
Species Diversity and Soil Interconstraints Exert Significant Influences on Plant Survival during Ecological Restoration in Semi-Arid Mining Areas

Abstract
:1. Introduction
2. Materials and Methods
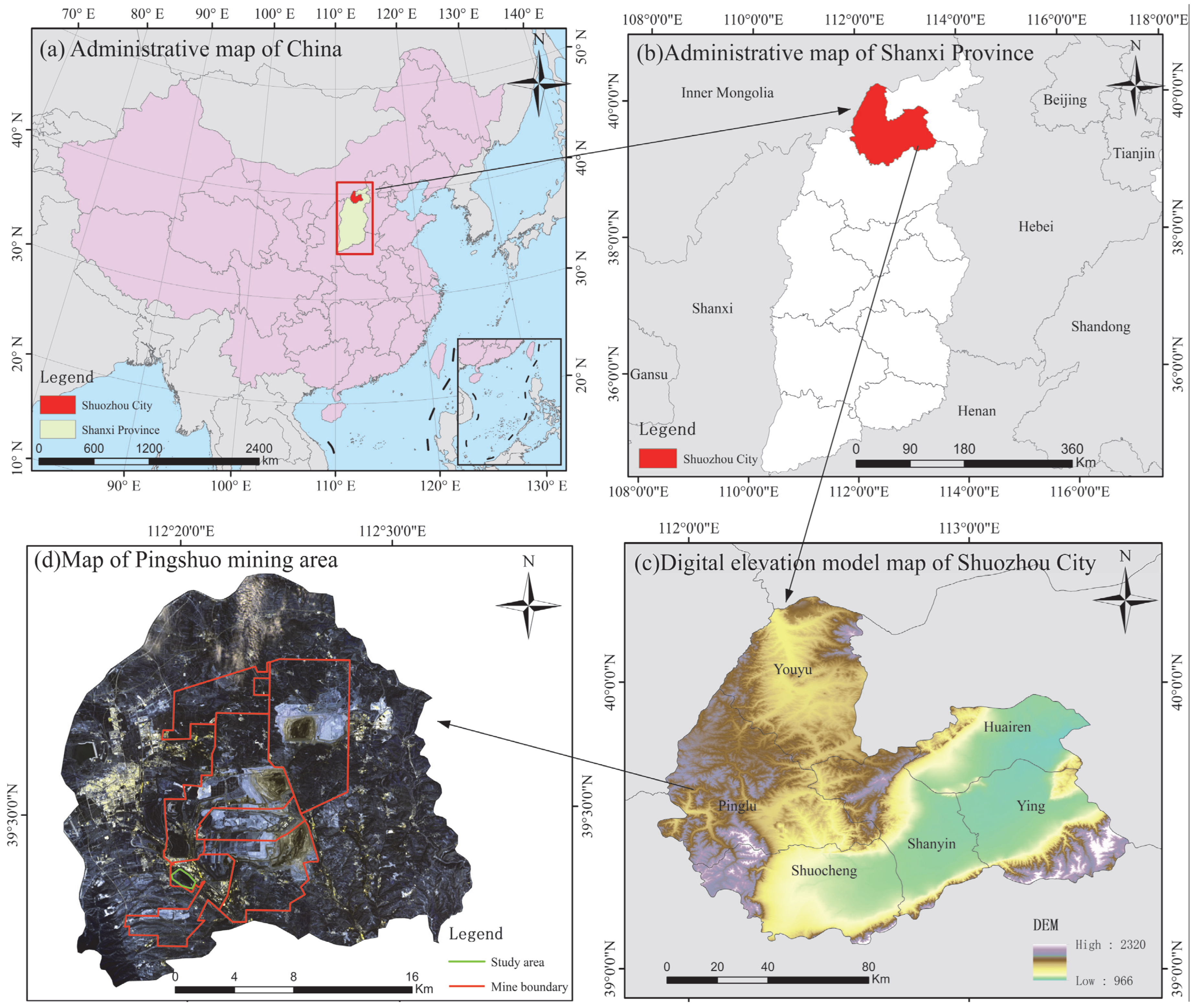
2.1. Study Sites
2.2. Sample Plot Setting
2.3. Sample Plot Survey
2.4. Statistical Analysis
3. Results
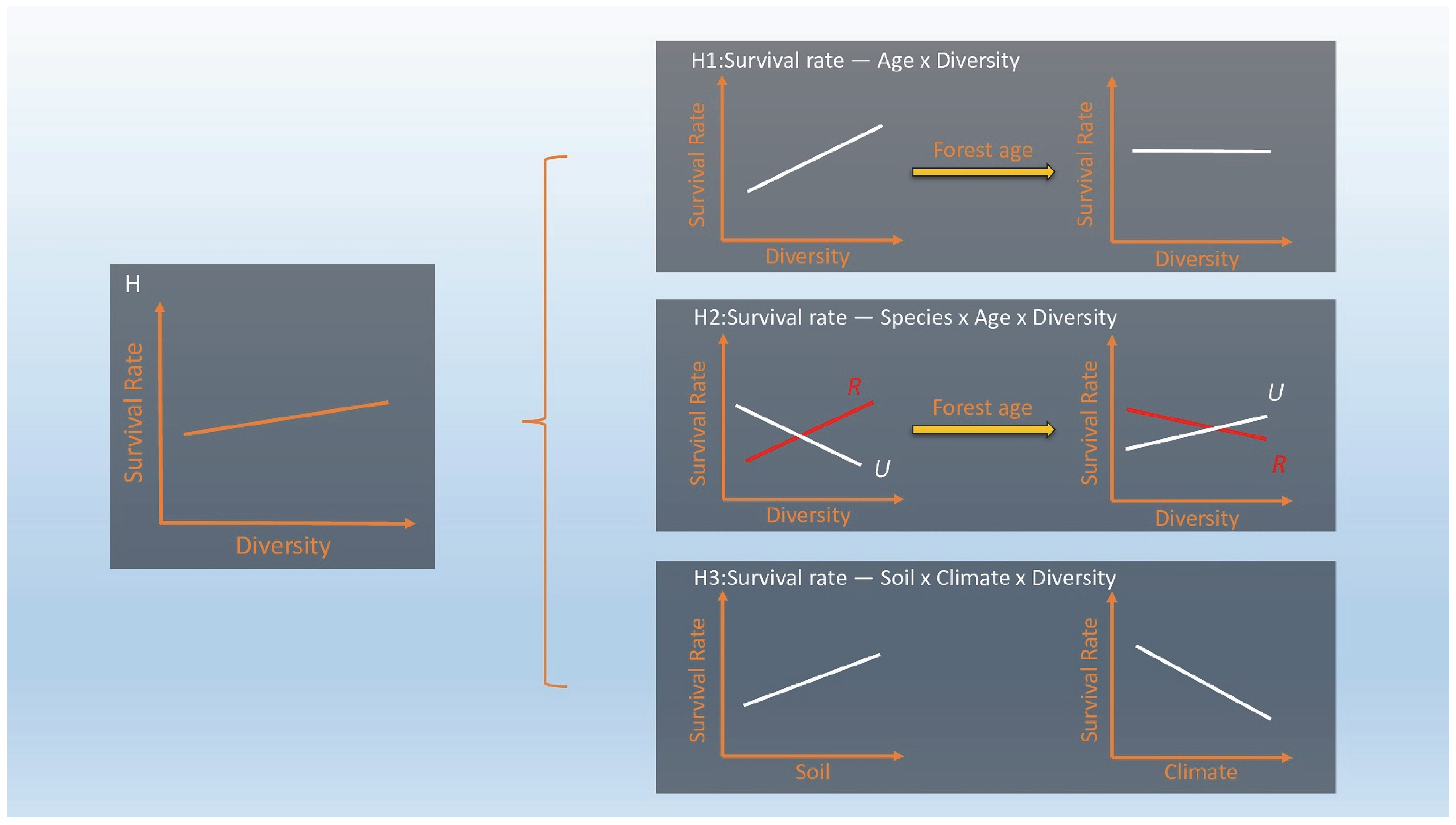
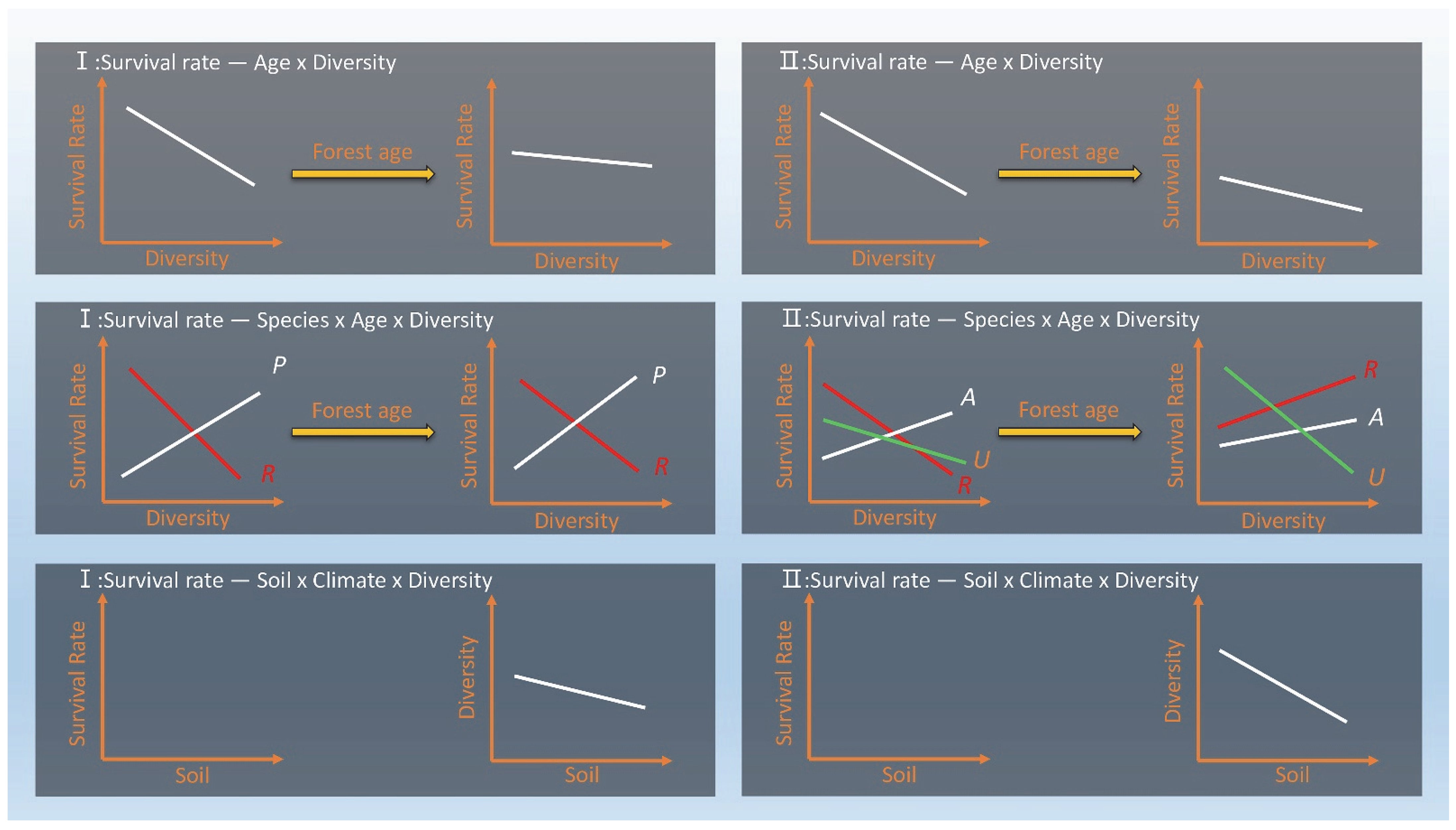
3.1. Survival Tends to Decrease with Stand Age and Diversity (H1)
3.2. Different Species Show Differential Survival Responses to Diversity and with Stand Age (H2)
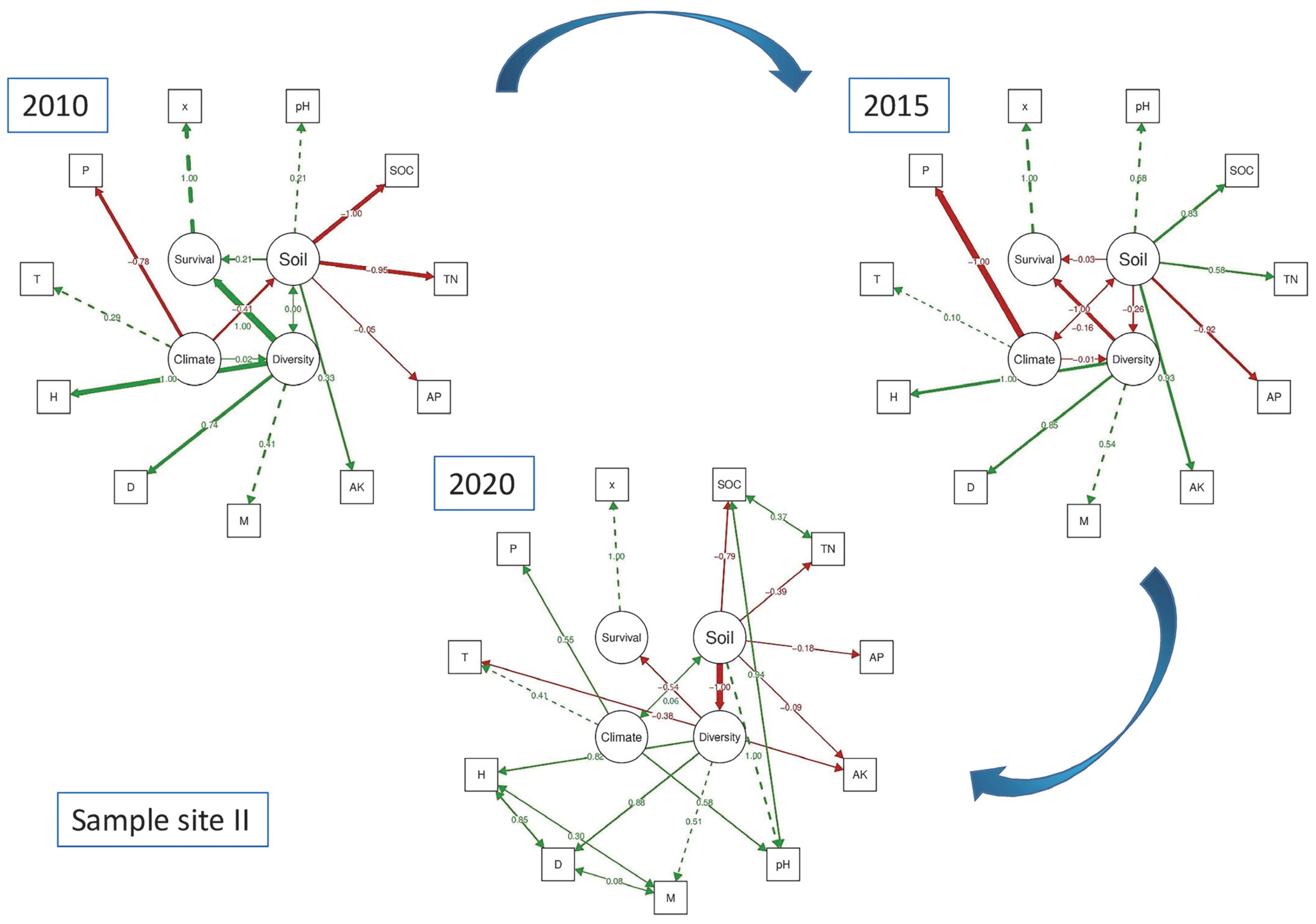
3.3. Different Survival Responses of Species to Diversity Always Depend on the Soil (H3)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, Q.; Zhao, B.; Guo, D. A review on vegetation restoration of opencast coal mine areas in northern China. Chin. J. Ecol. 2015, 34, 1152–1157. [Google Scholar] [CrossRef]
- Luo, Y.; Lü, Y.; Fu, B.; Zhang, Q.; Li, T.; Hu, W.; Comber, A. Half century change of interactions among ecosystem services driven by ecological restoration: Quantification and policy implications at a watershed scale in the Chinese Loess Plateau. Sci. Total Environ. 2019, 651, 2546–2557. [Google Scholar] [CrossRef]
- Rana, P.; Miller, D.C. Predicting the long-term social and ecological impacts of tree-planting programs: Evidence from northern India. World Dev. 2021, 140, 105367. [Google Scholar] [CrossRef]
- Brandt, M.; Hiernaux, P.; Rasmussen, K.; Mbow, C.; Kergoat, L.; Tagesson, T.; Ibrahim, Y.Z.; Wélé, A.; Tucker, C.J.; Fensholt, R. Assessing woody vegetation trends in Sahelian drylands using MODIS based seasonal metrics. Remote Sens. Environ. 2016, 183, 215–225. [Google Scholar] [CrossRef]
- Shen, Q.; Gao, G.; Han, F.; Xiao, F.; Ma, Y.; Wang, S.; Fu, B. Quantifying the effects of human activities and climate variability on vegetation cover change in a hyper-arid endorheic basin. Land Degrad. Dev. 2018, 29, 3294–3304. [Google Scholar] [CrossRef]
- Adman, B.; Wahyu Nugroho, A.; Yassir, I. The Growth of Local Tree Species on Post-Coal Mining Areas in East Kalimantan. Indones. J. For. Res. 2020, 7, 83–97. [Google Scholar] [CrossRef]
- Löf, M.; Dey, D.C.; Navarro, R.M.; Jacobs, D.F. Mechanical site preparation for forest restoration. New For. 2012, 43, 825–848. [Google Scholar] [CrossRef]
- Leibold, M.A.; Chase, J.M.; Ernest, S.K.M. Community assembly and the functioning of ecosystems: How metacommunity processes alter ecosystems attributes. Ecology 2017, 98, 909–919. [Google Scholar] [CrossRef]
- Hiroshima, T. Applying age-based mortality analysis to a natural forest stand in Japan. J. For. Res. 2014, 19, 379–387. [Google Scholar] [CrossRef]
- Óskarsson, H.; Sigurgeirsson, A.; Raulund-Rasmussen, K. Survival, growth, and nutrition of tree seedlings fertilized at planting on Andisol soils in Iceland: Six-year results. For. Ecol. Manag. 2006, 229, 88–97. [Google Scholar] [CrossRef]
- Osunkoya, O.O.; Othman, F.E.; Kahar, R.S. Growth and competition between seedlings of an invasive plantation tree, Acacia mangium, and those of a native Borneo heath-forest species, Melastoma beccarianum. Ecol. Res. 2005, 20, 205–214. [Google Scholar] [CrossRef]
- Hattori, D.; Kenzo, T.; Yamauchi, N.; Irino, K.O.; Kendawang, J.J.; Ninomiya, I.; Sakurai, K. Effects of environmental factors on growth and mortality of Parashorea macrophylla (Dipterocarpaceae) planted on slopes and valleys in a degraded tropical secondary forest in Sarawak, Malaysia. Soil Sci. Plant Nutr. 2013, 59, 218–228. [Google Scholar] [CrossRef]
- Van de Peer, T.; Verheyen, K.; Baeten, L.; Ponette, Q.; Muys, B.; Firn, J. Biodiversity as insurance for sapling survival in experimental tree plantations. J. Appl. Ecol. 2016, 53, 1777–1786. [Google Scholar] [CrossRef]
- Woodall, C.W.; Grambsch, P.L.; Thomas, W. Applying survival analysis to a large-scale forest inventory for assessment of tree mortality in Minnesota. Ecol. Model. 2005, 189, 199–208. [Google Scholar] [CrossRef]
- Bai, X.; Queenborough, S.A.; Wang, X.; Zhang, J.; Li, B.; Yuan, Z.; Xing, D.; Lin, F.; Ye, J.; Hao, Z. Effects of local biotic neighbors and habitat heterogeneity on tree and shrub seedling survival in an old-growth temperate forest. Oecologia 2012, 170, 755–765. [Google Scholar] [CrossRef]
- Frouz, J.; Dvorščík, P.; Vávrová, A.; Doušová, O.; Kadochová, Š.; Matějíček, L. Development of canopy cover and woody vegetation biomass on reclaimed and unreclaimed post-mining sites. Ecol. Eng. 2015, 84, 233–239. [Google Scholar] [CrossRef]
- Ahirwal, J.; Maiti, S.K.; Satyanarayana Reddy, M. Development of carbon, nitrogen and phosphate stocks of reclaimed coal mine soil within 8 years after forestation with Prosopis juliflora (Sw.) Dc. CATENA 2017, 156, 42–50. [Google Scholar] [CrossRef]
- Abebe, G.; Tsunekawa, A.; Haregeweyn, N.; Taniguchi, T.; Wondie, M.; Adgo, E.; Masunaga, T.; Tsubo, M.; Ebabu, k.; Mamedov, A.; et al. Effect of Soil Microbiome from Church Forest in the Northwest Ethiopian Highlands on the Growth of Olea europaea and Albizia gummifera Seedlings under Glasshouse Conditions. Sustainability 2020, 12, 4976. [Google Scholar] [CrossRef]
- Maloney, P.E.; Vogler, D.R.; Jensen, C.E.; Mix, A.D. Ecology of whitebark pine populations in relation to white pine blister rust infection in subalpine forests of the Lake Tahoe Basin, USA: Implications for restoration. For. Ecol. Manag. 2012, 280, 166–175. [Google Scholar] [CrossRef]
- Comita, L.S.; Engelbrecht, B.M. Seasonal and Spatial Variation in Water Availability Drive Habitat Associations in a Tropical Forest. Ecology 2009, 90, 2755–2765. [Google Scholar] [CrossRef]
- Navarro-Cerrillo, R.M.; Rodriguez-Vallejo, C.; Silveiro, E.; Hortal, A.; Palacios-Rodríguez, G.; Duque-Lazo, J.; Camarero, J.J. Cumulative Drought Stress Leads to a Loss of Growth Resilience and Explains Higher Mortality in Planted than in Naturally Regenerated Pinus pinaster Stands. Forests 2018, 9, 358. [Google Scholar] [CrossRef]
- Spathelf, P.; van der Maaten, E.; van der Maaten-Theunissen, M.; Campioli, M.; Dobrowolska, D. Climate change impacts in European forests: The expert views of local observers. Ann. For. Sci. 2014, 71, 131–137. [Google Scholar] [CrossRef]
- Tong, C.; Wu, J.; Yong, S.; Yang, J.; Yong, W. A landscape-scale assessment of steppe degradation in the Xilin River Basin, Inner Mongolia, China. J. Arid Environ. 2004, 59, 133–149. [Google Scholar] [CrossRef]
- Wang, X.; Guo, X.; Du, N.; Guo, W.; Pang, J. Rapid nitrogen fixation contributes to a similar growth and photosynthetic rate of Robinia pseudoacacia supplied with different levels of nitrogen. Tree Physiol. 2021, 41, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Lim, J.Y.; Yang, J.; Luskin, M.S. When do Janzen-Connell effects matter? A phylogenetic meta-analysis of conspecific negative distance and density dependence experiments. Ecol. Lett. 2021, 24, 608–620. [Google Scholar] [CrossRef]
- Searle, E.B.; Chen, H.Y.; Paquette, A. Higher tree diversity is linked to higher tree mortality. Proc. Natl. Acad. Sci. USA 2022, 119, e2013171119. [Google Scholar] [CrossRef]
- Garcia, L.C.; Hobbs, R.J.; Ribeiro, D.B.; Tamashiro, J.Y.; Santos, F.A.M.; Rodrigues, R.R.; Marrs, R. Restoration over time: Is it possible to restore trees and non-trees in high-diversity forests? Appl. Veg. Sci. 2016, 19, 655–666. [Google Scholar] [CrossRef]
- Chen, Y.; Cao, Y. Response of tree regeneration and understory plant species diversity to stand density in mature Pinus tabulaeformis plantations in the hilly area of the Loess Plateau, China. Ecol. Eng. 2014, 73, 238–245. [Google Scholar] [CrossRef]
- Thijs, K.W.; Aerts, R.; Van de Moortele, P.; Musila, W.; Gulinck, H.; Muys, B. Contrasting Cloud Forest Restoration Potential Between Plantations of Different Exotic Tree Species. Restor. Ecol. 2014, 22, 472–479. [Google Scholar] [CrossRef]
- Yachi, S.; Loreau, M. Biodiversity and ecosystem productivity in a fluctuating environment: The insurance hypothesis. Proc. Natl. Acad. Sci. USA 1999, 96, 1463–1468. [Google Scholar] [CrossRef]
- Tilman, D.; Isbell, F.; Cowles, J.M. Biodiversity and Ecosystem Functioning. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 471–493. [Google Scholar] [CrossRef]
- Benayas, J.M.R.; Newton, A.C.; Diaz, A.; Bullock, J.M. Enhancement of biodiversity and ecosystem services by ecological restoration: A meta-analysis. Science 2009, 325, 1121–1124. [Google Scholar] [CrossRef]
- Hutto, R.L.; Belote, R.T. Distinguishing four types of monitoring based on the questions they address. For. Ecol. Manag. 2013, 289, 183–189. [Google Scholar] [CrossRef]
- Hao, F.; Zhang, X.; Ouyang, W.; Skidmore, A.K.; Toxopeus, A.G. Vegetation NDVI Linked to Temperature and Precipitation in the Upper Catchments of Yellow River. Environ. Model. Assess. 2011, 17, 389–398. [Google Scholar] [CrossRef]
- Zhao, B.; Guo, D.; Bai, Z.; Zhao, Z. Community dynamics of artificial vegetation in a reclaimed spoil from a semi-arid open-cast coal mine in 2010–2015. Chin. J. Ecol. 2018, 37, 1636–1644. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps, 4th ed.; International Union of Soil Sciences (IUSS): Vienna, Austria, 2022; 234p. [Google Scholar]
- Shi, Z.; Bai, Z.; Guo, D.; Chen, M. Develop a Soil Quality Index to Study the Results of Black Locust on Soil Quality below Different Allocation Patterns. Land 2021, 10, 785. [Google Scholar] [CrossRef]
- Keylock, C.J. Simpson diversity and the Shannon–Wiener index as special cases of a generalized entropy. Oikos 2005, 109, 203–207. [Google Scholar] [CrossRef]
- Fan, Y.; Chen, J.; Shirkey, G.; John, R.; Wu, S.R.; Park, H.; Shao, C. Applications of structural equation modeling (SEM) in ecological studies: An updated review. Ecol. Process. 2016, 5, 19. [Google Scholar] [CrossRef]
- Grace, J.B.; Irvine, K.M. Scientist’s guide to developing explanatory statistical models using causal analysis principles. Ecology 2019, 101, e02962. [Google Scholar] [CrossRef]
- Rosseel, Y. Lavaan: An R package for structural equation modeling. J. Stat. Softw. 2012, 48, 1–36. [Google Scholar] [CrossRef]
- Cusack, D.; Montagnini, F. The role of native species plantations in recovery of understory woody diversity in degraded pasturelands of Costa Rica. For. Ecol. Manag. 2004, 188, 1–15. [Google Scholar] [CrossRef]
- Collet, C.; Le Moguedec, G. Individual seedling mortality as a function of size, growth and competition in naturally regenerated beech seedlings. Forestry 2007, 80, 359–370. [Google Scholar] [CrossRef]
- Laughlin, D.C.; Strahan, R.T.; Adler, P.B.; Moore, M.M. Survival rates indicate that correlations between community-weighted mean traits and environments can be unreliable estimates of the adaptive value of traits. Ecol. Lett. 2018, 21, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Chen, Y.; Castro-Izaguirre, N.; Baruffol, M.; Brezzi, M.; Lang, A.; Härdtle, W.; von Oheimb, G.; Yang, X.; Liu, X.; et al. Impacts of species richness on productivity in a large-scale subtropical forest experiment. Science 2018, 362, 80–83. [Google Scholar] [CrossRef]
- Grossman, J.J.; Vanhellemont, M.; Barsoum, N.; Bauhus, J.; Bruelheide, H.; Castagneyrol, B.; Cavender-Bares, J.; Eisenhauer, N.; Ferlian, O.; Gravel, D.; et al. Synthesis and future research directions linking tree diversity to growth, survival, and damage in a global network of tree diversity experiments. Environ. Exp. Bot. 2018, 152, 68–89. [Google Scholar] [CrossRef]
- De Sá, N.C.; Marchante, H.; Marchante, E.; Cabral, J.A.; Honrado, J.P.; Vicente, J.R. Can citizen science data guide the surveillance of invasive plants? A model-based test with Acacia trees in Portugal. Biol. Invasions 2019, 21, 2127–2141. [Google Scholar] [CrossRef]
- Shure, D.J.; Phillips, D.L.; Edward Bostick, P. Gap size and succession in cutover southern Appalachian forests: An 18 year study of vegetation dynamics. Plant Ecol. 2006, 185, 299–318. [Google Scholar] [CrossRef]
- Zhu, J.; Wu, A.; Zou, S.; Xiong, X.; Liu, S.; Chu, G.; Zhang, Q.; Liu, J.; Tang, X.; Yan, J.; et al. Relationships between tree diversity and biomass/productivity and their influence factors in a lower subtropical evergreen broad-leaved forest. Biodivers. Sci. 2021, 29, 1435–1446. [Google Scholar] [CrossRef]
- Huston, M.A. Hidden treatments in ecological experiments: Re-evaluating the ecosystem function of biodiversity. Oecologia 1997, 110, 449–460. [Google Scholar] [CrossRef]
- Vogt, D.R.; Murrell, D.J.; Stoll, P. Testing spatial theories of plant coexistence: No consistent differences in intra- and interspecific interaction distances. Am. Nat. 2010, 175, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Eilts, J.A.; Mittelbach, G.G.; Reynolds, H.L.; Gross, K.L. Resource heterogeneity, soil fertility, and species diversity: Effects of clonal species on plant communities. Am. Nat. 2011, 177, 574–588. [Google Scholar] [CrossRef]
- Gazol, A.; Tamme, R.; Price, J.N.; Hiiesalu, I.; Laanisto, L.; Partel, M. A negative heterogeneity-diversity relationship found in experimental grassland communities. Oecologia 2013, 173, 545–555. [Google Scholar] [CrossRef]
- Schoolmaster, D.R. Resource competition and coexistence in heterogeneous metacommunities: Many-species coexistence is unlikely to be facilitated by spatial variation in resources. PeerJ 2013, 1, e136. [Google Scholar] [CrossRef]
- Price, J.; Tamme, R.; Gazol, A.; de Bello, F.; Takkis, K.; Uria-Diez, J.; Kasari, L.; Pärtel, M. Within-community environmental variability drives trait variability in species-rich grasslands. J. Veg. Sci. 2017, 28, 303–312. [Google Scholar] [CrossRef]
- Williams, B.M.; Houseman, G.R. Experimental evidence that soil heterogeneity enhances plant diversity during community assembly. J. Plant Ecol. 2014, 7, 461–469. [Google Scholar] [CrossRef]
- Kraft, N.J.B.; Adler, P.B.; Godoy, O.; James, E.C.; Fuller, S.; Levine, J.M. Community assembly, coexistence and the environmental filtering metaphor. Funct. Ecol. 2015, 29, 592–599. [Google Scholar] [CrossRef]
- Aerts, R.; Chapin, F.S. The Mineral Nutrition of Wild Plants Revisited: A Re-evaluation of Processes and Patterns. Adv. Ecol. Res. 1999, 30, 1–67. [Google Scholar]
- Tang, Z.; Xu, W.; Zhou, G.; Bai, Y.; Li, J.; Tang, X.; Chen, D.; Liu, Q.; Ma, W.; Xiong, G.; et al. Patterns of plant carbon, nitrogen, and phosphorus concentration in relation to productivity in China’s terrestrial ecosystems. Proc. Natl. Acad. Sci. USA 2018, 115, 4033–4038. [Google Scholar] [CrossRef] [PubMed]
- Coomes, D.A.; Bentley, W.A.; Tanentzap, A.J.; Burrows, L.E. Soil drainage and phosphorus depletion contribute to retrogressive succession along a New Zealand chronosequence. Plant Soil 2013, 367, 77–91. [Google Scholar] [CrossRef]
- Deng, Q.; McMahon, D.E.; Xiang, Y.; Yu, C.L.; Jackson, R.B.; Hui, D. A global meta-analysis of soil phosphorus dynamics after afforestation. New Phytol. 2017, 213, 181–192. [Google Scholar] [CrossRef]
- Cao, S.; Chen, L.; Xu, C.; Liu, Z. Impact of three soil types on afforestation in China’s Loess Plateau: Growth and survival of six tree species and their effects on soil properties. Landsc. Urban Plan 2007, 83, 208–217. [Google Scholar] [CrossRef]
- Brooker, R.W.; Maestre, F.T.; Callaway, R.M.; Lortie, C.L.; Cavieres, L.A.; Kunstler, G.; Liancourt, P.; Tielbörger, K.; Travis, J.M.J.; Anthelme, F.; et al. Facilitation in plant communities: The past, the present, and the future. J. Ecol. 2008, 96, 18–34. [Google Scholar] [CrossRef]
- Devaney, J.L.; Pullen, J.; Cook-Patton, S.C.; Burghardt, K.T.; Parker, J.D. Tree diversity promotes growth of late successional species despite increasing deer damage in a restored forest. Ecology 2020, 101, e03063. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Parameter Interpretation |
|---|---|
| S | the total number of species |
| N | the total number of individuals of each species |
| InN | the natural logarithm of individuals |
| pi | the proportion of individuals of the ith species to all individuals |
| Stand Age/Year | Total Survival Rate/% | M | D | H’ |
|---|---|---|---|---|
| 0 | 100 | 0.300 | 1.237 | 0.340 |
| 17 | 58.66 | 0.412 | 1.584 | 0.711 |
| 22 | 57.23 | 0.551 | 1.812 | 0.794 |
| 27 | 57.01 | 0.631 | 2.245 | 0.913 |
| Stand Age/Year | Total Survival Rate/% | M | D | H’ |
|---|---|---|---|---|
| 0 | 100 | 0.435 | 3.000 | 1.099 |
| 17 | 33.02 | 0.988 | 2.393 | 0.988 |
| 22 | 31.73 | 0.993 | 2.673 | 1.054 |
| 27 | 47 | 0.592 | 1.784 | 0.756 |
| Survival Rate | Stand Age/Year | M | D | H’ |
|---|---|---|---|---|
| R | 17 | −0.508 ** | −0.507 ** | −0.573 ** |
| P | 0.598 ** | 0.609 ** | 0.701 ** | |
| Total | −0.349 ** | −0.345 ** | −0.385 ** | |
| R | 22 | −0.421 ** | −0.473 ** | −0.517 ** |
| P | 0.584 ** | 0.729 ** | 0.727 ** | |
| Total | −0.263 * | −0.272 * | −0.320 ** | |
| R | 27 | −0.531 ** | −0.438 ** | −0.540 ** |
| P | 0.786 ** | 0.686 ** | 0.796 ** | |
| Total | −0.311 ** | −0.243 * | −0.319 ** |
| Survival Rate | Stand Age/Year | M | D | H’ |
|---|---|---|---|---|
| R | 17 | −0.333 ** | −0.504 ** | −0.427 ** |
| U | −0.229 * | 0.149 | 0.158 | |
| A | 0.171 | 0.621 ** | 0.618 ** | |
| Total | −0.364 ** | 0.002 | 0.059 | |
| R | 22 | −0.125 | −0.125 | 0.009 |
| U | −0.298 ** | −0.317 ** | −0.311 ** | |
| A | 0.038 | 0.575 ** | 0.584 ** | |
| Total | −0.355 ** | −0.214 * | −0.133 | |
| R | 27 | −0.068 | 0.442 ** | 0.433 ** |
| U | −0.310 ** | −0.638 ** | −0.609 ** | |
| A | 0.258 ** | 0.539 ** | 0.567 ** | |
| Total | −0.301 ** | −0.469 ** | −0.437 ** |
| Site I | Stand Age/Year | pH | SOC (g/kg) | TN (g/kg) | AP (mg/kg) | AK (mg/kg) | T | P |
|---|---|---|---|---|---|---|---|---|
| Total survival | 17 | −0.020 | −0.261 * | −0.244 * | 0.285 * | 0.278 * | −0.143 | −0.024 |
| Total survival | 22 | 0.372 ** | −0.235 * | −0.143 | 0.022 | −0.257 * | 0.034 | −0.074 |
| Total survival | 27 | 0.039 | 0.134 | 0.117 | −0.039 | 0.014 | −0.218 | −0.217 |
| M | 17 | 0.023 | −0.087 | −0.092 | −0.111 | −0.028 | 0.018 | 0.044 |
| D | 0.080 | −0.097 | −0.105 | −0.039 | −0.042 | 0.045 | 0.006 | |
| H’ | 0.053 | −0.102 | −0.110 | −0.112 | −0.009 | 0.005 | −0.016 | |
| M | 22 | −0.095 | 0.109 | −0.009 | −0.177 | 0.092 | 0.169 | 0.023 |
| D | −0.095 | 0.103 | −0.065 | −0.153 | 0.065 | 0.052 | −0.024 | |
| H’ | −0.070 | 0.105 | −0.058 | −0.142 | 0.105 | 0.048 | −0.047 | |
| M | 27 | 0.045 | −0.191 | −0.104 | 0.164 | −0.136 | 0.021 | 0.058 |
| D | −0.026 | −0.020 | 0.069 | 0.181 | −0.036 | −0.011 | 0.021 | |
| H’ | −0.020 | −0.077 | 0.027 | 0.147 | −0.080 | 0.014 | 0.029 |
| Site II | Stand Age/Year | pH | SOC (g/kg) | TN (g/kg) | AP (mg/kg) | AK (mg/kg) | T | P |
|---|---|---|---|---|---|---|---|---|
| Total survival | 17 | 0.212 * | −0.246 * | −0.230 * | 0.035 | −0.009 | 0.246 | −0.329 |
| Total survival | 22 | 0.125 | 0.057 | −0.100 | −0.130 | 0.081 | 0.029 | −0.277 |
| Total survival | 27 | 0.228 * | −0.024 | −0.033 | −0.075 | −0.061 | −0.212 | 0.224 |
| M | 17 | −0.154 | 0.068 | 0.061 | −0.019 | 0.023 | 0.056 | 0.109 |
| D | 0.056 | −0.115 | −0.124 | −0.141 | 0.060 | 0.070 | 0.036 | |
| H’ | 0.091 | −0.070 | −0.074 | −0.116 | 0.094 | 0.108 | 0.085 | |
| M | 22 | −0.111 | −0.152 | −0.071 | 0.117 | −0.159 | 0.052 | 0.223 * |
| D | −0.039 | −0.255 * | −0.237 * | 0.117 | −0.174 | 0.121 | 0.160 | |
| H’ | −0.019 | −0.280 ** | −0.268 ** | 0.104 | −0.179 | 0.139 | 0.189 | |
| M | 27 | −0.187 | −0.088 | 0.113 | 0.056 | −0.102 | 0.106 | −0.024 |
| D | −0.187 | −0.048 | −0.036 | −0.253* | 0.012 | 0.086 | 0.168 | |
| H’ | −0.182 | −0.062 | −0.019 | −0.225 * | −0.021 | 0.087 | 0.142 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, Z.; Bai, Z.; Guo, D.; Li, S.; Chen, M. Species Diversity and Soil Interconstraints Exert Significant Influences on Plant Survival during Ecological Restoration in Semi-Arid Mining Areas. Diversity 2023, 15, 1100. https://doi.org/10.3390/d15101100
Shi Z, Bai Z, Guo D, Li S, Chen M. Species Diversity and Soil Interconstraints Exert Significant Influences on Plant Survival during Ecological Restoration in Semi-Arid Mining Areas. Diversity. 2023; 15(10):1100. https://doi.org/10.3390/d15101100
Chicago/Turabian StyleShi, Zeyu, Zhongke Bai, Donggang Guo, Shuai Li, and Meijing Chen. 2023. "Species Diversity and Soil Interconstraints Exert Significant Influences on Plant Survival during Ecological Restoration in Semi-Arid Mining Areas" Diversity 15, no. 10: 1100. https://doi.org/10.3390/d15101100
APA StyleShi, Z., Bai, Z., Guo, D., Li, S., & Chen, M. (2023). Species Diversity and Soil Interconstraints Exert Significant Influences on Plant Survival during Ecological Restoration in Semi-Arid Mining Areas. Diversity, 15(10), 1100. https://doi.org/10.3390/d15101100