
Systematics and Phylogenetic Interrelationships of the Enigmatic Late Jurassic Shark Protospinax annectans Woodward, 1918 with Comments on the Shark–Ray Sister Group Relationship

 , , , , , , , and
, , , , , , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Material
2.2. Phylogenetic Analyses
2.3. Reviewed Fossil Material
2.4. Reviewed Extant Material
2.5. Institutional Abbreviations
3. Results
3.1. Systematic Paleontology
3.1.1. Nomenclatural Notes
3.1.2. Revised Diagnosis
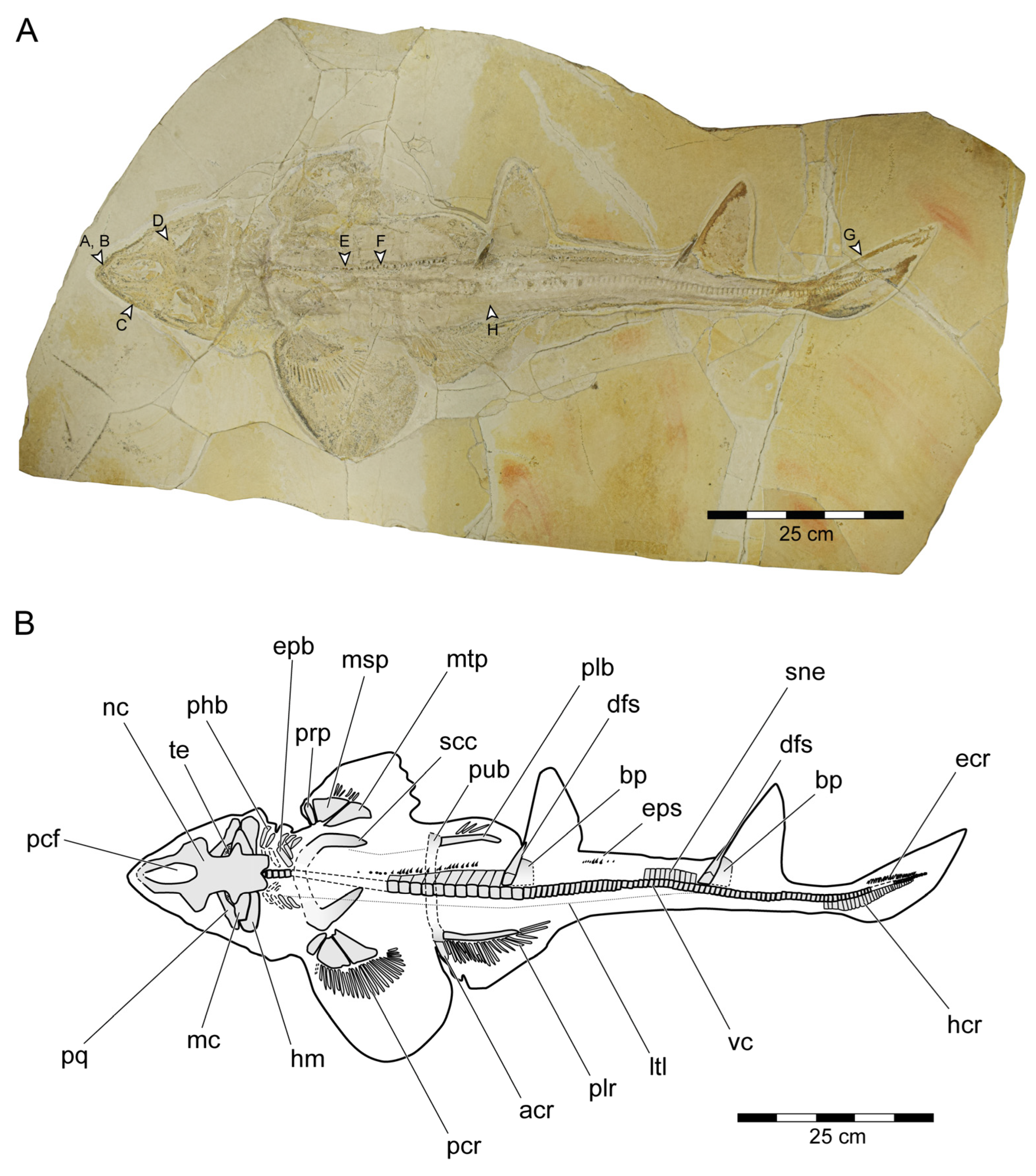
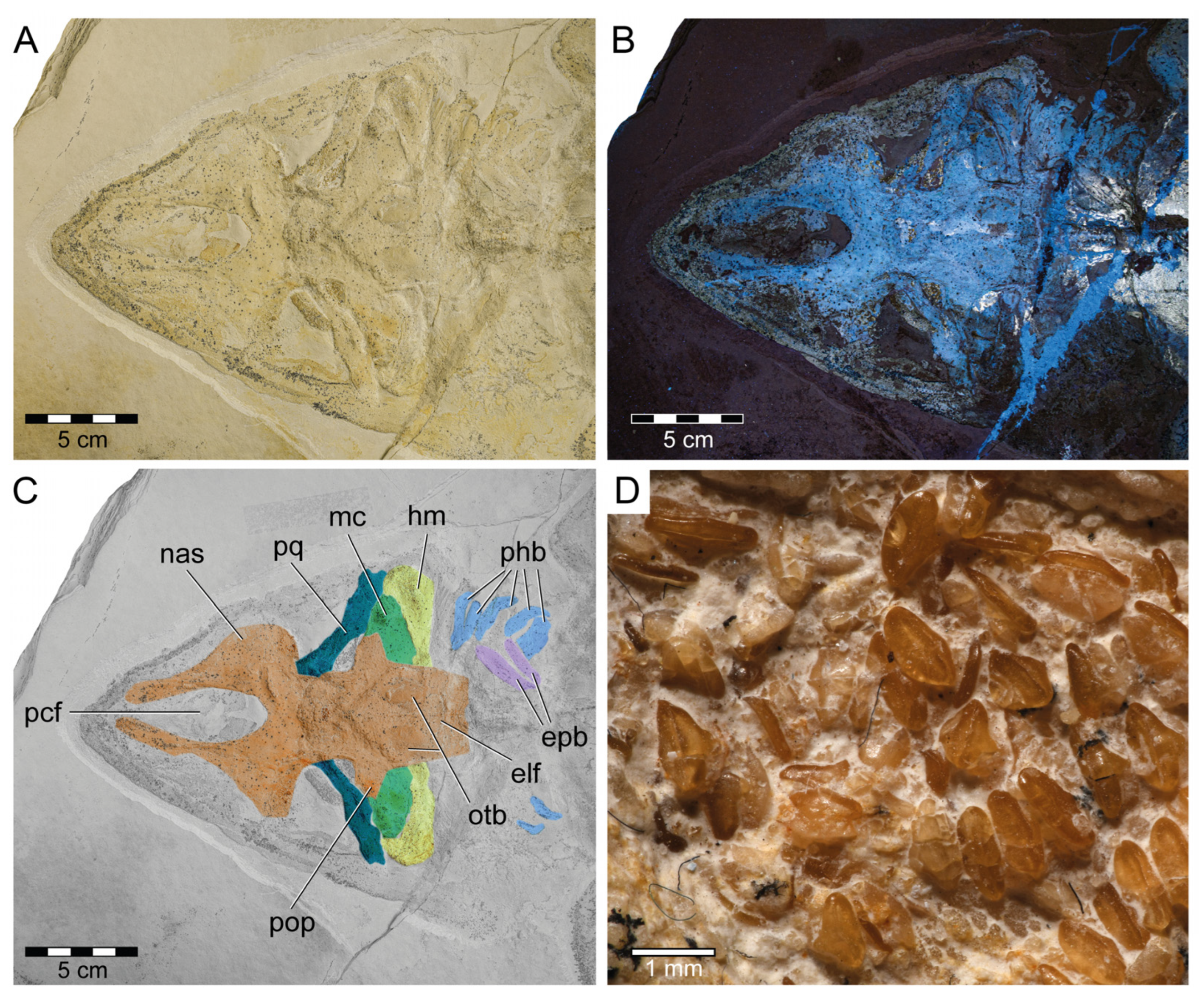
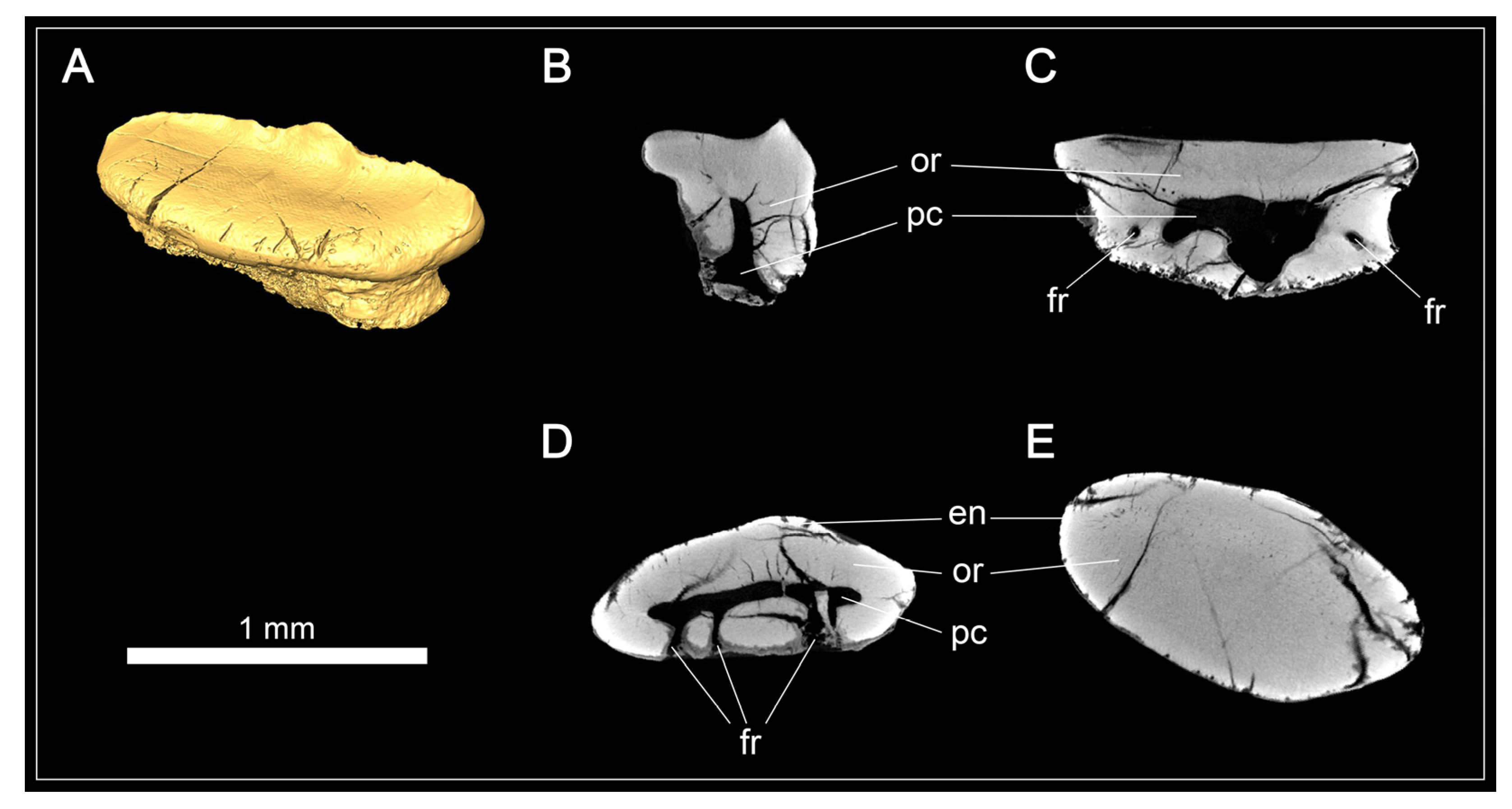
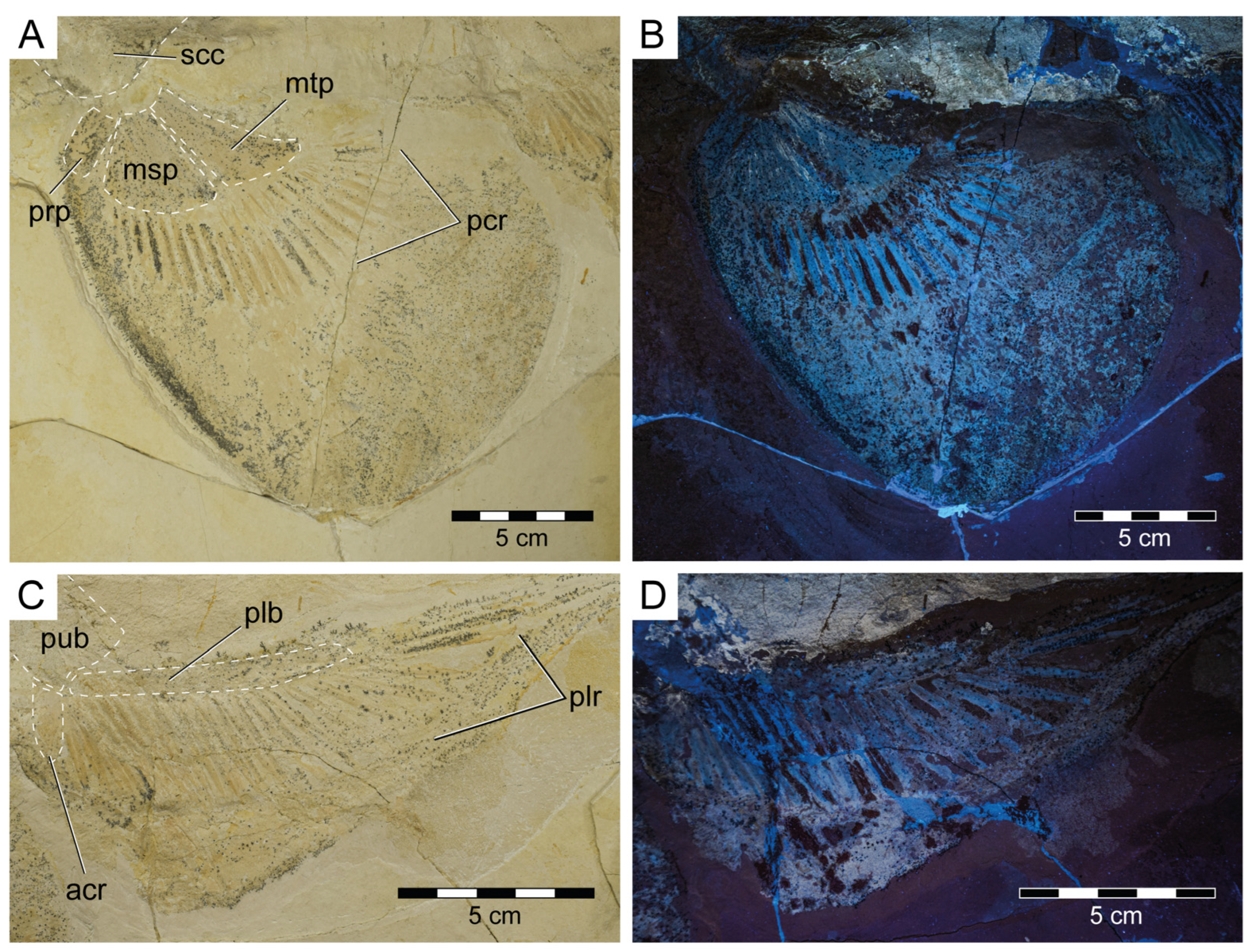
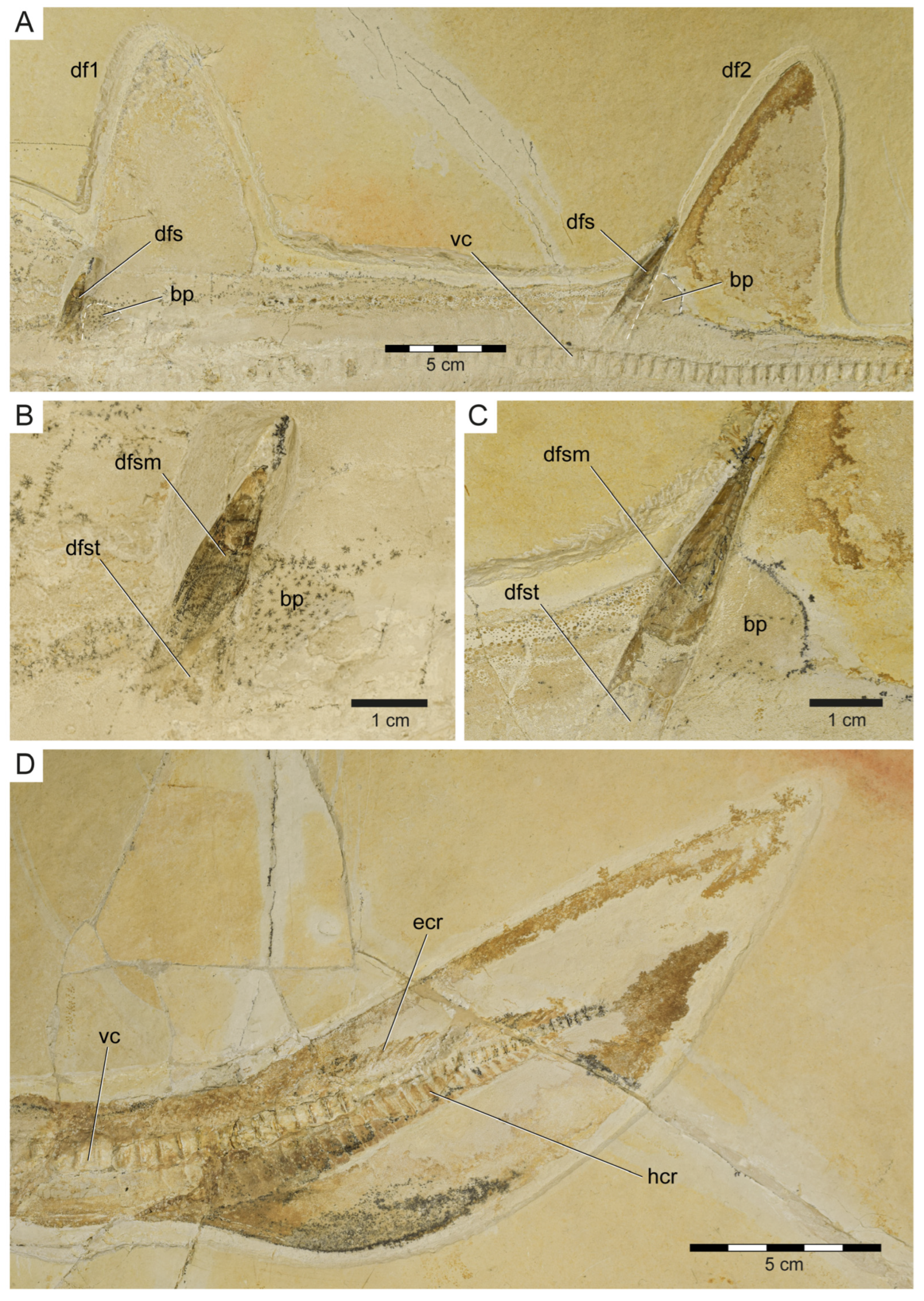
3.1.3. Description
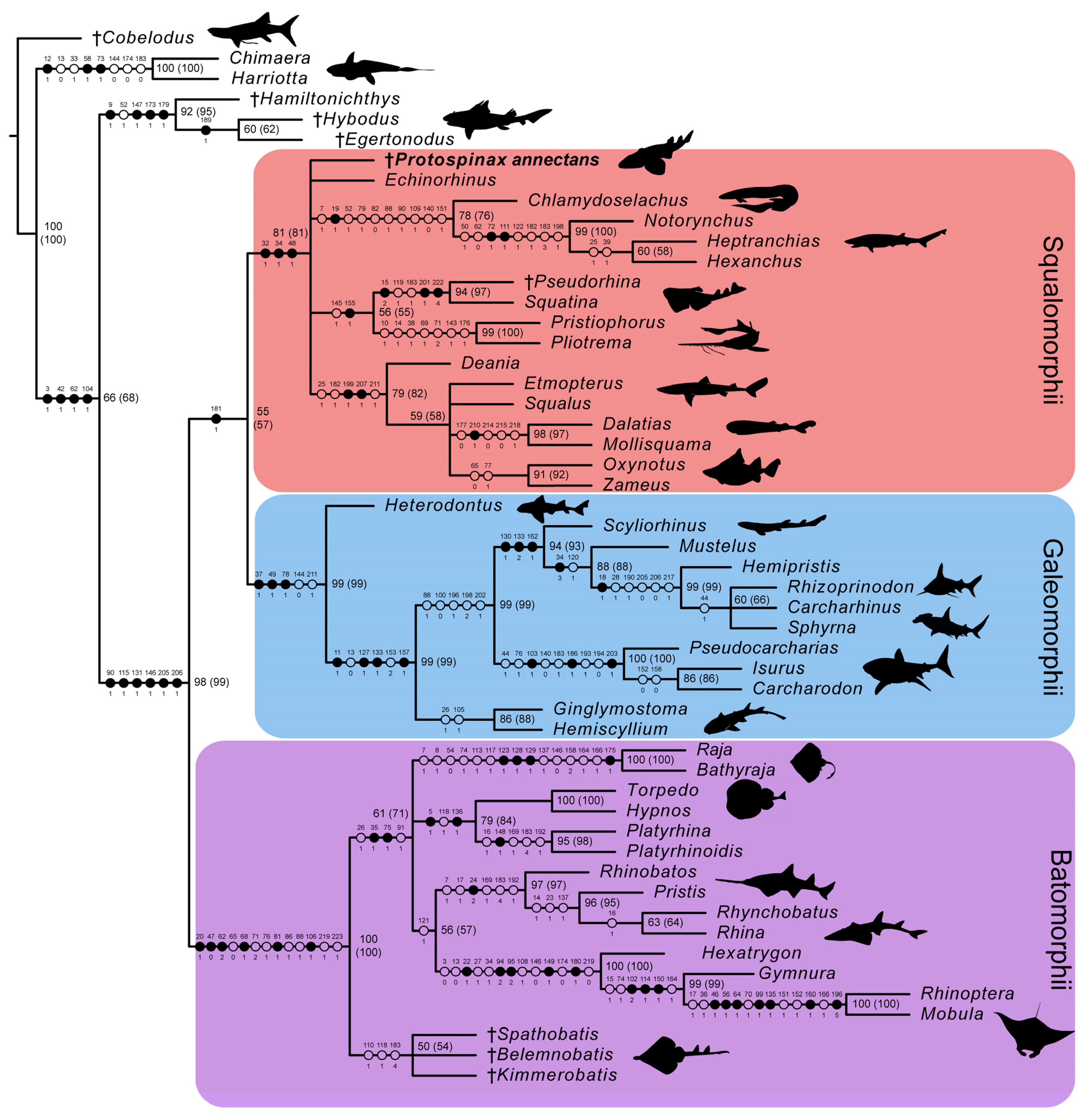
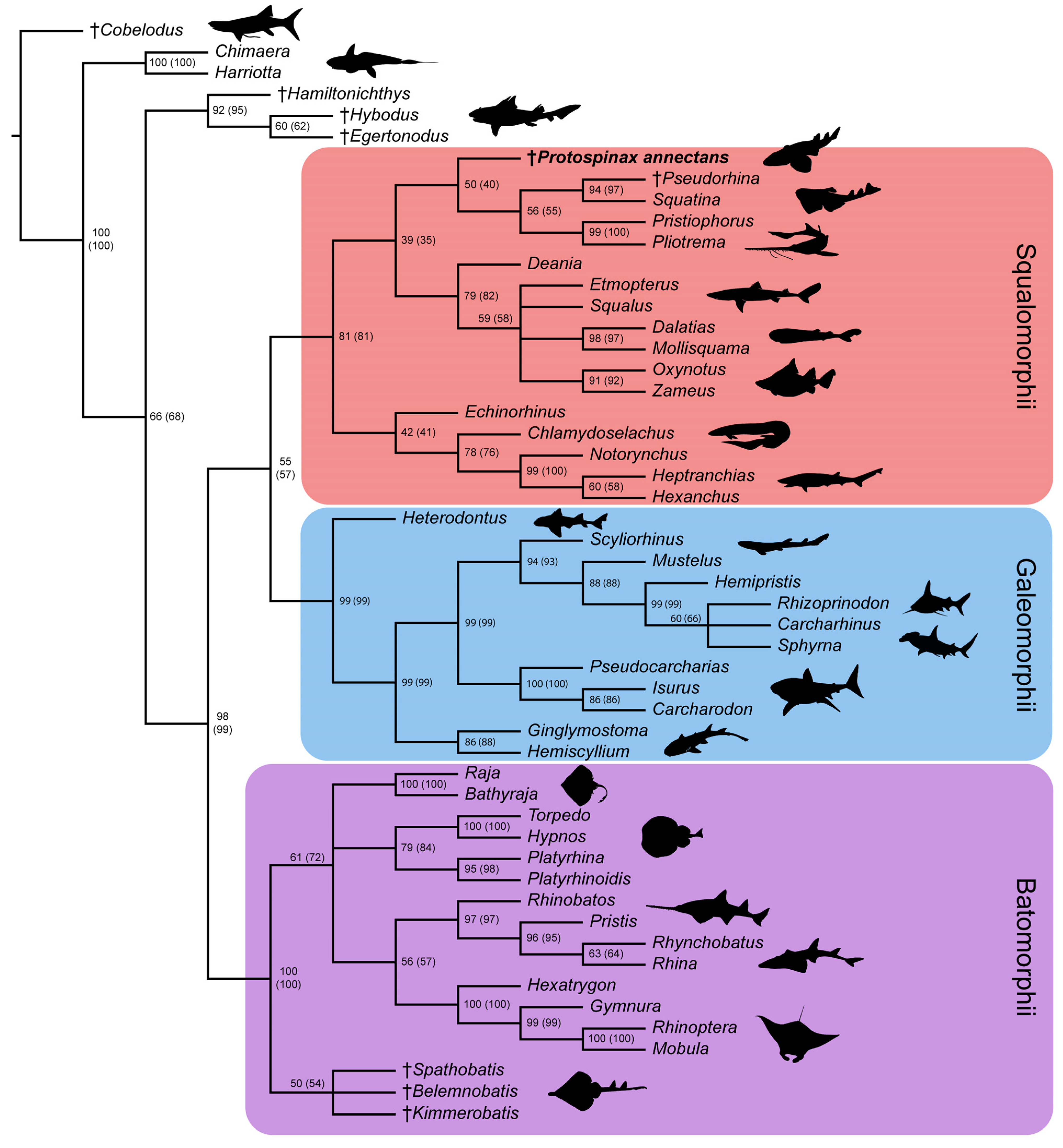
3.2. Phylogenetic Analysis
4. Discussion
4.1. Phylogenetic Implications for the Sister Group Relationship of Sharks and Rays
4.2. Phylogenetic Implications for the Superorders Batomorphii, Squalomorphii, and Galeomorphii
4.3. Phylogenetic Interrelationships of Protospinax annectans with Comments on the “Hypnosqualea” Clade
4.3.1. Revision of the Hypnosqualean Characters Proposed by Shirai [44]
4.3.2. Protospinax as a Basal Hypnosqualean as Proposed by de Carvalho and Maisey [47]
4.3.3. Protospinax as A Squaliform Shark as Proposed by Villalobos-Segura et al. [59]
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rabosky, D.L.; Slater, G.J.; Alfaro, M.E. Clade Age and Species Richness Are Decoupled Across the Eukaryotic Tree of Life. PLoS Biol. 2012, 10, e1001381. [Google Scholar] [CrossRef] [PubMed]
- Rabosky, D.L.; Chang, J.; Title, P.O.; Cowman, P.F.; Sallan, L.; Friedman, M.; Kaschner, K.; Garilao, C.; Near, T.J.; Coll, M.; et al. An Inverse Latitudinal Gradient in Speciation Rate for Marine Fishes. Nature 2018, 559, 392–395. [Google Scholar] [CrossRef] [PubMed]
- Upham, N.S.; Esselstyn, J.A.; Jetz, W. Inferring the Mammal Tree: Species-Level Sets of Phylogenies for Questions in Ecology, Evolution, and Conservation. PLoS Biol. 2019, 17, e3000494. [Google Scholar] [CrossRef] [PubMed]
- Zaman, W.; Saqib, S.; Ullah, F.; Ayaz, A.; Ye, J. COVID-19: Phylogenetic Approaches May Help in Finding Resources for Natural Cure. Phytother. Res. 2020, 34, 2783–2785. [Google Scholar] [CrossRef]
- Behl, A.; Nair, A.; Mohagaonkar, S.; Yadav, P.; Gambhir, K.; Tyagi, N.; Sharma, R.K.; Butola, B.S.; Sharma, N. Threat, Challenges, and Preparedness for Future Pandemics: A Descriptive Review of Phylogenetic Analysis Based Predictions. Infect. Genet. Evol. 2022, 98, 105217. [Google Scholar] [CrossRef]
- Edwards, E.J.; Still, C.J.; Donoghue, M.J. The Relevance of Phylogeny to Studies of Global Change. Trends Ecol. Evol. 2007, 22, 243–249. [Google Scholar] [CrossRef]
- Davis, C.C.; Willis, C.G.; Primack, R.B.; Miller-Rushing, A.J. The Importance of Phylogeny to the Study of Phenological Response to Global Climate Change. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3201–3213. [Google Scholar] [CrossRef]
- Lawing, A.M.; Polly, P.D. Pleistocene Climate, Phylogeny, and Climate Envelope Models: An Integrative Approach to Better Understand Species’ Response to Climate Change. PLoS ONE 2011, 6, e28554. [Google Scholar] [CrossRef]
- González-Orozco, C.E.; Pollock, L.J.; Thornhill, A.H.; Mishler, B.D.; Knerr, N.; Laffan, S.W.; Miller, J.T.; Rosauer, D.F.; Faith, D.P.; Nipperess, D.A.; et al. Phylogenetic Approaches Reveal Biodiversity Threats under Climate Change. Nat. Clim. Chang. 2016, 6, 1110–1114. [Google Scholar] [CrossRef]
- Isaac, N.J.B.; Turvey, S.T.; Collen, B.; Waterman, C.; Baillie, J.E.M. Mammals on the EDGE: Conservation Priorities Based on Threat and Phylogeny. PLoS ONE 2007, 2, e296. [Google Scholar] [CrossRef]
- Jetz, W.; Thomas, G.H.; Joy, J.B.; Redding, D.W.; Hartmann, K.; Mooers, A.O. Global Distribution and Conservation of Evolutionary Distinctness in Birds. Curr. Biol. 2014, 24, 919–930. [Google Scholar] [CrossRef] [PubMed]
- Faith, D.P. Phylogenetic Diversity, Functional Trait Diversity and Extinction: Avoiding Tipping Points and Worst-Case Losses. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140011. [Google Scholar] [CrossRef] [PubMed]
- Stein, R.W.; Mull, C.G.; Kuhn, T.S.; Aschliman, N.C.; Davidson, L.N.K.; Joy, J.B.; Smith, G.J.; Dulvy, N.K.; Mooers, A.O. Global Priorities for Conserving the Evolutionary History of Sharks, Rays and Chimaeras. Nat. Ecol. Evol. 2018, 2, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Springer, M.S.; Teeling, E.C.; Madsen, O.; Stanhope, M.J.; de Jong, W.W. Integrated Fossil and Molecular Data Reconstruct Bat Echolocation. Proc. Natl. Acad. Sci. USA 2001, 98, 6241–6246. [Google Scholar] [CrossRef] [PubMed]
- Parham, J.F.; Donoghue, P.C.J.; Bell, C.J.; Calway, T.D.; Head, J.J.; Holroyd, P.A.; Inoue, J.G.; Irmis, R.B.; Joyce, W.G.; Ksepka, D.T.; et al. Best Practices for Justifying Fossil Calibrations. Syst. Biol. 2012, 61, 346–359. [Google Scholar] [CrossRef] [PubMed]
- Kriwet, J.; Klug, S. Late Jurassic Selachians (Chondrichthyes, Elasmobranchii) from Southern Germany: Re-Evaluation on Taxonomy and Diversity. Zitteliana 2004, 44, 67–95. [Google Scholar] [CrossRef]
- Kriwet, J.; Klug, S. Knorpelfische (Chondrichthyes). In Solnhofen—Ein Fenster in die Jurazeit; Arratia, G., Schultze, H.-P., Tischlinger, H., Viohl, G., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2015; pp. 334–359. [Google Scholar]
- Klug, S.; Kriwet, J.; Böttcher, R.; Schweigert, G.; Dietl, G. Skeletal Anatomy of the Extinct Shark Paraorthacodus jurensis (Chondrichthyes; Palaeospinacidae), with Comments on Synechodontiform and Palaeospinacid Monophyly. Zool. J. Linn. Soc. 2009, 157, 107–134. [Google Scholar] [CrossRef]
- Thiollière, V. Troisième notice sur les gisements à poissons fossiles situés dans le Jura du département de l’Ain. Ann. Sci. Phys. Nat. Agric. Ind. 1852, 4, 353–446. [Google Scholar]
- de Saint-Seine, P. Les poissons des calcaires lithographiques de Cerin (Ain). Nouv. Arch. Muséum Hist. Nat. Lyon 1949, 2, 1–357. [Google Scholar] [CrossRef]
- Bernier, P.; Barale, G.; Bourseau, J.-P.; Buffetaut, E.; Gaillard, C.; Gall, J.-C.; Wenz, S. The Lithographic Limestones of Cerin (Southern Jura Mountains, France). A Synthetic Approach and Environmental Interpretation. Comptes Rendus Palevol 2014, 13, 383–402. [Google Scholar] [CrossRef]
- Forey, P.L.; Yi, L.; Patterson, C.; Davies, C.E. Fossil Fishes from the Cenomanian (Upper Cretaceous) of Namoura, Lebanon. J. Syst. Palaeontol. 2003, 1, 227–330. [Google Scholar] [CrossRef]
- Jambura, P.L.; Stumpf, S.; Kriwet, J. Skeletal Remains of the Oldest Known Pseudocoracid Shark Pseudocorax kindlimanni sp. nov. (Chondrichthyes, Lamniformes) from the Late Cretaceous of Lebanon. Cretac. Res. 2021, 125, 104842. [Google Scholar] [CrossRef] [PubMed]
- Pfeil, F.H. The New Family Mesiteiidae (Chondrichthyes, Orectolobiformes), Based on Mesiteia Emiliae Kramberger, 1884. In Ancient Fishes and their Living Relatives: A Tribute to John G. Maisey; Pradel, A., Denton, J.S.S., Janvier, P., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2021; pp. 101–182. [Google Scholar]
- Volta, G.S. Ittiolitologia Veronese del Museo Bozziano ora Annesso a Quello del Conte Giovambattista Gazola e di Altri Gabinetti di Fossili Veronesi: Con la Versione Latina; Dalla Stamperia Giuliari: Verona, Italy, 1796. [Google Scholar]
- Marramà, G.; Carnevale, G.; Engelbrecht, A.; Claeson, K.M.; Zorzin, R.; Fornasiero, M.; Kriwet, J. A Synoptic Review of the Eocene (Ypresian) Cartilaginous Fishes (Chondrichthyes: Holocephali, Elasmobranchii) of the Bolca Konservat-Lagerstätte, Italy. PalZ 2018, 92, 283–313. [Google Scholar] [CrossRef] [PubMed]
- Marramà, G.; Carnevale, G.; Kriwet, J. Diversity, Palaeoecology and Palaeoenvironmental Significance of the Eocene Chondrichthyan Assemblages of the Bolca Lagerstätte, Italy. Lethaia 2021, 54, 736–751. [Google Scholar] [CrossRef]
- de Carvalho, M.R.; Maisey, J.G.; Grande, L. Freshwater Stingrays of the Green River Formation of Wyoming (Early Eocene), with the Description of a New Genus and Species and an Analysis of Its Phylogenetic Relationships (Chondrichthyes: Myliobatiformes). Bull. Am. Mus. Nat. Hist. 2004, 284, 1–136. [Google Scholar] [CrossRef]
- Hovestadt, D.; Hovestadt-Euler, M.; Micklich, N. A Review of the Chondrichthyan Fauna of Grube Unterfeld (Frauenweiler) Clay Pit. Kaupia 2010, 17, 57–71. [Google Scholar]
- Hovestadt, D.C. A Partial Skeleton of Carcharias cuspidatus (Agassiz, 1843) (Chondrichthyes, Carchariidae) Including Embryos from the Oligocene of Germany. Cainozoic Res. 2022, 22, 25–36. [Google Scholar]
- Pharisat, A. La Paeoichthyofaune du Rupelien marin de Froidefontaine (Territoire de Belfort): Taxinomie et populations, genese du gisement, implications paleobiogeographiques. Ann. Sci. Univ. Fr. Comté Besançon Géologie 1991, 4, 13–97. [Google Scholar]
- Pharisat, A. Marine Fish Biodiversity in the Oligocene of Froidefontaine (Belfort Territory, France). Ital. J. Zool. 1998, 65, 189–191. [Google Scholar] [CrossRef]
- Pharisat, A.; Micklich, N. Oligocene Fishes in the Western Paratethys of the Rhine Valley Rift System. Ital. J. Zool. 1998, 65, 163–168. [Google Scholar] [CrossRef]
- Ehret, D.J.; Hubbell, G.; Macfadden, B.J. Exceptional Preservation of the White Shark Carcharodon (Lamniformes, Lamnidae) from the Early Pliocene of Peru. J. Vertebr. Paleontol. 2009, 29, 1–13. [Google Scholar] [CrossRef]
- Ehret, D.J.; Macfadden, B.J.; Jones, D.S.; Devries, T.J.; Foster, D.A.; Salas-Gismondi, R. Origin of the White Shark Carcharodon (Lamniformes: Lamnidae) Based on Recalibration of the Upper Neogene Pisco Formation of Peru. Palaeontology 2012, 55, 1139–1153. [Google Scholar] [CrossRef]
- Collareta, A.; Landini, W.; Chacaltana, C.; Bianucci, G. A Well Preserved Skeleton of the Fossil Shark Cosmopolitodus hastalis from the Late Miocene of Peru, Featuring Fish Reamins as Fossilized Stomach Contents. Riv. Ital. Paleontol. E Stratigr. 2017, 123, 11–22. [Google Scholar] [CrossRef]
- Naylor, G.J.P.; Caira, J.N.; Jensen, K.; Rosana, K.A.M.; Straube, N.; Lakner, C. Elasmobranch Phylogeny: A Mitochondrial Estimate Based on 595 Species. In Biology of Sharks and Their Relatives; Carrier, J.C., Musick, J.A., Heithaus, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 31–56. ISBN 978-1-4398-3924-9. [Google Scholar]
- Maisey, J.G.; Naylor, G.J.P.; Ward, D.J. Mesozoic Elasmobranchs, Neoselachian Phylogeny and the Rise of Modern Elasmobranch Diversity. In Mesozoic Fishes 3—Systematics, Palaeoenvironments and Biodiversity; Arratia, G., Tintori, A., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2004; pp. 17–56. ISBN 3-89937-053-8. [Google Scholar]
- Douady, C.J.; Dosay, M.; Shivji, M.S.; Stanhope, M.J. Molecular Phylogenetic Evidence Refuting the Hypothesis of Batoidea (Rays and Skates) as Derived Sharks. Mol. Phylogenet. Evol. 2003, 26, 215–221. [Google Scholar] [CrossRef]
- Human, B.A.; Owen, E.P.; Compagno, L.J.V.; Harley, E.H. Testing Morphologically Based Phylogenetic Theories within the Cartilaginous Fishes with Molecular Data, with Special Reference to the Catshark Family (Chondrichthyes; Scyliorhinidae) and the Interrelationships within Them. Mol. Phylogenet. Evol. 2006, 39, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Vélez-Zuazo, X.; Agnarsson, I. Shark Tales: A Molecular Species-Level Phylogeny of Sharks (Selachimorpha, Chondrichthyes). Mol. Phylogenet. Evol. 2011, 58, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Amaral, C.R.L.; Pereira, F.; Silva, D.A.; Amorim, A.; de Carvalho, E.F. The Mitogenomic Phylogeny of the Elasmobranchii (Chondrichthyes). Mitochondrial DNA Part A 2018, 29, 867–878. [Google Scholar] [CrossRef]
- Kousteni, V.; Mazzoleni, S.; Vasileiadou, K.; Rovatsos, M. Complete Mitochondrial DNA Genome of Nine Species of Sharks and Rays and Their Phylogenetic Placement among Modern Elasmobranchs. Genes 2021, 12, 324. [Google Scholar] [CrossRef]
- Shirai, S. Phylogenetic Relationships of the Angel Sharks, with Comments on Elasmobranch Phylogeny (Chondrichthyes, Squatinidae). Copeia 1992, 1992, 505–518. [Google Scholar] [CrossRef]
- Shirai, S. Squalean Phylogeny: A New Framework of “Squaloid” Sharks and Related Taxa; Hokkaido University Press: Sapporo, Japan, 1992; ISBN 978-4-8329-0276-3. [Google Scholar]
- Shirai, S. Phylogenetic Interrelationships of Neoselachians. In Interrelationships of Fishes; Stiassny, M.L.J., Parenti, L.R., Johnson, D.G., Eds.; Academic Press: London, UK; New York, NY, USA; San Diego, CA, USA, 1996; pp. 9–34. [Google Scholar]
- de Carvalho, M.R.; Maisey, J.G. Phylogenetic Relationships of the Late Jurassic Shark Protospinax Woodward 1919 (Chondrichthyes: Elasmobranchii). In Mesozoic Fishes—Systematics and Paleoecology; Arratia, G., Viohl, G., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 1996; pp. 9–46. ISBN 3-923871-90-2. [Google Scholar]
- de Carvalho, M.R. Higher-Level Elasmobranch Phylogeny, Basal Squaleans and Paraphyly. In Interrelationships of fishes; Stiassny, M.L.J., Parenti, L.R., Johnson, D.G., Eds.; Academic Press: London, UK; New York, NY, USA; San Diego, CA, USA, 1996; pp. 35–62. [Google Scholar]
- Underwood, C.J. Diversification of the Neoselachii (Chondrichthyes) during the Jurassic and Cretaceous. Paleobiology 2006, 32, 215–235. [Google Scholar] [CrossRef]
- Kriwet, J.; Kiessling, W.; Klug, S. Diversification Trajectories and Evolutionary Life-History Traits in Early Sharks and Batoids. Proc. R. Soc. B Biol. Sci. 2009, 276, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Stumpf, S.; Kriwet, J. A New Pliensbachian Elasmobranch (Vertebrata, Chondrichthyes) Assemblage from Europe, and Its Contribution to the Understanding of Late Early Jurassic Elasmobranch Diversity and Distributional Patterns. PalZ 2019, 93, 637–658. [Google Scholar] [CrossRef]
- Cappetta, H. Handbook of Paleoichthyology, Vol. 3E: Chondrichthyes—Mesozoic and Cenozoic Elasmobranchii: Teeth; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2012; p. 512. [Google Scholar]
- Woodward, A.S. On Two New Elasmobranch Fishes (Crossorhinus jurassicus, sp. nov., and Protospinax annectans, gen. et sp. nov.) from the Upper Jurassic Lithographic Stone of Bavaria. Proc. Zool. Soc. Lond. 1919, 13, 231–235. [Google Scholar] [CrossRef]
- Maisey, J.G. The Jurassic Selachian Fish Protospinax Woodward. Palaeontology 1976, 19, 733–747. [Google Scholar]
- White, E.G. Interrelationships of the Elasmobranchs with a Key to the Order Galea. Bull. Am. Mus. Nat. Hist. 1937, 74, 25–138. [Google Scholar]
- Compagno, L.J.V. Interrelationships of Living Elasmobranchs. Zool. J. Linn. Soc. 1973, 53, 15–61. [Google Scholar]
- Schaeffer, B. Comments on Elasmobranch Evolution. In Sharks, Skates and Rays; Gilbert, P.W., Mathewson, R.F., Rall, D.P., Eds.; John Hopkins Press: Baltimore, MD, USA, 1967; pp. 3–35. [Google Scholar]
- Thies, D. Jurazeitliche Neoselachier aus Deutschland und S-England. Cour. Forsch. Senckenberg 1983, 58, 1–116. [Google Scholar]
- Villalobos-Segura, E.; Marramà, G.; Carnevale, G.; Claeson, K.M.; Underwood, C.J.; Naylor, G.J.P.; Kriwet, J. The Phylogeny of Rays and Skates (Chondrichthyes: Elasmobranchii) Based on Morphological Characters Revisited. Diversity 2022, 14, 456. [Google Scholar] [CrossRef]
- Adnet, S.; Cappetta, H. A Palaeontological and Phylogenetical Analysis of Squaliform Sharks (Chondrichthyes: Squaliformes) Based on Dental Characters. Lethaia 2001, 34, 234–248. [Google Scholar] [CrossRef]
- Flammensbeck, C.K.; Pollerspöck, J.; Schedel, F.D.B.; Matzke, N.J.; Straube, N. Of Teeth and Trees: A Fossil Tip-Dating Approach to Infer Divergence Times of Extinct and Extant Squaliform Sharks. Zool. Scr. 2018, 47, 539–557. [Google Scholar] [CrossRef]
- Heinicke, M.P.; Naylor, G.J.P.; Hedges, S.B. Cartilaginous Fishes (Chondrichthyes). In the Timetree of Life; Hedges, S.B., Kumar, S., Eds.; Oxford University Press: New York, NY, USA, 2009; pp. 320–327. [Google Scholar]
- Lee, M.S.Y. Molecular Evidence and Marine Snake Origins. Biol. Lett. 2005, 1, 227–230. [Google Scholar] [CrossRef] [PubMed]
- Evers, S.W.; Benson, R.B.J. A New Phylogenetic Hypothesis of Turtles with Implications for the Timing and Number of Evolutionary Transitions to Marine Lifestyles in the Group. Palaeontology 2019, 62, 93–134. [Google Scholar] [CrossRef]
- Pugh, K.D.; Gilbert, C.C. Phylogenetic Relationships of Living and Fossil African Papionins: Combined Evidence from Morphology and Molecules. J. Hum. Evol. 2018, 123, 35–51. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Li, Y.; Feldberg, K.; Wang, Y.-D.; Yang, X.-J. Radula heinrichsii (Radulaceae, Porellales), a Leafy Liverwort from the Mid-Cretaceous of Myanmar. Palaeoworld 2022, 31, 679–687. [Google Scholar] [CrossRef]
- Schweigert, G. Ammonite Biostratigraphy as a Tool for Dating Upper Jurassic Lithographic Limestones from South Germany First Results and Open Questions. Neues Jahrb. Für Geol. Paläontol. Abh. 2007, 245, 117–125. [Google Scholar] [CrossRef]
- Schweigert, G. Biostratigraphie der Plattenkalke der südlichen Frankenalb. In Solnhofen–Ein Fenster in die Jurazeit; Arratia, G., Schultze, H.-P., Tischlinger, H., Viohl, G., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2015; pp. 63–66. [Google Scholar]
- Niebuhr, B.; Pürner, T. Plattenkalk und Frankendolomit—Lithostratigraphie der Weißjura-Gruppe der Frankenalb (außeralpiner Oberjura, Bayern). Schr. Dtsch. Ges. Für Geowiss. 2014, 83, 5–72. [Google Scholar] [CrossRef]
- Stumpf, S.; López-Romero, F.A.; Kindlimann, R.; Lacombat, F.; Pohl, B.; Kriwet, J. A Unique Hybodontiform Skeleton Provides Novel Insights into Mesozoic Chondrichthyan Life. Pap. Palaeontol. 2021, 7, 1479–1505. [Google Scholar] [CrossRef]
- Tischlinger, H.; Arratia, G. Ultraviolet Light as a Tool of Investigating Mesozoic Fishes with a Focus on the Ichthyofauna of the Solnhofen Limestone. In Mesozoic Fishes 5—Global Diversity and Evolution; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2013; pp. 549–562. [Google Scholar]
- Compagno, L.J.V. Sharks of the World: Part 1: Hexachiformes to Lamniformes; FAO Species Catalogue; FAO: Rome, Italy, 1984; Volume 4, ISBN 978-92-5-101384-7. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. 2021.
- Wilberg, E.W. What’s in an Outgroup? The Impact of Outgroup Choice on the Phylogenetic Position of Thalattosuchia (Crocodylomorpha) and the Origin of Crocodyliformes. Syst. Biol. 2015, 64, 621–637. [Google Scholar] [CrossRef]
- Grant, T. Outgroup Sampling in Phylogenetics: Severity of Test and Successive Outgroup Expansion. J. Zool. Syst. Evol. Res. 2019, 57, 748–763. [Google Scholar] [CrossRef]
- Landemaine, O.; Thies, D.; Waschkewitz, J. The Late Jurassic Shark Palaeocarcharias (Elasmobranchii, Selachimorpha)—Functional Morphology of Teeth, Dermal Cephalic Lobes and Phylogenetic Position. Palaeontogr. Abt. A 2018, 312, 103–165. [Google Scholar] [CrossRef]
- Klug, S. Monophyly, Phylogeny and Systematic Position of the †Synechodontiformes (Chondrichthyes, Neoselachii). Zool. Scr. 2010, 39, 37–49. [Google Scholar] [CrossRef]
- McEachran, J.D.; Miyake, T. Phylogenetic Interrelationships of Skates: A Working Hypothesis (Chondrichthyes, Rajoidei). In Elasmobranchs as Living Resources: Advances in the Biology, Ecology, Systematics, and the Status of the Fisheries; Pratt, H.L., Gruber, S.H., Taniuchi, T., Eds.; NOAA Technical Report NMFS; 1990; pp. 285–304. [Google Scholar]
- Nishida, K. Phylogeny of the Suborder Myliobatidoidei. Mem. Fac. Fish. Sci. Hokkaido Univ. 1990, 37, 1–108. [Google Scholar]
- Brito, P.M.; Seret, B. The New Genus Iansan (Chondrichthyes, Rhinobatoidea) from the Early Cretaceous of Brazil and Its Phylogenetic Relationships. In Mesozoic Fishes—Systematics and Paleoecology; Arratia, G., Viohl, G., Eds.; Verlag Dr. Friedrich Pfeil: München, Germany, 1996; pp. 47–62. [Google Scholar]
- McEachran, J.D.; Dunn, K.A. Phylogenetic Analysis of Skates, a Morphologically Conservative Clade of Elasmobranchs (Chondrichthyes: Rajidae). Copeia 1998, 1998, 271–290. [Google Scholar] [CrossRef]
- Brito, P.M.; Dutheil, D.B. A Preliminary Systematic Analysis of Cretaceous Guitarfishes from Lebanon. In Mesozoic Fishes 3—Systematics, Paleoenvironments and Biodiversity; Arratia, G., Tintori, A., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2004; Volume 3, pp. 101–108. [Google Scholar]
- de Carvalho, M.R. Late Cretaceous Thornback Ray from Southern Italy, with a Phylogenetic Reappraisal of the Platyrhinidae (Chondrichthyes: Batoidea). In Mesozoic Fishes 3—Systematics, Paleoenvironments and Biodiversity; Arratia, G., Tintori, A., Eds.; Verlag Dr. Friedrich Pfeil: München, Germany, 2004; pp. 75–100. [Google Scholar]
- Kriwet, J. The Systematic Position of the Cretaceous Sclerohynchid Sawfishes (Elasmobranchii, Pristiorajea). In Mesozoic Fishes 3—Systematics, Paleoenvironments and Biodiversity; Arratia, G., Tintori, A., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2004; pp. 57–73. ISBN 3-89937-053-8. [Google Scholar]
- McEachran, J.D.; Aschliman, N. Phylogeny of Batoidea. In Biology of Sharks and their Relatives; Carrier, J.C., Musick, J.A., Heithaus, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2004; pp. 79–113. [Google Scholar]
- Aschliman, N.C.; Claeson, K.M.; McEachran, J.D. Phylogeny of Batoidea. In Biology of Sharks and Their Relatives; CRC Press: Boca Raton, FL, USA, 2012; pp. 57–95. ISBN 978-0-429-10654-5. [Google Scholar]
- Brito, P.; Leal, M.; Gallo, V. A New Lower Cretaceous Guitarfish (Chondrichthyes, Batoidea) from the Santana Formation, Northeastern Brazil. Bol. Mus. Nac. Nova Ser. Geol. 2013, 76, 1–15. [Google Scholar]
- Brito, P.M.; Villalobos-Segura, E.; Alvarado-Ortega, J. A New Early Cretaceous Guitarfish (Chondrichthyes, Batoidea) from the Tlayúa Formation, Puebla, Mexico. J. South Am. Earth Sci. 2019, 90, 155–161. [Google Scholar] [CrossRef]
- Claeson, K.M.; Underwood, C.J.; Ward, D.J. Tingitanius tenuimandibulus, a New Platyrhinid Batoid from the Turonian (Cretaceous) of Morocco and the Cretaceous Radiation of the Platyrhinidae. J. Vertebr. Paleontol. 2013, 33, 1019–1036. [Google Scholar] [CrossRef]
- Johanson, Z.; Trinajstic, K.; Carr, R.; Ritchie, A. Evolution and Development of the Synarcual in Early Vertebrates. Zoomorphology 2013, 132, 95–110. [Google Scholar] [CrossRef]
- Claeson, K.M. The Impacts of Comparative Anatomy of Electric Rays (Batoidea: Torpediniformes) on Their Systematic Hypotheses. J. Morphol. 2014, 275, 597–612. [Google Scholar] [CrossRef]
- Marramà, G.; Klug, S.; de Vos, J.; Kriwet, J. Anatomy, Relationships and Palaeobiogeographic Implications of the First Neogene Holomorphic Stingray (Myliobatiformes: Dasyatidae) from the Early Miocene of Sulawesi, Indonesia, SE Asia. Zool. J. Linn. Soc. 2018, 184, 1142–1168. [Google Scholar] [CrossRef]
- Underwood, C.J.; Claeson, K.M. The Late Jurassic Ray Kimmerobatis etchesi gen. et sp. nov. and the Jurassic Radiation of the Batoidea. Proc. Geol. Assoc. 2019, 130, 345–354. [Google Scholar] [CrossRef]
- Villalobos-Segura, E.; Underwood, C.J.; Ward, D.J.; Claeson, K.M. The First Three-Dimensional Fossils of Cretaceous Sclerorhynchid Sawfish: Asflapristis cristadentis gen. et sp. nov., and Implications for the Phylogenetic Relations of the Sclerorhynchoidei (Chondrichthyes). J. Syst. Palaeontol. 2019, 17, 1847–1870. [Google Scholar] [CrossRef]
- McEachran, J.D.; Dunn, K.A.; Miyake, T. Interrelationships of the Batoid Fishes (Chondrichthyes: Batoidea). In Interrelationships of Fishes; Stiassny, M.L.J., Parenti, L.R., Johnson, G.D., Eds.; Academic Press: San Diego, CA, USA; London, UK, 1996; pp. 63–84. [Google Scholar]
- Pollerspöck, J.; Straube, N. Phylogenetic Placement and Description of an Extinct Genus and Species of Kitefin Shark Based on Tooth Fossils (Squaliformes: Dalatiidae). J. Syst. Palaeontol. 2022, 19, 1083–1096. [Google Scholar] [CrossRef]
- Goto, T. Comparative Anatomy, Phylogeny and Cladistic Classification of the Order Orectolobiformes (Chondrichthyes, Elasmobranchii). Mem. Grad. Sch. Fish. Sci. Hokkaido Univ. 2001, 48, 1–100. [Google Scholar]
- Shimada, K. Phylogeny of Lamniform Sharks (Chondrichthyes: Elasmobranchii) and the Contribution of Dental Characters to Lamniform Systematics. Paleontol. Res. 2005, 9, 55–72. [Google Scholar] [CrossRef]
- Stone, N.R.; Shimada, K. Skeletal Anatomy of the Bigeye Sand Tiger Shark, Odontaspis noronhai (Lamniformes: Odontaspididae), and Its Implications for Lamniform Phylogeny, Taxonomy, and Conservation Biology. Copeia 2019, 107, 632–652. [Google Scholar] [CrossRef]
- Coates, M.I.; Gess, R.W.; Finarelli, J.A.; Criswell, K.E.; Tietjen, K. A Symmoriiform Chondrichthyan Braincase and the Origin of Chimaeroid Fishes. Nature 2017, 541, 208–211. [Google Scholar] [CrossRef]
- Frey, L.; Coates, M.I.; Tietjen, K.; Rücklin, M.; Klug, C. A Symmoriiform from the Late Devonian of Morocco Demonstrates a Derived Jaw Function in Ancient Chondrichthyans. Commun. Biol. 2020, 3, 681. [Google Scholar] [CrossRef]
- Motta, P.J.; Wilga, C.D. Anatomy of the Feeding Apparatus of the Nurse Shark, Ginglymostoma cirratum. J. Morphol. 1999, 241, 33–60. [Google Scholar] [CrossRef]
- da Silva, J.P.C.B.; de Carvalho, M.R. Morphology and Phylogenetic Significance of the Pectoral Articular Region in Elasmobranchs (Chondrichthyes). Zool. J. Linn. Soc. 2015, 175, 525–568. [Google Scholar] [CrossRef]
- Maisey, J.G. The Anatomy and Interrelationships of Mesozoic Hybodont Sharks. Am. Mus. Novit. 1982, 2724, 1–48. [Google Scholar]
- Jambura, P.L.; Pfaff, C.; Underwood, C.J.; Ward, D.J.; Kriwet, J. Tooth Mineralization and Histology Patterns in Extinct and Extant Snaggletooth Sharks, Hemipristis (Carcharhiniformes, Hemigaleidae)—Evolutionary Significance or Ecological Adaptation? PLoS ONE 2018, 13, e0200951. [Google Scholar] [CrossRef] [PubMed]
- Jambura, P.L.; Kindlimann, R.; López-Romero, F.; Marramà, G.; Pfaff, C.; Stumpf, S.; Türtscher, J.; Underwood, C.J.; Ward, D.J.; Kriwet, J. Micro-Computed Tomography Imaging Reveals the Development of a Unique Tooth Mineralization Pattern in Mackerel Sharks (Chondrichthyes; Lamniformes) in Deep Time. Sci. Rep. 2019, 9, 9652. [Google Scholar] [CrossRef]
- Jambura, P.L.; Türtscher, J.; Kindlimann, R.; Metscher, B.; Pfaff, C.; Stumpf, S.; Weber, G.W.; Kriwet, J. Evolutionary Trajectories of Tooth Histology Patterns in Modern Sharks (Chondrichthyes, Elasmobranchii). J. Anat. 2020, 236, 753–771. [Google Scholar] [CrossRef] [PubMed]
- Casier, E. Constitution et évolution de Ia racine dentaire des Euselachii. I- Note préliminaire. Bull. Inst. R. Sci. Nat. Belg. Biol. 1947, 23, 1–15. [Google Scholar]
- Casier, E. Constitution et évolution de Ia racine dentaire des Euselachii. II- Etude comparative des types. Bull. Inst. R. Sci. Nat. Belg. Biol. 1947, 23, 1–32. [Google Scholar]
- Casier, E. Constitution et évolution de Ia racine dentaire des Euselachii. III—Evolution des principaux caractères morphologiques et conclusions. Bull. Inst. R. Sci. Nat. Belg. Biol. 1947, 23, 1–45. [Google Scholar]
- Compagno, L.J.V. Sharks of the Order Carcharhiniformes; Princeton University Press: Princeton, NJ, USA, 1988; ISBN 978-0-691-08453-4. [Google Scholar]
- Compagno, L.J.V. Alternative Life-History Styles of Cartilaginous Fishes in Time and Space. Environ. Biol. Fishes 1990, 28, 33–75. [Google Scholar] [CrossRef]
- Compagno, L.J.V. Relationships of the Megamouth Shark, Megachasma pelagios (Lamniformes: Megachasmidae), with Comments on Its Feeding Habits. In Elasmobranchs as Living Resources: Advances in the Biology, Ecology, Systematics, and the Status of the Fisheries; Pratt, H.L., Gruber, S.H., Taniuchi, T., Eds.; NOAA Technical Report NMFS; 1990; Volume 90, pp. 357–379. [Google Scholar]
- Herman, J.; Hovestadt-Euler, M.; Hovestadt, D.C. Contributions to the Study of the Comparative Morphology of Teeth and Other Relevant Ichthyodorulites in Living Supraspecific Taxa of Chondrichtyan Fishes. Part A: Selachii. No. 2a: Order: Carcharhiniformes—Family: Triakidae. Bull. Inst. R. Sci. Nat. Belg. Biol. 1988, 58, 99–126. [Google Scholar]
- Herman, J.; Hovestadt-Euler, M.; Hovestadt, D.C. Contributions to the Study of the Comparative Morphology of Teeth and Other Relevant Ichthyodorulites in Living Supraspecific Taxa of Chondrichtyan Fishes. Part A: Selachii. No. 3: Order: Squaliformes Families: Echinorhinidae, Oxynotidae and Squalidae. Bull. Inst. R. Sci. Nat. Belg. Biol. 1989, 59, 101–157. [Google Scholar]
- Herman, J.; Hovestadt-Euler, M.; Hovestadt, D.C. Contributions to the Study of the Comparative Morphology of Teeth and Other Relevant Ichthyodorulites in Living Supraspecific Taxa of Chondrichtyan Fishes. Part A: Selachii. No. 2b: Order: Carcharhiniformes—Family: Scyliorhinidae. Bull. Inst. R. Sci. Nat. Belg. Biol. 1990, 60, 181–230. [Google Scholar]
- Herman, J.; Hovestadt-Euler, M.; Hovestadt, D.C. Contributions to the Study of the Comparative Morphology of Teeth and Other Relevant Ichthyodorulites in Living Supraspecific Taxa of Chondrichtyan Fishes. Part A: Selachii. No. 2c: Order: Carcharhiniformes Families: Proscyllidae, Hemigaleidae, Pseudotriakidae, Leptochariidae and Carcharhinidae. Bull. Inst. R. Sci. Nat. Belg. Biol. 1991, 61, 73–120. [Google Scholar]
- Herman, J.; Hovestadt-Euler, M.; Hovestadt, D.C. Contributions to the Study of the Comparative Morphology of Teeth and Other Relevant Ichthyodorulites in Living Supraspecific Taxa of Chondrichthyan Fishes. Part A: Selachii. No. 4: Order: Orectolobiformes—Families: Brachaeluridae, Ginglymostomatidae, Hemiscylliidae, Orectolobidae, Parascylhidae, Rhiniodontidae, Stegostomatidae. Order: Pristiophoriformes—Family: Pristiophoridae. Order: Squatiniformes—Family: Squatinidae. Bull. Inst. R. Sci. Nat. Belg. Biol. 1992, 62, 193–254. [Google Scholar]
- Herman, J.; Hovestadt-Euler, M.; Hovestadt, D.C. Contributions to the Study of the Comparative Morphology of Teeth and Other Relevant Ichthyodorulites in Living Supraspecific Taxa of Chondrichthyan Fishes. Part A: Selachii. No. 1b: Order: Hexanchiformes—Family: Chlamydoselachidae; No. 5: Order Heterodontiformes—Family: Heterodontidae; No. 6: Order: Lamniformes—Families: Cetorhinidae, Megachasmidae; Addendum 1 to No. 3: Order Squaliformes; Addendum 1 to No. 4: Order: Orectolobiformes; General Glosary; Summary Part A. Bull. Inst. R. Sci. Nat. Belg. Biol. 1993, 63, 185–256. [Google Scholar]
- Herman, J.; Hovestadt-Euler, M.; Hovestadt, D.C.; Stehmann, M. Contributions to the Study of the Comparative Morphology of Teeth and Other Relevant Ichthyodorulites in Living Supraspecific Taxa of Chondrichtyan Fishes. Part B: Batomorphii. No. 1b: Order: Rajiformes—Suborder Rajoidei—Family: Rajidae—Genera and Subgenera: Bathyraja (with a Deep-Water, Shallow-Water and Transitional Morphotype), Psammobatis, Raja (Amblyraja), Raja (Dipturus), Raja (Leucoraja), Raja (Raja), Raja (Rajella) with Two Morphotypes), Raja (Rioraja), Raja (Rostroraja), Raja Lintea, and Sympterygia. Bull. Inst. R. Sci. Nat. Belg. Biol. 1995, 65, 237–307. [Google Scholar]
- Herman, J.; Hovestadt-Euler, M.; Hovestadt, D.C.; Stehmann, M. Contributions to the Study of the Comparative Morphology of Teeth and Other Relevant Ichthyodorulites in Living Supra-Specific Taxa of Chondrichthyan Fishes. Part B: Batomorphii No. 1c: Order Rajiformes—Suborder Rajoidei—Family: Rajidae—Genera and Subgenera: Arhynchobatis, Bathyraja Richardsoni-Type, Cruriraja, Irolita, Notoraja, Pavoraja (Insentiraja), Pavoraja (Pavoraja), Pseudoraja, Raja (Atlantoraja), Raja (Okamejei) and Rhinoraja. Bull. Inst. R. Sci. Nat. Belg. Biol. 1996, 66, 179–236. [Google Scholar]
- Herman, J.; Hovestadt-Euler, M.; Hovestadt, D.C.; Stehmann, M. Contributions to the Study of the Comparative Morphology of Teeth and Other Relevant Ichthyodorulites in Living Supra-Specific Taxa of Chondrichthyan Fishes. Part B: Batomorphii No. 2: Order Rajiformes—Suborder: Pristoidei—Family: Pristidae—Genera: Anoxypristis and Pristis No. 3: Suborder Rajoidei—Superfamily Rhinobatoidea—Families: Rhinidae—Genera: Rhina and Rhynchobatus and Rhinobatidae—Genera: Aptychotrema, Platyrhina, Platyrhinoidis, Rhinobatos, Trygonorrhina, Zanobatus and Zapteryx. Bull. Inst. R. Sci. Nat. Belg. Biol. 1997, 67, 107–162. [Google Scholar]
- Herman, J.S.; Hovestadt-Euler, M.; Hovestadt, D.C.; Stehmann, M. Contributions to the Study of the Comparative Morphology of Teeth and Other Relevant Ichthyodorulites in Living Supra-Specific Taxa of Chondrichthyan Fishes. Part B: Batomorphii 4a: Order Rajiformes—Suborder Myliobatoidei—Superfamily Dasyatoidea—Family Dasyatidae—Subfamily Dasyatinae—Genera: Amphotistius, Dasyatis, Himantura, Pastinachus, Pteroplatytrygon, Taeniura, Urogymnus and Urolophoides (Incl. Supraspecific Taxa of Uncertain Status and Validity), Superfamily Myliobatoidea—Family Gymnuridae—Genera: Aetoplatea and Gymnura, Superfamily Plesiobatoidea—Family Hexatrygonidae—Genus: Hexatrygon. Bull. Inst. R. Sci. Nat. Belg. Biol. 1998, 68, 145–197. [Google Scholar]
- Herman, J.; Hovestadt-Euler, M.; Hovestadt, D.C.; Stehmann, M. Contributions to the Study of the Comparative Morphology of Teeth and Other Relevant Ichthyodorulites in Living Supraspecific Taxa of Chondrichthyan Fishes. Part B: Batomorphii 4b: Order Rajiformes—Suborder Myliobatoidei—Superfamily Dasyatoidea—Family Dasyatidae—Subfamily Dasyatinae—Genera: Taeniura, Urogymnus, Urolophoides—Subfamily Potamotrygoninae—Genera: Disceus, Plesiotrygon, and Potamotrygon (Incl. Supraspecific Taxa of Uncertain Status and Validity), Family Urolophidae—Genera: Trygonoptera, Urolophus and Urotrygon—Superfamily Myliobatidea—Family: Gymnuridae—Genus Aetoplatea. Bull. Inst. R. Sci. Nat. Belg. Biol. 1999, 69, 161–200. [Google Scholar]
- Herman, J.; Hovestadt-Euler, M.; Hovestadt, D.C.; Stehmann Contributions to the Study of the Comparative Morphology of Teeth and Other Relevant Ichthyodorulites in Living Supraspecific Taxa of Chondrichthyan Fishes. Part B: Batomorphii 4c: Order Rajiformes—Suborder Myliobatoidei—Superfamily Dasyatoidea—Family Dasyatidae—Subfamily Dasyatinae—Genus: Urobatis, Subfamily Potamotrygoninae—Genus: Paratrygon, Superfamily Plesiobatoidea—Family Plesiobatidae—Genus: Plesiobatis, Superfamily Myliobatoidea—Family Myliobatidae—Subfamily Myliobatinae—Genera: Aetobatus, Aetoylaeus, Myliobatis, and Pteromylaeus, Subfamily Rhinopterinae—Genus: Rhinoptera and Subfamily Mobulinae—Genera: Manta and Mobula. Addendum 1 to 4a: Erratum to Genus Pteroplatytrygon. Bull. Inst. R. Sci. Nat. Belg. Biol. 2000, 70, 5–67. [Google Scholar]
- Herman, J.; Hovestadt-Euler, M.; Hovestadt, D.C.; Stehmann, M. Contributions to the Study of the Comparative Morphology of Teeth and Other Relevant Ichthyodorulites in Living Supraspecific Taxa of Chondrichthyan Fishes Part B: Batomorphii 4: Order Torpediniformes—Family Narcinidae—Subfamily Narcininae—Genera: Benthobatis, Diplobatis, Discopyge and Narcine, Subfamily Narkinae—Genera: Bengalichthys, Crassinarke, Heteronarce, Narke, Temera, and Typhlonarke, Family Torpedinidae—Subfamily Torpedininae—Genus: Torpedo—Subgenus: T.(Tetronarke) and T. (Torpedo) and Subfamily Hypninae—Genus: Hypnos. Bull. Inst. R. Sci. Nat. Belg. Biol. 2002, 72, 5–45. [Google Scholar]
- Herman, J.; Hovestadt-Euler, M.; Hovestadt, D.C. Contributions to the Study of the Comparative Morphology of Teeth and Other Relevant Ichthyodorulites in Living Supraspecific Taxa of Chondrichthyan Fishes. Part A: Selachii. Addendum to 1: Order Hexanchiformes—Family Hexanchidae, 2: Order Carcharhiniformes, 2a: Family Triakidae, 2b: Family Scyliorhinidae, 2c: Family Carcharhinidae, Hemigaleidae, Leptochariidae, Sphyrnidae, Proscyllidae and Pseudotriakidae, 3: Order Squaliformes: Family Echinorhinidae, Oxynotidae and Squalidae. Tooth Vascularization and Phylogenetic Interpretation. Bull. Inst. R. Sci. Nat. Belg. Biol. 2003, 73, 5–26. [Google Scholar]
- Maisey, J.G. Cranial Morphology of the Fossil Elasmobranch Synechodus dubrisiensis. Am. Mus. Novit. 1985, 2804, 1–28. [Google Scholar]
- Maisey, J.G. Morphology of the Braincase in the Broadnose Sevengill Shark Notorynchus (Elasmobranchii, Hexanchiformes), Based on CT Scanning. Am. Mus. Novit. 2004, 3429, 1–52. [Google Scholar] [CrossRef]
- de Carvalho, M.R.; Kriwet, J.; Thies, D. A Systematic and Anatomical Revision of Late Jurassic Angelsharks (Chondrichthyes: Squatinidae). In Mesozoic Fishes 4—Homology and Phylogeny; Arratia, G., Schultze, H.-P., Wilson, M.V.H., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2008; pp. 469–502. [Google Scholar]
- Denton, J.S.S.; Maisey, J.G.; Grace, M.; Pradel, A.; Doosey, M.H.; Bart Jr, H.L.; Naylor, G.J.P. Cranial Morphology in Mollisquama sp. (Squaliformes; Dalatiidae) and Patterns of Cranial Evolution in Dalatiid Sharks. J. Anat. 2018, 233, 15–32. [Google Scholar] [CrossRef]
- da Silva, J.P.C.B.; Vaz, D.; de Carvalho, M.R. Systematic Implications of the Anterior Pectoral Basals in Squaliform Sharks (Chondrichthyes: Elasmobranchii). Copeia 2015, 103, 874–885. [Google Scholar] [CrossRef]
- da Silva, J.P.C.B.; Vaz, D.F.B.; de Carvalho, M.R. Phylogenetic Inferences on the Systematics of Squaliform Sharks Based on Elasmobranch Scapular Morphology (Chondrichthyes: Elasmobranchii). Zool. J. Linn. Soc. 2018, 182, 614–630. [Google Scholar] [CrossRef]
- Moreira, R.A.; de Carvalho, M.R. Clasper Morphology of the Japanese Sawshark, Pristiophorus japonicus Günther, 1870 (Chondrichthyes: Elasmobranchii). Anat. Rec. 2019, 302, 1666–1670. [Google Scholar] [CrossRef]
- da Silva, J.P.C.B.; Datovo, A. The Coracoid Bar and Its Phylogenetic Importance for Elasmobranchs (Chondrichthyes). Zool. Anz. 2020, 287, 167–177. [Google Scholar] [CrossRef]
- da Silva, J.P.C.B.; Datovo, A. A Reappraisal of the Pectoral Skeleton of Lantern Sharks (Elasmobranchii: Squaliformes: Etmopteridae). J. Morphol. 2021, 282, 408–418. [Google Scholar] [CrossRef]
- Moyer, J.; Finucci, B.; Riccio, M.; Irschick, D. Dental Morphology and Microstructure of the Prickly Dogfish Oxynotus bruniensis (Squaliformes: Oxynotidae). J. Anat. 2020, 237, 916–932. [Google Scholar] [CrossRef] [PubMed]
- Soares, K.D.A.; de Carvalho, M.R. Phylogenetic Relationship of Catshark Species of the Genus Scyliorhinus (Chondrichthyes, Carcharhiniformes, Scyliorhinidae) Based on Comparative Morphology. Zoosyst. Evol. 2020, 96, 345–395. [Google Scholar] [CrossRef]
- Klimpfinger, C.; Kriwet, J. Comparative Morphology of Labial Cartilages in Sharks (Chondrichthyes, Elasmobranchii). Eur. Zool. J. 2020, 87, 741–753. [Google Scholar] [CrossRef]
- de Oliveira Lana, F.; Soares, K.D.A.; Hazin, F.H.V.; Gomes, U.L. Description of the Chondrocranium of the Silky Shark Carcharhinus falciformis with Comments on the Cranial Terminology and Phylogenetic Implications in Carcharhinids (Chondrichthyes, Carcharhiniformes, Carcharhinidae). J. Morphol. 2021, 282, 685–700. [Google Scholar] [CrossRef] [PubMed]
- Soares, K.D.A.; Mathubara, K. Combined Phylogeny and New Classification of Catsharks (Chondrichthyes: Elasmobranchii: Carcharhiniformes). Zool. J. Linn. Soc. 2022, 195, 761–814. [Google Scholar] [CrossRef]
- Staggl, M.A.; Ruthensteiner, B.; Straube, N. Head Anatomy of a Lantern Shark Wet-Collection Specimen (Chondrichthyes: Etmopteridae). J. Anat. 2023. [Google Scholar] [CrossRef]
- Zangerl, R.; Case, G. Cobelodus aculeatus (Cope), an Anacanthous Shark from Pennsylvanian Black Shales of North America. Palaeontogr. A 1976, 154, 107–157. [Google Scholar]
- Brazeau, M.D. Problematic Character Coding Methods in Morphology and Their Effects. Biol. J. Linn. Soc. 2011, 104, 489–498. [Google Scholar] [CrossRef]
- Maddison, W.P. Missing Data Versus Missing Characters in Phylogenetic Analysis. Syst. Biol. 1993, 42, 576–581. [Google Scholar] [CrossRef]
- Goloboff, P.A.; Catalano, S.A. TNT Version 1.5, Including a Full Implementation of Phylogenetic Morphometrics. Cladistics 2016, 32, 221–238. [Google Scholar] [CrossRef]
- Swafford, D.L. PAUP*. Phylogenetic Analysis Using Parsimony (* and Other Methods), Version 4; Sinauer Associates: Sunderland, MA, USA, 2003. [Google Scholar]
- Lewis, P.O. A Likelihood Approach to Estimating Phylogeny from Discrete Morphological Character Data. Syst. Biol. 2001, 50, 913–925. [Google Scholar] [CrossRef] [PubMed]
- Compagno, L.J.V. Endoskeleton. In Sharks, Skates, and Rays: The Biology of Elasmobranch Fishes; Hamlett, W.C., Ed.; The Johns Hopkins University Press: Baltimore, MD, USA, 1999; pp. 69–92. [Google Scholar]
- Moreira, R.A.; Gomes, U.L.; de Carvalho, M.R. Systematic Implications of the Caudal Fin Skeletal Anatomy in Ground Sharks, Order Carcharhiniformes (Chondrichthyes: Elasmobranchii). Zool. J. Linn. Soc. 2019, 185, 193–211. [Google Scholar] [CrossRef]
- Batchelor, T.J.; Ward, D.J. Fish Remains from a Temporary Exposure of Hythe Beds (Aptian-Lower Creaceous) near Godstone, Surrey. Mesoz. Res. 1990, 2, 181–203. [Google Scholar]
- Huxley, T.H. On the Application of the Laws of Evolution to the Arrangement of the Vertebrata, and More Particularly of the Mammalia. Proc. Zool. Soc. Lond. 1880, 1880, 649–661. [Google Scholar]
- Maisey, J.G. What Is an ‘Elasmobranch’? The Impact of Palaeontology in Understanding Elasmobranch Phylogeny and Evolution. J. Fish Biol. 2012, 80, 918–951. [Google Scholar] [CrossRef] [PubMed]
- Underwood, C.J.; Ward, D.J. Neoselachian Sharks and Rays from the British Bathonian (Middle Jurassic). Palaeontology 2004, 47, 447–501. [Google Scholar] [CrossRef]
- Underwood, C.J. Sharks, Rays and a Chimaeroid from the Kimmeridgian (Late Jurassic) of Ringstead, Southern England. Palaeontology 2002, 45, 297–325. [Google Scholar] [CrossRef]
- Delsate, D. Une Nouvelle Faune de Poissons et Requins Toarciens Du Sud Du Luxembourg (Dudelange) et de l’Allemagne (Schömberg). Bull. Académie Lorraine Sci. 2003, 42, 13–49. [Google Scholar]
- Underwood, C.J.; Ward, D.J. Environmental Distribution of Bathonian (Middle Jurassic) Neoselachians in Southern England. In Mesozoic Fishes 3—Systematics, Palaeoenvironments and Biodiversity; Tintori, A., Arratia, G., Eds.; Verlag Dr. Friedrich Pfeil: München, Germany, 2004; pp. 111–122. [Google Scholar]
- Delsate, D.; Lepage, J.C. Découverte d’une faune originale d’Elasmobranches dans les phosphates du Toarcien lorrain (couches à Coeloceras crassum). Bull. Académie Société Lorraines Sci. 1990, 29, 153–161. [Google Scholar]
- Thies, D. Some Problematical Sharks Teeth (Chondrichthyes, Neoselachii) from the Early and Middle Jurassic of Germany. Paläontol. Z. 1989, 63, 103–117. [Google Scholar] [CrossRef]
- Young, T. Neoselachian Sharks from the Middle Jurassic of England. Sheff. Univ. Geol. Soc. J. 1982, 8, 24–31. [Google Scholar]
- Rees, J. Palaeoecological Implications of Neoselachian Shark Teeth from the Bathonian (Middle Jurassic) Ore-Bearing Clays at Gnaszyn, Kraków-Silesia Homocline, Poland. Acta Geol. Pol. 2012, 62, 397–402. [Google Scholar] [CrossRef]
- Kriwet, J. Neoselachian Remains (Chondrichthyes, Elasmobranchii) from the Middle Jurassic of SW Germany and NW Poland. Acta Palaeontol. Pol. 2003, 48, 583–594. [Google Scholar]
- Popov, E.; Seltser, V.B.; Volkov, A. On Records of Elasmobranchs Teeth (Chondrichthyes: Elasmobranchii) from the Lower Callovian of Saratov, Russia [In Russian with English Summary and Explanations of Figures]. Trans. Sci. Res. Geol. Inst. N G Chernyshevskii Saratov State Univ. New Ser. 2004, 14, 133–154. [Google Scholar]
- Rees, J. Neoselachian Sharks from the Callovian-Oxfordian (Jurassic) of Ogrodzieniec, Zawiercie Region, Southern Poland. Palaeontology 2010, 53, 887–902. [Google Scholar] [CrossRef]
- Duffin, C.J. New Records of Late Jurassic Sharks Teeth from Southern Germany. Stuttg. Beitr. Zur Naturkunde Ser. B 1993, 193, 1–13. [Google Scholar]
- Mudroch, A. Fischzähne aus dem Oberjura Nordwesteuropas: Systematik, Biogeochemie und Palökologie. Doctoral Thesis, University of Hannover: Hannover, Germany, 2001. [Google Scholar]
- Klug, S.; Kriwet, J. An Offshore Fish Assemblage (Elasmobranchii, Actinopterygii) from the Late Jurassic of NE Spain. Palaontologische Z. 2013, 87, 235–257. [Google Scholar] [CrossRef]
- Candoni, L. Deux faunes inédites de sélaciens dans le Jurassique terminal Français-Premiers résultats stratigraphiques. Bull. Trimest. Société Géologique Normandie Amis Muséum Havre 1995, 82, 29–49. [Google Scholar]
- Kriwet, J. Late Jurassic Elasmobranch and Actinopterygian Fishes from Portugal and Spain. Cuad. Geol. Ibérica 1998, 24, 241–260. [Google Scholar]
- Barthel, K.W. Solnhofen: Ein Blick in die Erdgeschichte; Ott Verlag: Thun, Switzerland, 1978; ISBN 3-7225-6253-8. [Google Scholar]
- Cappetta, H. Handbook of Paleoichthyology, Vol. 3B: Chondrichthyes II.; Gustav Fischer Verlag: Stuttgart, Germany, 1987. [Google Scholar]
- Carroll, R.L. Vertebrate Paleontology and Evolution; W. H. Freeman and Company: Dallas, TX, USA, 1988. [Google Scholar]
- Frickhinger, K.A. Die Fossilien von Solnhofen: Dokumentation der aus den Plattenkalken bekannten Tiere und Pflanzen; Goldschneck-Verlag: Korb, Australia, 1994; ISBN 978-3-926129-15-4. [Google Scholar]
- Frickhinger, K.A. Die Fossilien von Solnhofen-Dokumentation der aus den Plattenkalken bekannten Tiere und Pflanzen, Band 2: Neue Funde, neue Details, neue Erkenntnisse; Goldschneck-Verlag: Korb, Australia, 1999; ISBN 3-926129-26-3. [Google Scholar]
- Thies, D.; Leidner, A. Sharks and Guitarfishes (Elasmobranchii) from the Late Jurassic of Europe. Palaeodiversity 2011, 4, 63–184. [Google Scholar]
- Leuzinger, L.; Cuny, G.; Popov, E.; Billon-Bruyat, J.-P. A New Chondrichthyan Fauna from the Late Jurassic of the Swiss Jura (Kimmeridgian) Dominated by Hybodonts, Chimaeroids and Guitarfishes. Pap. Palaeontol. 2017, 3, 471–511. [Google Scholar] [CrossRef]
- Sharp, D. The Zoological Record; Zoological Society of London: London, UK, 1918; Volume 55. [Google Scholar]
- Jordan, D.S.; Evermann, B.W. The Genera of Fishes. A Contribution to the Stability of Scientific Nomenclature; Stanford University Press: Redwood City, CA, USA, 1917. [Google Scholar]
- Dickinson, E.C.; Overstreet, L.K.; Dowsett, R.J.; Bruce, M.D. Priority! The Dating of Scientific Names in Ornithology: A Directory to the Literature and Its Reviewers; Aves Press: Northampton, UK, 2011; ISBN 978-0-9568611-1-5. [Google Scholar]
- ICZN, 1999, Internatl. Code Zool. Nomencl., Ed. 4|Amphibian Species of the World. Available online: https://amphibiansoftheworld.amnh.org/Bibliography/I/ICZN-1999-Internatl.-Code-Zool.-Nomencl.-Ed.-4 (accessed on 17 November 2022).
- Maisey, J.G. An Evaluation of Jaw Suspension in Sharks. Am. Mus. Novit. 1980, 2706, 1–17. [Google Scholar]
- Vullo, R.; Guinot, G. Denticle-Embedded Ampullary Organs in a Cretaceous Shark Provide Unique Insight into the Evolution of Elasmobranch Electroreceptors. Sci. Nat. 2015, 102, 65. [Google Scholar] [CrossRef] [PubMed]
- Vullo, R.; Guinot, G.; Barbe, G. The First Articulated Specimen of the Cretaceous Mackerel Shark Haimirichia amonensis gen. nov. (Haimirichiidae Fam. Nov.) Reveals a Novel Ecomorphological Adaptation within the Lamniformes (Elasmobranchii). J. Syst. Palaeontol. 2016, 14, 1003–1024. [Google Scholar] [CrossRef]
- Macías-Cuyare, M.; Oddone, M.C. Morphological Pattern of the Dermal Denticles of the Southern Sawtail Catshark Galeus Mincaronei Soto, 2001. J. Morphol. 2022, 283, 1120–1134. [Google Scholar] [CrossRef]
- Reif, W.-E. Morphogenesis and Histology of Large Scales of Batoids (Elasmobranchii). Paläontol. Z. 1979, 53, 26–37. [Google Scholar] [CrossRef]
- Vaz, D.; de Carvalho, M.R. New Species of Squatina (Squatiniformes: Squatinidae) from Brazil, with Comments on the Taxonomy of Angel Sharks from the Central and Northwestern Atlantic. Copeia 2018, 106, 144–160. [Google Scholar] [CrossRef]
- Reif, W.-E. Protective and Hydrodynamic Function of the Dermal Skeleton of Elasmobranchs. Neues Jahrb. Für Geol. Paläontol. Abh. 1978, 157, 133–141. [Google Scholar]
- Raschi, W.; Tabit, C. Functional Aspects of Placoid Scales: A Review and Update. Mar. Freshw. Res. 1992, 43, 123–147. [Google Scholar] [CrossRef]
- Reif, W.-E. Morphologie und Ultrastruktur des Hai- “Schmelzes”. Zool. Scr. 1973, 2, 231–250. [Google Scholar] [CrossRef]
- Enault, S.; Guinot, G.; Koot, M.B.; Cuny, G. Chondrichthyan Tooth Enameloid: Past, Present, and Future. Zool. J. Linn. Soc. 2015, 174, 549–570. [Google Scholar] [CrossRef]
- Cuny, G.; Guinot, G.; Enault, S. Evolution of Dental Tissues and Paleobiology in Selachians; Elsevier: Oxford, UK, 2018. [Google Scholar]
- Hampe, O.; Long, J.A. The Histology of Middle Devonian Chondrichthyan Teeth from Southern Victoria Land, Antarctica. Rec. West. Aust. Mus. 1999, 57, 23–36. [Google Scholar]
- Hampe, O. Revision of the Xenacanthida (Chondrichthyes: Elasmobranchii) from the Carboniferous of the British Isles. Trans. R. Soc. Edinb. Earth Sci. 2002, 93, 191–237. [Google Scholar] [CrossRef]
- Manzanares, E.; Rasskin-Gutman, D.; Botella, H. New Insights into the Enameloid Microstructure of Batoid Fishes (Chondrichthyes). Zool. J. Linn. Soc. 2016, 177, 621–632. [Google Scholar] [CrossRef]
- Enault, S.; Cappetta, H.; Adnet, S. Simplification of the Enameloid Microstructure of Large Stingrays (Chondrichthyes: Myliobatiformes): A Functional Approach: Enameloid Microstructure of Stingrays. Zool. J. Linn. Soc. 2013, 169, 144–155. [Google Scholar] [CrossRef]
- Greenfield, T.; Delsate, D.; Candoni, L. Toarcibatidae fam. nov., a Replacement for the Unavailable Name Archaeobatidae Delsate & Candoni, 2001 (Chondrichthyes, Batomorphii). Zootaxa 2022, 5195, 499–500. [Google Scholar] [CrossRef]
- Manzanares, E.; Botella, H.; Delsate, D. On the Enameloid Microstructure of Archaeobatidae (Neoselachii, Chondrichthyes). J. Iber. Geol. 2018, 44, 67–74. [Google Scholar] [CrossRef]
- Thies, D.; Vespermann, J.; Solcher, J. Two New Neoselachian Sharks (Elasmobranchii, Neoselachii, Synechodontiformes) from the Rhaetian (Late Triassic) of Europe. Palaeontogr. Abt. A 2014, 303, 137–167. [Google Scholar] [CrossRef]
- Hodnett, J.-P.M.; Elliott, D.K.; Olson, T.J.; Wittke, J.H. Ctenacanthiform Sharks from the Permian Kaibab Formation, Northern Arizona. Hist. Biol. 2012, 24, 381–395. [Google Scholar] [CrossRef]
- Guinot, G.; Adnet, S.; Cavin, L.; Cappetta, H. Cretaceous Stem Chondrichthyans Survived the End-Permian Mass Extinction. Nat. Commun. 2013, 4, 2669. [Google Scholar] [CrossRef]
- Feichtinger, I.; Engelbrecht, A.; Lukeneder, A.; Kriwet, J. New Chondrichthyans Characterised by Cladodont-like Tooth Morphologies from the Early Cretaceous of Austria, with Remarks on the Microstructural Diversity of Enameloid. Hist. Biol. 2020, 32, 823–836. [Google Scholar] [CrossRef]
- da Cunha, D.B.; da Silva Rodrigues-Filho, L.F.; de Luna Sales, J.B. A Review of the Mitogenomic Phylogeny of the Chondrichthyes. In Chondrichthyes Multidisciplinary Approach; da Silva Rodrigues-Filho, L.F., de Luna Sales, J.B., Eds.; Intech Open: Rijeka, Croatia, 2017; pp. 113–125. [Google Scholar]
- Li, C.; Matthes-Rosana, K.A.; Garcia, M.; Naylor, G.J.P. Phylogenetics of Chondrichthyes and the Problem of Rooting Phylogenies with Distant Outgroups. Mol. Phylogenet. Evol. 2012, 63, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Regan, C.T. A Classification of the Selachian Fishes. Proc. Zool. Soc. Lond. 1906, 722–758. [Google Scholar]
- Bigelow, H.B.; Schroeder, W.C. Sharks. In Fishes of the Western North Atlantic. Part 1; Tee-Van, J., Breder, C.M., Hildebrand, S.F., Parr, A.E., Schroeder, W.C., Eds.; Sears Foundation for Marine Research, Yale University: New Haven, CT, USA, 1948; pp. 59–576. [Google Scholar]
- Nelson, J.S. Fishes of the World, 1st ed.; Wiley-Interscience: New York, NY, USA, 1976. [Google Scholar]
- Nelson, J.S. Fishes of the World, 2nd ed.; John Wiley & Sons: New York, NY, USA, 1984. [Google Scholar]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the World, 5th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2016. [Google Scholar]
- Wu, E.H. Kinematic Analysis of Jaw Protrusion in Orectolobiform Sharks: A New Mechanism for Jaw Protrusion in Elasmobranchs. J. Morphol. 1994, 222, 175–190. [Google Scholar] [CrossRef] [PubMed]
- Véran, M. Les cartilages labiaux des Chondrichthyens sont-ils homologues de l’os labial des Actinoptérygiens primitifs fossiles? Geobios 1995, 28, 161–166. [Google Scholar] [CrossRef]
- Shimada, K.; Rigsby, C.K.; Kim, S.H. Labial Cartilages in the Smalltooth Sandtiger Shark, Odontaspis ferox (Lamniformes: Odontaspididae) and Their Significance to the Phylogeny of Lamniform Sharks. Anat. Rec. 2009, 292, 813–817. [Google Scholar] [CrossRef]
- Koken, E. Ueber Hybodus. Geol. Paläontol. Abh. 1907, 5, 259–276. [Google Scholar]
- Maisey, J.G. Anatomical Revision of the Fossil Shark Hybodus fraasi (Chondrichthyes: Elasmobranchii). Am. Mus. Novit. 1986, 2857, 1–16. [Google Scholar]
- Maisey, J.G. Cranial Anatomy of the Lower Jurassic Shark Hybodus reticulatus (Chondrichthyes: Elasmobranchii), with Comments on Hybodontid Systematics. Am. Mus. Novit. 1987, 2878, 1–39. [Google Scholar]
- Tapanila, L.; Pruitt, J.; Pradel, A.; Wilga, C.D.; Ramsay, J.B.; Schlader, R.; Didier, D.A. Jaws for a Spiral-Tooth Whorl: CT Images Reveal Novel Adaptation and Phylogeny in Fossil Helicoprion. Biol. Lett. 2013, 9, 20130057. [Google Scholar] [CrossRef]
- Coates, M.I.; Tietjen, K.; Olsen, A.M.; Finarelli, J.A. High-Performance Suction Feeding in an Early Elasmobranch. Sci. Adv. 2019, 5, eaax2742. [Google Scholar] [CrossRef] [PubMed]
- Grogan, E.D.; Lund, R. Debeerius Ellefseni (fam. nov., gen. nov., spec. nov.), an Autodiastylic Chondrichthyan from the Mississippian Bear Gulch Limestone of Montana (USA), the Relationships of the Chondrichthyes, and Comments on Gnathostome Evolution. J. Morphol. 2000, 243, 219–245. [Google Scholar] [CrossRef]
- Garman, S. The Plagiostomia (Sharks, Skates and Rays). Mem. Mus. Comp. Zoöl. Harv. Coll. 1913, 36, 1–528. [Google Scholar]
- Dean, M.N.; Huber, D.R.; Nance, H.A. Functional Morphology of Jaw Trabeculation in the Lesser Electric Ray Narcine brasiliensis, with Comments on the Evolution of Structural Support in the Batoidea. J. Morphol. 2006, 267, 1137–1146. [Google Scholar] [CrossRef]
- Tomita, T.; Nakamura, M.; Miyamoto, K.; Nozu, R.; Toda, M.; Murakumo, K.; Sato, K. Clasper Pocket: Adaptation of a Novel Morphological Feature by Lamnoid Sharks, Which Aids with Tuna-like Swimming. Zoomorphology 2021, 140, 365–371. [Google Scholar] [CrossRef]
- Pavan-Kumar, A.; Gireesh-Babu, P.; Babu, P.P.S.; Jaiswar, A.K.; Hari Krishna, V.; Prasasd, K.P.; Chaudhari, A.; Raje, S.G.; Chakraborty, S.K.; Krishna, G.; et al. Molecular Phylogeny of Elasmobranchs Inferred from Mitochondrial and Nuclear Markers. Mol. Biol. Rep. 2014, 41, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Ridgewood, W.G. Note on the Basibranchial Skeleton of Echinorhinus spinosus. Anat. Anz. 1899, 15, 346–348. [Google Scholar]
- Varoujean, D.H. Systematics of the Genus Echinorhinus Blainville, Based on a Study of the Prickly Shark Echinorhinus cookei Pietschmann. Unpublished Thesis, Fresno State College, Fresno, CA, USA, 1972. [Google Scholar]
- de Buen, F. Catálogo Ictiológico del Mediterráneo Español y de Marruecos: Recopilando lo Publicado Sobre Peces de las Costas Mediterránea y Próximas del Atlántico (Mar de España); “Mateu” Artes e Industrias Gráficas: Madrid, Spain, 1926. [Google Scholar]
- Bernardi, G.; Powers, D. Molecular Phylogeny of the Prickly Shark, Echinorhinus cookei, Based on a Nuclear (18s RRNA) and a Mitochondrial (Cytochrome b) Gene. Mol. Phylogenet. Evol. 1992, 1, 161–167. [Google Scholar] [CrossRef]
- Claeson, K.M.; Hilger, A. Morphology of the Anterior Vertebral Region in Elasmobranchs: Special Focus, Squatiniformes. Foss. Rec. 2011, 14, 129–140. [Google Scholar] [CrossRef]
- de Carvalho, M.R.; Faro, C.; Gomes, U.L. Comparative Neurocranial Morphology of Angelsharks from the South-Western Atlantic Ocean (Chondrichthyes, Elasmobranchii, Squatinidae): Implications for Taxonomy and Phylogeny. Acta Zool. 2012, 93, 171–183. [Google Scholar] [CrossRef]
- Maisey, J.G.; Ehret, D.J.; Denton, J.S.S. A New Genus of Late Cretaceous Angel Shark (Elasmobranchii; Squatinidae), with Comments on Squatinid Phylogeny. Am. Mus. Novit. 2020, 2020, 1–29. [Google Scholar] [CrossRef]
- Maisey, J.G. Higher Elasmobranch Phylogeny and Biostratigraphy. Zool. J. Linn. Soc. 1984, 82, 33–54. [Google Scholar] [CrossRef]
- Maisey, J.G.; Janvier, P.; Pradel, A.; Denton, J.S.S.; Bronson, A.; Miller, R.; Burrow, C.J. Doliodus and Pucapampellids: Contrasting Perspectives on Stem Chondrichthyan Morphology. In Evolution and Development of Fishes; Underwood, C., Richter, M., Johanson, Z., Eds.; Cambridge University Press: Cambridge, UK, 2019; pp. 87–109. ISBN 978-1-107-17944-8. [Google Scholar]
- Holmgren, N. Studies on the Head in Fishes. Embryological, Morphological, and Phylogenetical Researches. Part II. Comparative Anatomy of the Adult Selachian Skull, with Remarks on the Dorsal Fins in Sharks. Acta Zool. 1941, 22, 1–100. [Google Scholar] [CrossRef]
- Compagno, L.J.V. Phyletic Relationships of Living Sharks and Rays. Am. Zool. 1977, 17, 303–322. [Google Scholar] [CrossRef]
- Holmgren, N. Studies on the Head in Fishes. Embryological, Morphological, and Phylogenetical Researches. Part I. Development of the Skull in Sharks and Rays. Acta Zool. 1940, 21, 51–266. [Google Scholar] [CrossRef]
- Schaeffer, B. The Xenacanth Shark Neurocranium, with Comments on Elasmobranch Monophyly. Bull. Am. Mus. Nat. Hist. 1981, 169, 1–66. [Google Scholar]
- Allis, E.P. The Cranial Anatomy of Chlamydoselachus anguineus. Acta Zool. 1923, 4, 123–221. [Google Scholar] [CrossRef]
- Mollen, F.H.; van Bakel, B.W.M.; Jagt, J.W.M. A Partial Braincase and Other Skeletal Remains of Oligocene Angel Sharks (Chondrichthyes, Squatiniformes) from Northwest Belgium, with Comments on Squatinoid Taxonomy. Contrib. Zool. 2016, 85, 147–171. [Google Scholar] [CrossRef]
- Iselstöger, H. Das Neurocranium von Rhina squatina und einige Bemerkungen über ihre systematische Stellung. Zool. Jahrb. Abt. Für Anat. Ontog. Tiere 1937, 62, 349–394. [Google Scholar]
- Underwood, C.J.; Johanson, Z.; Welten, M.; Metscher, B.; Rasch, L.J.; Fraser, G.J.; Smith, M.M. Development and Evolution of Dentition Pattern and Tooth Order in the Skates And Rays (Batoidea; Chondrichthyes). PLoS ONE 2015, 10, e0122553. [Google Scholar] [CrossRef]
- Underwood, C.; Johanson, Z.; Smith, M.M. Cutting Blade Dentitions in Squaliform Sharks Form by Modification of Inherited Alternate Tooth Ordering Patterns. R. Soc. Open Sci. 2016, 3, 160385. [Google Scholar] [CrossRef]
- Smith, M.M.; Underwood, C.; Clark, B.; Kriwet, J.; Johanson, Z. Development and Evolution of Tooth Renewal in Neoselachian Sharks as a Model for Transformation in Chondrichthyan Dentitions. J. Anat. 2018, 232, 891–907. [Google Scholar] [CrossRef]
- Shirai, S.; Okamura, O. Anatomy of Trigonognathus kabeyai, with Comments on Feeding Mechanism and Phylogenetic Relationships (Elasmobranchii, Squalidae). Japan J. Ichthyol. 1992, 39, 139–150. [Google Scholar] [CrossRef]
- Naylor, G.J.P.; Ryburn, J.A.; Fedrigo, O.; Lopez, A. Phylogenetic Relationships among the Major Lineages of Modern Elasmobranchs. In Reproductive Biology and Phylogeny of Chondrichthyes: Sharks, Batoids, and Chimaeras; Hamlett, W.C., Ed.; CRC Press: Boca Raton, FL, USA, 2005; pp. 1–25. ISBN 978-1-57808-314-5. [Google Scholar]
- Adnet, S.; Cappetta, H.; Guinot, G.; Notarbartolo di Sciara, G. Evolutionary History of the Devilrays (Chondrichthyes: Myliobatiformes) from Fossil and Morphological Inference. Zool. J. Linn. Soc. 2012, 166, 132–159. [Google Scholar] [CrossRef]
- de Beaumont, G. Observations préliminaires sur trois Sélaciens nouveaux du calcaire lithographique d’Eichstätt (Bavière). Eclogae Geol. Helv. 1960, 53, 315–328. [Google Scholar]
- Duffin, C.J. The Upper Jurassic Selachian Palaeocarcharias de Beaumont (1960). Zool. J. Linn. Soc. 1988, 94, 271–286. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
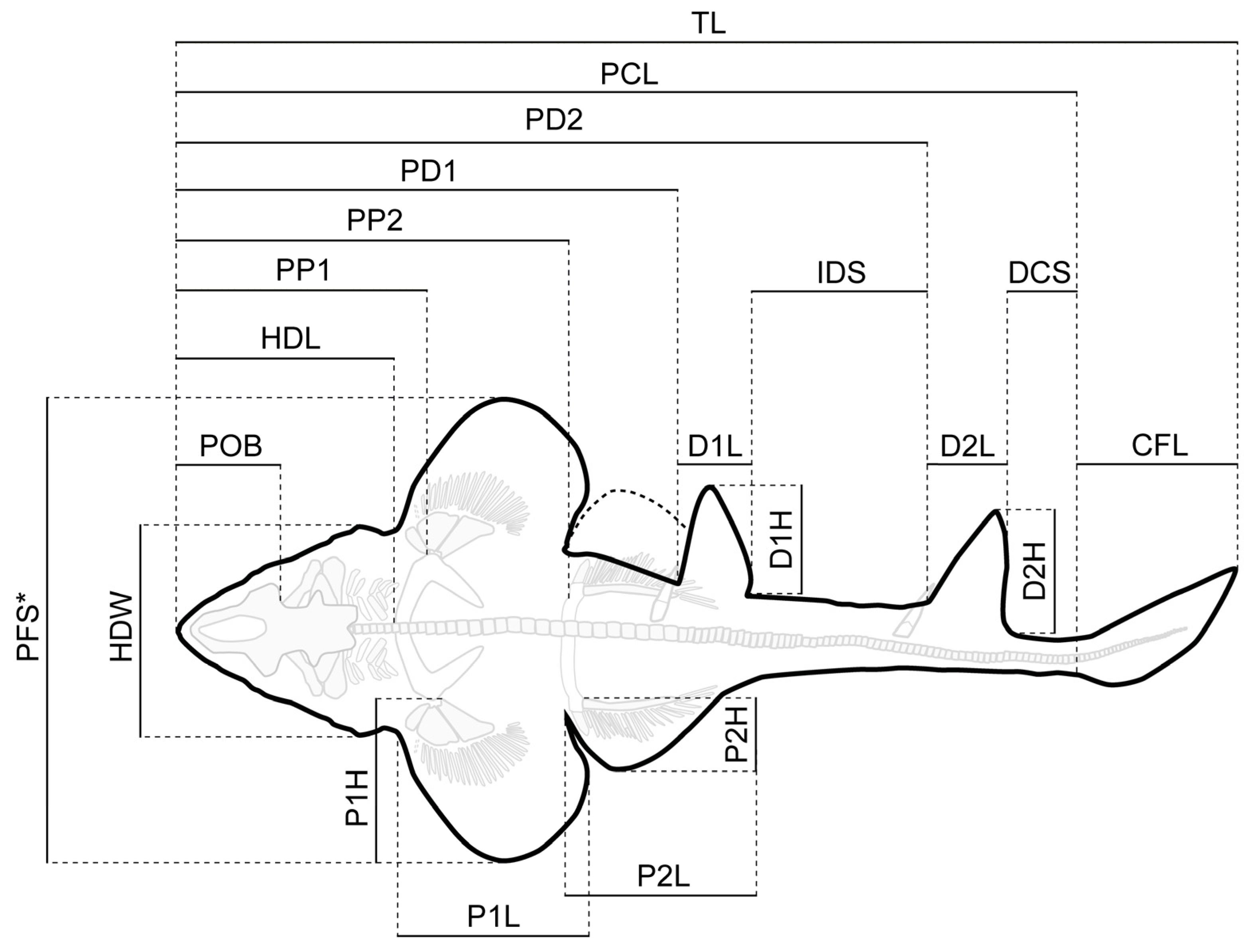
| PBP-SOL-8007 | MB 14-12-22-1 | UMN Uncatalogued | NHMUK PV P 8775 * | |||||
|---|---|---|---|---|---|---|---|---|
| Measurements | cm | % of PCL | cm | % of PCL | cm | % of PCL | cm | % of PCL |
| TL | 103.2 | 116.96 | - | - | 163.3 | 118.33 | - | - |
| PCL | 88.2 | 100.00 | 119.1 | 100.00 | 138.0 | 100.00 | 84.6 | 100 |
| POB | 9.8 | 11.05 | 13.4 | 11.21 | 15.0 | 10.86 | 10.6 | 12.54 |
| HDL | 22.3 | 25.31 | 30.4 | 25.55 | 33.9 | 24.58 | 27.5 | 32.6 |
| HDW | 17.0 | 19.29 | 20.7 | 17.37 | 22.9 | 16.56 | 19.9 | 23.5 |
| PP1 | 25.9 | 29.38 | 33.2 | 27.91 | 36.4 | 26.38 | - | - |
| P1L | 18.5 | 20.93 | 26.8 | 22.47 | 28.8 | 20.85 | 18.7 | 22.09 |
| P1H | 17.9 | 20.26 | 22.2 | 18.66 | 27.2 | 19.74 | 13.6 | 16.02 |
| PFS | 46.1 | 52.25 | 59.6 | 50.04 | 74.2 | 53.79 | 41.8 | 49.4 |
| PD1 | 48.9 | 55.40 | 63.6 | 53.39 | 77.6 | 56.21 | - | - |
| D1L | 8.4 | 9.48 | 12.5 | 10.45 | 12.0 | 8.73 | - | - |
| D1H | 12.5 | 14.18 | 13.6 | 11.40 | 15.2 | 10.98 | - | - |
| IDS | 15.8 | 17.95 | 23.6 | 19.85 | 24.4 | 17.65 | - | - |
| PD2 | 73.2 | 82.97 | 98.6 | 82.82 | 114.3 | 82.85 | - | - |
| D2L | 8.1 | 9.18 | 10.7 | 9.01 | 11.5 | 8.36 | - | - |
| D2H | 11.5 | 12.99 | 15.2 | 12.74 | 17.4 | 12.64 | - | - |
| DCS | 6.7 | 7.60 | 9.1 | 7.67 | 13.5 | 9.78 | - | - |
| PP2 | 40.1 | 45.51 | 50.4 | 42.27 | 56.7 | 41.08 | 41.4 | 48.96 |
| P2L | 19.0 | 21.57 | 28.6 | 24.04 | 29.5 | 21.37 | 15.7 | 18.61 |
| P2H | 7.3 | 8.21 | 11.5 | 9.64 | 10.5 | 7.63 | 7.5 | 8.91 |
| CFL | 15.0 | 16.96 | - | - | 25.0 | 18.13 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jambura, P.L.; Villalobos-Segura, E.; Türtscher, J.; Begat, A.; Staggl, M.A.; Stumpf, S.; Kindlimann, R.; Klug, S.; Lacombat, F.; Pohl, B.; et al. Systematics and Phylogenetic Interrelationships of the Enigmatic Late Jurassic Shark Protospinax annectans Woodward, 1918 with Comments on the Shark–Ray Sister Group Relationship. Diversity 2023, 15, 311. https://doi.org/10.3390/d15030311
Jambura PL, Villalobos-Segura E, Türtscher J, Begat A, Staggl MA, Stumpf S, Kindlimann R, Klug S, Lacombat F, Pohl B, et al. Systematics and Phylogenetic Interrelationships of the Enigmatic Late Jurassic Shark Protospinax annectans Woodward, 1918 with Comments on the Shark–Ray Sister Group Relationship. Diversity. 2023; 15(3):311. https://doi.org/10.3390/d15030311
Chicago/Turabian StyleJambura, Patrick L., Eduardo Villalobos-Segura, Julia Türtscher, Arnaud Begat, Manuel Andreas Staggl, Sebastian Stumpf, René Kindlimann, Stefanie Klug, Frederic Lacombat, Burkhard Pohl, and et al. 2023. "Systematics and Phylogenetic Interrelationships of the Enigmatic Late Jurassic Shark Protospinax annectans Woodward, 1918 with Comments on the Shark–Ray Sister Group Relationship" Diversity 15, no. 3: 311. https://doi.org/10.3390/d15030311
APA StyleJambura, P. L., Villalobos-Segura, E., Türtscher, J., Begat, A., Staggl, M. A., Stumpf, S., Kindlimann, R., Klug, S., Lacombat, F., Pohl, B., Maisey, J. G., Naylor, G. J. P., & Kriwet, J. (2023). Systematics and Phylogenetic Interrelationships of the Enigmatic Late Jurassic Shark Protospinax annectans Woodward, 1918 with Comments on the Shark–Ray Sister Group Relationship. Diversity, 15(3), 311. https://doi.org/10.3390/d15030311