Biwa, JapanZhanjiang, Guangdong province, China [36] |
| OTU109 | Uncultured eukaryote | AB695505 | 99 | Freshwater lake, East Antarctica [37] |
| OTU1 | Stephanodiscus sp. | AB430594 | 99 | Japan and South Korea [38] |
| OTU21 | Chrysophycean isolate | AY082970 | 94 | High-latitude Arctic lakes, Ellesmere Island [39] |
| OTU50 | Woloszynskia pascheri | EF058253 | 96 | Freshwater environment [40] |
| OTU444 | Paraphysomonas foraminifera strain TPC2 | AY651096 | 99 | Lake Mondsee, Austria [32] |
| OTU926 | Ankyra lanceolata strain Hg 1998-5 | AF302769 | 98 | Channel of Danube, Hungary [41] |
| OTU1131 | Choricystis sp. AS-29, green alga symbiont of sponges | AY195972 | 99 | Freshwater, free-living [42] Lake Baikal, sponge symbiont [43] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galachyants, Y.; Zakharova, Y.; Bashenkhaeva, M.; Petrova, D.; Kopyrina, L.; Likhoshway, Y. Microeukaryotic Communities of the Long-Term Ice-Covered Freshwater Lakes in the Subarctic Region of Yakutia, Russia. Diversity 2023, 15, 454. https://doi.org/10.3390/d15030454
Galachyants Y, Zakharova Y, Bashenkhaeva M, Petrova D, Kopyrina L, Likhoshway Y. Microeukaryotic Communities of the Long-Term Ice-Covered Freshwater Lakes in the Subarctic Region of Yakutia, Russia. Diversity. 2023; 15(3):454. https://doi.org/10.3390/d15030454
Chicago/Turabian StyleGalachyants, Yuri, Yulia Zakharova, Maria Bashenkhaeva, Darya Petrova, Liubov Kopyrina, and Yelena Likhoshway. 2023. "Microeukaryotic Communities of the Long-Term Ice-Covered Freshwater Lakes in the Subarctic Region of Yakutia, Russia" Diversity 15, no. 3: 454. https://doi.org/10.3390/d15030454
APA StyleGalachyants, Y., Zakharova, Y., Bashenkhaeva, M., Petrova, D., Kopyrina, L., & Likhoshway, Y. (2023). Microeukaryotic Communities of the Long-Term Ice-Covered Freshwater Lakes in the Subarctic Region of Yakutia, Russia. Diversity, 15(3), 454. https://doi.org/10.3390/d15030454