
Bats of the Tunisian Desert: Preliminary Data Using Acoustic Identification and First Record of Taphozous nudiventris in the Country


Abstract
:1. Introduction
2. Material and Methods
2.1. Data Collection and Species Identification
2.2. Data Analysis
3. Results
3.1. Species Identification
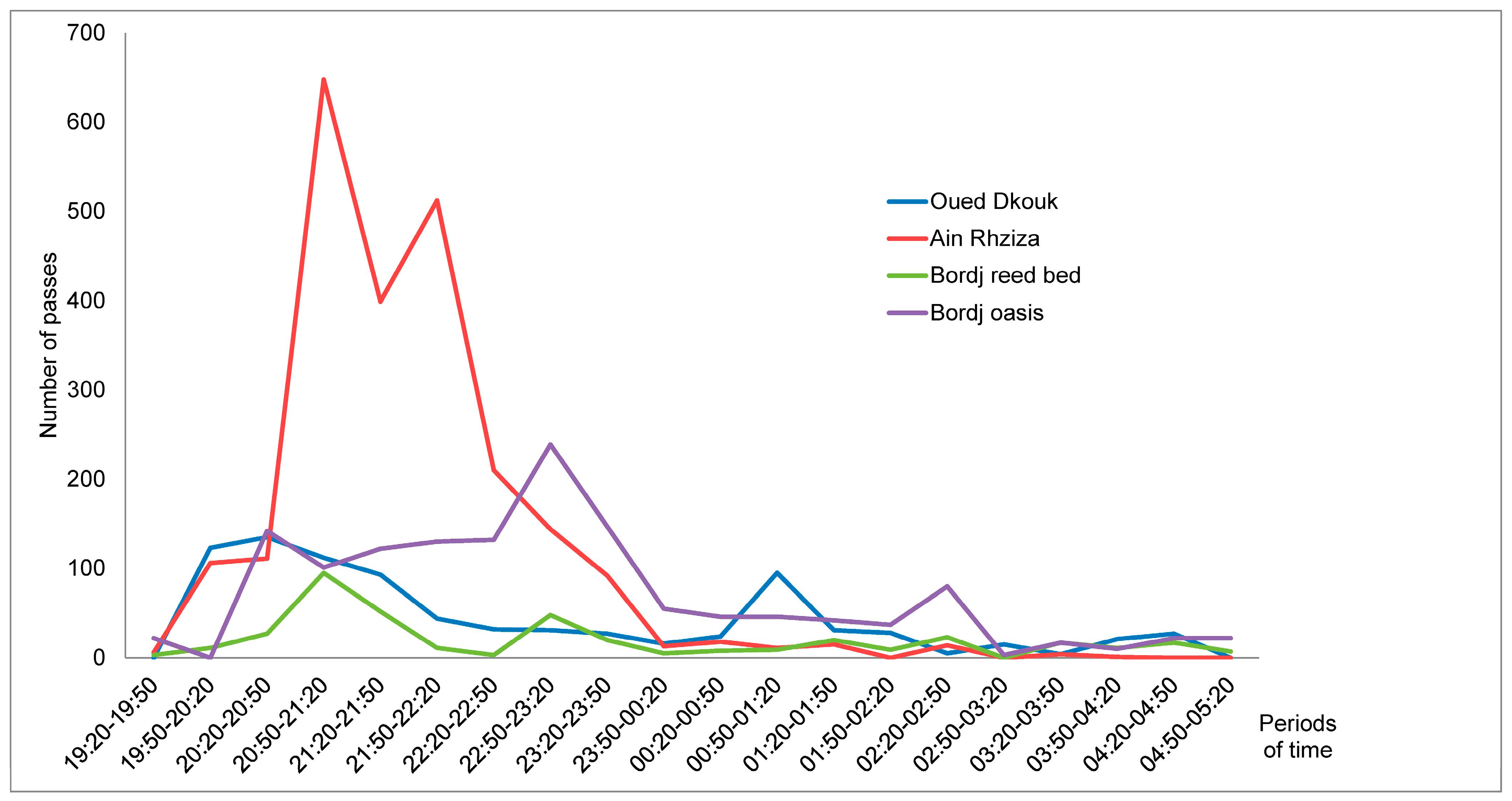
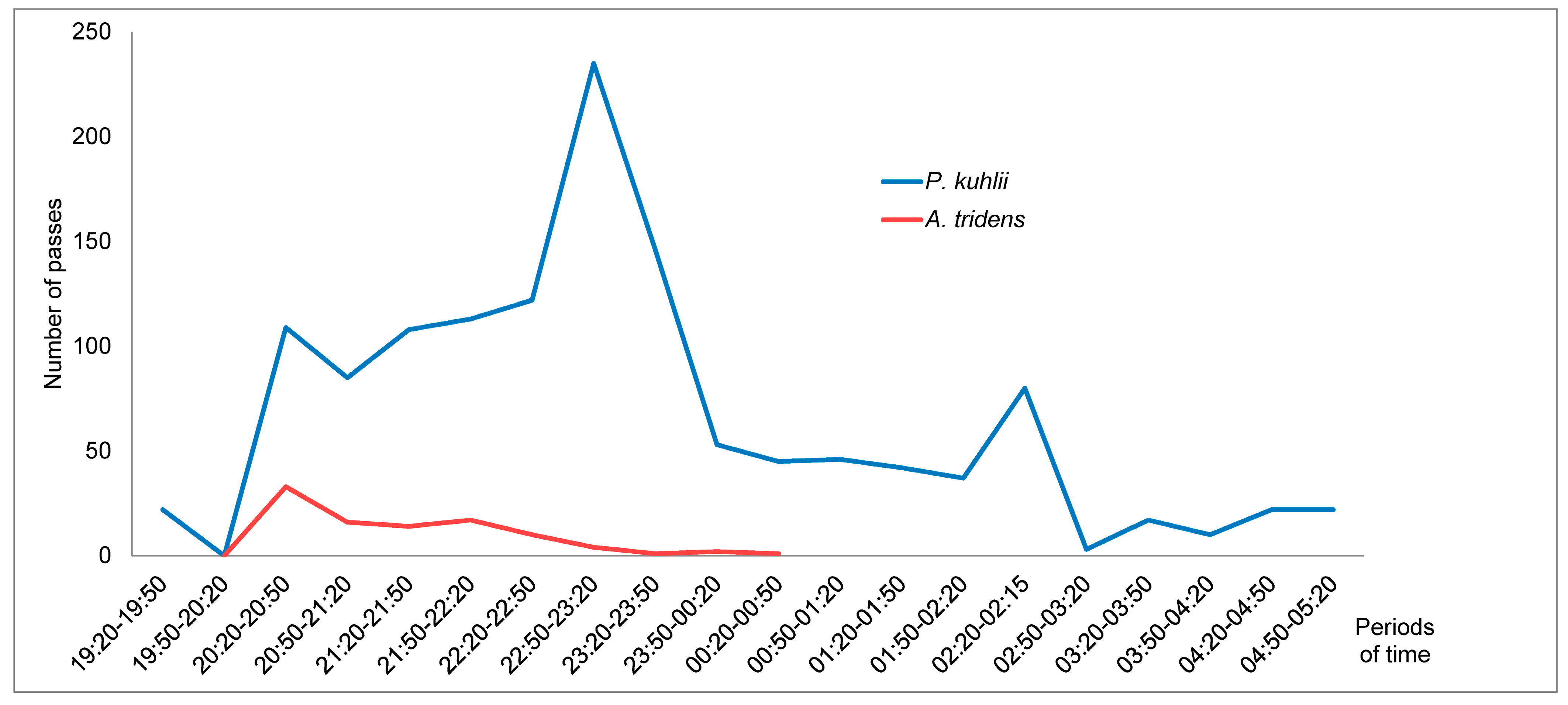
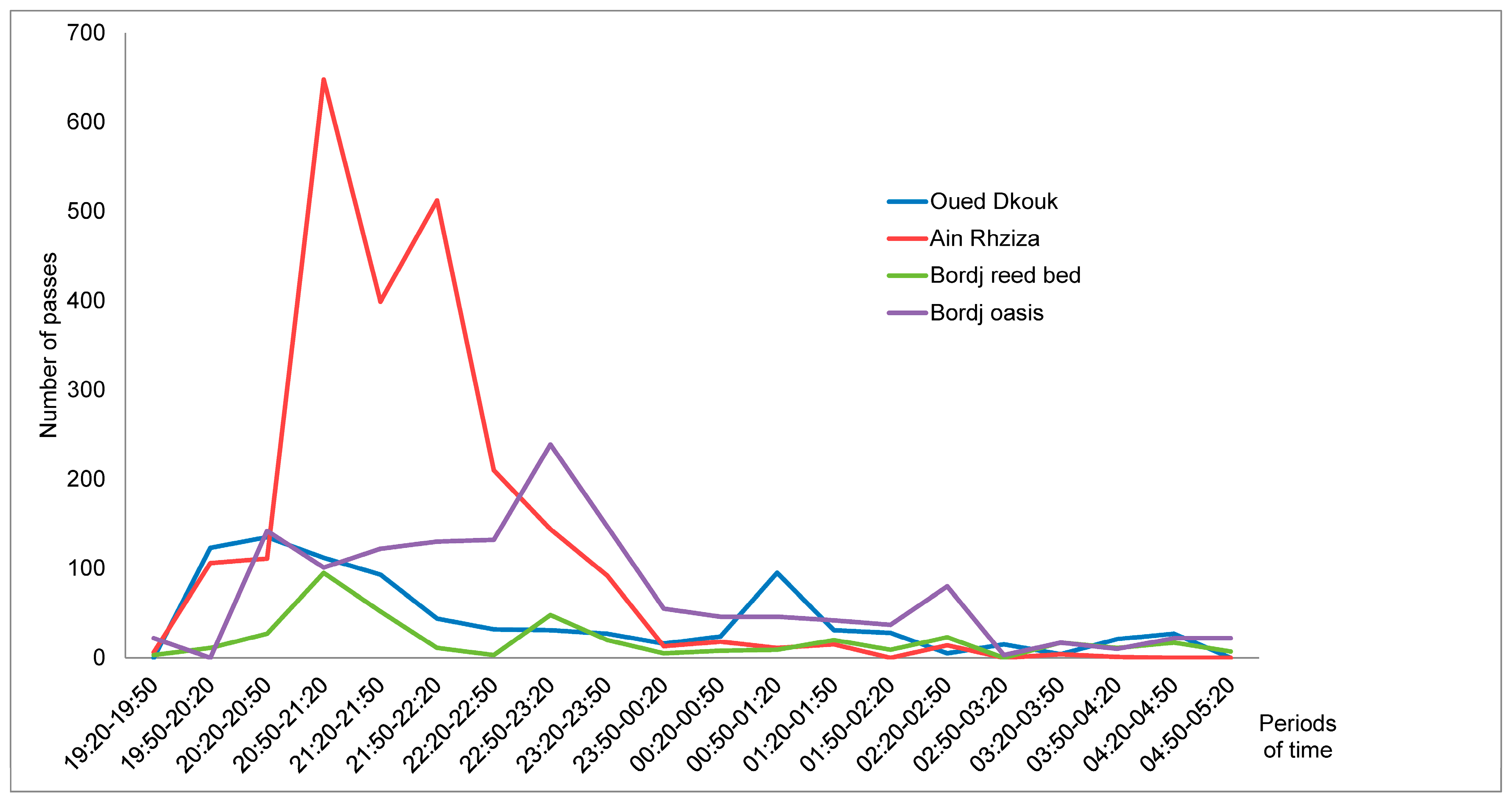
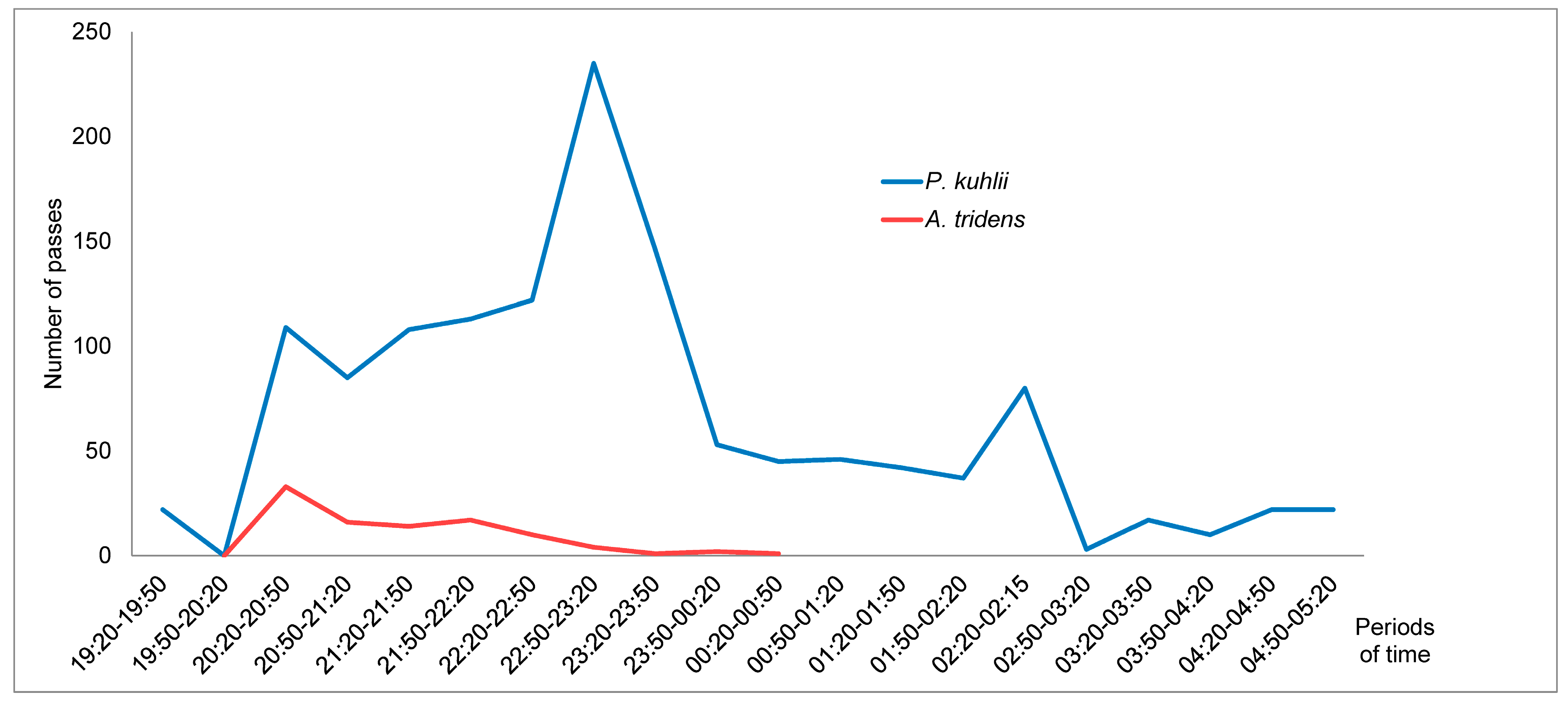
3.2. Species Richness and Bat Activity Patterns
4. Discussion
4.1. Echolocation Calls of Bats in the Tunisian Desert
4.2. Bat Assemblage and Activity Pattern
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Findley, J.S. Bats. A Community Perspective; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Herkt, K.M.B.; Barnikela, G.; Skidmore, A.K.; Fahr, J. A high-resolution model of bat diversity and endemism for continental Africa. Ecol. Model. 2016, 320, 9–28. [Google Scholar]
- Korine, C.; Pinshow, B. Guild structure, foraging space use, and distribution in a community of insectivorous bats in the Negev Desert. J. Zool. Lond. 2004, 262, 187–196. [Google Scholar]
- Adams, R.A.; Hayes, M.A. The importance of water availability to bats: Climate warming and increasing global aridity. In 50 Years of Bat Research. Foundations and New Frontiers; Lim, B.K., Fenton, M.B., Brigham, R.M., Mistry, S., Kurta, A., Gillam, E.H., Russell, A., Ortega, J., Eds.; Springer: Cham, Switzerland, 2021; pp. 105–120. [Google Scholar]
- Racey, P.A. The importance of the riparian environment as a habitat for British bats. Symp. Zool. Soc. Lond. 1998, 71, 69–91. [Google Scholar]
- Korine, C.; Adams, R.; Russo, D.; Fisher-Phelps, M.; Jacobs, D. Bats and water: Anthropogenic alterations threaten bat populations. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Voigt, C.C., Kingston, T., Eds.; Springer: Cham, Switzerland, 2016; pp. 215–241. [Google Scholar]
- Razgour, O.; Korine, C.; Saltz, D. Does interspecific competition drive patterns of habitat use in desert bat communities? Oecologia 2011, 167, 493–502. [Google Scholar]
- Razgour, O.; Korine, C.; Saltz, D. Pond characteristics as determinants of species diversity and community composition in desert bats. Anim. Conserv. 2010, 13, 505–513. [Google Scholar]
- Le Houérou, H.N. Définition et limites bioclimatiques du Sahara. Sécheresse 1990, 4, 246–259. [Google Scholar]
- Le Berre, M.; Le Guelte, L. Les Mammifères actuels dans l’espace saharien. Vie Milieu 1990, 40, 223–228. [Google Scholar]
- Happold, M.; Happold, D.C.D. (Eds.) Mammals of Africa. Volume IV. Hedgehogs, Shrews and Bats; Bloomsbury Publishing: London, UK, 2013. [Google Scholar]
- Simmons, N.B.; Cirranello, A.L. Bats of the World: A Taxonomic and Geographic Database. Version 1.3. Available online: https://batnames.org/ (accessed on 5 February 2023).
- Ahmim, M. Les Mammifères sauvages d’Algérie. Répartition et Biologie de la Conservation; Les Editions du Net: Saint-Ouen-sur-Seine, France, 2019; ISBN 978-2312068961. [Google Scholar]
- Aulagnier, S.; Cuzin, F.; Thévenot, M. Chiroptera. In Mammifères sauvages du Maroc. Peuplement, Répartition, Écologie; Aulagnier, S., Cuzin, F., Thévenot, M., Eds.; S.F.E.P.M.: Paris, France, 2017; pp. 117–154. [Google Scholar]
- Benda, P.; Spizenberger, F.; Hanák, V.; Andreas, M.; Reiter, A.; Ševčik, M.; Šmid, J.; Uhrin, M. Bats (Mammalia: Chiroptera) of the Eastern Mediterranean and Middle East. Part 11. On the bat fauna of Libya II. Acta Soc. Zool. Bohem. 2014, 78, 1–162. [Google Scholar]
- Benda, P.; Ševčík, M. Bats (Mammalia: Chiroptera) of the Eastern Mediterranean and Middle East. Part 16. Review of the distribution and taxonomy of bats in Egypt. Acta Soc. Zool. Bohem. 2020, 84, 115–279. [Google Scholar]
- Dalhoumi, R.; Aissa, P.; Aulagnier, S. Taxonomie et répartition des Chiroptères de Tunisie. Rev. Suisse Zool. 2011, 118, 265–292. [Google Scholar] [CrossRef]
- Puechmaille, S.; Hizem, W.M.; Allegrini, B.; Abiadh, A. Bat fauna of Tunisia: Review of records and new records, morphometrics and echolocation data. Vespertilio 2012, 16, 211–239. [Google Scholar]
- Dalhoumi, R.; Hedfi, A.; Aissa, P.; Aulagnier, S. Bats of Jebel Mghilla National Park (central Tunisia): First survey and habitat-related activity. Trop. Zool. 2014, 27, 53–62. [Google Scholar]
- Dalhoumi, R.; Aissa, P.; Aulagnier, S. Bat species richness and activity in the Bou Hedma National Park (central Tunisia). Turk. J. Zool. 2016, 40, 864–875. [Google Scholar]
- Dalhoumi, R.; Nefla, A.; Bedoui, W.; Ouni, R.; Aulagnier, S. Preliminary habitat related bat fauna of Mastouta-Bishshouk region (Northwest Tunisia). Vest. Zool. 2019, 53, 23–30. [Google Scholar] [CrossRef]
- Dalhoumi, R.; Aissa, P.; Beyrem, H.; Aulagnier, S. Bat species richness and activity in the desert Dghoumes National Park (Southwest Tunisia): A preliminary survey. Arxius Miscel. Zool. 2020, 18, 89–100. [Google Scholar]
- Bendjeddou, M.L.; Loumassine, H.E.; Metallaoui, W.; Chiheb, K.; Farfar, A.; Bounaceur, F.; Boukheroufa, F.; Bouslama, Z.; Dietz, C. First record of Nyctinomus aegyptiacus for Tunisia. Vespertilio 2016, 18, 23–27. [Google Scholar]
- Van Cakenberghe, V.; Benda, P.; Horaček, I. Pipistrellus deserti desert pipistrelle. In Mammals of Africa. Volume IV. Hedgehogs, Shrews and Bats; Happold, M., Happold, D.C.D., Eds.; Bloomsbury Publishing: London, UK, 2013; pp. 619–621. [Google Scholar]
- Benda, P.; Andriollo, T.; Ruedi, M. Systematic position and taxonomy of Pipistrellus deserti (Chiroptera: Vespertilionidae). Mammalia 2015, 79, 419–438. [Google Scholar]
- López-Baucells, A. Kuhl’s pipistrelle Pipistrellus kuhlii. In Handbook of the Mammals of the World. 9. Bats; Wilson, D.E., Mittermeier, R.A., Eds.; Lynx: Barcelona, Spain, 2019; p. 774. [Google Scholar]
- Burgin, C.J.; Wilson, D.E.; Mittermeier, R.A.; Rylands, A.B.; Lacher, T.E.; Sechrest, W. Illustrated Checklist of the Mammals of the World. Volume 2: Eulipotyphla to Carnivora; Lynx Edicions: Barcelona, Spain, 2020. [Google Scholar]
- Barataud, M. Ecologie Acoustique des Chiroptères d’Europe. Identification des Espèces, Étude de Leurs Habitats et Comportements de Chasse; Biotope—M.N.H.N.: Paris, France, 2012. [Google Scholar]
- Gamoun, M.; Louhaichi, M. Botanical composition and species diversity of arid and desert rangelands in Tataouine, Tunisia. Land 2021, 10, 313. [Google Scholar]
- Russ, J. The Bats of Britain and Ireland: Echolocation Calls, Sound Analysis, and Species Identification; Alana Ecology Books: Belfast, UK, 1999. [Google Scholar]
- Russo, D.; Ancillotto, L.; Cistrone, L.; Korine, C. The buzz of drinking on the wing in echolocating bats. Ethology 2016, 122, 226–235. [Google Scholar] [CrossRef]
- Russo, D.; Jones, G. Use of foraging habitats by bats (Mammalia: Chiroptera) in a Mediterranean area determined by acoustic surveys: Conservation implications. Ecography 2003, 26, 197–209. [Google Scholar]
- R Core Team. R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: http://www.R-project.org/ (accessed on 20 August 2023).
- Disca, T.; Allegrini, B.; Prié, V. Caractéristiques acoustiques des cris d’écholocation de 16 espèces de Chiroptères (Mammalia, Chiroptera) du Maroc. Vespère 2014, 3, 209–229. [Google Scholar]
- Loumassine, H.; Allegrini, B.; Bounaceur, F.; Peyre, O.; Aulagnier, S. A new mammal species for Algeria, Rhinopoma microphyllum (Chiroptera: Rhinopomatidae): Morphological and acoustic identification. Mammalia 2018, 82, 85–88. [Google Scholar] [CrossRef]
- Gustafson, Y.; Schnitzler, H.U. Echolocation and obstacle avoidance in the hipposiderid bat Asellia tridens. J. Comp. Physiol. 1979, 131, 161–167. [Google Scholar] [CrossRef]
- Ahmim, M.; Dalhoumi, R.; Ştefan Măntoiu, D. First data on the acoustic characteristics of some Chiropteran species from northern Algeria. Bioacoustics 2020, 29, 499–517. [Google Scholar] [CrossRef]
- Mifsud, C.M.; Vella, A. Mitochondrial genetic diversity of bat species from the Maltese Islands and applications for their conservation. Nat. Engin. Sci. 2019, 4, 276–292. [Google Scholar] [CrossRef]
- Moores, R.; Brown, D. New records of bats from southern Morocco (Atlantic Sahara) and notes on echolocation. Mammalia 2017, 81, 611–614. [Google Scholar] [CrossRef]
- Horta, P.; Raposeira, H.; Santos, H.; Alves, P.; Palmeirim, J.; Godinho, R.; Jones, G.; Rebelo, H. Bats’ echolocation call characteristics of cryptic Iberian Eptesicus species. Eur. J. Wildl. Res. 2015, 61, 813–818. [Google Scholar] [CrossRef]
- Loumassine, H.E.; Bonnot, N.; Allegrini, B.; Bendjeddou, M.L.; Bounaceur, F.; Aulagnier, S. How arid environments affect spatial and temporal activity of bats. J. Arid Environ. 2020, 180, 104206. [Google Scholar] [CrossRef]
- Russo, D.; Jones, G. Identification of twenty-two bat species (Mammalia: Chiroptera) from Italy by analysis of time-expanded recordings of echolocation calls. J. Zool. Lond. 2002, 258, 91–103. [Google Scholar] [CrossRef]
- Fichera, G.; Mucedda, M.; Russo, D.; Tomassini, A.; Kiefer, A.; Veith, M.; Ancillotto, L. Pantelleria island (Sicily, Italy): A biogeographic crossroad for bats between Africa and Europe. Hystrix Ital. J. Mammal. 2022, 33, 134–137. [Google Scholar]
- Papadatou, E.; Butlin, R.K.; Altringham, J.D. Identification of bat species in Greece from their echolocation calls. Acta Chiropterol. 2008, 10, 127–143. [Google Scholar] [CrossRef]
- Hackett, T.D.; Holderied, M.W.; Korine, C. Echolocation call description of 15 species of Middle-Eastern desert dwelling insectivorous bats. Bioacoustics 2017, 26, 217–235. [Google Scholar] [CrossRef]
- Benda, P.; Lucan, R.K.; Obuch, J.; Reiter, A.; Andreas, M.; Backor, P.; Bohnenstengel, T.; Eid, E.K.; Ševcík, M.; Vallo, P.; et al. Bats (Mammalia: Chiroptera) of the eastern Mediterranean and Middle East. Part 8. Bats of Jordan: Fauna, ecology, echolocation, ectoparasites. Acta Soc. Zool. Bohem. 2010, 74, 185–353. [Google Scholar]
- Benda, P.; Andreas, M.; Kock, D.; Horový, J.; Lučan, R.K.; Maltby, A.; Meakin, K.; Truscott, J.; Vallo, P. Bats (Mammalia: Chiroptera) of the Eastern Mediterranean and Middle East. Part 6. Bats of Sinai (Egypt) with some taxonomic, ecological and echolocation data on that fauna. Acta Soc. Zool. Bohem. 2008, 72, 1–103. [Google Scholar]
- Berger-Tal, O.; Berger-Tal, R.; Korine, C.; Holderied, M.W.; Fenton, M.B. Echolocation calls produced by Kuhl’s pipistrelles in different flight situations. J. Zool. Lond. 2008, 274, 59–64. [Google Scholar] [CrossRef]
- Benda, P.; Dietz, C.; Andreas, M.; Kock, D.; Lučan, R.K.; Munclinger, P.; Nová, P.; Obuch, J.; Ochman, K.; Reiter, A.; et al. Bats (Mammalia: Chiroptera) of the Eastern Mediterranean. Part 4. Bat fauna of Syria: Distribution, systematics, ecology. Acta Soc. Zool. Bohem. 2006, 70, 1–329. [Google Scholar]
- Spitzenberger, F. The systematic status of the Libyan bats of the genus Plecotus (Chiroptera: Vespertilionidae). Lynx Ser. Nova 2020, 51, 219–221. [Google Scholar] [CrossRef]
- Benda, P.; Cervený, J.; Konecný, A.; Reiter, A.; Ševcík, M.; Uhrin, M.; Vallo, P. Some new records of bats from Morocco (Chiroptera). Lynx Ser. Nova 2010, 41, 151–166. [Google Scholar]
- Pereswiet-Soltan, A. Relation between climate and bat fauna in Europe. Trav. Du Muséum D’histoire Nat. Rigore Antipa 2007, 50, 505–515. [Google Scholar]
- Hanák, V.; Elgadi, A. On the bat fauna (Chiroptera) of Lybia. Vĕst. Čs. Společ. Zool. 1984, 48, 165–187. [Google Scholar]
- Benda, P.; Hanák, V.; Andreas, M.; Reiter, A.; Uhrin, M. Two new species of bats (Chiroptera) for the fauna of Libya: Rhinopoma hardwickii and Pipistrellus rueppellii. Myotis 2004, 41–42, 109–124. [Google Scholar]
- Benda, P.; Lučan, R.K.; Shohdi, W.M.; Porteš, M.; Horáček, I. Microbats of the western oases of Egypt, Libyan Desert. Vespertilio 2014, 17, 45–58. [Google Scholar]
- Feldman, R.; Whitaker, J.O.; Yom-Tov, Y. Dietary composition and habitat use in a desert insectivorous bat community in Israel. Acta Chiropterol. 2000, 2, 15–22. [Google Scholar]
- Hackett, T.D.; Korine, C.; Holderied, M.W. The importance of Acacia trees for insectivorous bats and arthropods in the Arava Desert. PLoS ONE 2013, 8, e52999. [Google Scholar]
- Korine, C.; Adams, R.; Shamir, U.; Gross, A. Effect of water quality on species richness and activity of desert-dwelling bats. Mammal. Biol. 2015, 80, 185–190. [Google Scholar] [CrossRef]
- Horáček, I. Otonycteris hemprichii Hemprich’s desert bat. In Mammals of Africa. Volume IV. Hedgehogs, Shrews and Bats; Happold, M., Happold, D.C.D., Eds.; Bloomsbury Publishing: London, UK, 2013; pp. 598–599. [Google Scholar]
- Kowalski, K.; Rzebik-Kowalska, B. Mammals of Algeria; Ossolineum: Wroclaw, Poland, 1991. [Google Scholar]
- Bendjeddou, M.L.; Bouam, I.; Khelfaoui, F. First photographed record of the naked-rumped tomb bat, Taphozous nudiventris Cretzschmar, 1830 (Chiroptera: Emballonuridae), in the Grand Maghreb. Afr. J. Ecol. 2020, 58, 852–854. [Google Scholar] [CrossRef]
- Aulagnier, S.; Denys, C. Présence du Taphien à ventre nu, Taphozous nudiventris, (Chiroptera, Emballonuridae) au Maroc. Mammalia 2000, 64, 116–118. [Google Scholar]
- Djilali, K.; Sekour, M.; Souttou, K.; Ababsa, L.; Guezoul, O.; Denys, C.; Doumandji, S. Diet of short-eared owl Asio flammeus (Pontoppidan, 1763) in desert area at Hassi El Gara (El Golea, Algeria). Zool. Ecol. 2016, 26, 159–165. [Google Scholar] [CrossRef]
- Rebelo, H.; Brito, J.C. Bat guild structure and habitat use in the Sahara desert. Afr. J. Ecol. 2006, 45, 228–230. [Google Scholar] [CrossRef]
- Williams, A.J.; Dickman, C.R. The ecology of insectivorous bats in the Simpson Desert central Australia: Habitat use. Austral. Mammal. 2004, 26, 205–214. [Google Scholar] [CrossRef]
- Rabe, M.J.; Rosenstock, S.S. Influence of water size and type on bat captures in the lower Sonoran Desert. West. North Am. Nat. 2005, 65, 87–90. [Google Scholar]
- Adams, R.A.; Kwiecinski, G.G. Sonar surveys for bat species richness and activity in the Southern Kalahari Desert, Kgalagadi Transfrontier Park, South Africa. Diversity 2018, 10, 103. [Google Scholar]
- Egert-Berg, K.; Hurme, E.R.; Greif, S.; Goldstein, A.; Harten, L.; Flores-Martínez, J.J.; Valdés, A.T.; Johnston, D.S.; Eitan, O.; Borissov, I.; et al. Resource ephemerality drives social foraging in bats. Curr. Biol. 2018, 28, 3667–3673. [Google Scholar] [PubMed]
- de Jong, J.; Millon, L.; Håstad, O.; Victorsson, J. Activity pattern and correlation between bat and insect abundance at wind turbines in south Sweden. Animals 2021, 11, 3269. [Google Scholar] [PubMed]
- Greenfeld, A.; Saltz, D.; Kapota, D.; Korine, C. Managing anthropogenic driven range expansion behaviourally: Mediterranean bats in desert ecosystems. Eur. J. Wildl. Res. 2018, 64, 24. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
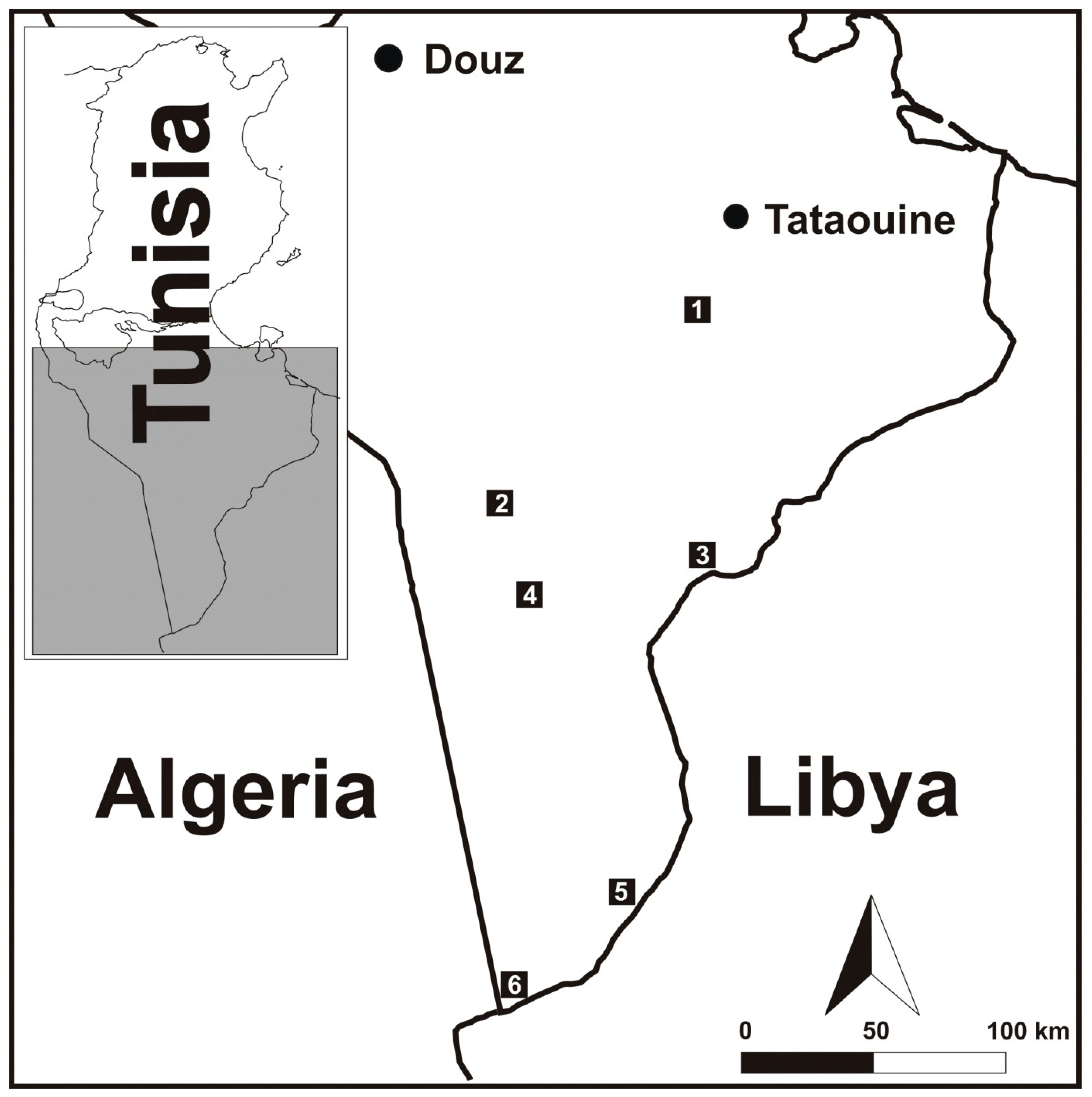
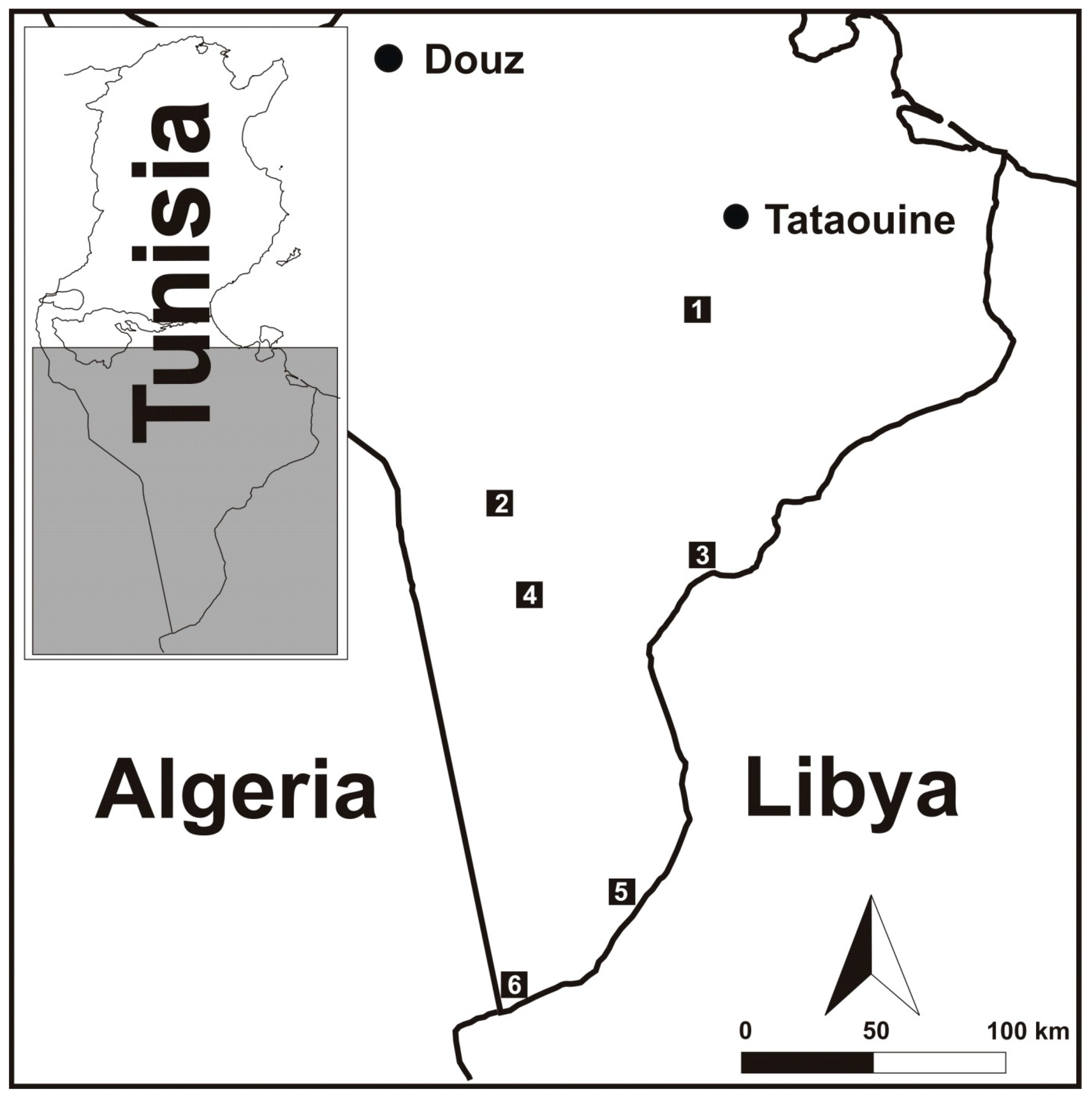
| Habitats | GPS Coordinates | Biotopes | |
|---|---|---|---|
| Oued Dkouk Nature Reserve | 32.624197, 10.308822 | Open habitat, Oued Daghsen bed with small lakes of salt water from Aïn Edkouk. The vegetation is sparse and represented by Tamarix gallica, Limonium pruinosum, Phragmites australis, Sarcocornia fruticosa, and Frankenia pulverulenta. | |
| Senghar Jabbes National Park | Lighted place | 31.958655, 9.547339 | Open habitat lighted with a lamp fixed to a wall. |
| Small trough | 31.940659, 9.528980 | Open habitat including an artificial small trough. | |
| Lorzet | 31.765947, 10.347375 | Small artificial trough near old human buildings. | |
| Ain Rhziza | 31.657026, 9.659167 | Open habitat including a water lake (50 m2) surrounded by groves of Tamarix aphylla and T. gallica. | |
| Ain Skhouna | 30.622722, 10.031079 | Open habitat corresponding to an isolated greenery (less than 0.5 ha) in the desert close to a hot spring. The vegetation cover includes Tamarix aphylla, Phragmites australis, Juncus maritimus, Tetraena alba, Phoenix dactylifera, Retama sphaerocarpa, Imperata cylindrica, Stipagrostis pungens, and Oudneya africana. | |
| Bordj el Khadra | Reed bed | 30.254260, 9.558299 | Open habitat including a small sewage water body. The vegetation includes Phragmites australis, Tetraena alba, T. geslinii, and Tamarix gallica. |
| Oasis | 30.250482, 9.558967 | Semi-open habitat inan old abandoned oasis. The vegetation cover is dominated by Phoenix dactylifera, Eucalyptus spp., Acacia spp., Heliotropium curassavicum, Tamarix aphylla, Cistanche violacea, Tetraena alba, T. geslinii, and Frankenia pulverulenta. | |
| Locality | Oued Dkouk Nature Reserve | Ain Rhziza | Ain Skhouna | Bordj el Khadra | ||
|---|---|---|---|---|---|---|
| Reed Bed | Oasis | |||||
| Species | ||||||
| Asellia tridens D | - | - | - | 5 | 81 (17) | |
| Tadarida teniotis M | 21 | - | - | - | - | |
| Eptesicus isabellinus M | 113 (3) | - | - | - | - | |
| Pipistrellus kuhlii M, D | 702 (36) | 2304 (621–42) | 18 (4) | 379 (18) | 1212 (105) | |
| Plecotus gaisleri M | 3 | - | - | 2 | - | |
| Otonycteris hemprichii D | 12 (1) | - | - | 10 | - | |
| Chiroptera sp. | 12 | - | - | - | - | |
| Total | 903 | 2967 | 22 | 414 | 1415 | |
| Species richness | 6 | 1 | 1 | 4 | 2 | |
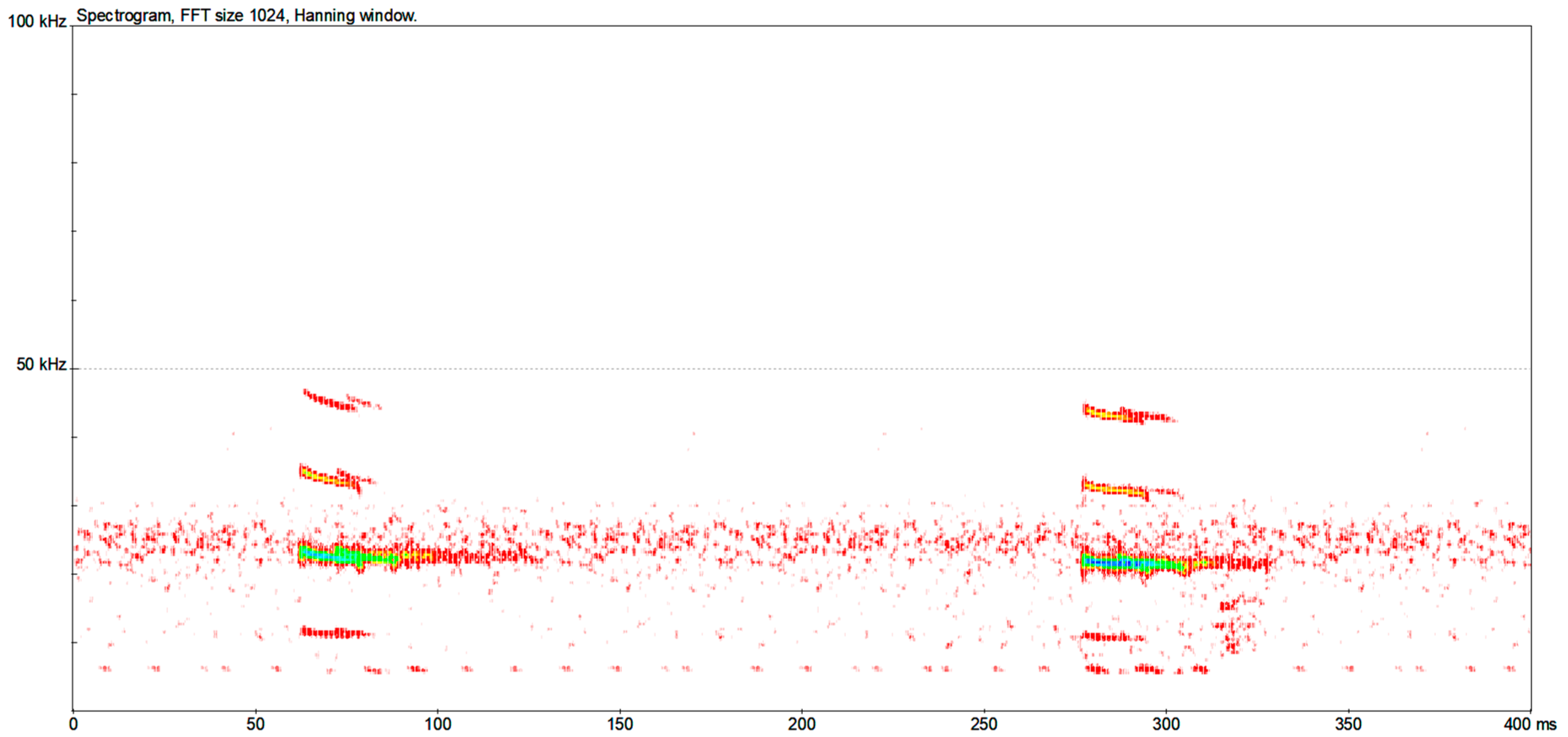
| Species | CS | N (S) | SF (kHz) | EF (kHz) | FMAXE (kHz) | D (ms) | IPI (ms) |
|---|---|---|---|---|---|---|---|
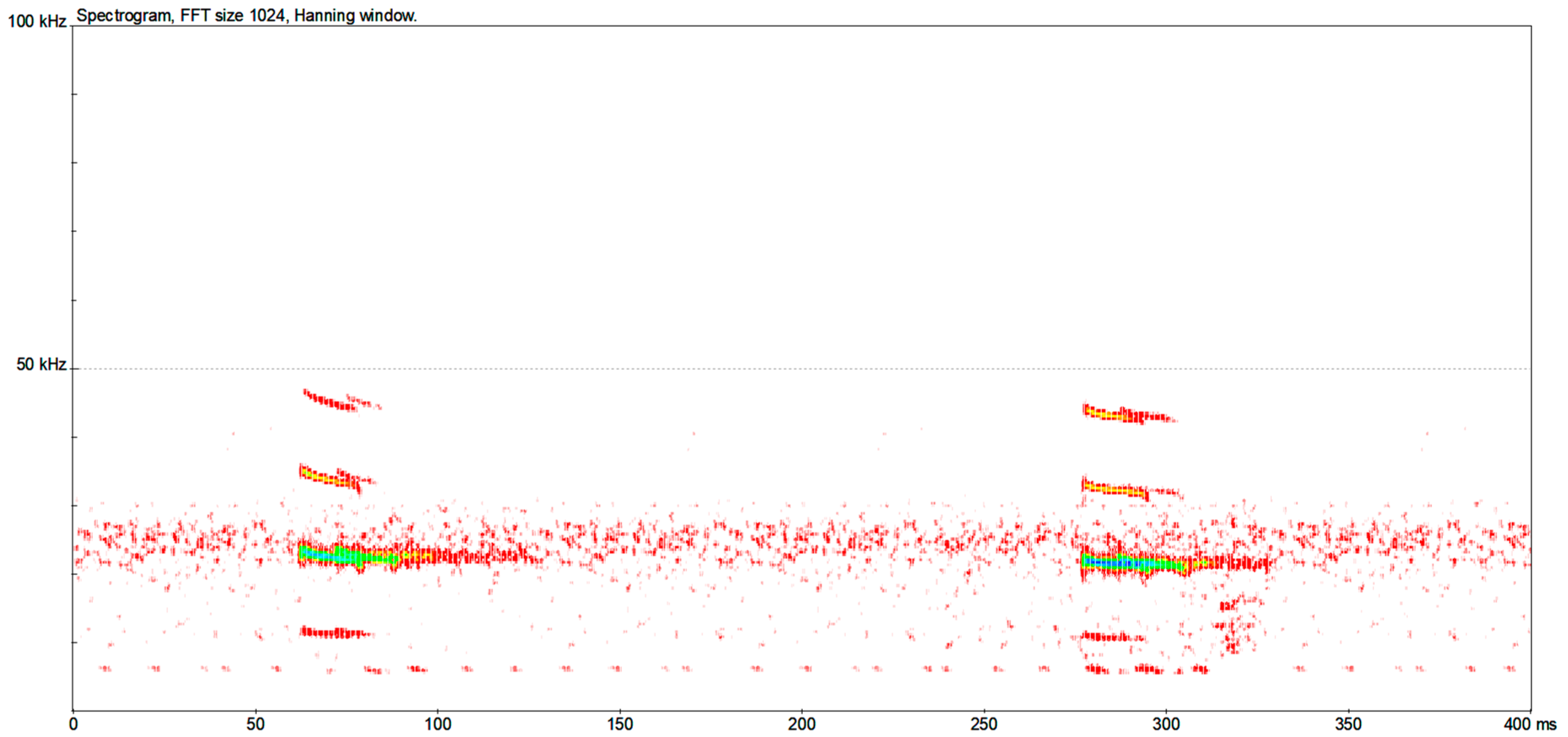
| Aselia tridens | CF/FM | 94 (94) | - | 105.22 ± 3.56 [93.4–112.3] | 118.90 ± 1.54 [113.0–122.3] | 9.47 ± 1.74 [5.7–12.8] | 48.60 ± 13.33 [23.8–89.2] |
| Tadarida teniotis | FM/QCF | 11 (11) | 13.20 ± 1.76 [11.4–16.7] | 11.71 ± 0.70 [11.1–12.8] | 12.16 ± 0.81 [11.3–13.0] | 14.25 ± 2.06 [10.7–17.3] | 772.13 ± 179.48 [634.9–1095.0] |
| Eptesicus isabellinus | FM/QCF | 109 (109) | 38.74 ± 8.21 [25.8–61.8] | 24.28 ± 1.01 [21.8–26.4] | 25.76 ± 1.03 [23.3–27.9] | 12.44 ± 2.63 [7.2–18.5] | 241.01 ± 74.08 [104.5–483.2] |
| Pipistrellus kuhlii | FM/QCF | 1940 (1940) | 59.60 ± 10.97 [39.7–101.6] | 42.13 ± 1.91 [36.8–48.4] | 42.91 ± 1.91 [36.9–49.5] | 6.10 ± 1.18 [3.6–11.8] | 181.81 ± 82.16 [8.4–716.5] |
| Otonycteris hemprichii | FM | 18 (18) | 46.68 ± 6.35 [30.7–55.9] | 17.72 ± 1.40 [15.7–18.9] | 22.97 ± 2.80 [18.9–27.7] | 6.49 ± 1.54 [4.0–8.4] | 200.92 ± 61.65 [112.2–313.9] |
| Plecotus gaisleri | FM | 5 (5) | 43.00 ± 4.27 [38.1–48.3] | 20.06 ± 3.46 [15.7–24.3] | 28.10 ± 1.20 [26.4–29.7] | 3.44 ± 0.66 [2.3–4.0] | 135.30 ± 42.12 [85.6–186.2] |
| Parameters | Mean ± SD | Range | Median | Q1 | Q3 | Jordan | Near East |
|---|---|---|---|---|---|---|---|
| SF (kHz) | 23.67 ± 1.09 | 21.8–26.3 | 23.4 | 23.2 | 23.6 | 26.1 ± 1.0 | 24.39 ± 1.55 |
| EF (kHz) | 22.42 ± 0.72 | 21.7–23.6 | 22.0 | 21.8 | 23.3 | 22.2 ± 0.6 | 22.80 ± 1.29 |
| FMAXE (kHz) | 23.17 ± 0.80 | 21.7–24.9 | 23.3 | 23.2 | 23.5 | 24.5 ± 0.5 | 23.38 ± 1.31 |
| D (ms) | 18.86 ± 3.32 | 13.1–26.4 | 18.5 | 16.6 | 20.6 | 15.4 ± 2.6 | 12.99 ± 3.50 |
| IPI (ms) | 420.17 ± 168.4 | 153.1–719.0 | 398.8 | 344.6 | 548.0 | 363.9 ± 115.1 | - |
| Oued Dkouk N. R. N = 541 | Ain Rhziza N = 143 | Bordj el Khadra Reed Bed N = 196 | Bordj el Khadra Oasis N = 1037 | |
|---|---|---|---|---|
| 58.66 ± 11.02 | 64.37 ± 13.28 | 58.99 ± 12.83 | 59.39 ± 9.96 | |
| SF (kHz) | 38.4–101.6 | 47.3–100.8 | 41.2–92.0 | 39.8–100.5 |
| 57.3 | 60.4 | 57.2 | 57.8 | |
| 40.36 ± 1.32 | 40.82 ± 1.75 | 42.65 ± 1.45 | 43.14 ± 1.48 | |
| EF (kHz) | 36.8–45.6 | 37.8–45.6 | 38.4–45.6 | 39.7–48.4 |
| 39.9 | 40.2 | 42.8 | 43.3 | |
| 41.19 ± 1.38 | 42.27 ± 1.75 | 43.41 ± 1.53 | 43.81 ± 1.58 | |
| FMAXE (kHz) | 36.9–46.3 | 38.6–46.3 | 39.9–47.3 | 39.7–49.5 |
| 41.3 | 41.8 | 42.8 | 43.4 | |
| 6.00 ± 1.14 | 5.70 ± 0.85 | 5.86 ± 1.36 | 6.25 ± 1.19 | |
| D (ms) | 4.0–11.8 | 4.0–9.5 | 4.0–11.0 | 3.6–10.6 |
| 5.9 | 5.7 | 5.6 | 6.2 | |
| 165.16 ± 76.28 | 138.42 ± 58.35 | 169.64 ± 76.18 | 198.76 ± 85.03 | |
| IPI (ms) | 8.4–563.5 | 42.4–275.6 | 61.3–716.5 | 56.5–528.6 |
| 169.5 | 111.5 | 184.2 | 195.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dalhoumi, R.; El Mokni, R.; Ouni, R.; Beyrem, H.; Aulagnier, S. Bats of the Tunisian Desert: Preliminary Data Using Acoustic Identification and First Record of Taphozous nudiventris in the Country. Diversity 2023, 15, 1108. https://doi.org/10.3390/d15111108
Dalhoumi R, El Mokni R, Ouni R, Beyrem H, Aulagnier S. Bats of the Tunisian Desert: Preliminary Data Using Acoustic Identification and First Record of Taphozous nudiventris in the Country. Diversity. 2023; 15(11):1108. https://doi.org/10.3390/d15111108
Chicago/Turabian StyleDalhoumi, Ridha, Ridha El Mokni, Ridha Ouni, Hamouda Beyrem, and Stéphane Aulagnier. 2023. "Bats of the Tunisian Desert: Preliminary Data Using Acoustic Identification and First Record of Taphozous nudiventris in the Country" Diversity 15, no. 11: 1108. https://doi.org/10.3390/d15111108
APA StyleDalhoumi, R., El Mokni, R., Ouni, R., Beyrem, H., & Aulagnier, S. (2023). Bats of the Tunisian Desert: Preliminary Data Using Acoustic Identification and First Record of Taphozous nudiventris in the Country. Diversity, 15(11), 1108. https://doi.org/10.3390/d15111108