
Assessing the Influence of Anthropogenic Land-Use Changes on Bird Diversity and Feeding Guilds—A Case Study of Kalametiya Lagoon (Southern Sri Lanka)

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
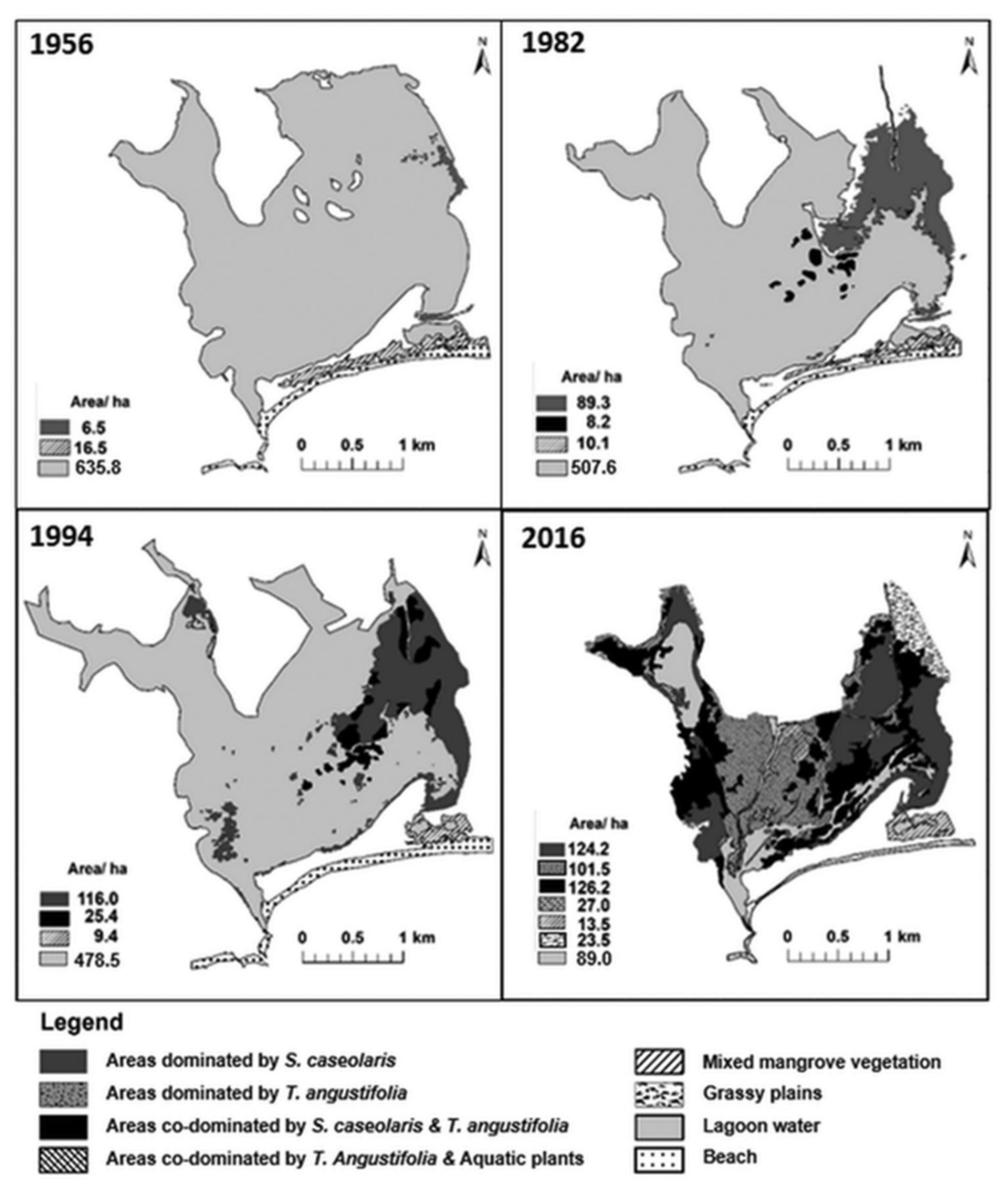
2.1. Study Area
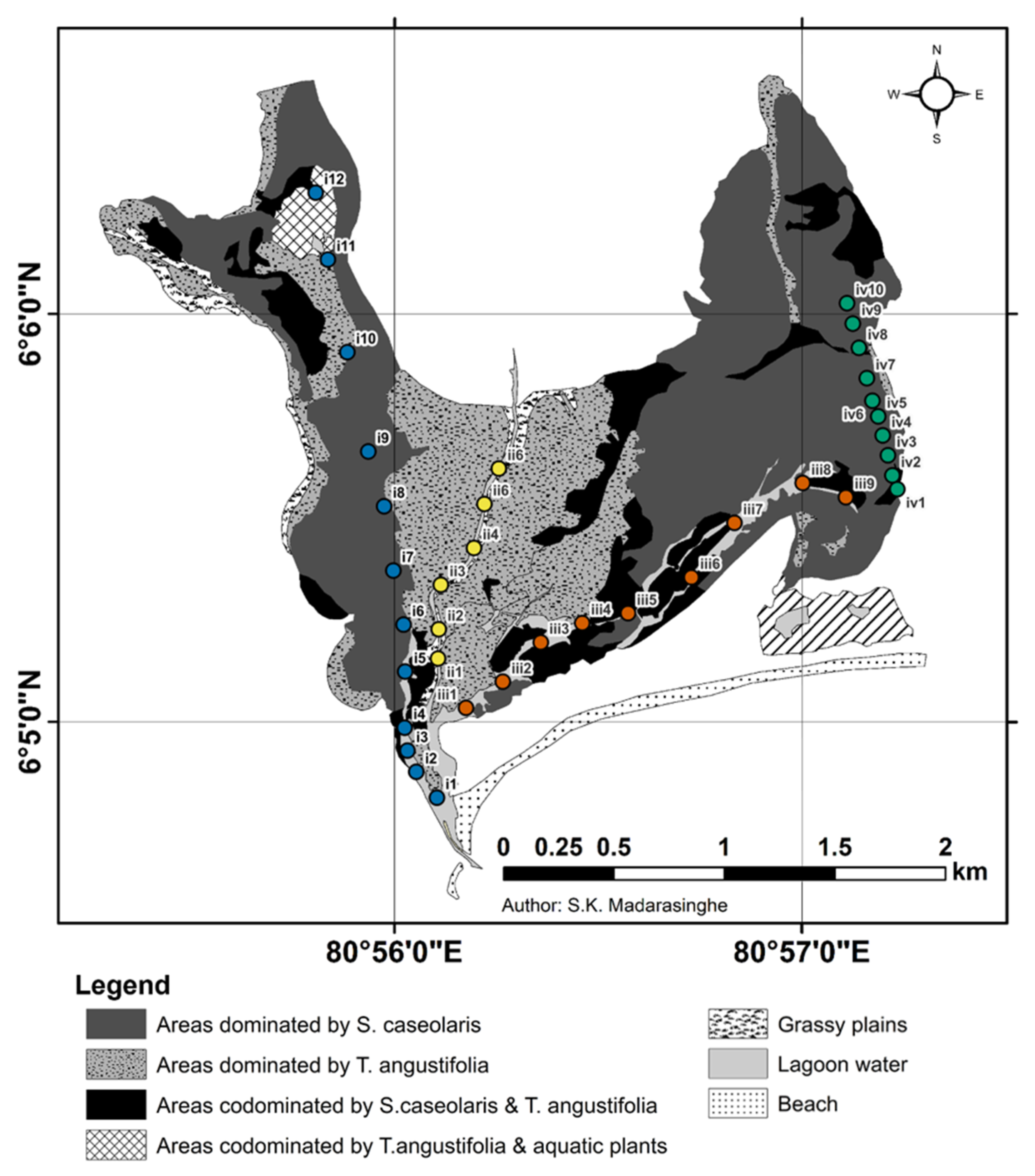
2.2. Sampling Design
2.3. Bird Surveys
2.4. Vegetation Sampling
2.5. Environmental Sampling
2.6. Habitat Affiliation
2.7. Avifauna Analysis
- (i)
- Shannon–Wiener’s Index:
- (ii)
- Simpson’s Index:
- (iii)
- Pielou’s Evenness:
- (iv)
- Taxonomic Distinctness:
- (v)
- Taxonomic Breadth:
- (vi)
- Taxonomic Distinctness Variation:
3. Results
3.1. Habitat Affiliation
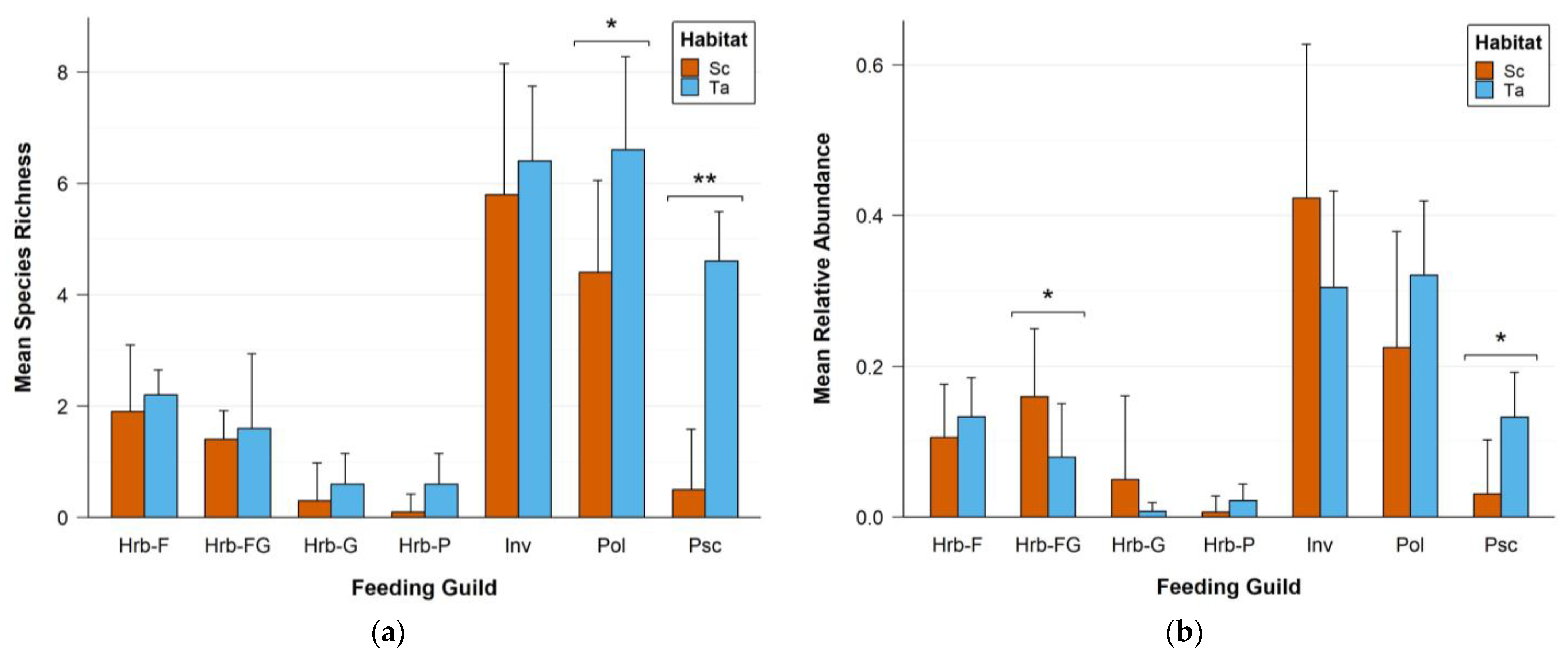
3.2. Diversity Analysis
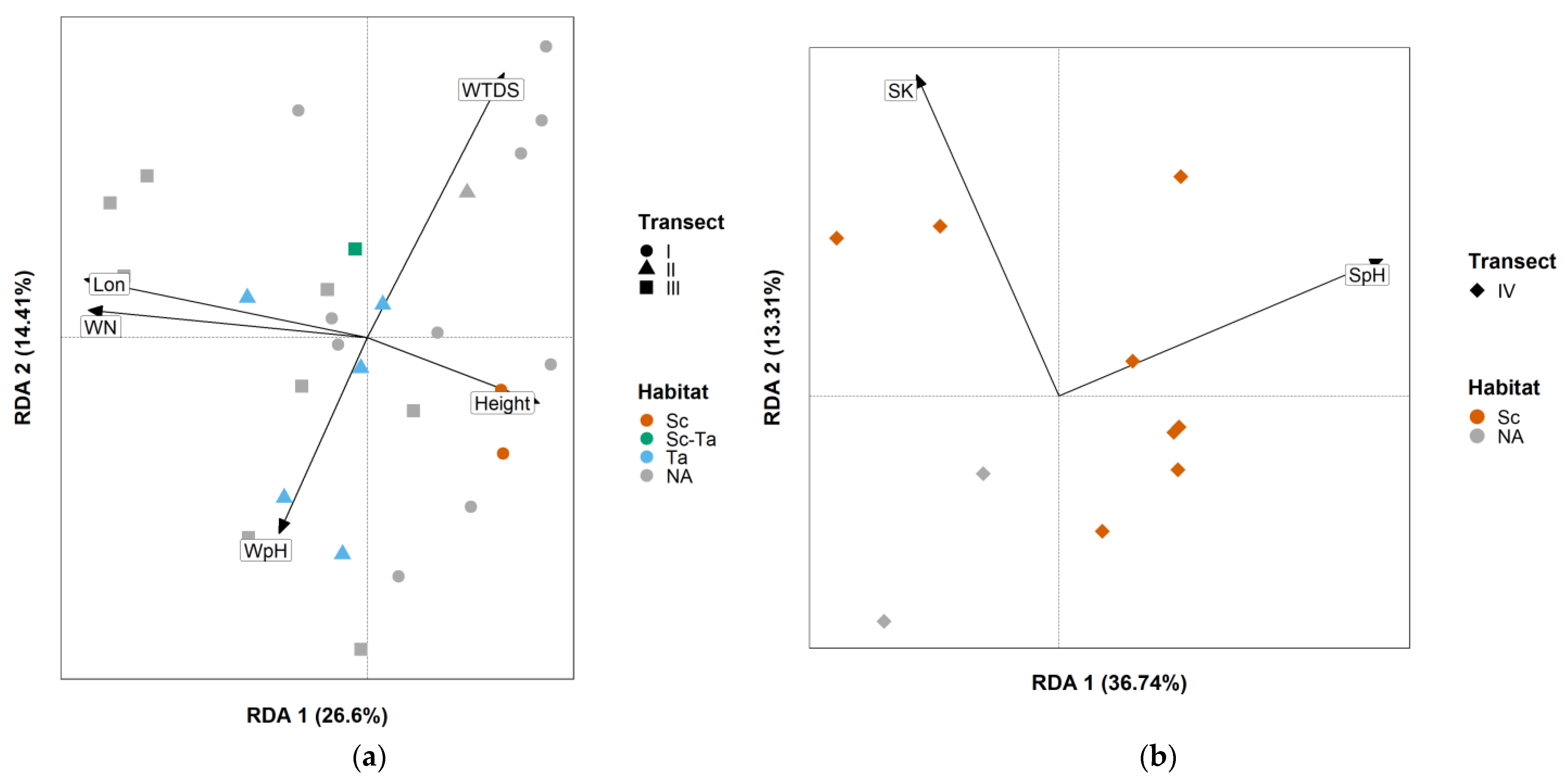
3.3. Multivariate Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Silva, E.I.L.; Katupotha, J.; Amarasinghe, O.; Manthrithilake, H.; Ariyaratna, R. Lagoons of Sri Lanka: From the Origins to the Present; International Water Management Institute: Colombo, Sri Lanka, 2013; pp. 6–8. [Google Scholar]
- Balasuriya, A. Coastal Area Management: Biodiversity and Ecological Sustainability in Sri Lankan Perspective. In Biodiversity and Climate Change Adaptation in Tropical Islands; Sivaperuman, C., Velmurugan, A., Singh, K.A., Jaisankar, I., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 701–724. [Google Scholar]
- Burke, L.; Kura, Y.; Kasem, K.; Revenga, C.; Spalding, M.; McAllister, D. Pilot Analysis of Global Ecosystems: Coastal Ecosystems; World Resources Institute: Washington, DC, USA, 2001. [Google Scholar]
- Newton, A.; Brito, A.C.; Icely, J.D.; Derolez, V.; Clara, I.; Angus, S.; Schernewski, G.; Inácio, M.; Lillebø, A.I.; Sousa, A.I.; et al. Assessing, Quantifying and Valuing the Ecosystem Services of Coastal Lagoons. J. Nat. Conserv. 2018, 44, 50–65. [Google Scholar] [CrossRef]
- Martínez, M.L.; Intralawan, A.; Vázquez, G.; Pérez-Maqueo, O.; Sutton, P.; Landgrave, R. The Coasts of Our World: Ecological, Economic and Social Importance. Ecol. Econ. 2007, 63, 254–272. [Google Scholar] [CrossRef]
- Lee, S.Y.; Primavera, J.H.; Dahdouh-Guebas, F.; McKee, K.; Bosire, J.O.; Cannicci, S.; Diele, K.; Fromard, F.; Koedam, N.; Marchand, C.; et al. Ecological Role and Services of Tropical Mangrove Ecosystems: A Reassessment. Glob. Ecol. Biogeogr. 2014, 23, 726–743. [Google Scholar] [CrossRef]
- Mukherjee, N.; Sutherland, W.J.; Dicks, L.; Hugé, J.; Koedam, N.; Dahdouh-Guebas, F. Ecosystem Service Valuations of Mangrove Ecosystems to Inform Decision Making and Future Valuation Exercises. PLoS ONE 2014, 9, e107706. [Google Scholar] [CrossRef]
- Ermgassen, P.S.E.z.; Mukherjee, N.; Worthington, T.A.; Acosta, A.; Araujo, A.R.d.R.; Beitl, C.M.; Castellanos-Galindo, G.A.; Cunha-Lignon, M.; Dahdouh-Guebas, F.; Diele, K.; et al. Fishers Who Rely on Mangroves: Modelling and Mapping the Global Intensity of Mangrove-Associated Fisheries. Estuar. Coast. Shelf Sci. 2020, 247, 106975. [Google Scholar] [CrossRef]
- Satyanarayana, B.; Stocken, T.V.d.; Rans, G.; Kodikara, K.A.S.; Ronsmans, G.; Jayatissa, L.P.; Husain, M.-L.; Koedam, N.; Dahdouh-Guebas, F. Island-Wide Coastal Vulnerability Assessment of Sri Lanka Reveals That Sand Dunes, Planted Trees and Natural Vegetation May Play a Role as Potential Barriers against Ocean Surges. Glob. Ecol. Conserv. 2017, 12, 144–157. [Google Scholar] [CrossRef]
- Gunarathne, K.; Kodikara, K.; Kokuhennadige, H.; Madarasinghe, S.; Loku Pulukkuttige, J. Diversity and Ecosystem Health of Inland Mangrove Forest in Garanduwa Lagoon, Southern Province, Sri Lanka. In Proceedings of the 15th Academic Sessions, University of Ruhuna, Matara, Sri Lanka, 7 March 2018. [Google Scholar]
- Madarasinghe, S.K.; Yapa, K.K.A.S.; Satyanarayana, B.; Udayakantha, P.M.P.; Kodikara, S.; Jayatissa, L.P. Inland Irrigation Project Causes Disappearance of Coastal Lagoon: The Trajectory of Kalametiya Lagoon, Sri Lanka from 1956 to 2016. Coast. Manag. 2020, 48, 188–209. [Google Scholar] [CrossRef]
- Madarasinghe, S.K.; Amarasinghe, Y.W.P.; Liyanage, C.H.; Gunathilake, H.M.S.A.T.; Jayasingha, J.A.I.K.; Jayasingha, M.; Priyankara, W.K.L.; Kodikara, K.A.S.; Koedam, N.; Dahdouh-Guebas, F.; et al. Retrospective Study on Changes in Dondra Lagoon (2006–2017) Resulting from Tsunami Impact and Post-Tsunami Development. J. Coast. Conserv. 2020, 24, 58. [Google Scholar] [CrossRef]
- Dahdouh-Guebas, F.; Hettiarachchi, S.; Lo Seen, D.; Batelaan, O.; Sooriyarachchi, S.; Jayatissa, L.P.; Koedam, N. Transitions in Ancient Inland Freshwater Resource Management in Sri Lanka Affect Biota and Human Populations in and around Coastal Lagoons. Curr. Biol. 2005, 15, 579–586. [Google Scholar] [CrossRef]
- Ekanayake, S.P.; Bambaradeniya, C.N.B.; Perera, W.P.N.; Perera, M.S.J.; Rodrigo, R.K.; Samarawickrema, V.A.M.P.K.; Peiris, T.N. A Biodiversity Status Profile of Lunama—Kalametiya Wetland Sanctuary; IUCN—Sri Lanka: Colombo, Sri Lanka, 2005. [Google Scholar]
- Jayatissa, L.P.; Guero, M.-C.; Hettiarachchi, S.; Koedam, N. Changes in Vegetation Cover and Socio-Economic Transitions in a Coastal Lagoon (Kalametiya, Sri Lanka), As Observed by Teledetection and Ground Truthing, Can Be Attributed to an Upstream Irrigation Scheme. Environ. Dev. Sustain. 2002, 4, 167–183. [Google Scholar] [CrossRef]
- Duke, N.; Ball, M.; Ellison, J. Factors Influencing Biodiversity and Distributional Gradients in Mangroves. Glob. Ecol. Biogeogr. Lett. 1998, 7, 27–47. [Google Scholar] [CrossRef]
- Siddig, A.A.H.; Ellison, A.M.; Ochs, A.; Villar-Leeman, C.; Lau, M.K. How Do Ecologists Select and Use Indicator Species to Monitor Ecological Change? Insights from 14 Years of Publication in Ecological Indicators. Ecol. Indic. 2016, 60, 223–230. [Google Scholar] [CrossRef]
- Birkhofer, K.; Rusch, A.; Andersson, G.K.S.; Bommarco, R.; Dänhardt, J.; Ekbom, B.; Jönsson, A.; Lindborg, R.; Olsson, O.; Rader, R.; et al. A Framework to Identify Indicator Species for Ecosystem Services in Agricultural Landscapes. Ecol. Indic. 2018, 91, 278–286. [Google Scholar] [CrossRef]
- Cairns, J.; McCormick, P.V.; Niederlehner, B.R. A Proposed Framework for Developing Indicators of Ecosystem Health. Hydrobiologia 1993, 263, 1–44. [Google Scholar] [CrossRef]
- Godefroid, S.; Koedam, N. Identifying Indicator Plant Species of Habitat Quality and Invasibility as a Guide for Peri-Urban Forest Management. Biodivers. Conserv. 2003, 12, 1699–1713. [Google Scholar] [CrossRef]
- Dahdouh-Guebas, F.; Friess, D.A.; Lovelock, C.E.; Connolly, R.M.; Feller, I.C.; Rogers, K.; Cannicci, S. Cross-Cutting Research Themes for Future Mangrove Forest Research. Nat. Plants 2022, 8, 1131–1135. [Google Scholar] [CrossRef]
- Järvinen, O.; Väisänen, R.A. Changes in Bird Populations as Criteria of Environmental Changes. Ecography 1979, 2, 75–80. [Google Scholar] [CrossRef]
- Morrison, M.L. Bird Populations as Indicators of Environmental Change. In Current Ornithology: Volume 3; Johnston, R.F., Ed.; Springer US: Boston, MA, USA, 1986; pp. 429–451. [Google Scholar] [CrossRef]
- Koskimies, P. Birds as a Tool in Environmental Monitoring. Ann. Zool. Fenn. 1989, 26, 153–166. [Google Scholar]
- Becker, P.H. Chapter 19 Biomonitoring with Birds. In Trace Metals and other Contaminants in the Environment; Markert, B.A., Breure, A.M., Zechmeister, H.G., Eds.; Elsevier: Amsterdam, The Netherlands, 2003; Volume 6, pp. 677–736. [Google Scholar] [CrossRef]
- Amat, J.A.; Green, A.J. Waterbirds as Bioindicators of Environmental Conditions. In Conservation Monitoring in Freshwater Habitats: A Practical Guide and Case Studies; Hurford, C., Schneider, M., Cowx, I., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2010; pp. 45–52. [Google Scholar] [CrossRef]
- Jayathilake, M.B.; Chandrasekara, W.U. Variation of Avifaunal Diversity in Relation to Land-Use Modifications around a Tropical Estuary, the Negombo Estuary in Sri Lanka. J. Asia-Pac. Biodivers. 2015, 8, 72–82. [Google Scholar] [CrossRef]
- Chandana, E.; Amarasinghe, N.; Samayawardhena, L. Factors Affecting the Avi-Faunal Distribution in the Three Lagoons (Malala, Embillakala and Bundala Lewaya) of Bundala National Park (A Ramsar Wetland) in Sri Lanka. Ruhuna J. Sci. 2008, 3, 34–43. [Google Scholar]
- Bellio, M.; Kingsford, R.T. Alteration of Wetland Hydrology in Coastal Lagoons: Implications for Shorebird Conservation and Wetland Restoration at a Ramsar Site in Sri Lanka. Biol. Conserv. 2013, 167, 57–68. [Google Scholar] [CrossRef]
- Fernández, J.M.; Selma, M.A.E.; Aymerich, F.R.; Sáez, M.T.P.; Fructuoso, M.F.C. Aquatic Birds as Bioindicators of Trophic Changes and Ecosystem Deterioration in the Mar Menor Lagoon (SE Spain). Hydrobiologia 2005, 550, 221–235. [Google Scholar] [CrossRef]
- Paszkowski, C.A.; Tonn, W.M. Foraging Guilds of Aquatic Birds on Productive Boreal Lakes: Environmental Relations and Concordance Patterns. Hydrobiologia 2006, 567, 19–30. [Google Scholar] [CrossRef]
- Asefa, A.; Mengesha, G.; Sori, T.; Mamo, Y. Local- and Landscape-Level Effects of Land Use Change on Bird Diversity in Abiata-Shalla Lakes National Park, Ethiopia. Afr. J. Ecol. 2019, 57, 51–58. [Google Scholar] [CrossRef]
- CEA; Euroconsult. Wetland Site Report and Conservation Management Plan: Kalametiya and Lunama Lagoons; Central Environmental Authority: Sri Jayawardenapura Kotte, Sri Lanka, 1995. [Google Scholar]
- Punyawardena, B.V.R. Climate. In The Soils of Sri Lanka; Mapa, R.B., Ed.; Springer International Publishing: Cham, Switzerland, 2020; pp. 13–22. [Google Scholar] [CrossRef]
- Bibby, C.J.; Jones, M.; Marsden, S. Expedition Field Techniques: Bird Surveys; Expedition Advisory Centre: London, UK, 1998. [Google Scholar]
- Huff, M.H.; Bettinger, K.A.; Ferguson, H.L.; Brown, M.J.; Altman, B. A Habitat-Based Point-Count Protocol for Terrestrial Birds, Emphasizing Washington and Oregon; U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 2000. [Google Scholar] [CrossRef]
- Gregory, R.D.; Gibbons, D.W.; Donald, P.F. Bird Census and Survey Techniques. In Bird Ecology and Conservation: A Handbook of Techniques; Gibbons Gregory, W.J., Newton, I., Green, R., Eds.; Oxford University Press: Oxford, UK, 2004; pp. 17–56. [Google Scholar] [CrossRef]
- Gibbons, D.W.; Gregory, R.D. Birds. In Ecological Census Techniques: A Handbook, 2nd ed.; Sutherland, W.J., Ed.; Cambridge University Press: Cambridge, UK, 2006; pp. 308–350. [Google Scholar]
- De Silva Wijeyeratne, G.; Warakagoda, D. Birds of Sri Lanka; Bloomsbury Publishing Plc: London, UK, 2016. [Google Scholar]
- Gill, F.; Donsker, D.; Rasmussen, P. IOC World Bird List (V12.1). Available online: https://www.worldbirdnames.org/new/ (accessed on 30 January 2023).
- The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org/ (accessed on 30 January 2023).
- MOE. The National Red List 2021—Conservation status of the birds of Sri Lanka (2021); Biodiversity Secretariat, Ministry of Environment: Battaramulla, Sri Lanka, 2021. [Google Scholar]
- Birds of the World. Available online: https://birdsoftheworld.org/bow/home (accessed on 30 January 2023).
- Bezzalla, A.; Houhamdi, M.; Chenchouni, H. Bird Ecological Status of Two Internationally Important Wetlands ‘Ramsar Sites and IBA’ in Algeria. Estuar. Coast. Shelf Sci. 2019, 227, 106308. [Google Scholar] [CrossRef]
- Wijeratne, G.G.N.K.; Ranawaka, D.P.D.; Gunathilaka, D.U.V.; Wijesundara, W.M.I.C.; Abeysinghe, N.K.; Thilakarathna, N.D.S.D.; Perera, A.J.D.; Dissanayake, N.P.; Andrieu, J.; Kodikara, K.A.S. Soil Organic Carbon in Mixed Mangroves and Monospecific Stands; A Case Study from Rekawa and Kalametiya Lagoons in Southern, Sri Lanka. In Proceedings of the International Symposium on Agriculture and Environment 2022, University of Ruhuna, Matara, Sri Lanka, 13 May 2022. [Google Scholar]
- Burnham, K.P.; Overton, W.S. Robust Estimation of Population Size When Capture Probabilities Vary Among Animals. Ecology 1979, 60, 927–936. [Google Scholar] [CrossRef]
- Chao, A. Estimating the Population Size for Capture-Recapture Data with Unequal Catchability. Biometrics 1987, 43, 783–791. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N.; Somerfield, P.J.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 3rd ed.; PRIMER-E: Plymouth, UK, 2014. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundations Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Oksanen, J.; Simpson, G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. Vegan: Community Ecology Package. R Package Version 2.6-2. 2022. Available online: https://cran.r-project.org/package=vegan (accessed on 25 July 2022).
- Khun, M. Caret: Classification and Regression Training. R Package Version 6.0-92. 2022. Available online: https://CRAN.R-project.org/package=caret (accessed on 30 January 2023).
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Rao, C.R. A Review of Canonical Coordinates and an Alternative to Correspondence Analysis Using Hellinger Distance. Qüestiió 1995, 19, 23–63. [Google Scholar]
- Blanchet, F.G.; Legendre, P.; Borcard, D. Forward Selection of Explanatory Variables. Ecology 2008, 89, 2623–2632. [Google Scholar] [CrossRef]
- Seneviratne, S.S.; Dayananda, S.K. History, Taxonomy and Evolutionary Status of Sri Lankan Avifauna. In The National Red List 2021—Conservation Status of the Birds of Sri Lanka (2021); Biodiversity Secretariat, Ministry of Environment: Battaramulla, Sri Lanka, 2021. [Google Scholar]
- IUCN; CEA. National Wetland Directory of Sri Lanka; IUCN—Sri Lanka: Colombo, Sri Lanka, 2006. [Google Scholar]
- Perera, N.; Perera, S.; Rodrigo, R.; Pelris, N.; Samarawickrama, P.; Ekanayake, S.; Bambaradeniya, C. An Assessment of Biodiversity in the Rekawa, Ussangoda and Kalametiya Inland Coastal Ecosystems in Southern Sri Lanka. In Proceedings of the Ninth Annual Forestry and Environment Symposium of the Department of Forestry and Environmental Science, University of Sri Jayewardenepura, Colombo, Sri Lanka, 29 December 2003. [Google Scholar] [CrossRef]
- Rosenstock, S.S.; Anderson, D.R.; Giesen, K.M.; Leukering, T.; Carter, M.F. Landbird Counting Techniques: Current Practices and an Alternative. Auk 2002, 119, 46–53. [Google Scholar] [CrossRef]
- Sólymos, P.; Matsuoka, S.M.; Stralberg, D.; Barker, N.K.S.; Bayne, E.M. Phylogeny and species traits predict bird detectability. Ecography 2018, 41, 1595–1603. [Google Scholar] [CrossRef]
- Buelow, C.; Sheaves, M. A Birds-Eye View of Biological Connectivity in Mangrove Systems. Estuar. Coast. Shelf Sci. 2015, 152, 33–43. [Google Scholar] [CrossRef]
- Michael, A.G.; Baldauf, S.L.; Mayhew, P.J.; Hill, J.K. The Response of Avian Feeding Guilds to Tropical Forest Disturbance. Conserv. Biol. 2007, 21, 133–141. [Google Scholar]
- Boulinier, T.; Nichols, J.D.; Sauer, J.R.; Hines, J.E.; Pollock, K.H. Estimating Species Richness: The Importance of Heterogeneity in Species Detectability. Ecology 1998, 79, 1018–1028. [Google Scholar] [CrossRef]
- Johnston, A.; Newson, S.E.; Risely, K.; Musgrove, A.J.; Massimino, D.; Baillie, S.R.; Pearce-Higgins, J.W. Species Traits Explain Variation in Detectability of UK Birds. Bird Study 2014, 61, 340–350. [Google Scholar] [CrossRef]
- Alwis, N.S.; Perera, P.; Dayawansa, N.P. Response of Tropical Avifauna to Visitor Recreational Disturbances: A Case Study from the Sinharaja World Heritage Forest, Sri Lanka. Avian Res. 2016, 7, 15. [Google Scholar] [CrossRef]
- Mohd-Azlan, J.; Lawes, M.J. The Effect of the Surrounding Landscape Matrix on Mangrove Bird Community Assembly in North Australia. Biol. Conserv. 2011, 144, 2134–2141. [Google Scholar] [CrossRef]
- Mohd-Azlan, J.; Noske, R.A.; Lawes, M.J. Avian Species-Assemblage Structure and Indicator Bird Species of Mangroves in the Australian Monsoon Tropics. Emu Austral Ornithol. 2012, 112, 287–297. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Latin Name | Vernacular Name | Relative Abundance | Number of Observations | Feeding Guild | Habitat |
|---|---|---|---|---|---|
| Himantopus himantopus | Black-winged Stilt | 10.29% | 252 | Inv | Sc/Ta |
| Tringa totanus | Common Redshank | 9.35% | 229 | Inv | Sc |
| Corvus splendens | House Crow | 5.67% | 139 | Pol | Sc/Sc-Ta/Ta |
| Leptocoma zeylonica | Purple-rumped Sunbird | 5.22% | 128 | Inv | Sc/Sc-Ta/Ta |
| Spilopelia chinensis | Spotted Dove | 4.57% | 112 | Hrb-FG | Sc/Ta |
| Acridotheres tristis | Common Myna | 4.41% | 108 | Pol | Sc/Ta |
| Vanellus indicus | Red-wattled Lapwing | 3.59% | 88 | Inv | Sc/Ta |
| Merops philippinus | Blue-tailed Bee-Eater | 3.31% | 81 | Inv | Sc/Sc-Ta/Ta |
| Plegadis falcinellus | Glossy Ibis | 3.18% | 78 | Pol | __ |
| Microcarbo niger | Little Cormorant | 2.90% | 71 | Psc | Sc-Ta/Ta |
| Ardea purpurea | Purple Heron | 2.90% | 71 | Pol | Sc/Sc-Ta/Ta |
| Hirundo rustica | Barn Swallow | 2.16% | 53 | Inv | Sc/Ta |
| Ducula aenea | Green Imperial Pigeon | 2.12% | 52 | Hrb-F | Sc/Sc-Ta/Ta |
| Ardea intermedia | Intermediate Egret | 2.12% | 52 | Psc | Sc/Sc-Ta/Ta |
| Egretta garzetta | Little Egret | 2.12% | 52 | Pol | Sc/Ta |
| Psittacula krameri | Rose-ringed Parakeet | 2.12% | 52 | Hrb-FG | Sc/Ta |
| Porphyrio poliocephalus | Grey-headed Swamphen | 2.08% | 51 | Hrb-P | Sc/Ta |
| Dendrocygna javanica | Lesser Whistling-Duck | 1.88% | 46 | Pol | Sc/Sc-Ta/Ta |
| Eudynamys scolopaceus | Asian Koel | 1.84% | 45 | Hrb-F | Sc/Ta |
| Ardeola Grayii | Indian Pond Heron | 1.76% | 43 | Pol | Sc/Ta |
| Sternula albifrons | Little Tern | 1.76% | 43 | Pol | Sc/Ta |
| Psilopogon zeylanicus | Brown-headed Barbet | 1.51% | 37 | Hrb-F | Sc/Sc-Ta/Ta |
| Ardea cinerea | Grey Heron | 1.47% | 36 | Psc | Ta |
| Lonchura punctulata | Scaly-breasted Munia | 1.39% | 34 | Hrb-G | Sc |
| Threskiornis melanocephalus | Black-headed Ibis | 1.31% | 32 | Pol | Sc/Sc-Ta/Ta |
| Columba livia | Rock Dove | 1.31% | 32 | Hrb-FG | Sc/Ta |
| Pelecanus philippensis | Spot-billed Pelican | 1.31% | 32 | Psc | Sc/Ta |
| Lonchura malacca | Tricolored munia | 1.10% | 27 | Hrb-G | Sc/Ta |
| Orthotomus sutorius | Common Tailorbird | 1.06% | 26 | Inv | Sc/Ta |
| Anthus rufulus | Paddyfield Pipit | 1.02% | 25 | Inv | Sc/Sc-Ta |
| Hydrophasianus chirurgus | Pheasant-tailed Jacana | 0.82% | 20 | Inv | Ta |
| Pycnonotus cafer | Red-vented Bulbul | 0.82% | 20 | Hrb-F | Sc |
| Dinopium psarodes | Red-backed Flameback | 0.78% | 19 | Inv | Sc/Ta |
| Anastomus oscitans | Asian Openbill | 0.69% | 17 | Inv | Sc/Ta |
| Anhinga melanogaster | Oriental Darter | 0.69% | 17 | Psc | Sc/Sc-Ta/Ta |
| Halcyon smyrnensis | White-throated Kingfisher | 0.69% | 17 | Pol | Sc/Sc-Ta/Ta |
| Argya affinis | Yellow-billed Babbler | 0.65% | 16 | Pol | Sc/Ta |
| Centropus sinensis | Greater Coucal | 0.61% | 15 | Pol | Sc/Sc-Ta/Ta |
| Tringa glareola | Wood Sandpiper | 0.57% | 14 | Pol | Sc |
| Prinia socialis | Ashy Prinia | 0.53% | 13 | Inv | Ta |
| Sterna hirundo | Common Tern | 0.49% | 12 | Pol | Ta |
| Mycteria leucocephala | Painted Stork | 0.49% | 12 | Psc | Sc/Ta |
| Chalcophaps indica | Common Emerald Dove | 0.45% | 11 | Hrb-FG | Ta |
| Pycnonotus luteolus | White-browed Bulbul | 0.45% | 11 | Hrb-F | Sc |
| Lalage melanoptera | Black-headed Cuckooshrike | 0.41% | 10 | Inv | Sc/Ta |
| Oriolus xanthornus | Black-hooded Oriole | 0.37% | 9 | Hrb-F | Sc/Ta |
| Alcedo atthis | Common Kingfisher | 0.37% | 9 | Psc | Sc/Ta |
| Prinia inornata | Plain Prinia | 0.37% | 9 | Inv | Sc |
| Ixobrychus sinensis | Yellow Bittern | 0.37% | 9 | Inv | Ta |
| Vanellus malabricus | Yellow-wattled Lapwing | 0.37% | 9 | Inv | Sc |
| Pavo cristatus | Indian Peafowl | 0.33% | 8 | Pol | Sc/Ta |
| Ceryle rudis | Pied Kingfisher | 0.29% | 7 | Psc | Ta |
| Motacilla cinerea | Grey Wagtail | 0.24% | 6 | Inv | Sc |
| Gallus lafayettii | Sri Lanka Junglefowl | 0.24% | 6 | Pol | __ |
| Rhipidura aureola | White-browed Fantail | 0.20% | 5 | Inv | Ta |
| Merops orientalis | Asian Green Bee-Eater | 0.16% | 4 | Inv | Sc |
| Copsychus saularis | Oriental Magpie-Robin | 0.16% | 4 | Inv | Sc |
| Gallinula chloropus | Common Moorhen | 0.12% | 3 | Pol | __ |
| Lanius cristatus | Brown Shrike | 0.08% | 2 | Inv | Sc |
| Treron bicinctus | Orange-breasted Green Pigeon | 0.08% | 2 | Hrb-F | Sc |
| Pelargopsis capensis | Stork-billed Kingfisher | 0.08% | 2 | Pol | Sc |
| Amaurornis phoenicurus | White-breasted Waterhen | 0.08% | 2 | Pol | Sc |
| Dicaeum vincens | Legge’s Flowerpecker | 0.08% | 2 | Hrb-F | Sc |
| Ardea alba | Great Egret | 0.04% | 1 | Psc | __ |
| Feeding Guild | N | RA | S | H | 1−λ | J | Δ* | Δ+ | Λ+ |
|---|---|---|---|---|---|---|---|---|---|
| Overall | 2450 | __ | 64 | 3.584 | 0.9594 | 0.8618 | 95.66 | 95.51 | 140.40 |
| Inv | 1009 | 41.18% | 21 | 2.263 | 0.8503 | 0.7434 | 91.38 | 91.07 | 214.9 |
| Pol | 707 | 28.86% | 19 | 2.483 | 0.8942 | 0.8432 | 93.05 | 96.05 | 141.6 |
| Psc | 237 | 9.67% | 9 | 1.842 | 0.8108 | 0.8383 | 88.95 | 89.58 | 499.1 |
| Hrb-F | 178 | 7.27% | 8 | 1.70 | 0.79 | 0.82 | 97.54 | 91.07 | 322.1 |
| Hrb-FG | 207 | 8.45% | 4 | 1.12 | 0.62 | 0.81 | 80.47 | 75 | 625 |
| Hrb-G | 61 | 2.49% | 2 | 0.69 | 0.49 | 0.99 | 25 | 25 | __ |
| Hrb-P | 51 | 2.08% | 1 | __ | __ | __ | __ | __ | __ |
| Categories | N | S | Chao2 (±SD) | Jacknife2 | ||
|---|---|---|---|---|---|---|
| Total | (±SD) | Total | (±SD) | |||
| Overall | 2450 | 66.22 (56.04) | 64 | 15.59 (4.01) | 65 (1.46) | 64.32 |
| Sc | 619 | 61.9 (9.86) | 51 | 14.4 * (2.17) | 69.18 (10.69) | 77.18 |
| Ta | 372 | 74.4 (13.59) | 44 | 22.6 * (2.61) | 48.55 (3.83) | 53.05 |
| Categories | (±SD) | (±SD) | (±SD) | (±SD) | (±SD) | (±SD) |
|---|---|---|---|---|---|---|
| Sc | 2.39 * (0.16) | 0.88 (0.03) | 0.9 (0.04) | 93.12 (3.41) | 94.03 (2.61) | 167.13 (65.62) |
| Ta | 2.83 * (0.31) | 0.92 (0.05) | 0.91 (0.08) | 94.72 (1.62) | 95.9 (0.71) | 152.86 (28.31) |
| Sc | Ta | ||
|---|---|---|---|
| Common Name | RA | Common Name | RA |
| Leptocoma zeylonica | 13.57% | Himantopus himantopus | 9.14% |
| Spilopelia chinensis | 12.12% | Ardea purpurea | 5.91% |
| Vanellus indicus | 7.43% | Leptocoma zeylonica | 5.65% |
| Acridotheres tristis | 5.98% | Ducula aenea | 5.38% |
| Merops philippinus | 4.52% | Corvus splendens | 4.57% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernard, T.; Kodikara, K.A.S.; Sleutel, J.; Wijeratne, G.G.N.K.; Hugé, J.; Kumara, M.P.; Weerasinghe, M.A.Y.N.; Ranakawa, D.P.D.; Thakshila, W.A.K.G.; Dahdouh-Guebas, F. Assessing the Influence of Anthropogenic Land-Use Changes on Bird Diversity and Feeding Guilds—A Case Study of Kalametiya Lagoon (Southern Sri Lanka). Diversity 2023, 15, 383. https://doi.org/10.3390/d15030383
Bernard T, Kodikara KAS, Sleutel J, Wijeratne GGNK, Hugé J, Kumara MP, Weerasinghe MAYN, Ranakawa DPD, Thakshila WAKG, Dahdouh-Guebas F. Assessing the Influence of Anthropogenic Land-Use Changes on Bird Diversity and Feeding Guilds—A Case Study of Kalametiya Lagoon (Southern Sri Lanka). Diversity. 2023; 15(3):383. https://doi.org/10.3390/d15030383
Chicago/Turabian StyleBernard, Tom, K. A. S. Kodikara, Jani Sleutel, G. G. N. K. Wijeratne, Jean Hugé, M. P. Kumara, M. A. Y. N. Weerasinghe, D. P. D. Ranakawa, W. A. K. G. Thakshila, and Farid Dahdouh-Guebas. 2023. "Assessing the Influence of Anthropogenic Land-Use Changes on Bird Diversity and Feeding Guilds—A Case Study of Kalametiya Lagoon (Southern Sri Lanka)" Diversity 15, no. 3: 383. https://doi.org/10.3390/d15030383
APA StyleBernard, T., Kodikara, K. A. S., Sleutel, J., Wijeratne, G. G. N. K., Hugé, J., Kumara, M. P., Weerasinghe, M. A. Y. N., Ranakawa, D. P. D., Thakshila, W. A. K. G., & Dahdouh-Guebas, F. (2023). Assessing the Influence of Anthropogenic Land-Use Changes on Bird Diversity and Feeding Guilds—A Case Study of Kalametiya Lagoon (Southern Sri Lanka). Diversity, 15(3), 383. https://doi.org/10.3390/d15030383