


Species Diversity and Community Structure of Macrobenthos in the Cosmonaut Sea, East Antarctica

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
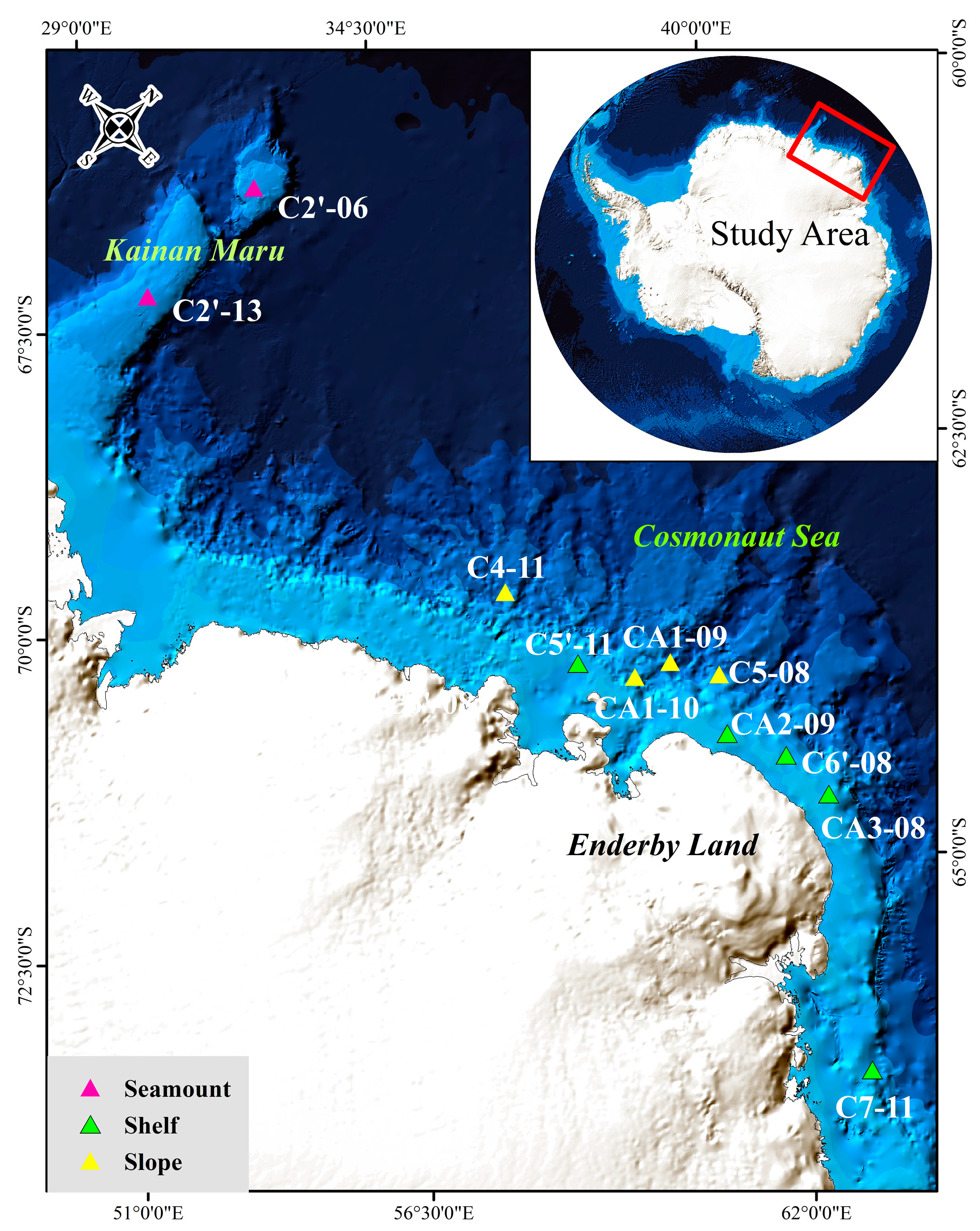
2.1. Study Area
2.2. Sample Processing
2.3. Data Processing
2.4. Classification of Macrobenthos Communities
3. Results
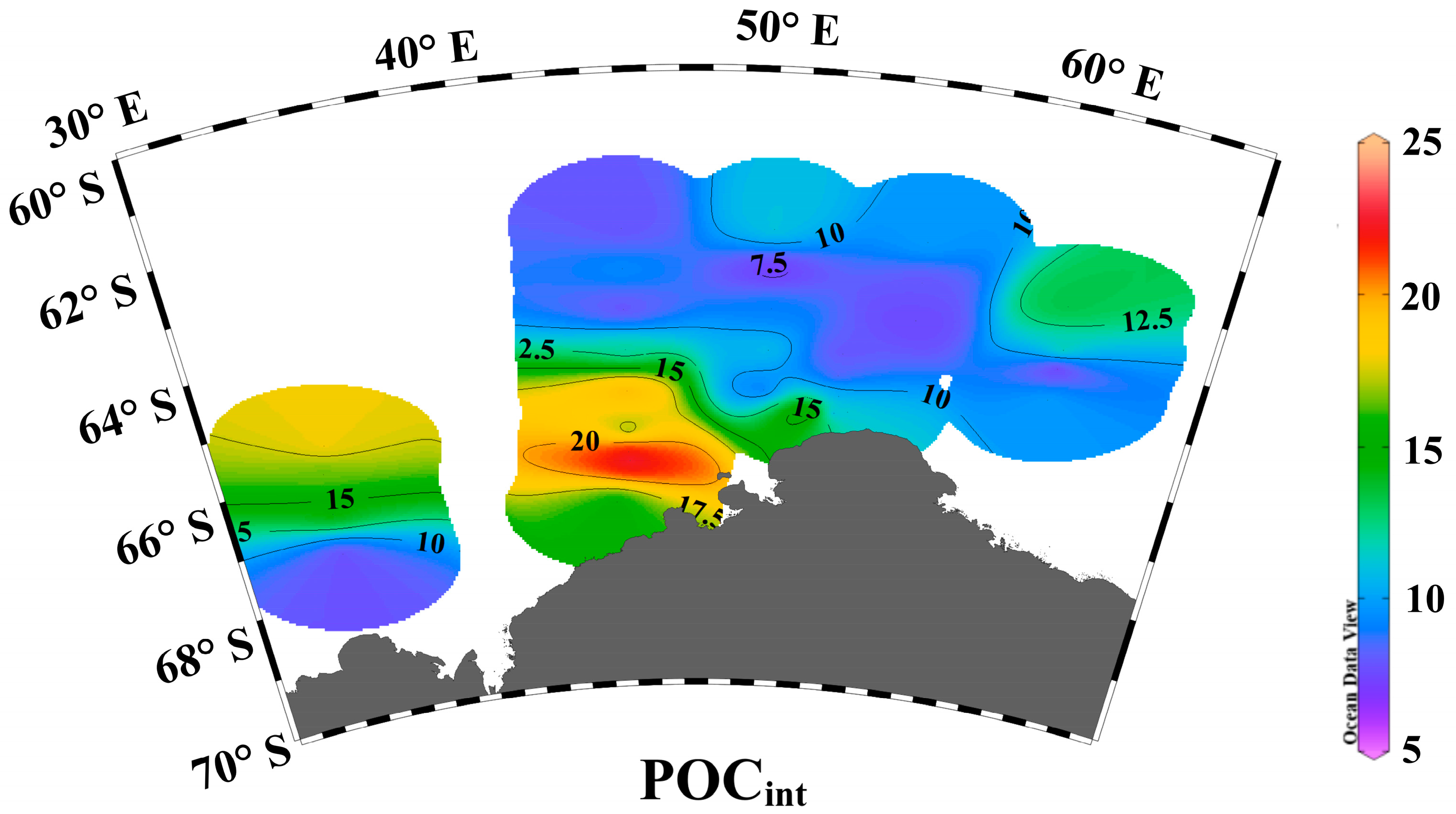
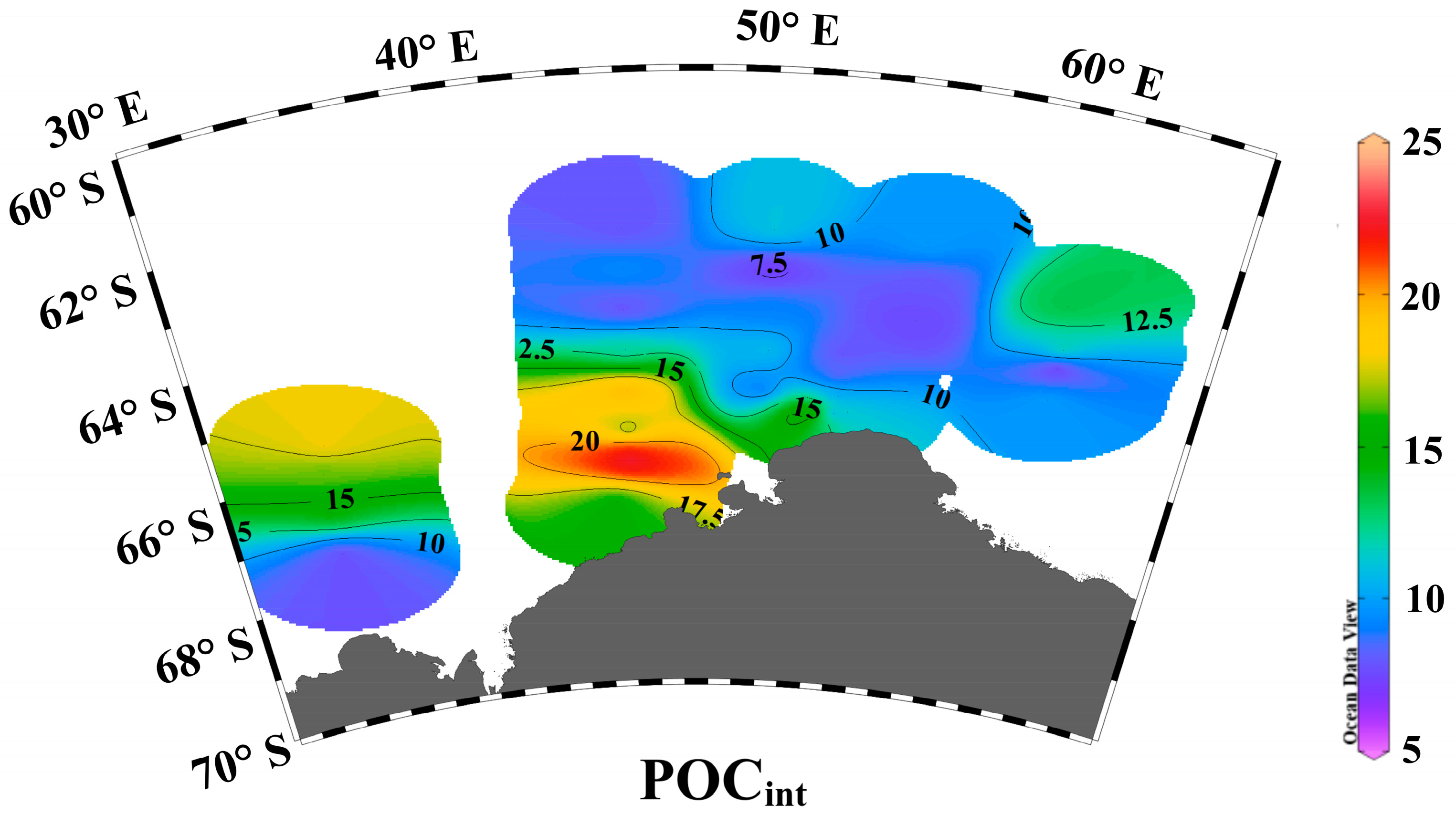
3.1. Environmental Parameters
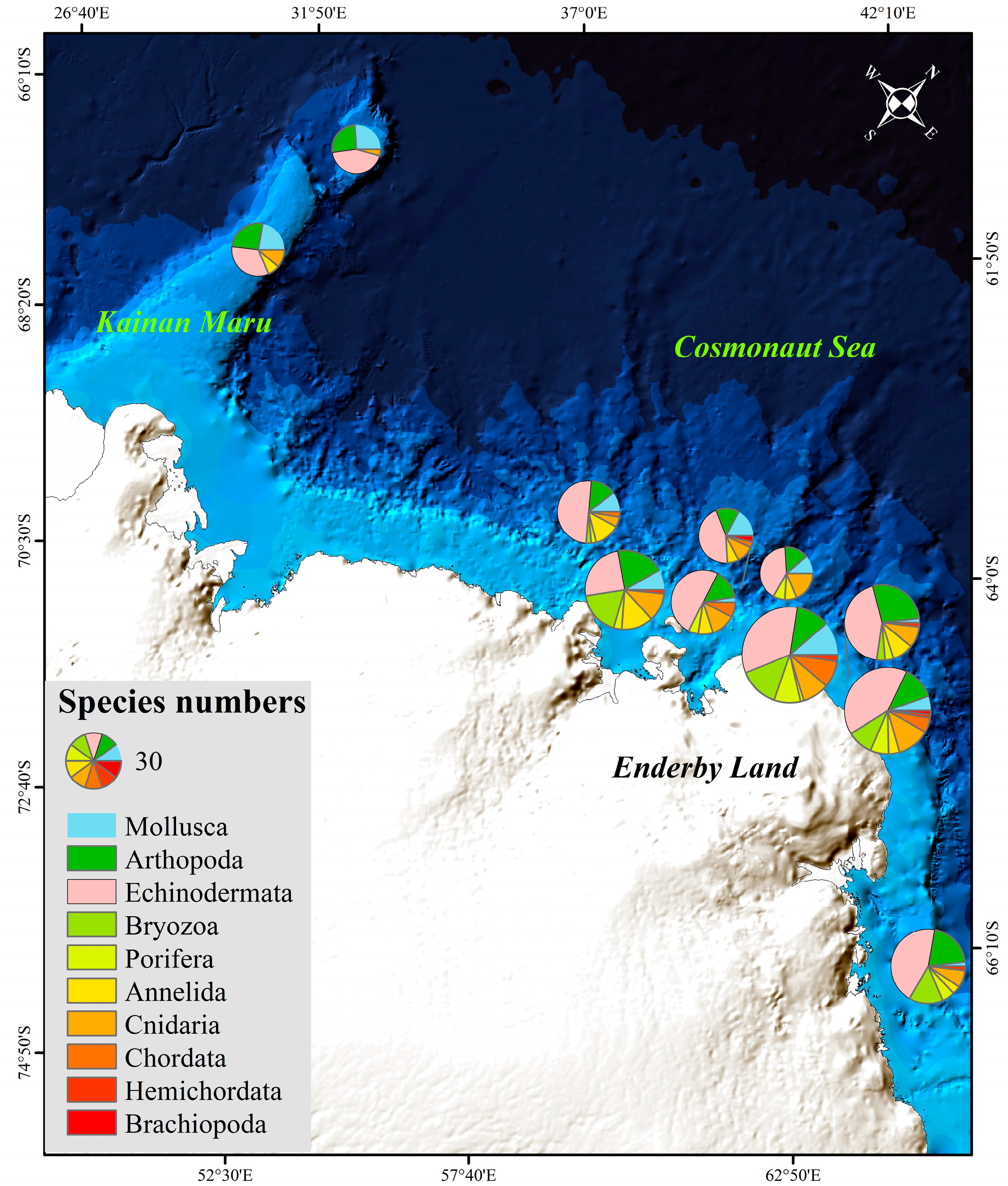
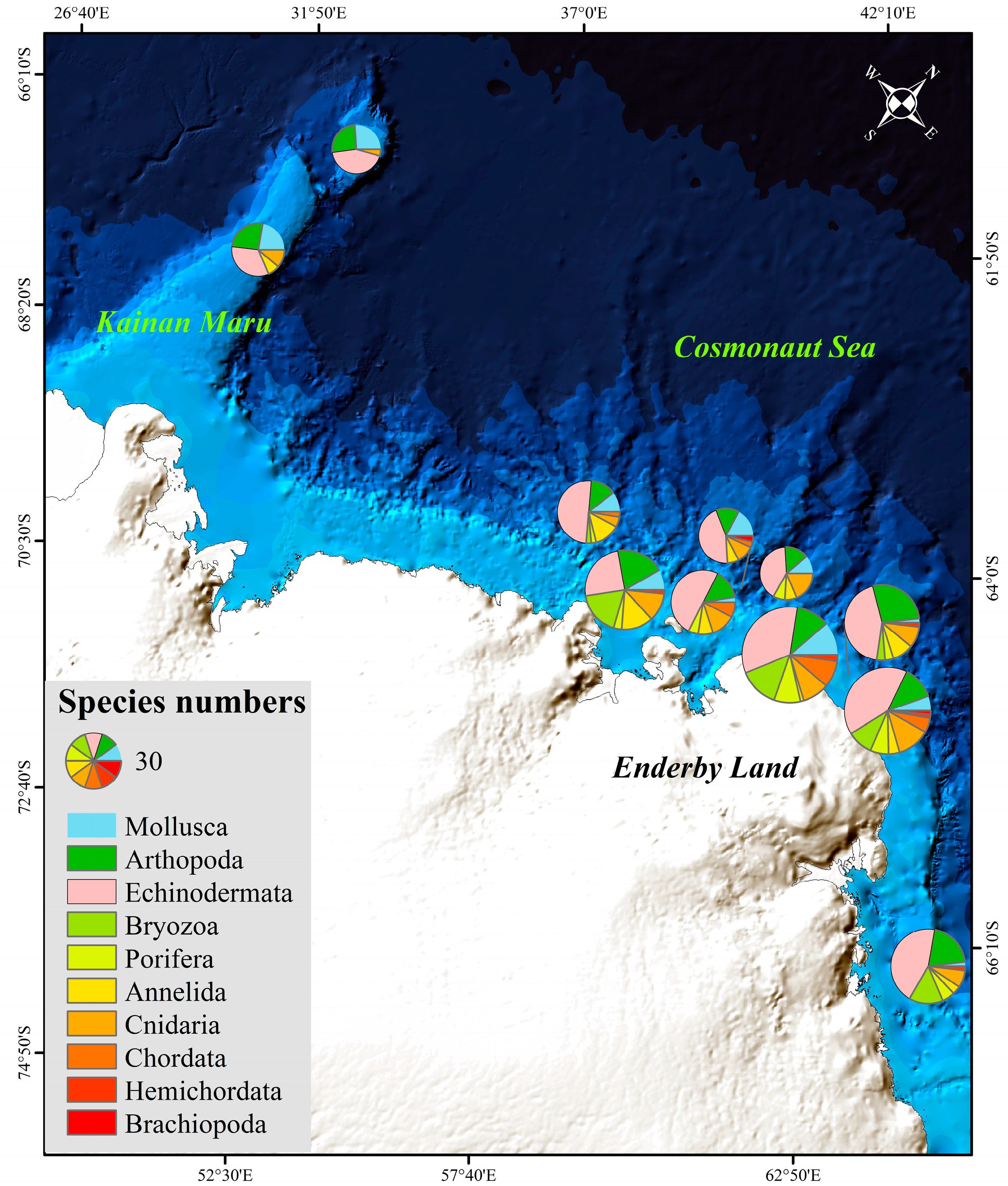
3.2. Macrobenthos
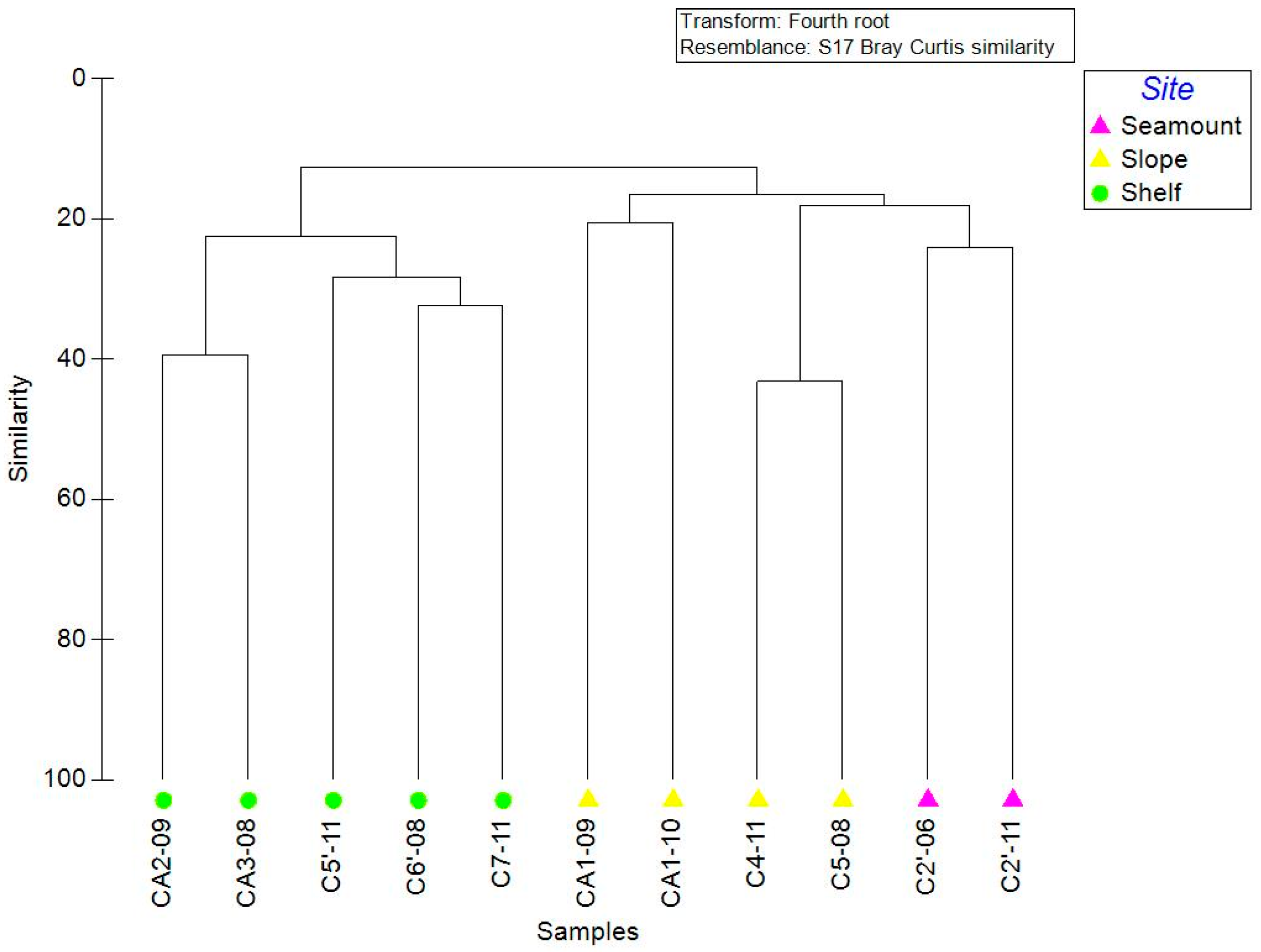
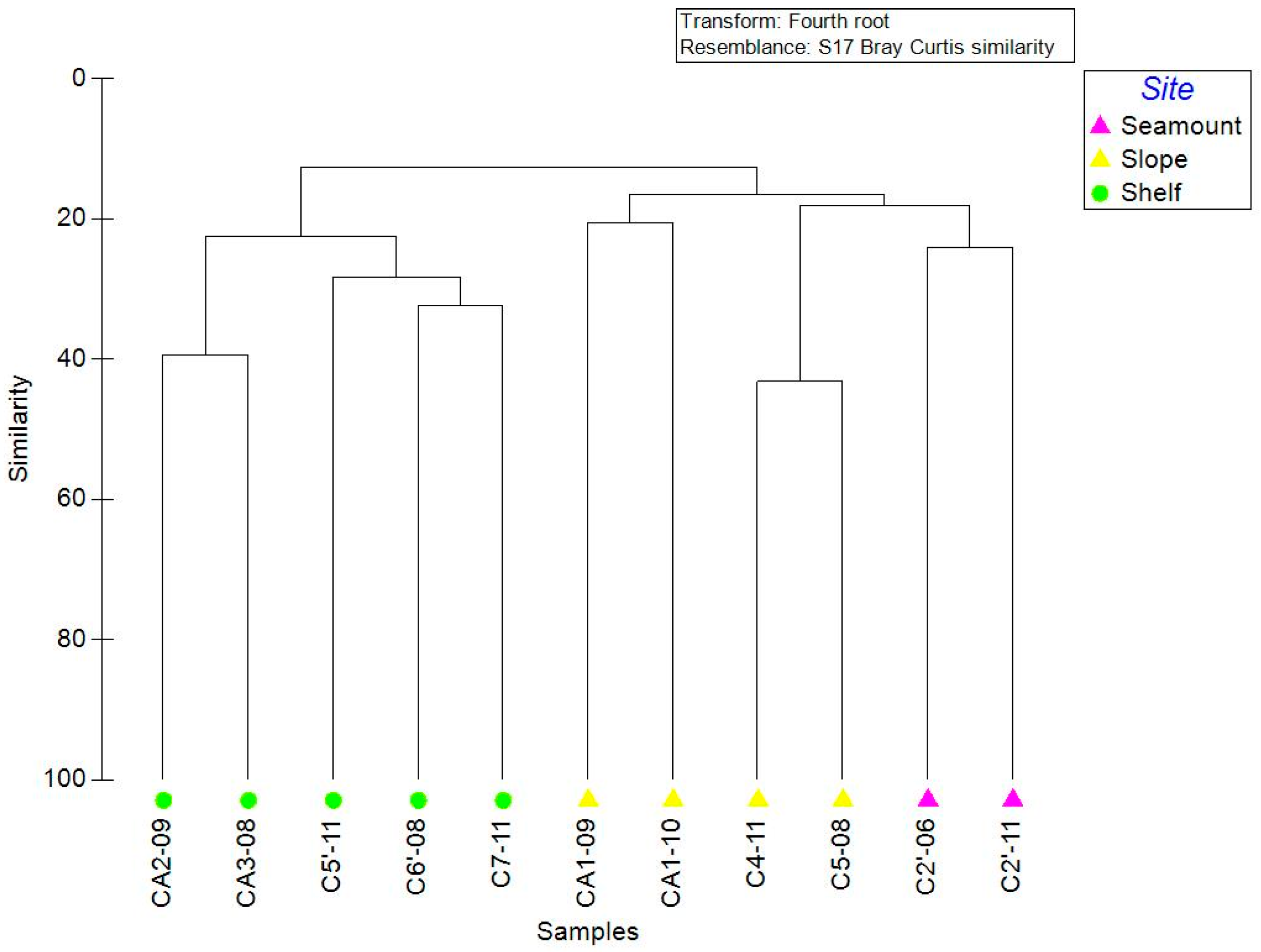
3.3. Community Structure
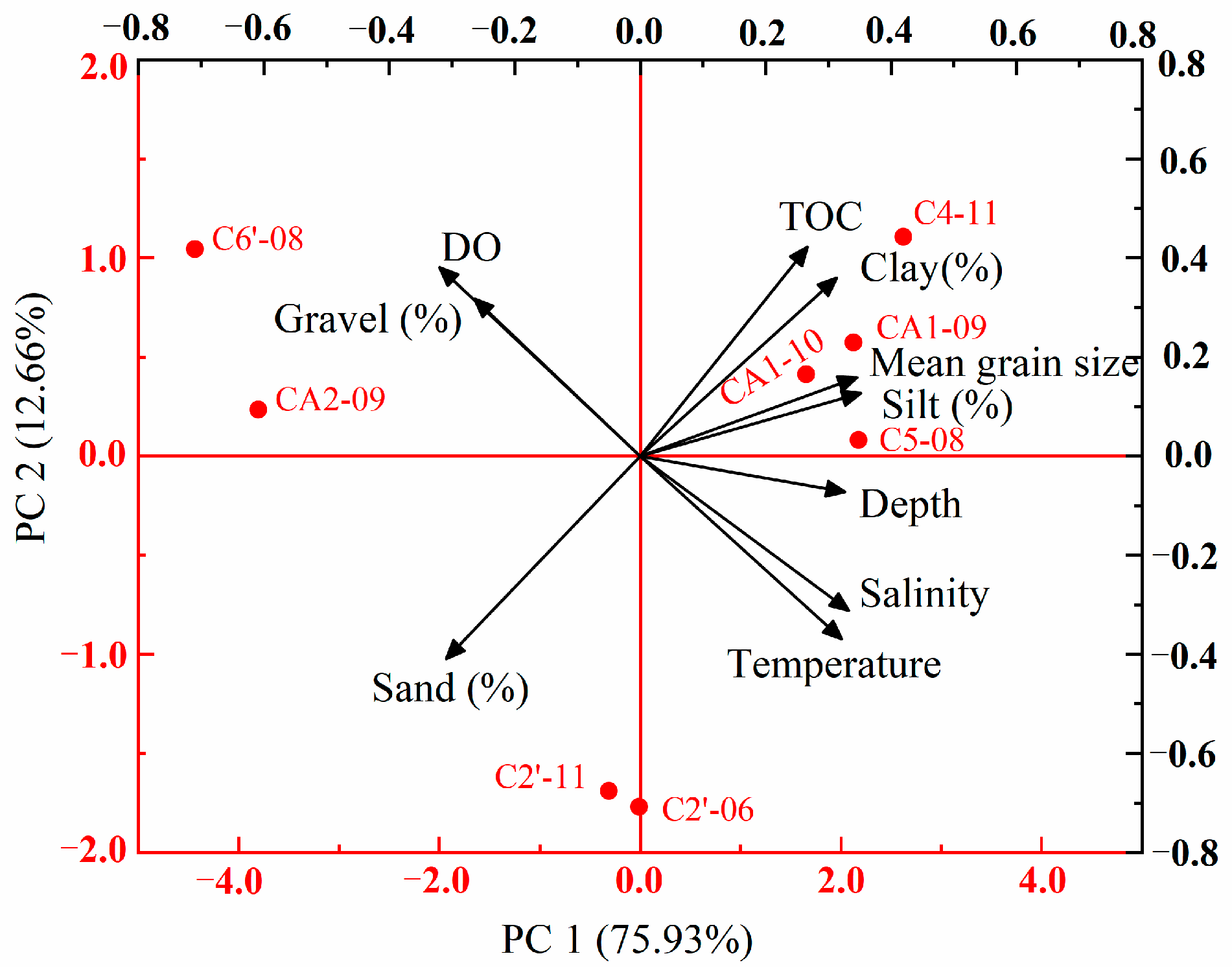
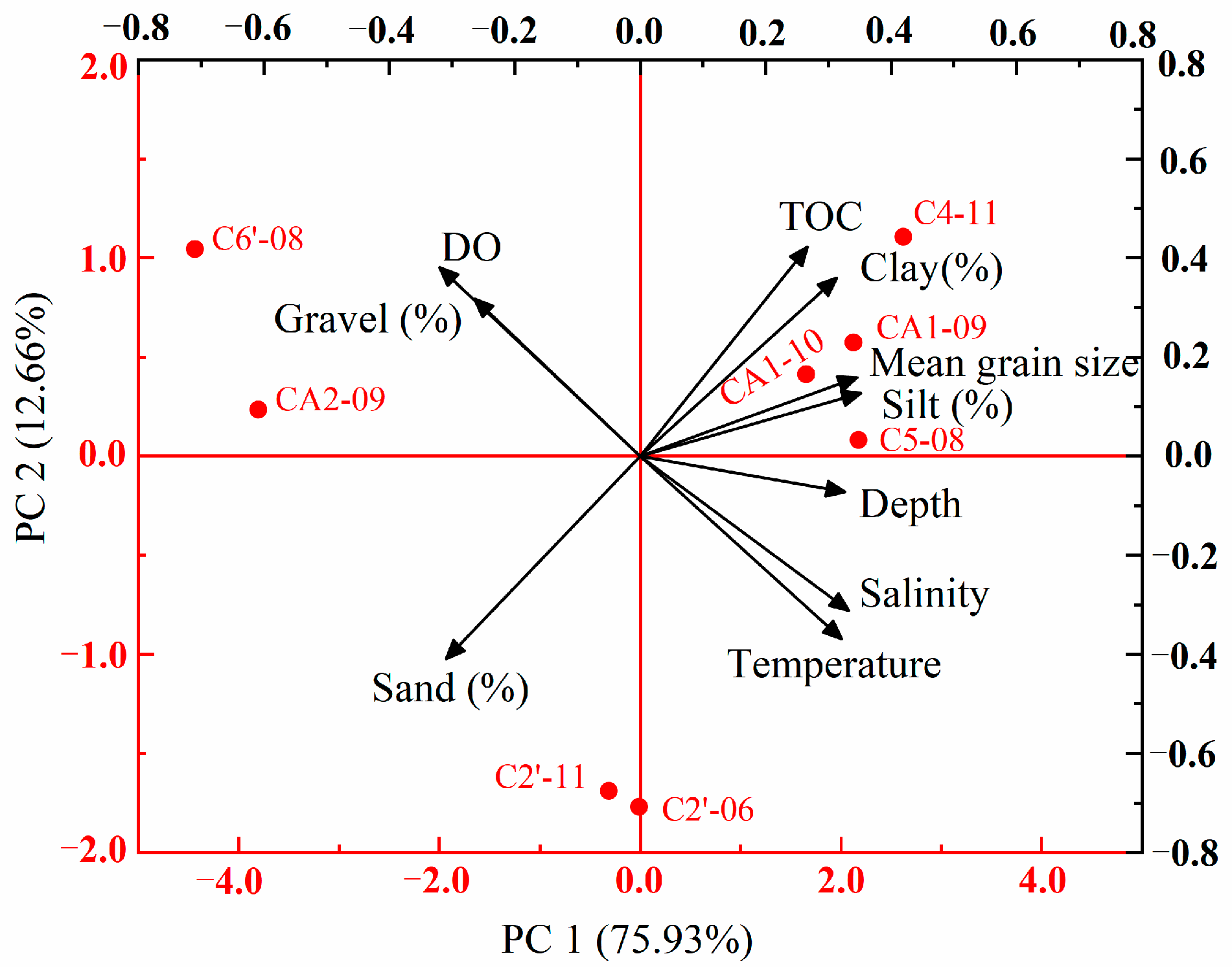
3.4. Relationship between the Macrobenthic Community and Environmental Factors
4. Discussion
4.1. Taxon Richness
4.2. Abundance
4.3. Community Structure
4.4. Species Distribution and Environmental Factors
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hunt, B.P.V.; Pakhomov, E.A.; Trotsenko, B.G. The macrozooplankton of the Cosmonaut Sea, east Antarctica (30° E–60° E), 1987–1990. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2007, 54, 1042–1069. [Google Scholar] [CrossRef]
- Ran, Q.; Duan, M.; Wang, P.; Ye, Z.; Mou, J.; Wang, X.; Tian, Y.; Zhang, C.; Qiao, H.; Zhang, J. Predicting the current habitat suitability and future habitat changes of Antarctic jonasfish Notolepis coatsorum in the Southern Ocean. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2022, 199, 105077. [Google Scholar] [CrossRef]
- Huang, W.H.; Yang, X.F.; Zhao, J. Dissolved nutrient distributions in the Antarctic Cosmonaut Sea in austral summer 2021. Adv. Polar Sci. 2022, 33, 267–290. [Google Scholar]
- Wright, S.W.; van den Enden, R.L.; Pearce, I.; Davidson, A.T.; Scott, F.J.; Westwood, K.J. Phytoplankton community structure and stocks in the Southern Ocean (30–80° E) determined by CHEMTAX analysis of HPLC pigment signatures. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 758–778. [Google Scholar] [CrossRef]
- Chimitza, V.A. Investigation of geostrophic currents in the Antarctic zone of the Indian Ocean. Oceanology 1976, 16, 234–238. [Google Scholar]
- Lubimova, T.G.; Makarov, R.R.; Maslennikov, V.V.; Samyshev, E.Z.; Bibik, V.A.; Tarverdieva, T.G. Interdisciplinary Investigations of Pelagic Ecosystem in the Commonwealth and Cosmonaut Seas; Collected Papers; VNIRO Publishers: Moscow, Russia, 1988; pp. 1–241. (In Russian) [Google Scholar]
- Davidson, A.T.; Scott, F.j.; Nash, G.V.; Wright, S.W.; Raymond, B. Physical and biological control of protistan community composition, distribution and abundance in the seasonal ice zone of the Southern Ocean between 30 and 80° E. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 828–848. [Google Scholar] [CrossRef]
- Swading, K.M.; Kawaguchi, S.; Hosie, G.W. Antarctic mesozooplankton community structure during BROKE-West (30° E–80° E), January–February 2006. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 887–904. [Google Scholar] [CrossRef]
- Westwood, K.J.; Griffiths, F.B.; Meiners, K.M.; Williams, G.D. Primary productivity off the Antarctic coast from 30° E–80° E.; BROKE-West survey, 2006. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 794–814. [Google Scholar] [CrossRef]
- Gutt, J.; Griffiths, H.J.; Jones, C.D. Circumpolar overview and spatial heterogeneity of Antarctic macrobenthic communities. Mar. Biodivers. 2013, 43, 481–487. [Google Scholar] [CrossRef]
- Linse, K.; Griffiths, H.J.; Barnes, D.K.A.; Brandt, A.; Davey, N.; David, B.; De Grave, S.; D′Acoz, C.D.; Eléaume, M.; Glover, A.G.; et al. The macro- and megabenthic fauna on the continental shelf of the eastern Amundsen Sea, Antarctica. Cont. Shelf Res. 2013, 68, 80–90. [Google Scholar] [CrossRef]
- Gambi, M.C.; Bussotti, S. Composition, abundance and stratification of softbottom macrobenthos from selected areas of the Ross Sea shelf (Antarctica). Polar Biol. 1999, 21, 347–354. [Google Scholar] [CrossRef]
- Brandt, A.; Gooday, A.J.; Brandão, S.N.; Brix, S.; Brökeland, W.; Cedhagen, T.; Choudhury, M.; Cornelius, N.; Danis, B.; De Mesel, I.; et al. First insights into the biodiversity and biogeography of the Southern Ocean deep sea. Nature 2007, 447, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Clarke, A.; Johnston, N.M. Antarctic marine benthic diversity. Oceanogr. Mar. Biol. 2003, 41, 47–114. [Google Scholar]
- Barnes, D.K.A.; Clarke, A. Antarctic marine biology. Curr. Biol. 2011, 21, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Mou, J.F.; Liu, K.; Huang, Y.Q.; He, X.; Zhang, S.; Wang, J.; Lin, J.; Lin, H.; Liu, W. The macro-and megabenthic fauna on the continental shelf of Prydz Bay, east Antarctica. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2022, 198, 105052. [Google Scholar] [CrossRef]
- Sahade, R.; Lagger, C.; Torre, L.; Momo, F.; Monien, P.; Schloss, I.; Barnes, D.K.A.; Servetto, N.; Tarantelli, S.; Tatián, M.; et al. Climate change and glacier retreat drive shifts in an Antarctic benthic ecosystem. Mar. Biol. 2015, 1, e1500050. [Google Scholar] [CrossRef] [PubMed]
- Arndt, J.E.; Schenke, H.W.; Jakobsson, M.; Nitsche, F.O.; Buys, G.; Goleby, B.; Rebesco, M.; Bohoyo, F.; Hong, J.; Black, J.; et al. The International Bathymetric Chart of the Southern Ocean (IBCSO) Version 1.0—A new bathymetric compilation covering circum-Antarctic water. Geophys. Res. Lett. 2013, 40, 3111–3117. [Google Scholar] [CrossRef]
- Gutt, J.; Alvaro, M.C.; Barco, A.; Böhmer, A.; Bracher, A.; David, B.; De Ridder, C.; Dorschel, B.; Eléaume, M.; Janussen, D.; et al. Macroepibenthic communities at the tip of the Antarctic Peninsula, an ecological survey at different spatial scales. Polar Biol. 2016, 39, 829–849. [Google Scholar] [CrossRef]
- Griffiths, H.J.; Linse, K.; Barnes, D.K.A. Distribution of macrobenthic taxa across the Scotia Arc, Antarctica. Antarct. Sci. 2008, 20, 213–226. [Google Scholar] [CrossRef]
- Chapman, M.G. Relationships between spatial patterns of benthic assemblages in a mangrove forest using different levels of taxonomic resolution. Mar. Ecol. Prog. Ser. 1998, 162, 71–78. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. PRIMER V6: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2006. [Google Scholar]
- Gutt, J.; Barnes, D.K.A.; Lockhart, S.J.; van de Putte, A. Antarctic macrobenthic communities: A compilation of circumpolar information. Nat. Conserv. 2013, 4, 1–13. [Google Scholar] [CrossRef]
- Gutt, J.; Sirenko, B.I.; Smirnov, I.S.; Arntz, W.E. How many macrobenthic species might inhabit the Antarctic shelf? Antarct. Sci. 2004, 16, 11–16. [Google Scholar] [CrossRef]
- Voβ, J. Zoogeographie und Gemeinschaftsanalyse des Makrozoobenthos des Weddellmeeres (Antarktis). Ber. Polarforsch. 1988, 45, 1–145. [Google Scholar]
- Arntz, W.E.; Thatje, S.; Linse, K.; Avila, C.; Ballesteros, M.; Barnes, D.K.; Cope, T.; Cristobo, F.J.; de Broyer, C.; Gutt, J.; et al. Sampling the marine fauna of remote Bouvet Island, Southern Ocean. Polar Biol. 2006, 29, 83–96. [Google Scholar] [CrossRef]
- Arntz, W.E.; Thatje, S.; Gerdes, D.; Gili, J.M.; Gutt, J.; Jacob, U.; Montiel, A.; Orejas, C.; Teixidó, N. The Antarctic-Magellan connection: Macrobenthos ecology on the shelf and upper slope, a progress report. Sci. Mar. 2005, 69, 237–269. [Google Scholar] [CrossRef]
- Basher, Z.; Bowden, D.A.; Costello, M.J. Diversity and Distribution of Deep-Sea Shrimps in the Ross Sea Region of Antarctica. PLoS ONE 2014, 9, e103195. [Google Scholar] [CrossRef]
- Post, A.L.; Beaman, R.J.; O′Brien, P.E.; Eléaume, M.; Riddle, M.J. Community structure and benthic habitats across the George V Shelf, East Antarctica: Trends through space and time. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2010, 58, 105–118. [Google Scholar] [CrossRef]
- Gutt, J. Antarctic macro-zoobenthic communities: A review and an ecological classification. Antarct. Sci. 2007, 19, 165–182. [Google Scholar] [CrossRef]
- Siegert, M.; Atkinson, A.; Banwell, A.; Brandon, M.; Convey, P.; Davies, B.; Downie, R.; Edwards, T.; Hubbard, B.; Marshall, G.; et al. The Antarctic Peninsula under a 1.5 °C global warming scenario. Front. Environ. Sci. 2019, 7, 102. [Google Scholar] [CrossRef]
- Lockhart, S.J.; Hocevar, J. Combined Abundance of AllVulnerable Marine Ecosystem Indicator Taxa Inadequate as Sole Determiner of Vulnerability, Antarctic Peninsula. Front. Mar. Sci. 2021, 8, 577761. [Google Scholar] [CrossRef]
- Li, Y.; Li, D.; Zhao, J.; Pan, J.; Hu, J.; Yu, P.; Zhang, C.; Yang, X.; Zhang, H.; Han, Z.; et al. Factors controlling the phytoplankton crops, taxonomic composition and particulate organic carbon stocks in the Cosmonaut Sea, East Antarctica. J. Oceanol. Limnol. 2023. submitted. [Google Scholar]
- Mou, J.; He, X.; Liu, K.; Huang, Y.; Zhang, S.; Zu, Y.; Liu, Y.; Cao, S.; Lan, M.; Miao, X.; et al. Benthic biodiversity by baited camera observations on the Cosmonaut Sea shelf of East Antarctica. Polar Bio. 2023. underview. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C2’-06 | C2’-11 | C4-11 | C5-08 | C5’-11 | CA1-09 | CA1-10 | C6’-08 | CA2-09 | CA3-08 | C7-11 | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Depth (m) | 1591 | 1152 | 1831 | 1924 | 445 | 1994 | 1070 | 486 | 207 | 330 | 701 |
| Temperature (℃) | 0.17 | 0.14 | −0.04 | 0.19 | −0.76 | −0.09 | 0.22 | −1.44 | −1.47 | −1.46 | 0.16 |
| Salinity (psu) | 34.67 | 34.66 | 34.66 | 34.66 | 34.43 | 34.66 | 34.66 | 34.24 | 34.20 | 34.34 | 34.64 |
| DO (mg/L) | 6.66 | 6.74 | 6.99 | 6.80 | 8.47 | 7.18 | 6.76 | 9.51 | 9.60 | 9.13 | 7.05 |
| Gravel (%) | 1.67 | 0.57 | 0.00 | 0.36 | - | 0.00 | 0.00 | 39.64 | 4.6 | - | - |
| Sand (%) | 50.06 | 55.92 | 4.78 | 6.28 | - | 7.37 | 19.41 | 49.26 | 72.06 | - | - |
| Silt (%) | 43.20 | 35.58 | 71.51 | 78.84 | - | 67.56 | 63.61 | 9.44 | 19.85 | - | - |
| Clay(%) | 5.05 | 7.92 | 23.70 | 14.51 | - | 25.07 | 16.99 | 1.65 | 3.51 | - | - |
| Mean grain size (phi) | 4.35 | 4.36 | 6.77 | 6.25 | - | 6.21 | 5.34 | 2.42 | 3.61 | - | - |
| TOC (%) | 0.11 | 0.12 | 0.61 | 0.36 | - | 0.31 | 0.67 | 0.10 | 0.08 | - | - |
| Phyla | C2’-06 | C2’-11 | C4-11 | C5-08 | C5’-11 | CA1-09 | CA1-10 | C6’-08 | CA2-09 | CA3-08 | C7-11 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Abundance (ind./1000 m2) | Annelida | 0 | 3 | 6 | 1 | 22 | 41 | 40 | 21 | 1 | 7 | 16 |
| Mollusca | 10 | 3 | 12 | 2 | 4 | 2 | 1 | 0 | 52 | 38 | 0 | |
| Arthopoda | 44 | 17 | 94 | 38 | 12 | 111 | 324 | 18 | 865 | 31 | 33 | |
| Echinodermata | 464 | 27 | 211 | 46 | 114 | 105 | 119 | 196 | 101 | 83 | 117 | |
| Porifera | 0 | 0 | 0 | 1 | 1 | 0 | 8 | 1 | 4 | 6 | 1 | |
| Cnidaria | 0 | 2 | 1 | 2 | 5 | 1 | 24 | 5 | 30 | 10 | 2 | |
| Bryozoa | 0 | 0 | 0 | 0 | 5 | 0 | 0 | 1 | 6 | 8 | 4 | |
| Brachiopoda | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Hemichordata | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 7 | 3 | 0 | |
| Chordata | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 0 | 155 | 9 | 0 | |
| Total | 519 | 52 | 326 | 91 | 164 | 262 | 522 | 243 | 1221 | 197 | 175 | |
| Biomass (g/1000 m2) | Annelida | 0 | 3.97 | 29.21 | 3.10 | 29.82 | 666.04 | 122.97 | 30.56 | 8.51 | 7.58 | 62.82 |
| Mollusca | 4.36 | 49.68 | 16.54 | 12.82 | 4.30 | 3.97 | 60.90 | 5.42 | 201.68 | 2110.95 | 0.16 | |
| Arthopoda | 40.84 | 49.54 | 706.06 | 220.38 | 10.61 | 774.64 | 1682.50 | 11.50 | 759.21 | 62.65 | 24.04 | |
| Echinodermata | 675.91 | 310.76 | 992.46 | 123.44 | 603.94 | 859.93 | 805.09 | 862.11 | 2187.06 | 548.21 | 176.61 | |
| Porifera | 0 | 0 | 3.64 | 12.26 | 7.19 | 0 | 142.70 | 642.73 | 7699.92 | 1372.75 | 438.14 | |
| Cnidaria | 0.31 | 45.23 | 27.86 | 5.28 | 39.07 | 0.84 | 86.01 | 88.31 | 387.86 | 46.79 | 3.77 | |
| Bryozoa | 0 | 0 | 0.76 | 0 | 159.15 | 0 | 0 | 12.67 | 187,636.23 | 782.02 | 12.06 | |
| Brachiopoda | 0 | 0 | 0 | 0 | 0 | 0.19 | 0 | 0 | 0 | 0.75 | 0 | |
| Hemichordata | 0 | 0 | 0 | 0 | 49.09 | 0 | 0 | 0.60 | 3877.76 | 128.97 | 660.15 | |
| Chordata | 0 | 0 | 3.76 | 0 | 0 | 5.31 | 31.98 | 0 | 3613.54 | 272.66 | 0 | |
| Total | 721.42 | 459.19 | 1780.28 | 377.28 | 903.18 | 2310.92 | 2932.15 | 1653.90 | 206,371.78 | 5333.34 | 1377.74 | |
| d | 3.16 | 5.58 | 5.69 | 4.98 | 10.32 | 4.62 | 5.60 | 8.70 | 11.25 | 12.01 | 9.02 | |
| J’ | 0.48 | 0.77 | 0.52 | 0.61 | 0.74 | 0.58 | 0.53 | 0.70 | 0.39 | 0.81 | 0.74 | |
| H’(log2) | 2.16 | 3.67 | 2.72 | 2.90 | 4.42 | 2.84 | 2.83 | 4.03 | 2.55 | 5.00 | 4.25 |
| Taxa | Average Abundance | Average Similarity | Similarity/Standard Deviation | Contribution% | Cumulative% |
|---|---|---|---|---|---|
| Group deep-water (Average similarity: 19.76) | |||||
| Nematocarcinus lanceopes | 3.3 | 5.94 | 5.91 | 30.03 | 30.03 |
| Ceratoserolis meridionalis | 1.8 | 3.23 | 5.93 | 16.33 | 46.36 |
| Ophiacantha sp. | 1.18 | 1.58 | 0.78 | 7.99 | 54.35 |
| Group shallow-water (Average similarity: 26.41) | |||||
| Ophioplinthus sp. | 1.76 | 1.76 | 3.73 | 6.65 | 6.65 |
| Echinisis sp.2 | 1.41 | 1.3 | 4.66 | 4.91 | 11.55 |
| Amphiophiura sp.1 | 1.81 | 1.2 | 0.98 | 4.55 | 16.1 |
| Reteporella hippocrepis | 1.16 | 1.14 | 7.11 | 4.31 | 20.41 |
| Ophioplinthus brevirima | 1.53 | 1.01 | 1.07 | 3.82 | 24.23 |
| Polynoidae undefined | 1.41 | 1.01 | 1.03 | 3.82 | 28.05 |
| Ophiacantha sp. | 1.4 | 0.85 | 0.62 | 3.23 | 31.28 |
| Colossendeis sp. | 1.09 | 0.8 | 1.13 | 3.03 | 34.31 |
| Henricia sp. | 0.94 | 0.72 | 1.14 | 2.72 | 37.02 |
| Cephalodiscus nigrescens | 0.99 | 0.7 | 1.13 | 2.65 | 39.67 |
| Rossella sp. | 0.8 | 0.66 | 1.14 | 2.5 | 42.17 |
| Fasciculipora ramosa | 0.8 | 0.66 | 1.14 | 2.49 | 44.67 |
| Hornera sp. | 0.8 | 0.66 | 1.14 | 2.49 | 47.16 |
| Orthoporidra sp. | 0.8 | 0.66 | 1.14 | 2.49 | 49.65 |
| Terebellidae undefined | 0.99 | 0.51 | 0.59 | 1.93 | 51.59 |
| Number of Variables | Correlation | Best Variables |
|---|---|---|
| 1 | 0.629 | depth |
| 3 | 0.615 | depth, DO, clay |
| 4 | 0.613 | depth, salinity, DO, clay |
| 3 | 0.604 | depth, salinity, clay |
| 2 | 0.603 | depth, DO |
| 4 | 0.594 | depth, temperature, DO, clay |
| 2 | 0.592 | depth, clay |
| 4 | 0.592 | depth, temperature, salinity, clay |
| 5 | 0.590 | depth, temperature, salinity, DO, clay |
| 4 | 0.589 | depth, salinity, silt, clay |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mou, J.; Liu, K.; Huang, Y.; Lin, J.; He, X.; Zhang, S.; Li, D.; Zu, Y.; Chen, Z.; Fu, S.; et al. Species Diversity and Community Structure of Macrobenthos in the Cosmonaut Sea, East Antarctica. Diversity 2023, 15, 1197. https://doi.org/10.3390/d15121197
Mou J, Liu K, Huang Y, Lin J, He X, Zhang S, Li D, Zu Y, Chen Z, Fu S, et al. Species Diversity and Community Structure of Macrobenthos in the Cosmonaut Sea, East Antarctica. Diversity. 2023; 15(12):1197. https://doi.org/10.3390/d15121197
Chicago/Turabian StyleMou, Jianfeng, Kun Liu, Yaqin Huang, Junhui Lin, Xuebao He, Shuyi Zhang, Dong Li, Yongcan Zu, Zhihua Chen, Sujing Fu, and et al. 2023. "Species Diversity and Community Structure of Macrobenthos in the Cosmonaut Sea, East Antarctica" Diversity 15, no. 12: 1197. https://doi.org/10.3390/d15121197
APA StyleMou, J., Liu, K., Huang, Y., Lin, J., He, X., Zhang, S., Li, D., Zu, Y., Chen, Z., Fu, S., Lin, H., & Liu, W. (2023). Species Diversity and Community Structure of Macrobenthos in the Cosmonaut Sea, East Antarctica. Diversity, 15(12), 1197. https://doi.org/10.3390/d15121197