
Floristic Homogenization and Differentiation under Deer Grazing in a Subalpine Zone in Central Japan


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design
2.3. Study of Plant Diversity
2.4. Data Analysis
3. Results
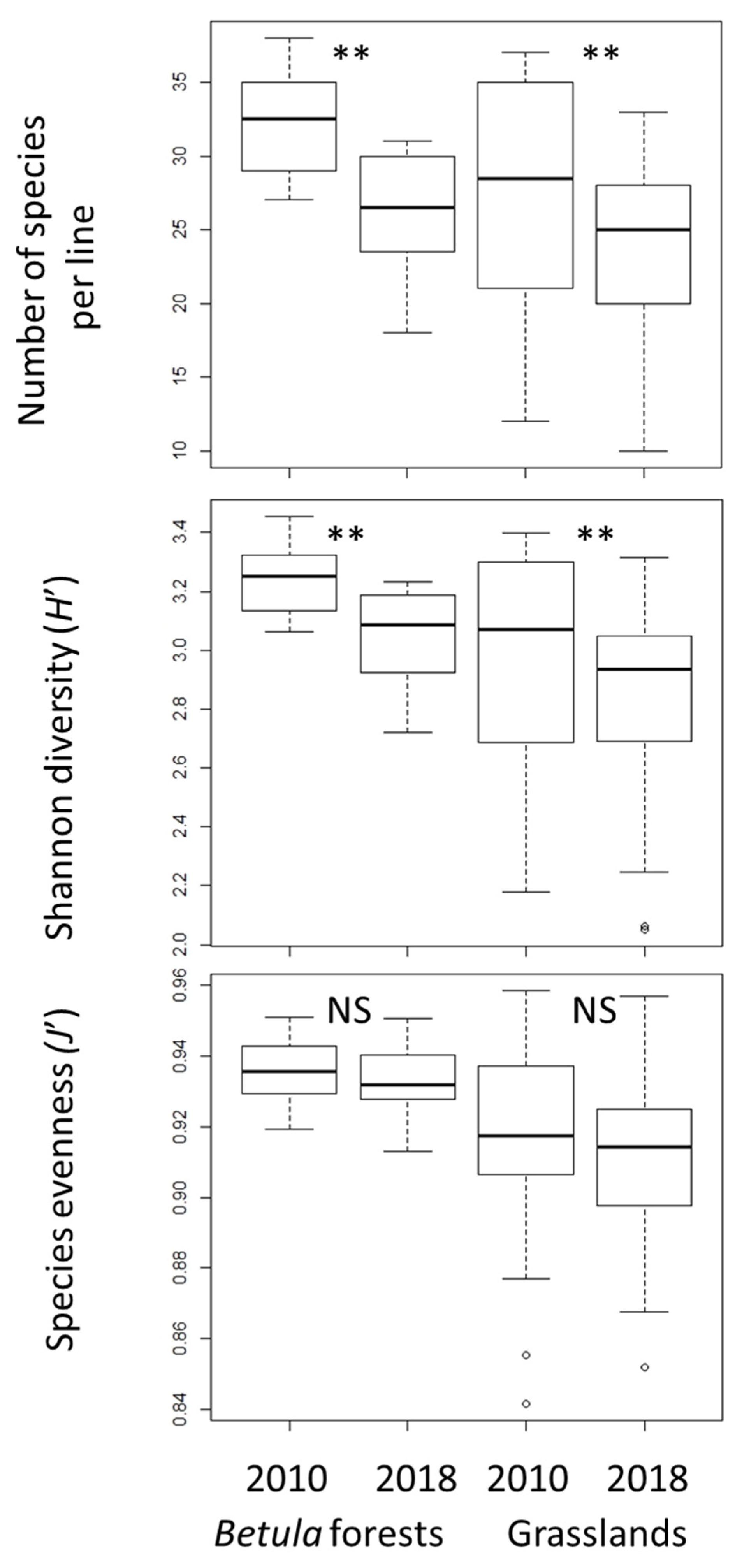
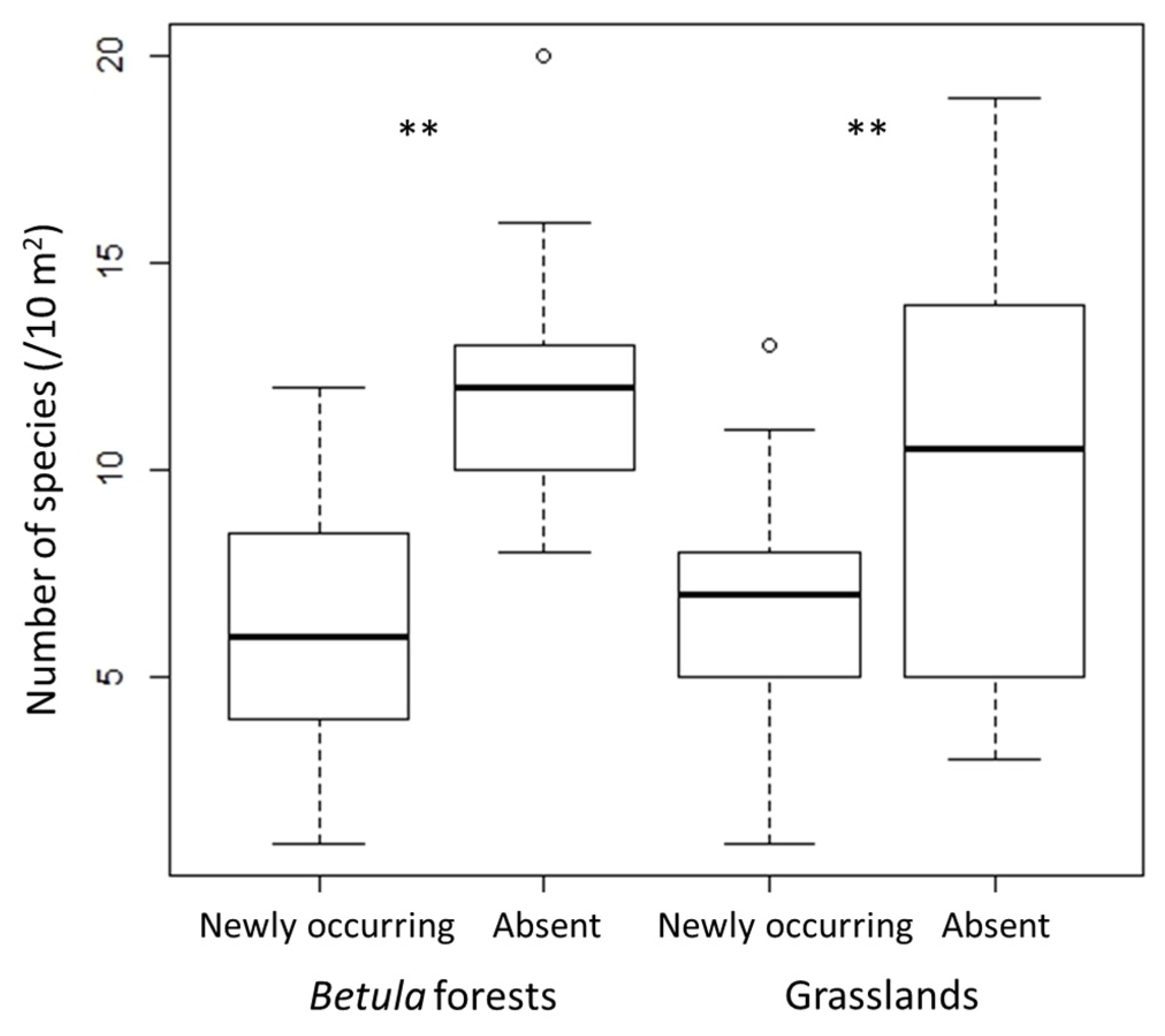
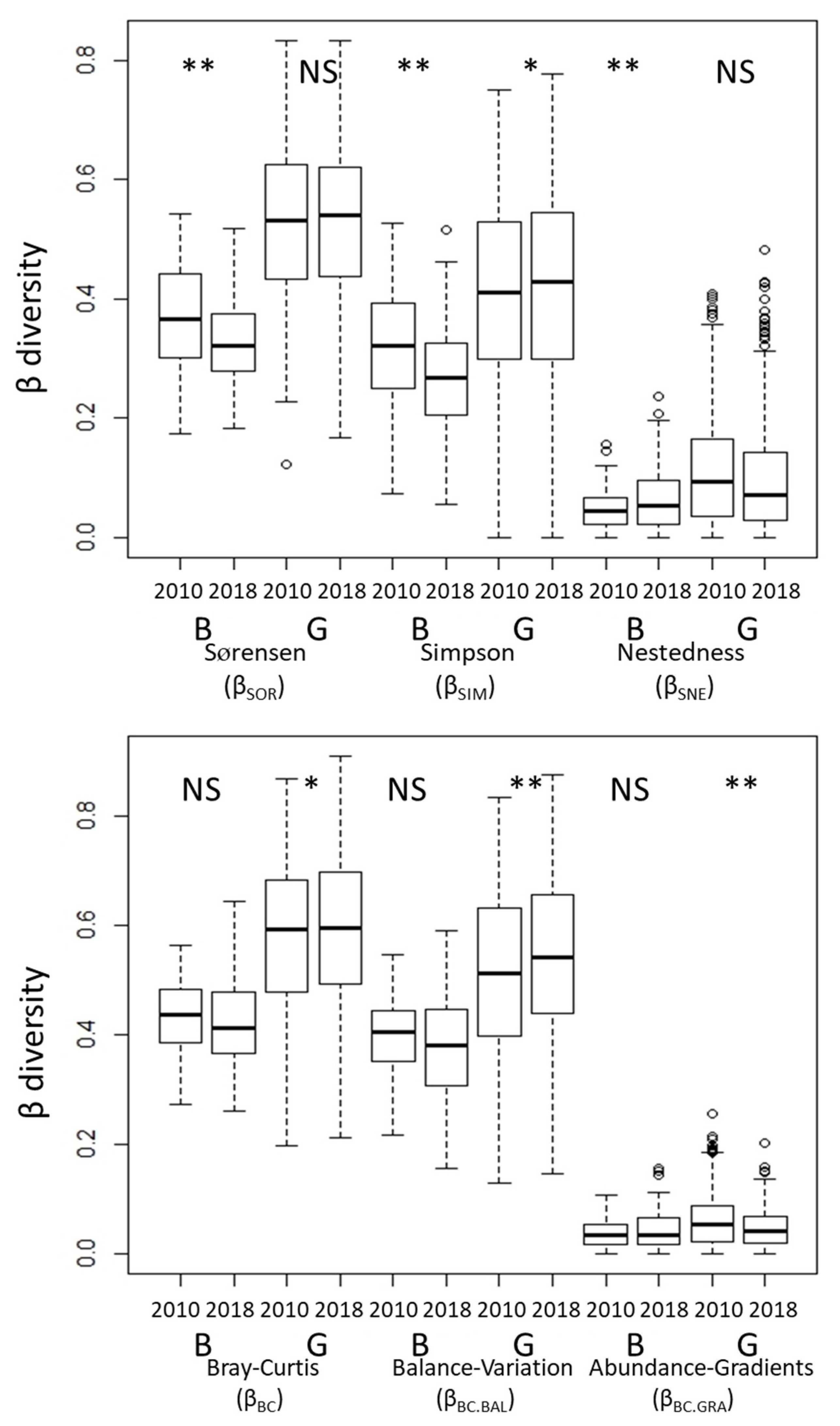
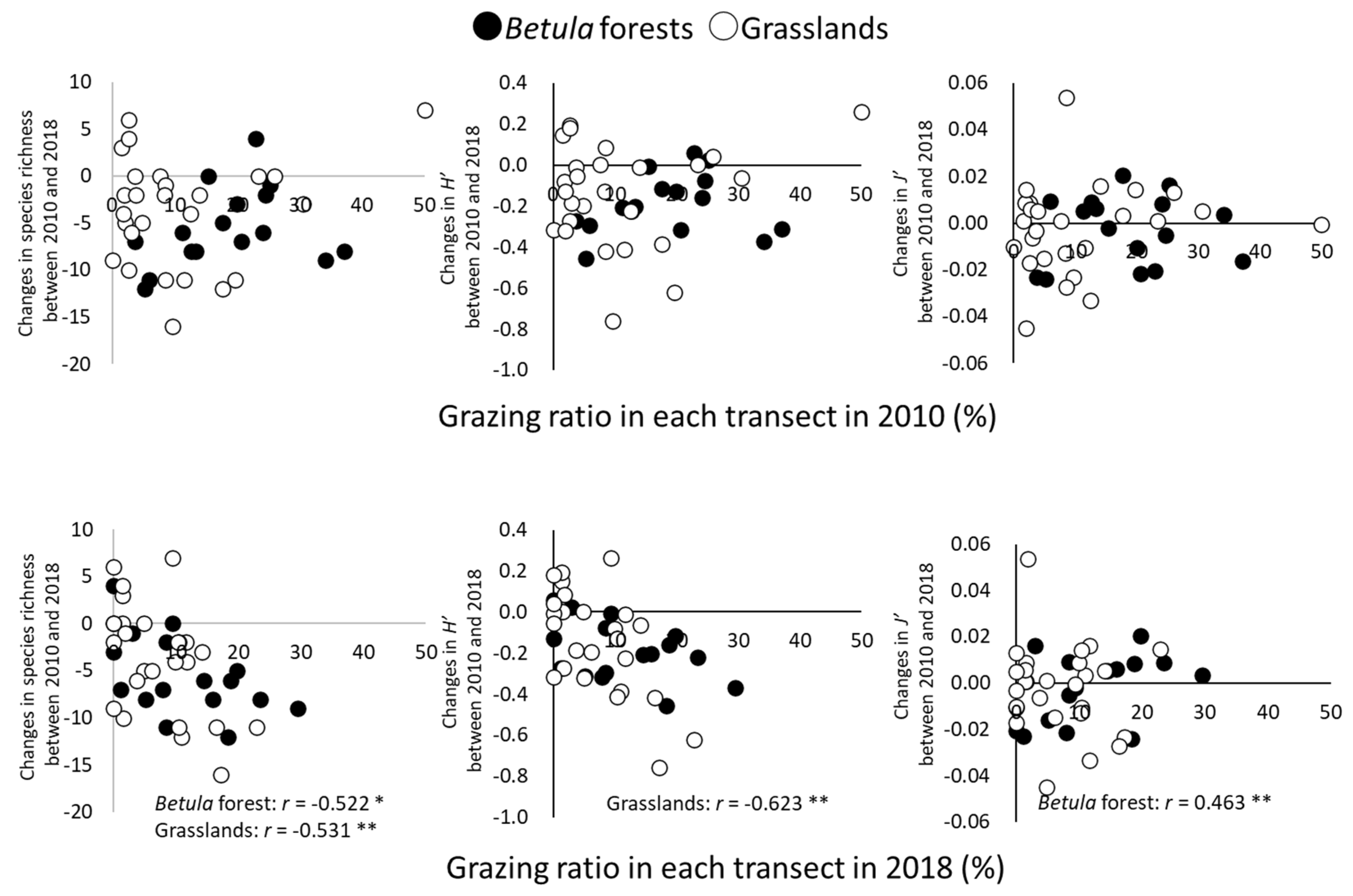
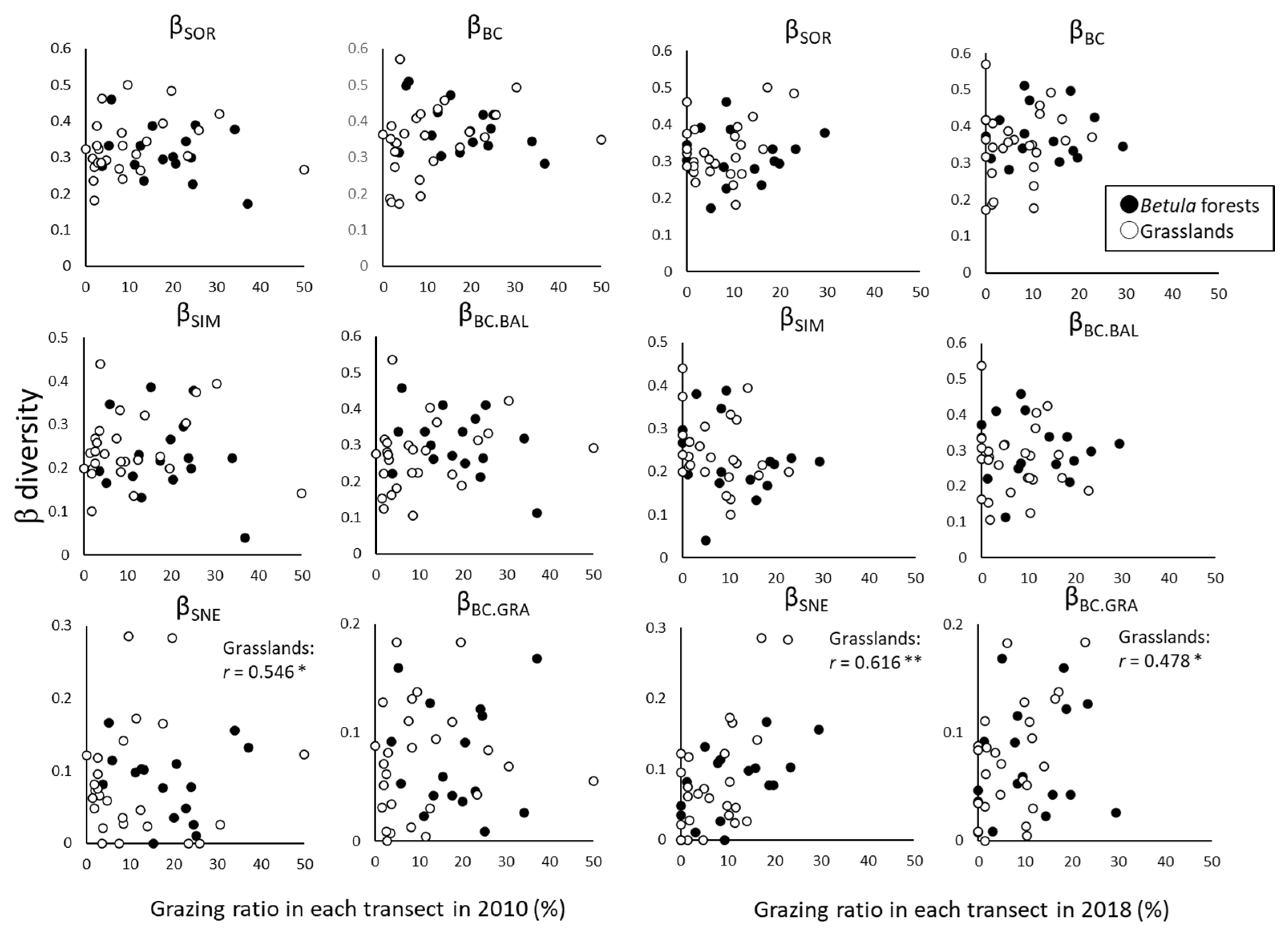
3.1. Changes in α- and β-Diversity
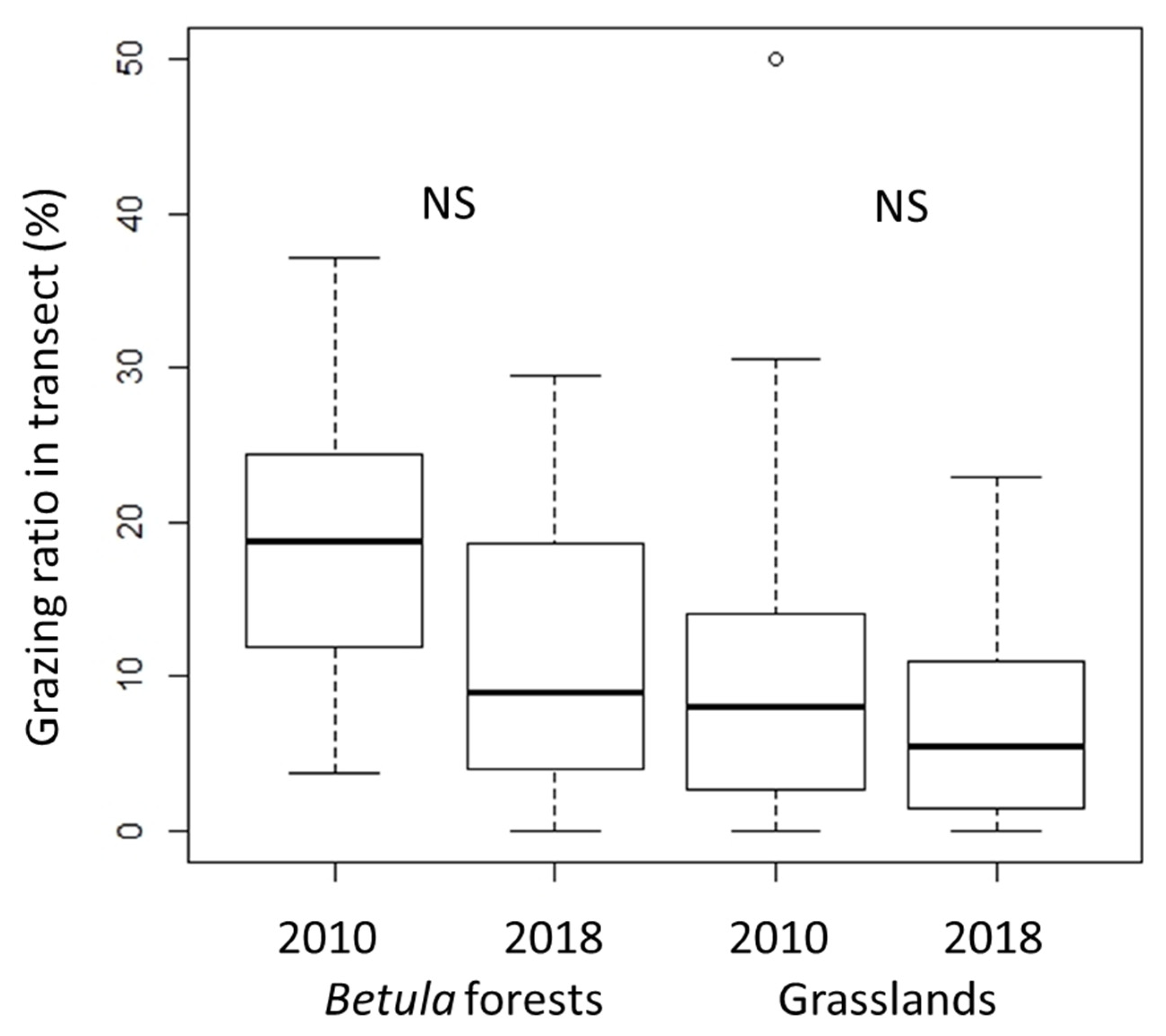
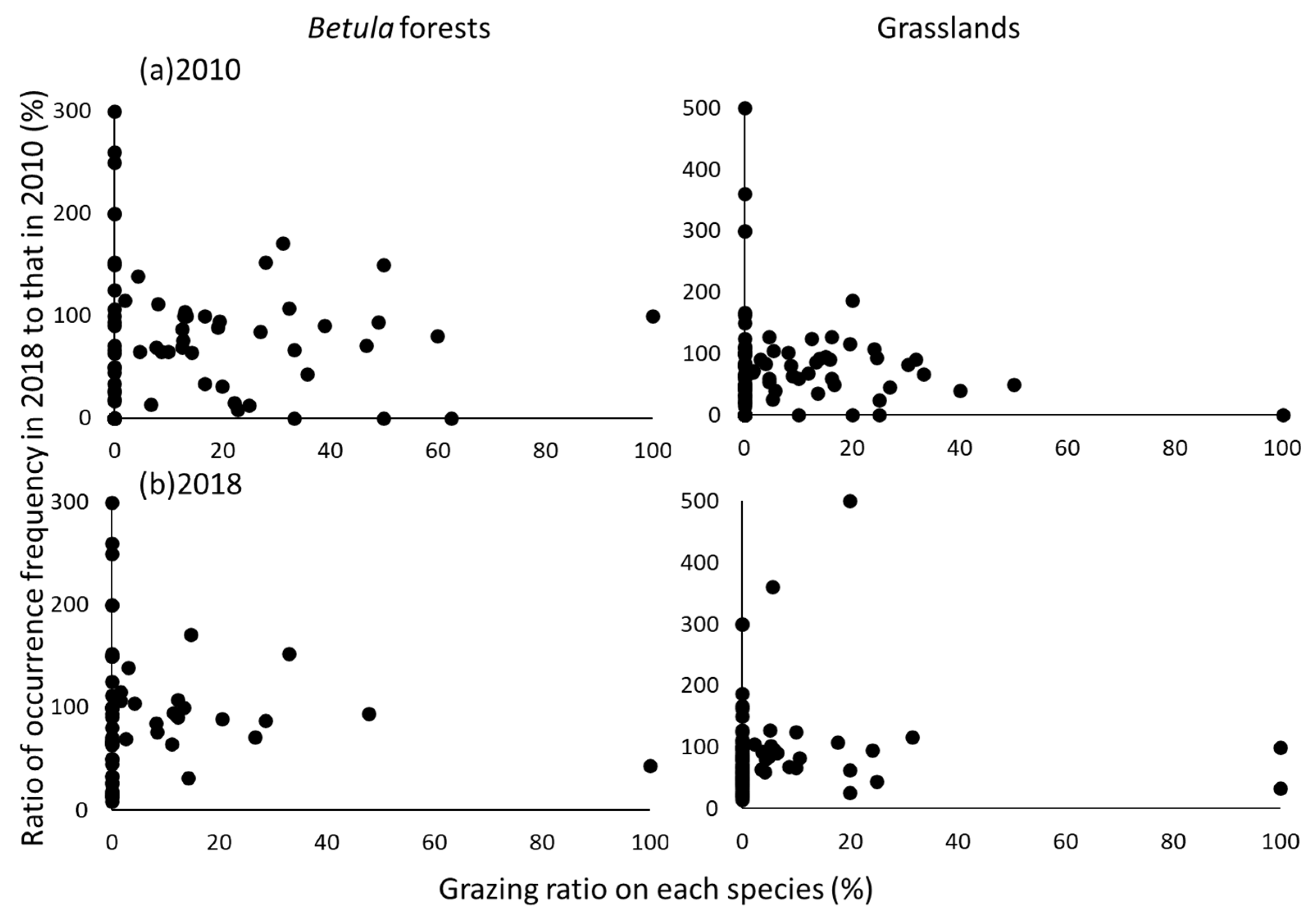
3.2. Grazing by Sika Deer
4. Discussion
4.1. Changes in α- and β-Diversity
4.2. Effects of Grazing on α- and β-Diversity
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Speed, J.D.M.; Austrheim, G.; Mysterud, A. The response of plant diversity to grazing varies along an elevational gradient. J. Ecol. 2013, 101, 1225–1236. [Google Scholar] [CrossRef]
- Li, C.; Hao, X.; Zhao, M.; Han, G.; Willms, W.D. Influence of historic sheep grazing on vegetation and soil properties of a desert steppe in inner Mongolia. Agric. Ecosyst. Environ. 2008, 128, 109–116. [Google Scholar] [CrossRef]
- Sasaki, T.; Okayasu, T.; Jamsran, U.; Takeuchi, K. Threshold changes in vegetation along a grazing gradient in Mongolian rangelands. J. Ecol. 2008, 96, 145–154. [Google Scholar] [CrossRef]
- Holmes, S.A.; Webster, C.R. Herbivore-induced expansion of generalist species as a driver of homogenization in post-disturbance plant communities. Plant Ecol. 2011, 212, 753–768. [Google Scholar] [CrossRef]
- Oldén, A.; Halme, P. Grazers increase β-diversity of vascular plants and bryophytes in wood pastures. J. Veg. Sci. 2016, 27, 1084–1093. [Google Scholar] [CrossRef]
- Perotti, E.; Probo, M.; Pittarello, M.; Lonati, M.; Lombardi, G. A 5-year rotational grazing changes the botanical composition of sub-alpine and alpine grasslands. App. Veg. Sci. 2018, 21, 647–657. [Google Scholar] [CrossRef]
- Suding, K.N.; Hobbs, R.J. Threshold models in restoration and conservation: A developing framework. Trends Ecol. Evol. 2009, 24, 271–279. [Google Scholar] [CrossRef]
- Rooney, T.P. High white-tailed deer densities benefit graminoids and contribute to biotic homogenization of forest ground-layer vegetation. Plant Ecol. 2009, 202, 103–111. [Google Scholar] [CrossRef]
- Speed, J.D.M.; Cooper, E.J.; Jónsdóttir, I.S.; Van Der Wal, R.; Woodin, S.J. Plant community properties predict vegetation resilience to herbivore disturbance in the Arctic. J. Ecol. 2010, 98, 1002–1013. [Google Scholar] [CrossRef]
- Nagaike, T.; Ohkubo, E.; Hirose, K. Vegetation recovery in response to the exclusion of grazing by sika deer (Cervus nippon) in seminatural grassland on Mt. Kushigata, Japan. ISRN Biodivers. 2014, 2014, 493495. [Google Scholar] [CrossRef]
- Pendergast, T.H.; Hanlon, S.M.; Long, Z.M.; Royo, A.A.; Carson, W.P. The legacy of deer overabundance: Long-term delays in herbaceous understory recovery. Can. J. For. Res. 2016, 46, 362–369. [Google Scholar] [CrossRef]
- Olden, J.D.; Rooney, T.P. On defining and quantifying biotic homogenization. Glob. Ecol. Biogeogr. 2006, 15, 113–120. [Google Scholar] [CrossRef]
- Ross, L.C.; Woodin, S.J.; Hester, A.J.; Thompson, D.B.A.; Birks, J.B. Biotic homogenization of upland vegetation: Patterns and drivers at multiple spatial scales over five decades. J. Veg. Sci. 2012, 23, 755–770. [Google Scholar] [CrossRef]
- Ohashi, H.; Hoshino, Y. Disturbance by large herbivores alters the relative importance of the ecological processes that influence the assembly pattern in heterogeneous meta-community. Ecol. Evol. 2014, 4, 766–775. [Google Scholar] [CrossRef]
- Nascimbene, J.; Benesperi, R.; Brunialti, G.; Catalano, I.; Vedove, M.D.; Grillo, M.; Isocrono, D.; Matteucci, E.; Potenza, G.; Puntillo, D.; et al. Patterns and derivers of ꞵ-diversity and similarity of Lobaria pulmonaria communities in Italian forests. J. Ecol. 2013, 101, 493–505. [Google Scholar] [CrossRef]
- Prach, J.; Kopecký, M. Landscape-scale vegetation homogenization in central European sub-montane forests over the past 50 years. App. Veg. Sci. 2018, 21, 373–384. [Google Scholar] [CrossRef]
- Johnson, S.E.; Mudrak, E.L.; Waller, D.M. Local increases in diversity accompany community homogenization in floodplain forest understories. J. Veg. Sci. 2014, 25, 885–896. [Google Scholar] [CrossRef]
- Trentanovi, G.; Von der Lippe, M.; Sitzia, T.; Ziechmann, U.; Kowarik, I.; Cierjacks, A. Biotic homogenization at the community scale: Disentangling the roles of urbanization and plant invasion. Div. Dis. 2013, 19, 738–748. [Google Scholar] [CrossRef]
- Lososová, Z.; Chytrý, M.; Danihelka, J.; Tichý, L.; Ricotta, C. Biotic homogenization of urban floras by alien species: The role of species turnover and richness differences. J. Veg. Sci. 2016, 27, 452–459. [Google Scholar] [CrossRef]
- McCune, J.L.; Vellend, M. Gains in native species promote biotic homogenization over four decades in a human-dominated landscape. J. Ecol. 2013, 101, 1542–1551. [Google Scholar] [CrossRef]
- Peper, J.; Jansen, F.; Pietzsch, D.; Manthey, M. Patterns of plant species turnover along grazing gradients. J. Veg. Sci. 2011, 22, 457–466. [Google Scholar] [CrossRef]
- Naaf, T.; Wulf, M. Habitat specialists and generalists drive homogenization and differentiation of temperate forest plant communities at the regional scale. Biol. Conserv. 2010, 143, 848–855. [Google Scholar] [CrossRef]
- Bernhardt-Römermann, M.; Baeten, L.; Craven, D.; De Frenne, P.; Hédl, R.; Lenoir, J.; Bert, D.; Brunet, J.; Chudomelová, M.; Decocq, G.; et al. Drivers of temporal changes in temperate forest plant diversity vary across spatial scales. Glob. Chang. Biol. 2015, 21, 3726–3737. [Google Scholar] [CrossRef] [PubMed]
- Chase, J.M.; Myers, J.A. Disentangling the importance of ecological niches from stochastic processes across scales. Phil. Trans. Royal Soc. B 2011, 366, 2351–2363. [Google Scholar] [CrossRef]
- Alberti, J.; Bakker, E.S.; Van Klink, R.; Olff, H.; Smit, C. Herbivore exclusion promotes a more stochastic plant community assembly in a natural grassland. Ecology 2017, 98, 961–970. [Google Scholar] [CrossRef]
- Måren, I.E.; Kapfer, J.; Aarrestad, P.A.; Grytnes, J.A.; Vandvik, V. Changing contributions of stochastic and deterministic processes in community assembly over a successional gradient. Ecology 2018, 99, 148–157. [Google Scholar] [CrossRef]
- Nishizawa, K.; Tatsumi, S.; Kitagawa, R.; Mori, A.S. Deer herbivory affects the functional diversity of forest floor plants via changes in competition-mediated assembly rules. Ecol. Res. 2016, 31, 569–578. [Google Scholar] [CrossRef]
- Takatsuki, S. Effects of sika deer on vegetation in Japan: A review. Biol. Conserv. 2009, 142, 1922–1929. [Google Scholar] [CrossRef]
- Iijima, H.; Nagaike, T.; Honda, T. Estimation of deer population dynamics using a Bayesian state–space model with multiple abundance indices. J. Wildl. Manag. 2013, 77, 1038–1047. [Google Scholar] [CrossRef]
- Nagaike, T.; Hayashi, A. Bark-stripping by sika deer (Cervus nippon) in Larix kaempferi plantations in central Japan. For. Ecol. Manag. 2003, 175, 563–572. [Google Scholar] [CrossRef]
- Nagaike, T. Effects of heavy, repeated bark stripping by Cervus nippon on survival of Abies veitchii in a subalpine coniferous forest in central Japan. J. For. Res. 2020, 39, 1139–1145. [Google Scholar] [CrossRef]
- Nagaike, T. Effects of browsing by sika deer (Cervus nippon) on subalpine vegetation at Mt. Kita, central Japan. Ecol. Res. 2012, 27, 467–473. [Google Scholar] [CrossRef]
- Kamei, T.; Takeda, K.; Izumiyama, S.; Ohshima, K. The effect of hunting on the behavior and habitat utilization of sika deer (Cervus nippon). Mammal Study 2010, 35, 235–241. [Google Scholar] [CrossRef]
- Takii, A.; Izumiyama, S.; Taguchi, M. Partial migration and effects of climate on migratory movements of sika deer in Kirigamine Highland, central Japan. Mammal Study 2012, 37, 331–340. [Google Scholar] [CrossRef]
- Otsu, C.; Iijima, H.; Nagaike, T.; Hoshino, Y. Evidence of extinction debt through the survival and colonization of each species in semi-natural grasslands. J. Veg. Sci. 2017, 28, 464–474. [Google Scholar] [CrossRef]
- Koyama, A.; Uchida, K.; Ozeki, M.; Iwasaki, T.; Nakahama, N.; Suka, T. Conservation of endanger and rare plants requires strategies additional to deer-proof fencing for conservation of sub-alpine plant diversity. Appl. Veg. Sci. 2021, 24, e12553. [Google Scholar] [CrossRef]
- Ishida, Y.; Takyu, M.; Nakamura, Y. Species composition and distribution pattern of Betula ermanii forests on Mt. Kitadake, Japanese Southern Alps. Veg. Sci. 2014, 31, 129–142, (In Japanese with English Summary). [Google Scholar]
- Mizuno, K. Habitat types of Ohanabatake vegetation in the Akaishi mountains, central Japan. Geogr. Rev. Jpn. 1984, 6, 384–402, (In Japanese with English Summary). [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring Biological Diversity; Blackwell: Oxford, UK, 2004. [Google Scholar]
- Baselga, A. Separating the two components of abundance-based dissimilarity: Balanced changes in abundance vs. abundance gradients. Methods Ecol. Evol. 2013, 4, 552–557. [Google Scholar] [CrossRef]
- Baselga, A. Partitioning abundance-based multiple-site dissimilarity into components: Balanced variation in abundance and abundance gradients. Methods Ecol. Evol. 2017, 8, 799–808. [Google Scholar] [CrossRef]
- Roberts, M.R. Effects of forest plantation management on herbaceous-layer composition and diversity. Can. J. Bot. 2002, 80, 378–389. [Google Scholar] [CrossRef]
- Peterson, E.B.; McCune, B. Diversity and succession of epiphytic macrolichen communities in low-elevation managed conifer forests in Western Oregon. J. Veg. Sci. 2001, 12, 511–524. [Google Scholar] [CrossRef]
- De Cáceres, M.; Jansen, F.; Dell, N. Indicator Species Analysis. 2020. Available online: https://cran.r-project.org/web/packages/indicspecies/vignettes/indicspeciesTutorial.pdf (accessed on 4 February 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 1 December 2020).
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Aolymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package. Version 2.6-4. Available online: https://cran.r-project.org/web/packages/vegan/vegan.pdf (accessed on 1 December 2022).
- Hothorn, T. Exactranktests: Exact Distributions for Rank and Permutation Tests. 2019. Available online: https://cran.r-project.org/web/packages/exactRankTests/exactRankTests.html (accessed on 1 June 2020).
- Baselga, A.; Orme, C.D.L. betapart: An R package for the study of beta diversity. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]
- Yonekura, K.; Kajita, T. BG Plants: Japanese and Scientific Name Index (YList). 2003. Available online: http://ylist.info/ (accessed on 1 December 2018).
- Bühler, C.; Roth, T. Spread of common species results in local-scale floristic homogenization in grassland of Switzerland. Div. Dis. 2011, 17, 1089–1098. [Google Scholar] [CrossRef]
- Reinecke, J.; Klemm, G.; Heinken, T. Vegetation change and homogenization of species composition in temperate nutrient deficient Scots pine forests after 45 years. J. Veg. Sci. 2014, 25, 113–121. [Google Scholar] [CrossRef]
- Amici, V.; Rocchini, D.; Geri, F.; Bacaro, G.; Marcantonio, M.; Chiarucci, A. Effects of an afforestation process on plant species richness: A retrogressive analysis. Ecol. Comp. 2012, 9, 55–62. [Google Scholar] [CrossRef]
- Arroyo-Rodríguez, V.; Rös, M.; Escobar, F.; Melo, F.P.L.; Santos, B.A.; Tabarelli, M.; Chazdon, R. Plant beta-diversity in fragmented rain forests: Testing floristic homogenization and differentiation hypothesis. J. Ecol. 2013, 101, 1449–1458. [Google Scholar] [CrossRef]
- Baeten, L.; Vangansbeke, P.; Hermy, M.; Peterken, G.; Vanhuyse, K.; Verheyen, K. Distinguishing between turnover and nestedness in the quantification of biotic homogenization. Biodivers. Conserv. 2012, 21, 1339–1409. [Google Scholar] [CrossRef]
- Nuttle, T.; Ristau, T.E.; Royo, A.A. Long-term biological legacies of herbivore density in a landscape-scale experiment: Forest understoreys reflect past deer density treatments for at least 20 years. J. Ecol. 2014, 102, 221–228. [Google Scholar] [CrossRef]
- Suzuki, M.; Miyashita, T.; Kabaya, H.; Ochiai, K.; Asada, M.; Kikvidze, Z. Deer herbivory as an important driver of divergence of ground vegetation communities in temperate forests. Oikos 2013, 122, 104–110. [Google Scholar] [CrossRef]
- Lilleeng, M.S.; Hegland, S.J.; Rydgren, K.; Moe, S.R. Red deer mediate spatial and temporal plant heterogeneity in boreal forests. Ecol. Res. 2016, 31, 777–784. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Incidence Data | ||
|---|---|---|
| βSOR | Sørensen dissimilarity. The total difference in species composition between censuses in a transect and between two transects in each vegetation type | |
| βSIM | Simpson dissimilarity. Turnover | |
| βSNE | Nestedness-driven dissimilarity. Nestedness | |
| Abundance data | ||
| βBC | Bray–Curtis dissimilarity. The total difference in species composition | |
| βBC.BAL | Replacement and balanced variation in abundance. The individuals of some species in one transect or census are replaced by the same number of individuals of a different species in another transect or census; substitution | |
| βBC.GRA | Nestedness in abundance gradients. Some individuals are lost from one transect to the other or between census periods; subset | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagaike, T. Floristic Homogenization and Differentiation under Deer Grazing in a Subalpine Zone in Central Japan. Diversity 2023, 15, 192. https://doi.org/10.3390/d15020192
Nagaike T. Floristic Homogenization and Differentiation under Deer Grazing in a Subalpine Zone in Central Japan. Diversity. 2023; 15(2):192. https://doi.org/10.3390/d15020192
Chicago/Turabian StyleNagaike, Takuo. 2023. "Floristic Homogenization and Differentiation under Deer Grazing in a Subalpine Zone in Central Japan" Diversity 15, no. 2: 192. https://doi.org/10.3390/d15020192
APA StyleNagaike, T. (2023). Floristic Homogenization and Differentiation under Deer Grazing in a Subalpine Zone in Central Japan. Diversity, 15(2), 192. https://doi.org/10.3390/d15020192