

Large Remaining Forest Habitat Patches Help Preserve Wild Bee Diversity in Cultivated Blueberry Bush


Abstract
1. Introduction
2. Materials and Methods
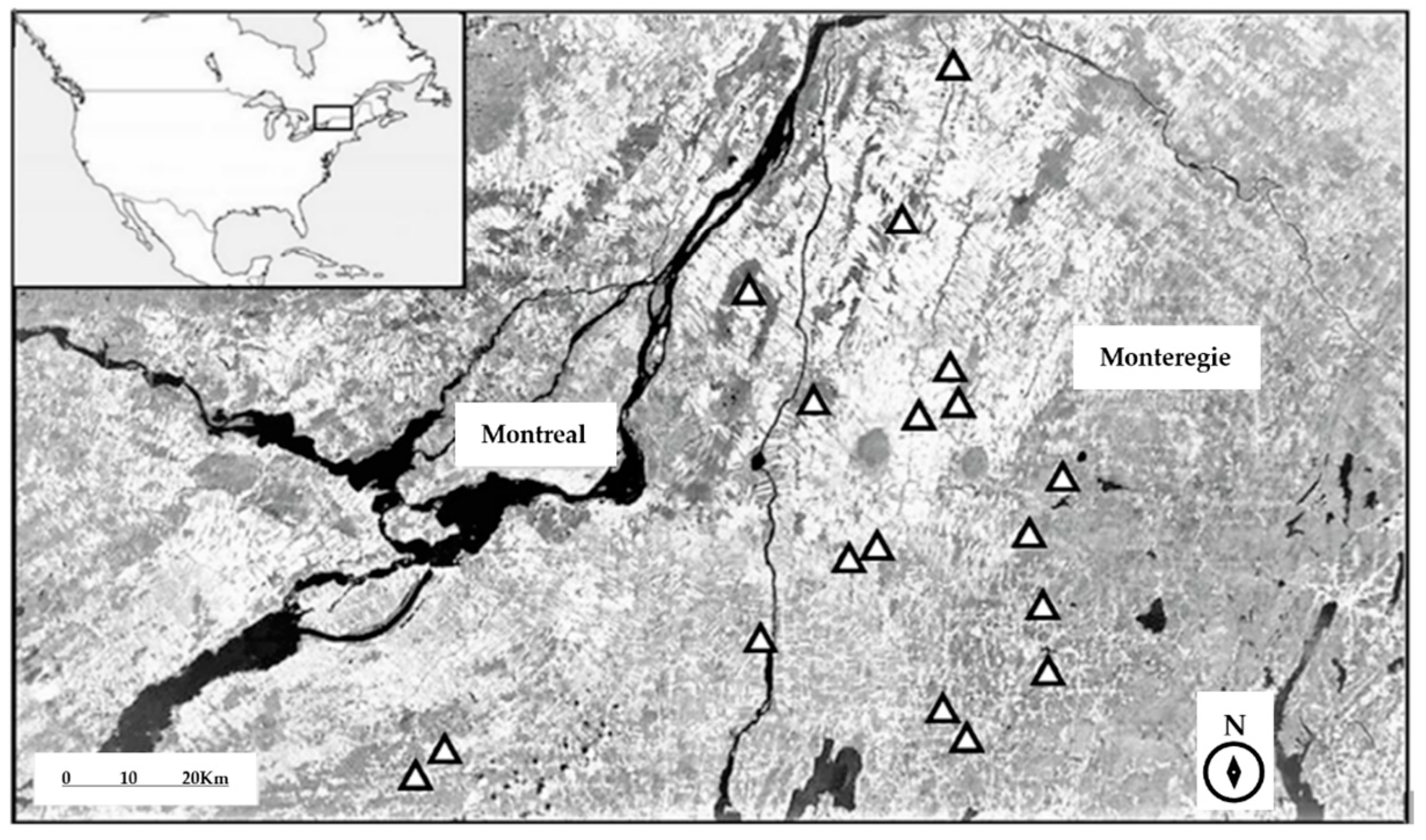
2.1. Study Area
2.2. Wild Bee Sampling
2.3. Environmental Variables
2.4. Surrounding Land Cover
2.5. Statistical Analysis
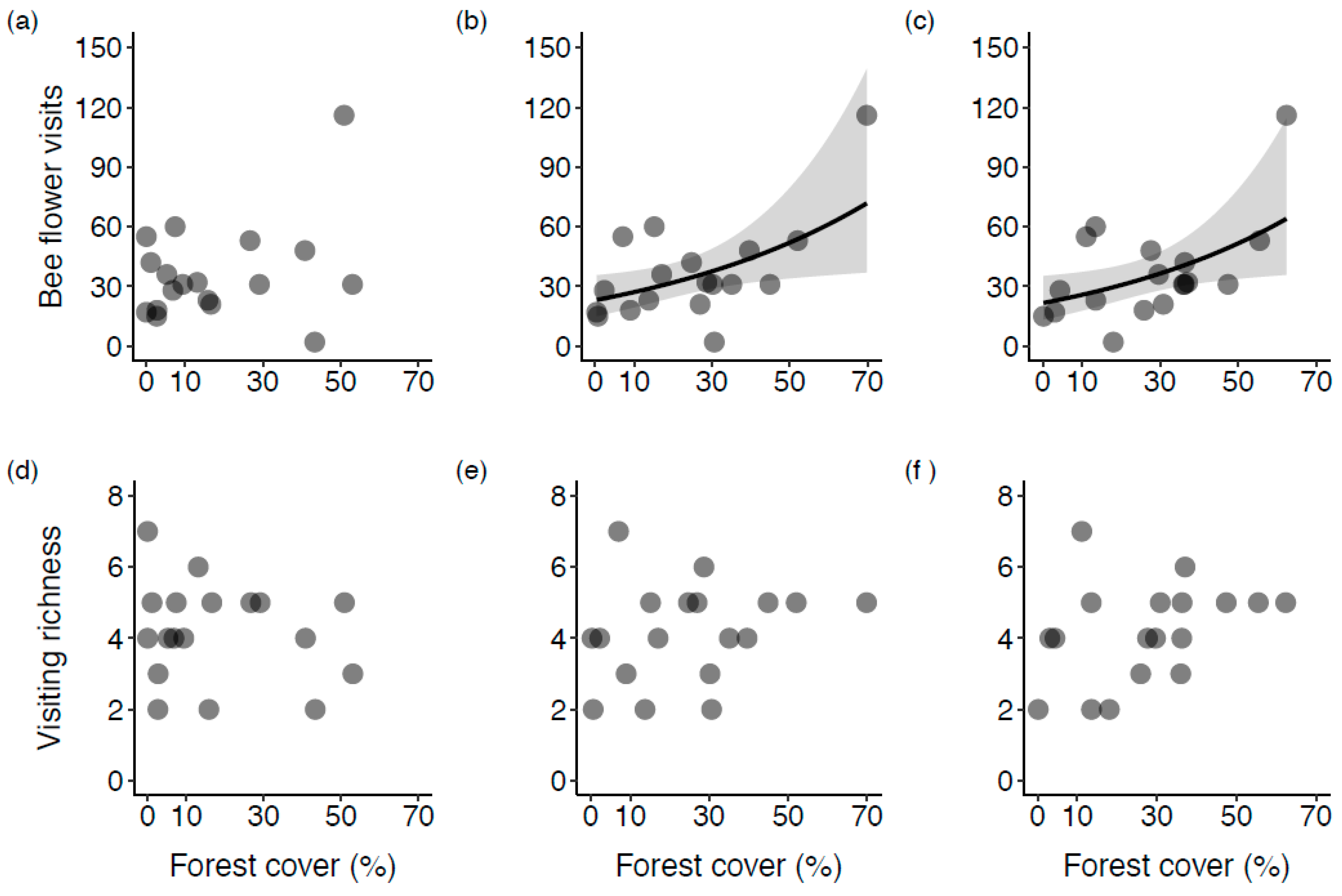
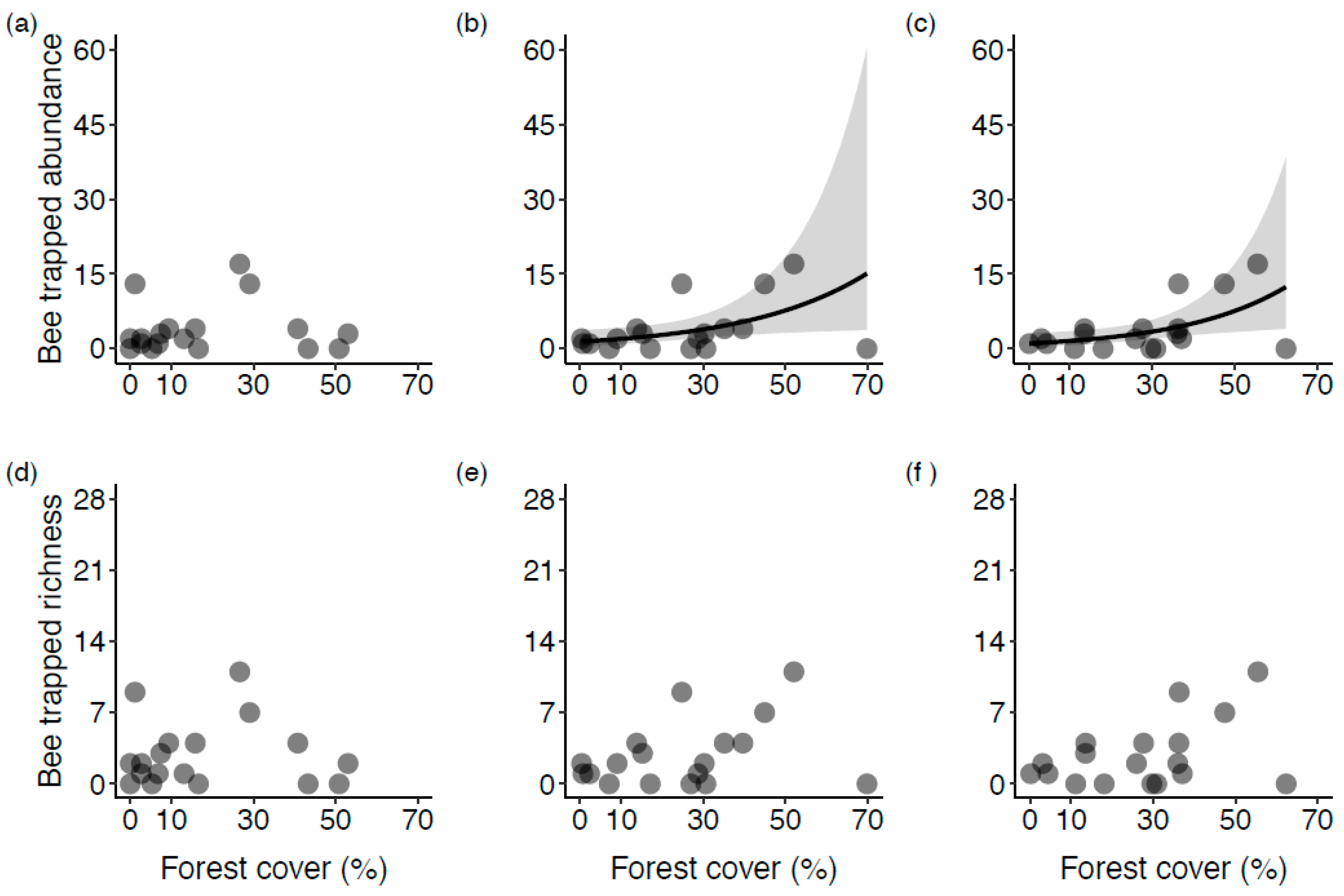
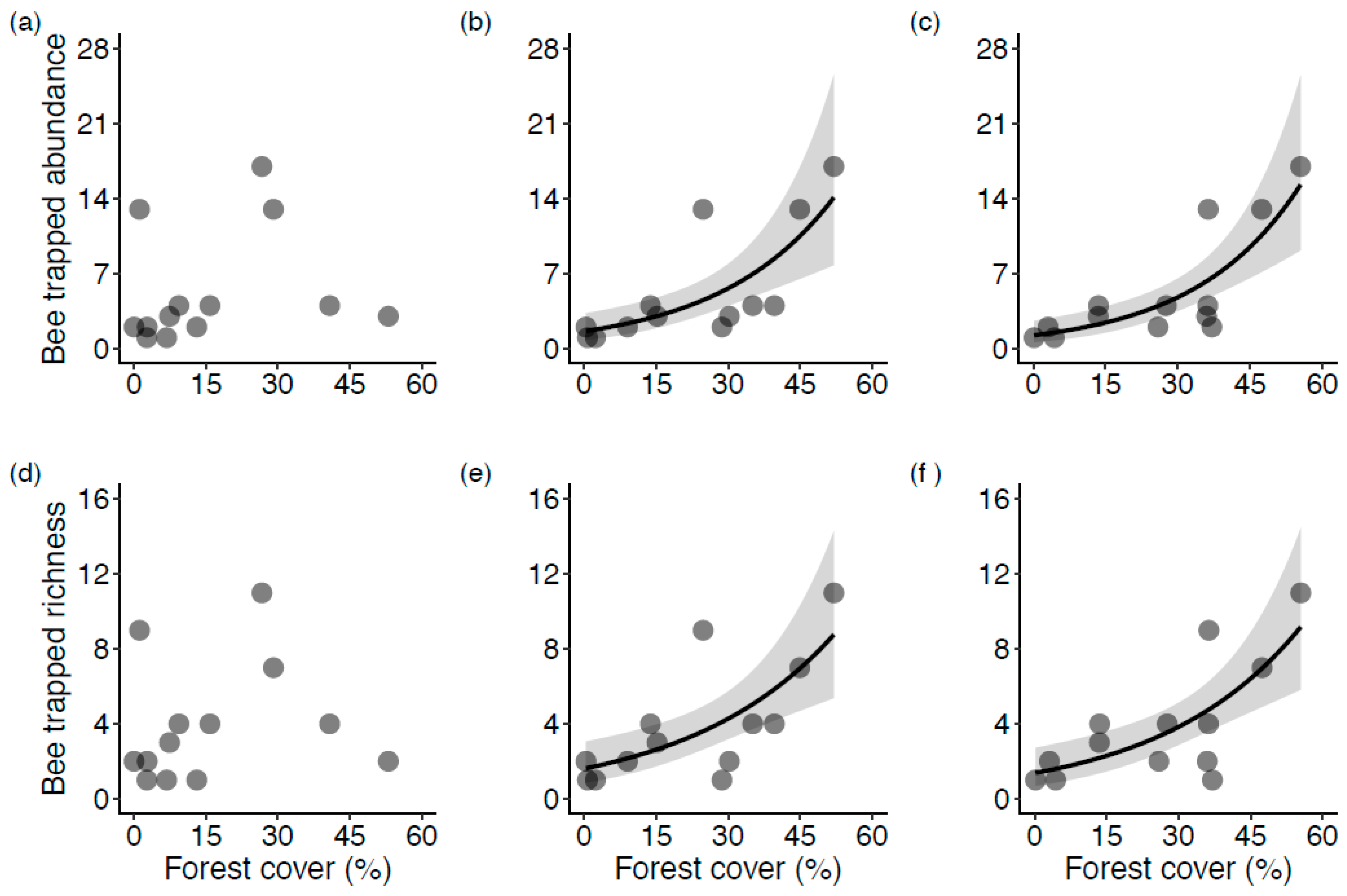
3. Results
3.1. Surrounding Land Cover
3.2. The Influence of Natural Habitat
4. Discussion
4.1. The Effect of Forest Proportion on Wild Bee Diversity in Blueberry Fields
4.2. The Role of Bumble Bees and Incidental Visitors
4.3. Assessing the Effective Spatial Scale of Natural Habitat Preservation
4.4. Landscape Management and Conservation Perspectives
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global Pollinator Declines: Trends, Impacts and Drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Kennedy, C.M.; Lonsdorf, E.; Neel, M.C.; Williams, N.M.; Ricketts, T.H.; Winfree, R.; Bommarco, R.; Brittain, C.; Burley, A.L.; Cariveau, D.; et al. A Global Quantitative Synthesis of Local and Landscape Effects on Wild Bee Pollinators in Agroecosystems. Ecol. Lett. 2013, 16, 584–599. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Kremen, C.; Morales, J.M.; Bommarco, R.; Cunningham, S.A.; Carvalheiro, L.G.; Chacoff, N.P.; Dudenhöffer, J.H.; Greenleaf, S.S.; et al. Stability of Pollination Services Decreases with Isolation from Natural Areas despite Honey Bee Visits. Ecol. Lett. 2011, 14, 1062–1072. [Google Scholar] [CrossRef]
- Hevia, V.; Bosch, J.; Azcárate, F.M.; Fernández, E.; Rodrigo, A.; Barril-Graells, H.; González, J.A. Bee Diversity and Abundance in a Livestock Drove Road and Its Impact on Pollination and Seed Set in Adjacent Sunflower Fields. Agric. Ecosyst. Environ. 2016, 232, 336–344. [Google Scholar] [CrossRef]
- Winfree, R.; Williams, N.M.; Dushoff, J.; Kremen, C. Native Bees Provide Insurance against Ongoing Honey Bee Losses. Ecol. Lett. 2007, 10, 1105–1113. [Google Scholar] [CrossRef]
- Hanley, N.; Breeze, T.D.; Ellis, C.; Goulson, D. Measuring the Economic Value of Pollination Services: Principles, Evidence and Knowledge Gaps. Ecosyst. Serv. 2015, 14, 124–132. [Google Scholar] [CrossRef]
- Klein, A.-M.; Vaissiere, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of Pollinators in Changing Landscapes for World Crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef]
- Williams, N.M.; Kremen, C. Resource Distributions among Habitats Determine Solitary Bee Offspring Production in a Mosaic Landscape. Ecol. Appl. 2007, 17, 910–921. [Google Scholar] [CrossRef]
- Moisan-Deserres, J.; Girard, M.; Chagnon, M.; Fournier, V. Pollen Loads and Specificity of Native Pollinators of Lowbush Blueberry. J. Econ. Entomol. 2014, 107, 1156–1162. [Google Scholar] [CrossRef]
- Statistics Canada. Table 32-10-0364-01 Area, Production and Farm Gate Value of Marketed Fruits. 2021. Available online: https://www150.statcan.gc.ca/t1/tbl1/en/tv.action?pid=3210036401&pickMembers%5B0%5D=1.1&pickMembers%5B1%5D=2.2&pickMembers%5B2%5D=4.5&cubeTimeFrame.startYear=2015&cubeTimeFrame.endYear=2020&referencePeriods=20150101%2C20200101 (accessed on 20 March 2021).
- Daly, K.; Pacheco, M.; Poplack, A.; Johnson, C.; Maxon, M.; Kopec, K.; Cypel, B. Comparing Apis Mellifera and Bombus Spp. Pollination Efficiencies on Willamette Valley Blueberry Farms. Or. Undergrad. Res. J. 2013, 4, 23–34. [Google Scholar] [CrossRef]
- Finnamore, A.T.; Neary, M.E. Blueberry Pollinators of Nova Scotia, with a Checklist of the Blueberry Pollinators in Eastern Canada Northeastern United States. Ann. Société Entomolgique Que. 1978, 433, 309–312. [Google Scholar] [CrossRef]
- Javorek, S.K.; Mackenzie, K.E.; Vander Kloet, S.P. Comparative Pollination Effectiveness Among Bees (Hymenoptera: Apoidea) on Lowbush Blueberry (Ericaceae: Vaccinium Angustifolium). Ann. Entomol. Soc. Am. 2002, 95, 345–351. [Google Scholar] [CrossRef]
- Jones, M.S.; Vanhanen, H.; Peltola, R.; Drummond, F. A Global Review of Arthropod-Mediated Ecosystem-Services in Vaccinium Berry Agroecosystems. Terr. Arthropod Rev. 2014, 7, 41–78. [Google Scholar] [CrossRef]
- Isaacs, R.; Kirk, A.K. Pollination Services Provided to Small and Large Highbush Blueberry Fields by Wild and Managed Bees. J. Appl. Ecol. 2010, 47, 841–849. [Google Scholar] [CrossRef]
- MAPAQ. Available online: https://www.mapaq.gouv.qc.ca/fr/Productions/Agroenvironnement/Pages/Agroenvironnement.aspx (accessed on 4 August 2018).
- Geldmann, J.; González-Varo, J.P. Conserving Honey Bees Does Not Help Wildlife. Science 2018, 359, 392–393. [Google Scholar] [CrossRef]
- Benjamin, F.E.; Reilly, J.R.; Winfree, R. Pollinator Body Size Mediates the Scale at Which Land Use Drives Crop Pollination Services. J. Appl. Ecol. 2014, 51, 440–449. [Google Scholar] [CrossRef]
- Bushmann, S.L.; Drummond, F.A. Abundance and Diversity of Wild Bees (Hymenoptera: Apoidea) Found in Lowbush Blueberry Growing Regions of Downeast Maine. Environ. Entomol. 2015, 44, 975–989. [Google Scholar] [CrossRef]
- Packer, L. The Bee Genera of Eastern Canada. Can. J. Arthropod Identif. 2007, 3, 1–32. [Google Scholar] [CrossRef]
- Michener, C.D. The Bees of the World; Johns Hopkins University Press: Baltimore, MD, USA, 2000; Volume 85, ISBN 978-0-8018-6133-8. [Google Scholar]
- Williams, P.H.; Thorp, R.W.; Richardson, L.L.; Colla, S.R. Bumble Bees of North America: An Identification Guide; Princeton University Press: Princeton, NJ, USA, 2014; ISBN 978-0-691-15222-6. [Google Scholar]
- La Financiére Agricole Du Quèbec Base de Données Des Parcelles et Productions Agricoles Déclarées. Available online: https://www.fadq.qc.ca/documents/donnees/base-de-donnees-des-parcelles-et-productions-agricoles-declarees/ (accessed on 20 March 2021).
- Benjamin, K.; Domon, G.; Bouchard, A. Vegetation Composition and Succession of Abandoned Farmland: Effects of Ecological, Historical and Spatial Factors. Landsc. Ecol. 2005, 20, 627–647. [Google Scholar] [CrossRef]
- Li, S.; Dragicevic, S.; Veenendaal, B. Advances in Web-Based GIS, Mapping Services and Applications; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Kremen, C.; Williams, N.S.G.N.M.; Bugg, R.L.; Fay, J.P.; Thorp, R.W.; de Brito, V.L.G.; Rech, A.R.; Ollerton, J.; Sazima, M.; Boreux, V.; et al. The Area Requirements of an Ecosystem Service: Crop Pollination by Native Bee Communities in California. Agric. Ecosyst. Environ. 2015, 7, 1109–1119. [Google Scholar] [CrossRef]
- Seavy, N.; Quader, S.; Alexander, J.; Ralph, C. Generalized Linear Models and Point Count Data: Statistical Considerations for the Design and Analysis of Monitoring Studies. Bird Conserv. Implement. Integr. Am. 2005, 744, 753. [Google Scholar]
- Kleiber, C.; Zeileis, A. Applied Econometrics with R; Springer: New York, NY, USA, 2008. [Google Scholar]
- Zeileis, A.; Kleiber, C.; Jackman, S. Regression Models for Count Data in R. J. Stat. Softw. 2008, 27, 1–25. [Google Scholar] [CrossRef]
- Lever, J.; Krzywinski, M.; Altman, N. Model Selection and Overfitting. Nat. Methods 2016, 703–704. [Google Scholar] [CrossRef]
- Brewer, M.J.; Butler, A.; Cooksley, S.L. The Relative Performance of AIC, AICC and BIC in the Presence of Unobserved Heterogeneity. Methods Ecol. Evol. 2016, 7, 679–692. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2002; ISBN 978-0-387-22456-5. [Google Scholar]
- Kreft, H.; Jetz, W.; Mutke, J.; Kier, G.; Barthlott, W. Global Diversity of Island Floras from a Macroecological Perspective. Ecol. Lett. 2007, 11, 116–127. [Google Scholar] [CrossRef]
- Nagelkerke, N.J.D. A Note on a General Definition of the Coefficient of Determination. Biometrika 1991, 78, 691–692. [Google Scholar] [CrossRef]
- R Core Team. The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 24 September 2020).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S-Plus; Springer: New York, NY, USA, 1999; ISBN 978-1-4757-3123-1. [Google Scholar]
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. Available online: https://cran.r-project.org/web/packages/DHARMa/vignettes/DHARMa.html (accessed on 20 March 2021).
- MacKenzie, K.E.; Winston, M.L. Diversity and Abundance of Native Bee Pollinators on Berry Crops and Natural Vegetation in the Lower Fraser Valley, British Columbia. Can. Entomol. 1984, 116, 965–974. [Google Scholar] [CrossRef]
- Scott-Dupree, C.D.; Winston, M.L. Wild Bee Pollinator Diversity and Abundance in the Orchard and Uncultivated Habitats in the Okanagan Valley, British Columbia. Can. Entomol. 1987, 119, 735–745. [Google Scholar] [CrossRef]
- Ricketts, T.H.; Regetz, J.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Bogdanski, A.; Gemmill-Herren, B.; Greenleaf, S.S.; Klein, A.M.; Mayfield, M.M.; et al. Landscape Effects on Crop Pollination Services: Are There General Patterns? Ecol. Lett. 2008, 11, 499–515. [Google Scholar] [CrossRef]
- Watson, J.C.; Wolf, A.T.; Ascher, J.S. Forested Landscapes Promote Richness and Abundance of Native Bees (Hymenoptera: Apoidea: Anthophila) in Wisconsin Apple Orchards. Environ. Entomol. 2011, 40, 621–632. [Google Scholar] [CrossRef]
- Martins, K.T.; Albert, C.H.; Lechowicz, M.J.; Gonzalez, A. Complementary Crops and Landscape Features Sustain Wild Bee Communities. Ecol. Appl. 2018, 28, 1093–1105. [Google Scholar] [CrossRef]
- Brown, M.J.F.; Paxton, R.J. The Conservation of Bees: A Global Perspective. Apidologie 2009, 40, 410–416. [Google Scholar] [CrossRef]
- Gathmann, A.; Tscharntke, T. Foraging Ranges of Solitary Bees. J. Anim. Ecol. 2002, 71, 757–764. [Google Scholar] [CrossRef]
- Osborne, J.L.; Martin, A.P.; Carreck, N.L.; Swain, J.L.; Knight, M.E.; Goulson, D.; Hale, R.J.; Sanderson, R.A. Bumblebee Flight Distances in Relation to the Forage Landscape. J. Anim. Ecol. 2008, 77, 406–415. [Google Scholar] [CrossRef]
- Benjamin, F.E.; Winfree, R. Lack of Pollinators Limits Fruit Production in Commercial Blueberry (Vaccinium corymbosum). Environ. Entomol. 2014, 43, 1574–1583. [Google Scholar] [CrossRef]
- Buchmann, S.L. Buzz Pollination in Angiosperms. In Handbook of Experimental Pollination Biology; Van Nostrand Reinhold Company: New York, NY, USA, 1983; pp. 73–113. ISBN 0-442-24676-5. [Google Scholar]
- Dogterom, M. 1946-Pollination by Four Species of Bees on Highbush Blueberry; National Library of Canada = Bibliothèque nationale du Canada: Ottawa, ON, Canada, 2001. [Google Scholar]
- Jacobson, M.M.; Tucker, E.M.; Mathiasson, M.E.; Rehan, S.M. Decline of Bumble Bees in Northeastern North America, with Special Focus on Bombus Terricola. Biol. Conserv. 2018, 217, 437–445. [Google Scholar] [CrossRef]
- Geib, J.C.; Strange, J.P.; Galen, A. Bumble Bee Nest Abundance, Foraging Distance, and Host-Plant Reproduction: Implications for Management and Conservation. Ecol. Appl. 2015, 25, 768–778. [Google Scholar] [CrossRef]
- Brown, J.C.; Albrecht, C. The Effect of Tropical Deforestation on Stingless Bees of the Genus Melipona (Insecta: Hymenoptera: Apidae: Meliponini) in Central Rondonia, Brazil. J. Biogeogr. 2001, 28, 623–634. [Google Scholar] [CrossRef]
- Walther-Hellwig, K.; Frankl, R. Foraging Habitats and Foraging Distances of Bumblebees, Bombus Spp. (Hym., Apidae), in an Agricultural Landscape. J. Appl. Entomol. 2000, 124, 299–306. [Google Scholar] [CrossRef]
- Williams, N.M.; Ward, K.L.; Pope, N.; Isaacs, R.; Wilson, J.; May, E.A.; Ellis, J.; Daniels, J.; Pence, A.; Ullmann, K.; et al. Native Wildflower Plantings Support Wild Bee Abundance and Diversity in Agricultural Landscapes across the United States. Ecol. Appl. 2015, 25, 2119–2131. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pollinator | Flower Visits | Flower Visits (%) |
|---|---|---|
| * Apis melifera | 293 | - |
| Andrena | 152 | 23.07 |
| Bombus bimaculatus | 4 | 0.61 |
| Bombus impatiens | 374 | 56.75 |
| Bombus perplexus | 1 | 0.15 |
| Bombus ternarius | 57 | 8.65 |
| Bombus terricola | 1 | 0.15 |
| Halictid green | 12 | 1.82 |
| Metallic black bee | 8 | 1.21 |
| Small black bee | 50 | 7.59 |
| Pollinator | Trapped Richness | Trapped Abundance | Relative Abundance (%) |
|---|---|---|---|
| * Apis melifera | 1 | 5 | - |
| Andrena | 8 | 14 | 20.29 |
| Augochlorella | 1 | 4 | 5.8 |
| Ceratina | 2 | 2 | 2.9 |
| Colletes | 1 | 2 | 2.9 |
| Lasioglossum | 11 | 32 | 46.38 |
| Nomada | 2 | 4 | 5.8 |
| Osmia | 5 | 6 | 8.7 |
| Sphecodes | 1 | 5 | 7.25 |
| Flower Visits | Forest | Covariate | |||
|---|---|---|---|---|---|
| Radius (m) | Model | IRR | 95% CI | IRR | 95% CI |
| 2000 | |||||
| Forest | 1.018 | 1.004–1.032 | |||
| Forest + Abandoned Fields | 1.017 | 1.004–1.032 | 1.002 | 0.984–1.022 | |
| Forest + Temperature | 1.016 | 0.999–1.034 | 0.979 | 0.849–1.130 | |
| Forest + Shrub Density | 1.018 | 1.004–1.032 | 1.000 | 1.000–1.000 | |
| 1000 | |||||
| Forest | 1.016 | 1.004–1.030 | |||
| Forest + Abandoned Fields | 1.016 | 1.004–1.030 | 1.000 | 0.983–1.018 | |
| Forest + Temperature | 1.015 | 1.000–1.031 | 0.977 | 0.853–1.120 | |
| Forest + Shrub Density | 1.016 | 1.004–1.030 | 1.000 | 1.000–1.000 | |
| 500 | |||||
| Forest | 1.010 | 0.995–1.027 | |||
| Forest + Abandoned Fields | 1.010 | 0.995–1.026 | 1.006 | 0.991–1.021 | |
| Forest + Temperature | 1.005 | 0.986–1.024 | 0.923 | 0.794–1.069 | |
| Forest + Shrub Density | 1.010 | 0.995–1.027 | 1.000 | 1.000–1.000 | |
| Visiting Richness | |||||
| 2000 | |||||
| Forest | 1.007 | 0.993–1.020 | |||
| Forest + Abandoned Fields | 1.006 | 0.993–1.019 | 1.003 | 0.986–1.019 | |
| Forest + Temperature | 1.009 | 0.993–1.025 | 1.035 | 0.908–1.180 | |
| Forest + Shrub Density | 1.006 | 0.993–1.020 | 1.000 | 1.000–1.000 | |
| 1000 | |||||
| Forest | 1.004 | 0.992–1.016 | |||
| Forest + Abandoned Fields | 1.004 | 0.990–1.016 | 1.003 | 0.988–1.018 | |
| Forest + Temperature | 1.005 | 0.991–1.020 | 1.019 | 0.896–1.160 | |
| Forest + Shrub Density | 1.004 | 0.991–1.016 | 1.000 | 1.000–1.000 | |
| 500 | |||||
| Forest | 0.997 | 0.983–1.010 | |||
| Forest + Abandoned Fields | 0.997 | 0.983–1.010 | 1.003 | 0.991–1.013 | |
| Forest + Temperature | 0.995 | 0.980–1.010 | 0.973 | 0.862–1.099 | |
| Forest + Shrub Density | 0.997 | 0.983–1.010 | 1.000 | 1.000–1.000 | |
| Trapped Abundance | Forest | Covariate | |||
|---|---|---|---|---|---|
| Radius (m) | Model | IRR | 95% CI | IRR | 95% CI |
| 2000 | |||||
| Forest | 1.040 | 1.009–1.075 | |||
| Forest + Abandoned Fields | 1.037 | 1.008–1.070 | 1.028 | 0.986–1.078 | |
| Forest + Temperature | 1.041 | 1.006–1.082 | 1.011 | 0.760–1.340 | |
| Forest + Shrub Density | 1.042 | 1.010–1.077 | 1.000 | 1.000–1.000 | |
| 1000 | |||||
| Forest | 1.035 | 1.001–1.072 | |||
| Forest + Abandoned Fields | 1.021 | 0.991–1.054 | 1.044 | 1.002–1.091 | |
| Forest + Temperature | 1.032 | 0.997–1.072 | 0.950 | 0.717–1.250 | |
| Forest + Shrub Density | 1.036 | 1.003–1.074 | 1.000 | 1.000–1.000 | |
| 500 | |||||
| Forest | 1.006 | 0.966–1.050 | |||
| Forest + Abandoned Fields | 0.999 | 0.966–1.033 | 1.047 | 1.013–1.086 | |
| Forest + Temperature | 1.003 | 0.960–1.048 | 0.871 | 0.635–1.185 | |
| Forest + Shrub Density | 1.007 | 0.967–1.052 | 1.000 | 1.000–1.000 | |
| Trapped Richness | |||||
| 2000 | |||||
| Forest | 1.028 | 0.999–1.061 | |||
| Forest + Abandoned Fields | 1.026 | 0.999–1.055 | 1.029 | 0.992–1.072 | |
| Forest + Temperature | 1.028 | 0.995–1.066 | 0.994 | 0.756–1.303 | |
| Forest + Shrub Density | 1.029 | 1.000–1.062 | 1.000 | 1.000–1.000 | |
| 1000 | |||||
| Forest | 1.024 | 0.994–1.057 | |||
| Forest + Abandoned Fields | 1.012 | 0.985–1.041 | 1.040 | 1.004–1.081 | |
| Forest + Temperature | 1.022 | 0.989–1.058 | 0.956 | 0.731–1.243 | |
| Forest + Shrub Density | 1.026 | 0.996–1.060 | 1.000 | 1.000–1.000 | |
| 500 | |||||
| Forest | 1.001 | 0.966–1.038 | |||
| Forest + Abandoned Fields | 0.997 | 0.968–1.026 | 1.040 | 1.013–1.072 | |
| Forest + Temperature | 0.996 | 0.959–1.034 | 0.883 | 0.667–1.156 | |
| Forest + Shrub Density | 1.002 | 0.968–1.040 | 1.000 | 1.000–1.000 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vega, S.; Vázquez-Rivera, H.; Normandin, É.; Fournier, V.; Lessard, J.-P. Large Remaining Forest Habitat Patches Help Preserve Wild Bee Diversity in Cultivated Blueberry Bush. Diversity 2023, 15, 405. https://doi.org/10.3390/d15030405
Vega S, Vázquez-Rivera H, Normandin É, Fournier V, Lessard J-P. Large Remaining Forest Habitat Patches Help Preserve Wild Bee Diversity in Cultivated Blueberry Bush. Diversity. 2023; 15(3):405. https://doi.org/10.3390/d15030405
Chicago/Turabian StyleVega, Sergio, Héctor Vázquez-Rivera, Étienne Normandin, Valérie Fournier, and Jean-Philippe Lessard. 2023. "Large Remaining Forest Habitat Patches Help Preserve Wild Bee Diversity in Cultivated Blueberry Bush" Diversity 15, no. 3: 405. https://doi.org/10.3390/d15030405
APA StyleVega, S., Vázquez-Rivera, H., Normandin, É., Fournier, V., & Lessard, J.-P. (2023). Large Remaining Forest Habitat Patches Help Preserve Wild Bee Diversity in Cultivated Blueberry Bush. Diversity, 15(3), 405. https://doi.org/10.3390/d15030405