



Is Shrub Encroachment Driving the Decline of Small Mammal Diversity in Pyrenean Grasslands? A Preliminary Study



Abstract
1. Introduction
2. Materials and Methods
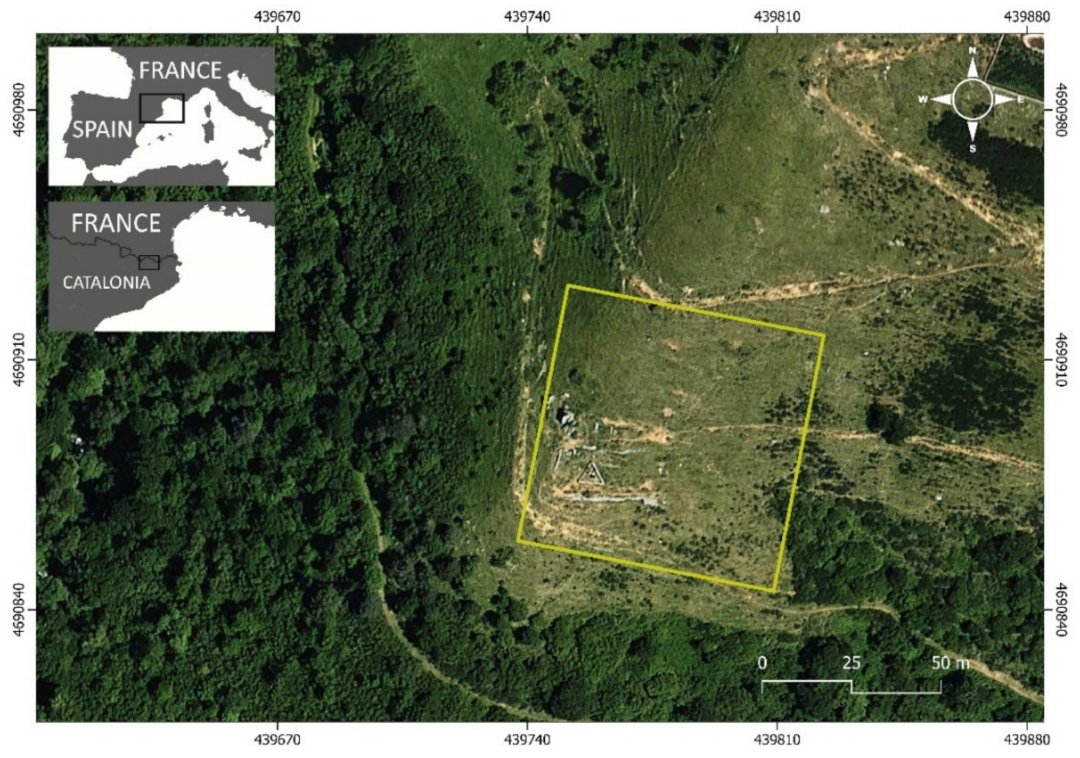
2.1. Study Area and Sampling Design
2.2. Data Analysis
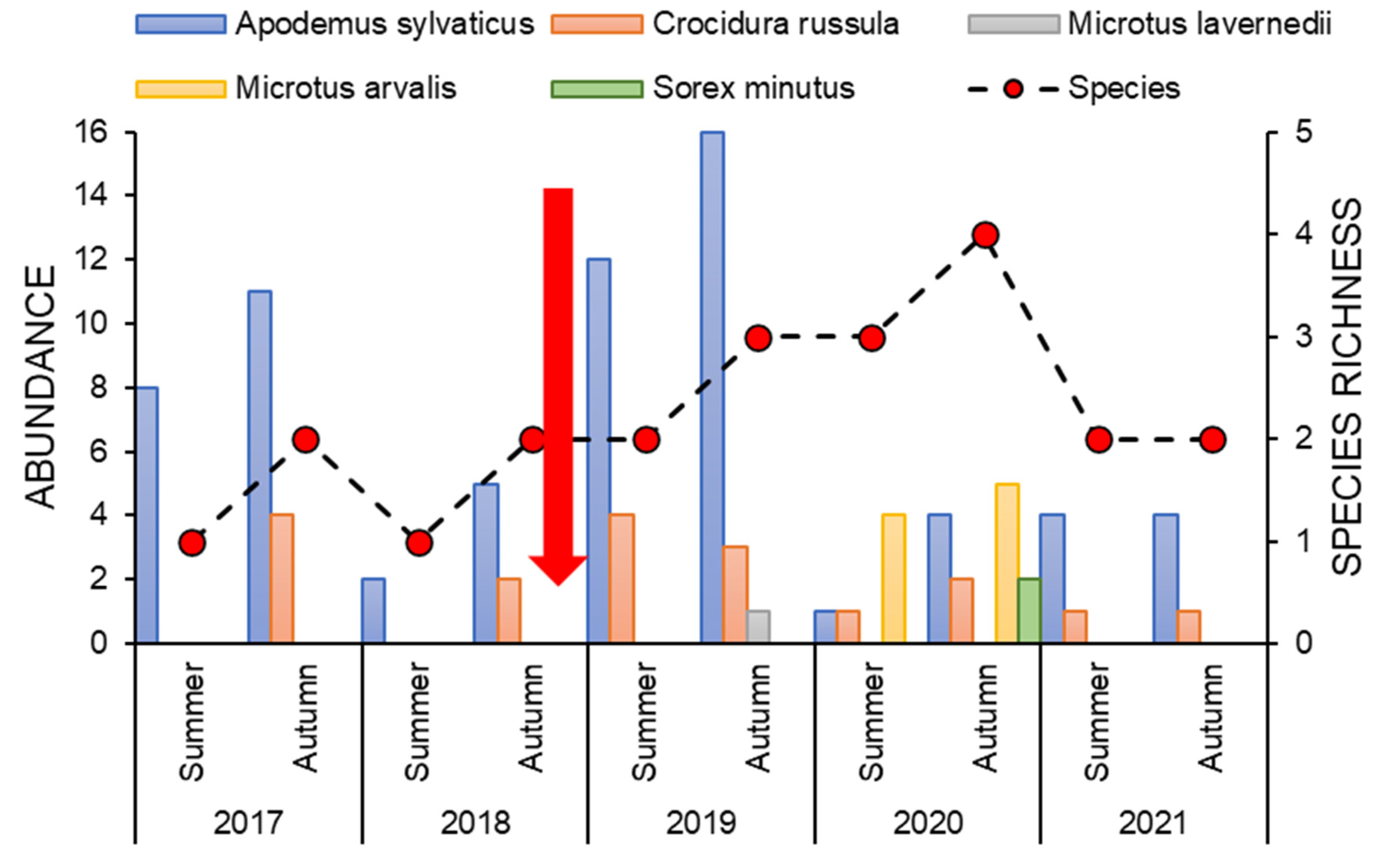
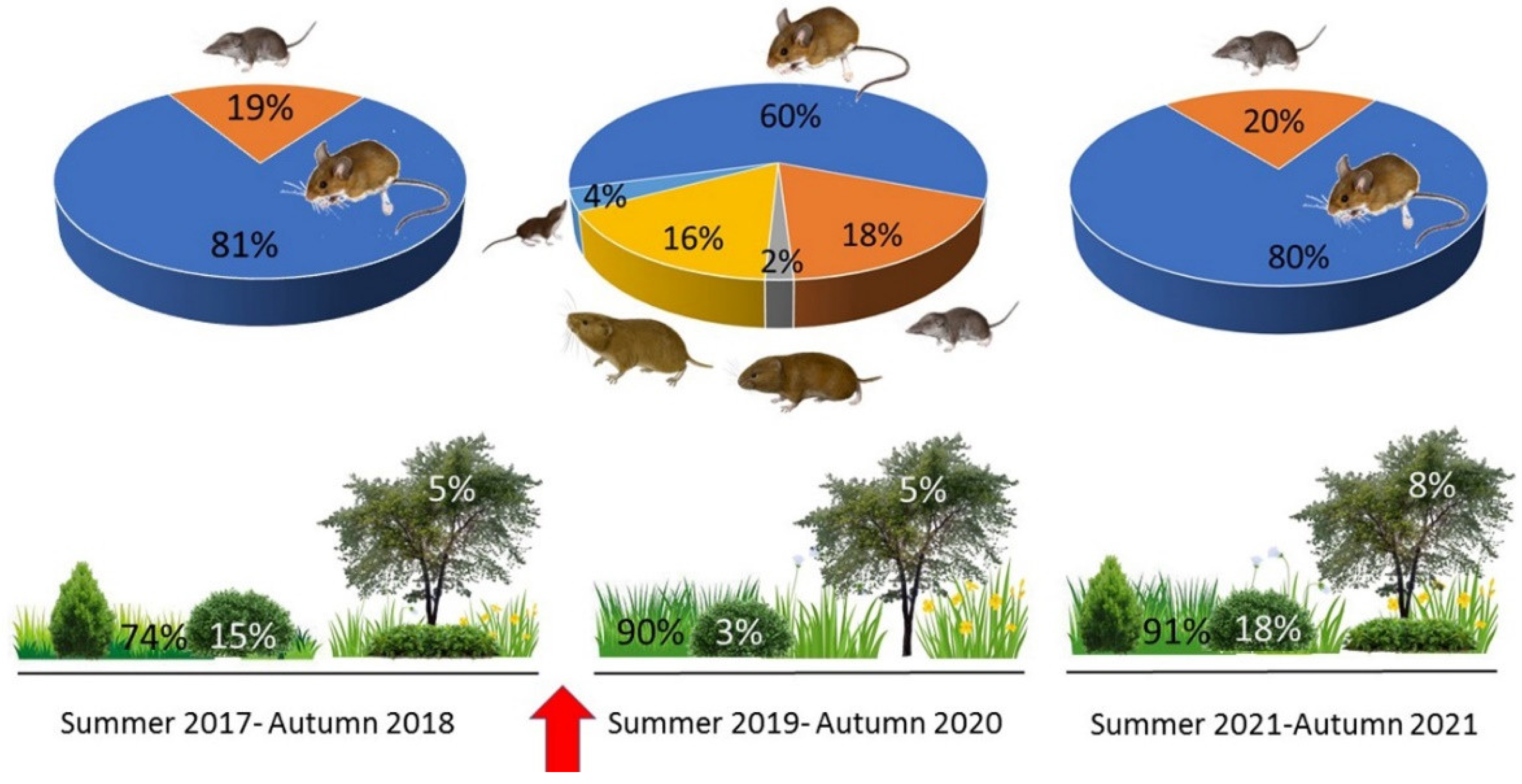
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blondel, J.; Aronson, J.; Bodiou, J.-Y.; Boeuf, G.; The Mediterranean Region. Biological Diversity in Space and Time. Available online: https://global.oup.com/academic/product/the-mediterranean-region-9780199557998?cc=es&lang=en& (accessed on 8 December 2021).
- Pascual, L.-L.; Maiorano, L.; Alessandra, F.; Emilio, B.; Luigi, B. Hotspots of species richness, threat and endemism for terrestrial vertebrates in SW Europe. Acta Oecologica 2011, 37, 399–412. [Google Scholar] [CrossRef]
- Flojgaard, C.; Normand, S.; Skov, F.; Svenning, J.-C. Deconstructing the mammal species richness pattern in Europe—Towards an understanding of the relative importance of climate, biogeographic history, habitat heterogeneity and humans. Glob. Ecol. Biogeogr. 2011, 20, 218–230. [Google Scholar] [CrossRef]
- Baquero, R.A.; Telleria, J.L. Species richness, rarity and endemicity of European mammals: A biogeographical approach. Biodivers. Conserv. 2001, 10, 29–44. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Pausas, J.G. Does plant richness influence animal richness? the mammals of Catalonia (NE Spain). Divers. Distrib. 2004, 10, 247–252. [Google Scholar] [CrossRef]
- Sans-Fuentes, M.A.; Ventura, J. Distribution patterns of the small mammals (Insectivora and Rodentia) in a transitional zone between the Eurosiberian and the Mediterranean regions. J. Biogeogr. 2000, 27, 755–764. [Google Scholar] [CrossRef]
- Davidson, A.D.; Detling, J.K.; Brown, J.H. Ecological roles and conservation challenges of social, burrowing, herbivorous mammals in the world’s grasslands. Front. Ecol. Environ. 2012, 10, 477–486. [Google Scholar] [CrossRef]
- Luque-Larena, J.J.; Mougeot, F.; Viñuela, J.; Jareño, D.; Arroyo, L.; Lambin, X.; Arroyo, B. Recent large-scale range expansion and outbreaks of the common vole (Microtus Arvalis) in NW Spain. Basic Appl. Ecol. 2013, 14, 432–441. [Google Scholar] [CrossRef]
- Araújo, M.B.; Guilhaumon, F.; Rodrigues Neto, D.; Pozo Ortego, I.; Calmaestra, R.G. Impactos, Vulnerabilidad y Adaptación al Cambio Climático de la Biodiversidad Española 2. Fauna de Vertebrados; Dirección General de Medio Natural y Política Forestal, Ministerio de Medio Ambiente, y Medio Rural y Marino: Madrid, Spain, 2011. [Google Scholar]
- Roura-Pascual, N.; Pons, P.; Etienne, M.; Lambert, B. Transformation of a rural landscape in the Eastern Pyrenees between 1953 and 2000. Mt. Res. Dev. 2005, 25, 252–261. [Google Scholar] [CrossRef]
- Taüll, M. Valoració i Diagnosi Pastoral del Parc Natural de les Capçaleres del Ter i del Freser; Ciència i Tecnologia Forestal a Catalunya (CTFC): Lleida, Spain, 2017; pp. 1–110. [Google Scholar]
- Aldezabal, A.; García-González, R.; Gómez, D.; Fillat, F. El papel de los herbívoros en la conservación de los pastos. Ecosistemas 2002, 11, 1–9. [Google Scholar]
- Minuartia. Directrius per a la Recuperació D’espais Oberts en el Parc Natural de les Capçaleres del Ter i del Freser; Minuartia: Barcelona, Spain, 2019. [Google Scholar]
- Álvarez-Martínez, J.; Gómez-Villar, A.; Lasanta, T. The Use of Goats Grazing to Restore Pastures Invaded by Shrubs and Avoid Desertification: A Preliminary Case Study in the Spanish Cantabrian Mountains. Land Degrad. Dev. 2016, 27, 3–13. [Google Scholar] [CrossRef]
- Alados, C.L.; Saiz, H.; Nuche, P.; Gartzia, M.; Komac, B.; Frutos, D.E.; Pueyo, Y. Clearing vs. burning for restoring Pyrenean grasslands after shrub encroachment. Cuad. Investig. Geográfica 2019, 45, 441–468. [Google Scholar] [CrossRef]
- Torre, I.; Freixas, L.; Arrizabalaga, A.; Díaz, M. The efficiency of two widely used commercial live-traps to develop monitoring protocols for small mammal biodiversity. Ecol. Indic. 2016, 66, 481–487. [Google Scholar] [CrossRef]
- Machtinger, E.T.; Williams, S.C. Practical guide to trapping Peromyscus leucopus (Rodentia: Cricetidae) and Peromyscus maniculatus for vector and vector-borne pathogen surveillance and ecology. J. Insect Sci. 2020, 20, 5. [Google Scholar] [CrossRef]
- Gurnell, J.; Flowerdew, J.R. Live Trapping Small Mammals: A Practical Guide; Mammal Society: Southampton, UK, 2006; ISBN 9780906282540. [Google Scholar]
- Sikes, R.S. Animal Care and use Committee of the American Society of Guidelines of the American Society of Mammalogists for the use of wild mammals in research. J. Mammal. 2016, 97, 663–688. [Google Scholar] [CrossRef]
- Wilson, D.E.; Lacher, T.E.; Mittermeier, R.A. Handbook of the Mammals of the World, Volume 7: Rodents II; Lynx: Barcelona, Spain, 2017; Volume 1008. [Google Scholar]
- Zuur, A.F.; Leno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer Science+Business Media: Berlin/Heidelberg, Germany, 2009; pp. 1–574. ISBN 9780387874579. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Tarjuelo, R.; Morales, M.B.; Traba, J. Breadth and specialization in microhabitat selection: The case of the algerian mouse (Mus spretus) in central Spain. Rev. d’Ecologie (La Terre la Vie) 2011, 66, 145–156. [Google Scholar] [CrossRef]
- Torre, I.; Díaz, M.; Martínez-Padilla, J.; Bonal, R.; Viñuela, J.; Fargallo, J.A. Cattle grazing, raptor abundance and small mammal communities in Mediterranean grasslands. Basic Appl. Ecol. 2007, 8, 565–575. [Google Scholar] [CrossRef]
- Harrison, X.A.; Donaldson, L.; Correa-Cano, M.E.; Evans, J.; Fisher, D.N.; Goodwin, C.E.D.; Robinson, B.S.; Hodgson, D.J.; Inger, R. A brief introduction to mixed effects modelling and multi-model inference in ecology. PeerJ 2018, 6, e4794. [Google Scholar] [CrossRef]
- Komac, B.; Kefi, S.; Nuche, P.; Escós, J.; Alados, C.L. Modeling shrub encroachment in subalpine grasslands under different environmental and management scenarios. J. Environ. Manage. 2013, 121, 160–169. [Google Scholar] [CrossRef]
- Casasús, I.; Bernués, A.; Sanz, A.; Villalba, D.; Riedel, J.L.; Revilla, R. Vegetation dynamics in Mediterranean forest pastures as affected by beef cattle grazing. Agric. Ecosyst. Environ. 2007, 121, 365–370. [Google Scholar] [CrossRef]
- Sebastià, M.T.; De Bello, F.; Puig, L.; Taull, M. Grazing as a factor structuring grasslands in the Pyrenees. Appl. Veg. Sci. 2008, 11, 215–222. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Colwell, R.K. Quantifying biodiversity: Procedures and pitfalls in the measurement and comparison of species richness. Ecol. Lett. 2001, 4, 379–391. [Google Scholar] [CrossRef]
- Jacob, J.; Manson, P.; Barfknecht, R.; Fredricks, T. Common vole (Microtus arvalis) ecology and management: Implications for risk assessment of plant protection products. Pest Manag. Sci. 2014, 70, 869–878. [Google Scholar] [CrossRef] [PubMed]
- Torre, I.; Jaime-González, C.; Díaz, M. Habitat Suitability for Small Mammals in Mediterranean Landscapes: How and Why Shrubs Matter. Sustainability 2022, 14, 1562. [Google Scholar] [CrossRef]
- Stefanescu, C.; Torre, I.; Jubany, J.; Paramo, F. Recent trends in butterfly populations from north-east Spain and Andorra in the light of habitat and climate change. J. Insect Conserv. 2011, 15, 83–93. [Google Scholar] [CrossRef]
- Borghi, C.E.; Giannoni, S.M.; Martinez-Rica, J.P. Habitat segregation of three sympatric fossorial rodents in the Spanish Pyrenees. Z. Fuer Saugetierkd. 1994, 59, 52–57. [Google Scholar]
- Lambin, X.; MacKinnon, J. The relative efficiency of two commercial live-traps for small mammals. J. Zool. 1997, 242, 400–404. [Google Scholar] [CrossRef]
- Fargallo, J.A.; Martinez-Padilla, J.; Vinuela, J.; Blanco, G.; Torre, I.; Vergara, P.; De Neve, L. Kestrel-Prey Dynamic in a Mediterranean Region: The Effect of Generalist Predation and Climatic Factors. PLoS ONE 2009, 4, e4311. [Google Scholar] [CrossRef]
- Torre, I.; Díaz, M. Assessing the Effects of Landscape Change on the Occupancy Dynamics of the Greater White-Toothed Shrew Crocidura russula. Life 2022, 12, 1230. [Google Scholar] [CrossRef]
- Torre, I.; Bastardas-Llabot, J.; Arrizabalaga, A.; Díaz, M. Population dynamics of small endotherms under global change: Greater white-toothed shrews Crocidura russula in Mediterranean habitats. Sci. Total Environ. 2020, 705, 135799. [Google Scholar] [CrossRef]
- Paliga, R. Landscapes of Stone: How the Landscape Affects Small Mammal Use of Dry Stone Walls; University of Guelph: Guelph, Canada, 2011. [Google Scholar]
- Torre, I.; Raspall, A.; Arrizabalaga, A.; Díaz, M. SEMICE: An unbiased and powerful monitoring protocol for small mammals in the Mediterranean Region. Mamm. Biol. 2018, 88, 161–167. [Google Scholar] [CrossRef]
- García, F.J.; Díaz, M.; De Alba, J.M.; Alonso, C.L.; Carbonell, R.; De Carrión, M.L.; Monedero, C.; Santos, T. Edge effects and patterns of winter abundance of wood mice Apodemus sylvaticus in Spanish fragmented forests. Acta Theriol. 1998, 43, 255–262. [Google Scholar] [CrossRef]
- Sunyer, P.; Muñoz, A.; Mazzerole, M.J.; Bonal, R.; Espelta, J.M. Wood mouse population dynamics: Interplay among seed abundance seasonality, shrub cover and wild boar interference. Mamm. Biol. 2016, 81, 372–379. [Google Scholar] [CrossRef]
- Stefanescu, C.; Soldevila, A.; Gutiérrez, C.; Torre, I.; Ubach, A.; Miralles, M. Explosions demogràfiques de l’eruga peluda del suro, Lymantria dispar (Linnaeus, 1758), als boscos del Montnegre el 2019 i 2020: Possibles causes, impactes i idoneïtat dels tractaments per combatre la plaga. Butll. Inst. Catalana Història Nat. 2020, 84, 267–279. [Google Scholar]
- Oro, D.; Sanz-Aguilar, A.; Carbonell, F.; Grajera, J.; Torre, I. Multi-species prey dynamics influences local survival in resident and wintering generalist predators. Oecologia 2021, 197, 437–446. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Herbaceous Cover (%) | Shrub Cover (<1.5 m, %) | Shrub Cover (>1.5 m, %) | |||||
|---|---|---|---|---|---|---|---|
| Estimate | SE | Estimate | SE | Estimate | SE | ||
| Treatment | (Intercept) | −0.67 *** | 0.13 | 0.19 | 0.13 | −0.12 | 0.14 |
| Post-1 | 0.97 *** | 0.10 | −0.71 *** | 0.13 | 0.05 | 0.13 | |
| Post-2 | 1.02 *** | 0.10 | 0.14 | 0.13 | 0.31 * | 0.13 | |
| AICc | 496.73 | 565.69 | 566.18 | ||||
| R2 marginal | 0.22 | 0.14 | 0.02 | ||||
| R2 conditional | 0.61 | 0.40 | 0.44 | ||||
| A. sylvaticus | C. russula | M. arvalis | Total | Species | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Estimate | SE | Estimate | SE | Estimate | SE | Estimate | SE | Estimate | SE | ||
| (Intercept) | 0.95 *** | 0.19 | −0.05 | 0.08 | −0.08 | 0.08 | 0.85 *** | 0.22 | 0.43 ** | 0.15 | |
| Treatment | Post-1 | 0.11 | 0.08 | 0.02 | 0.04 | 0.07 * | 0.03 | 0.23 * | 0.09 | 0.15 * | 0.06 |
| Post-2 | −0.04 | 0.08 | −0.05 | 0.04 | −0.01 | 0.03 | −0.10 | 0.09 | −0.08 | 0.06 | |
| Vegetation | Herb | −0.08 ** | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | −0.01 | 0.00 | 0.00 | 0.00 |
| Shrub (<1.5 m) | 0.00 | 0.00 | 0.00 ** | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Shrub (>1.5 m) | 0.00 *** | 0.00 | 0.00 ** | 0.00 | 0.00 | 0.00 | 0.01 *** | 0.00 | 0.01 *** | 0.00 | |
| Stone wall | −0.01 | 0.01 | −0.01 * | 0.00 | 0.01 | 0.00 | −0.01 | 0.01 | −0.01 | 0.01 | |
| Season | Summer | −0.10 | 0.05 | −0.04 | 0.02 | −0.02 | 0.02 | −0.18 ** | 0.06 | −0.10 * | 0.04 |
| AICc | 285.92 | −37.15 | −95.42 | 354.97 | 184.03 | ||||||
| R2 marginal | 0.22 | 0.08 | 0.07 | 0.21 | 0.22 | ||||||
| R2 conditional | 0.30 | 0.10 | 0.12 | 0.28 | 0.32 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torre, I.; Palau, O. Is Shrub Encroachment Driving the Decline of Small Mammal Diversity in Pyrenean Grasslands? A Preliminary Study. Diversity 2023, 15, 232. https://doi.org/10.3390/d15020232
Torre I, Palau O. Is Shrub Encroachment Driving the Decline of Small Mammal Diversity in Pyrenean Grasslands? A Preliminary Study. Diversity. 2023; 15(2):232. https://doi.org/10.3390/d15020232
Chicago/Turabian StyleTorre, Ignasi, and Oriol Palau. 2023. "Is Shrub Encroachment Driving the Decline of Small Mammal Diversity in Pyrenean Grasslands? A Preliminary Study" Diversity 15, no. 2: 232. https://doi.org/10.3390/d15020232
APA StyleTorre, I., & Palau, O. (2023). Is Shrub Encroachment Driving the Decline of Small Mammal Diversity in Pyrenean Grasslands? A Preliminary Study. Diversity, 15(2), 232. https://doi.org/10.3390/d15020232