





Stability of C3 and C4 Grass Patches in Woody Encroached Rangeland after Fire and Simulated Grazing

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Fire Treatments
2.3. Patch Identification and Clip Treatments
2.4. Analysis
3. Results
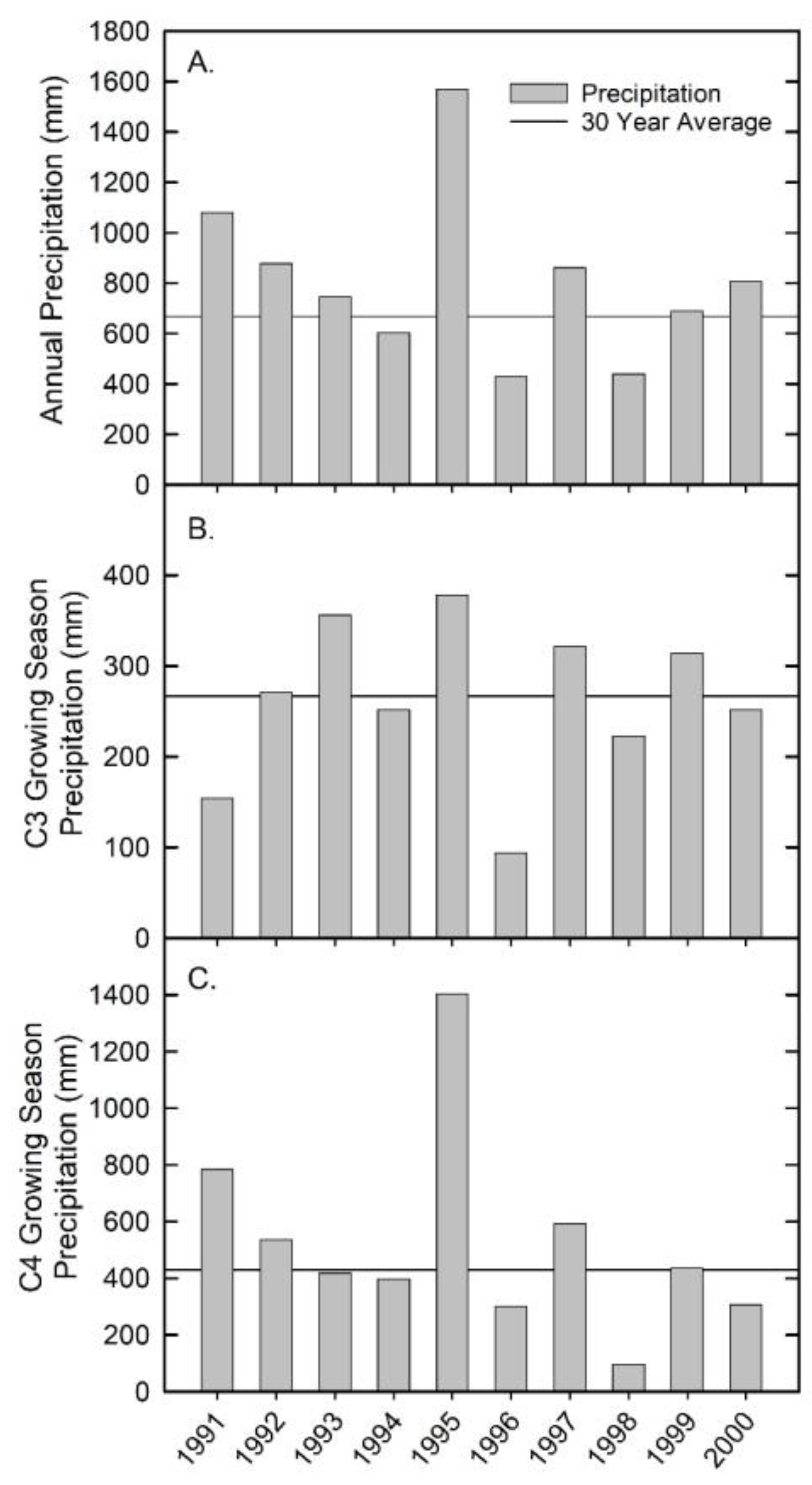
3.1. Precipitation
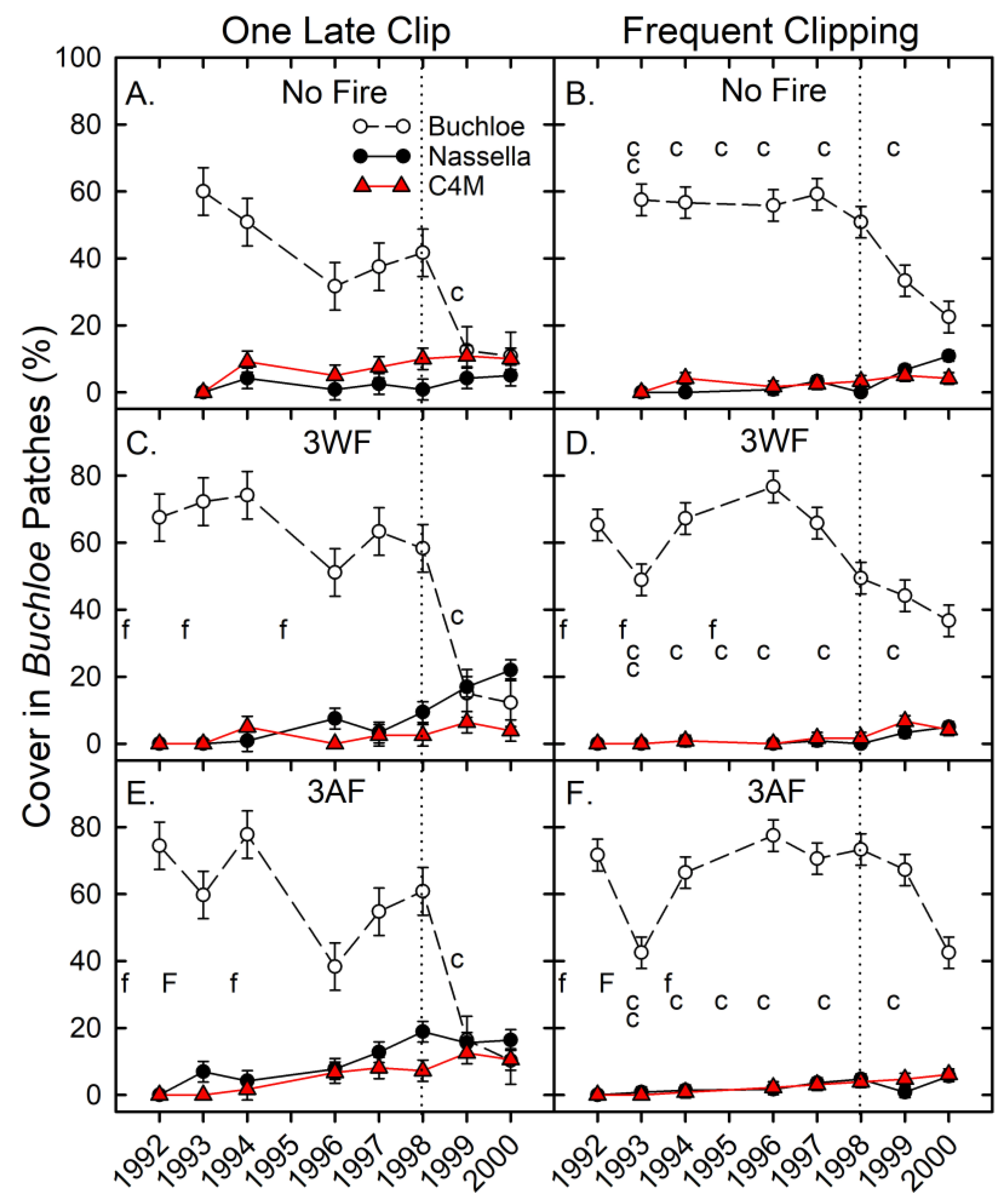
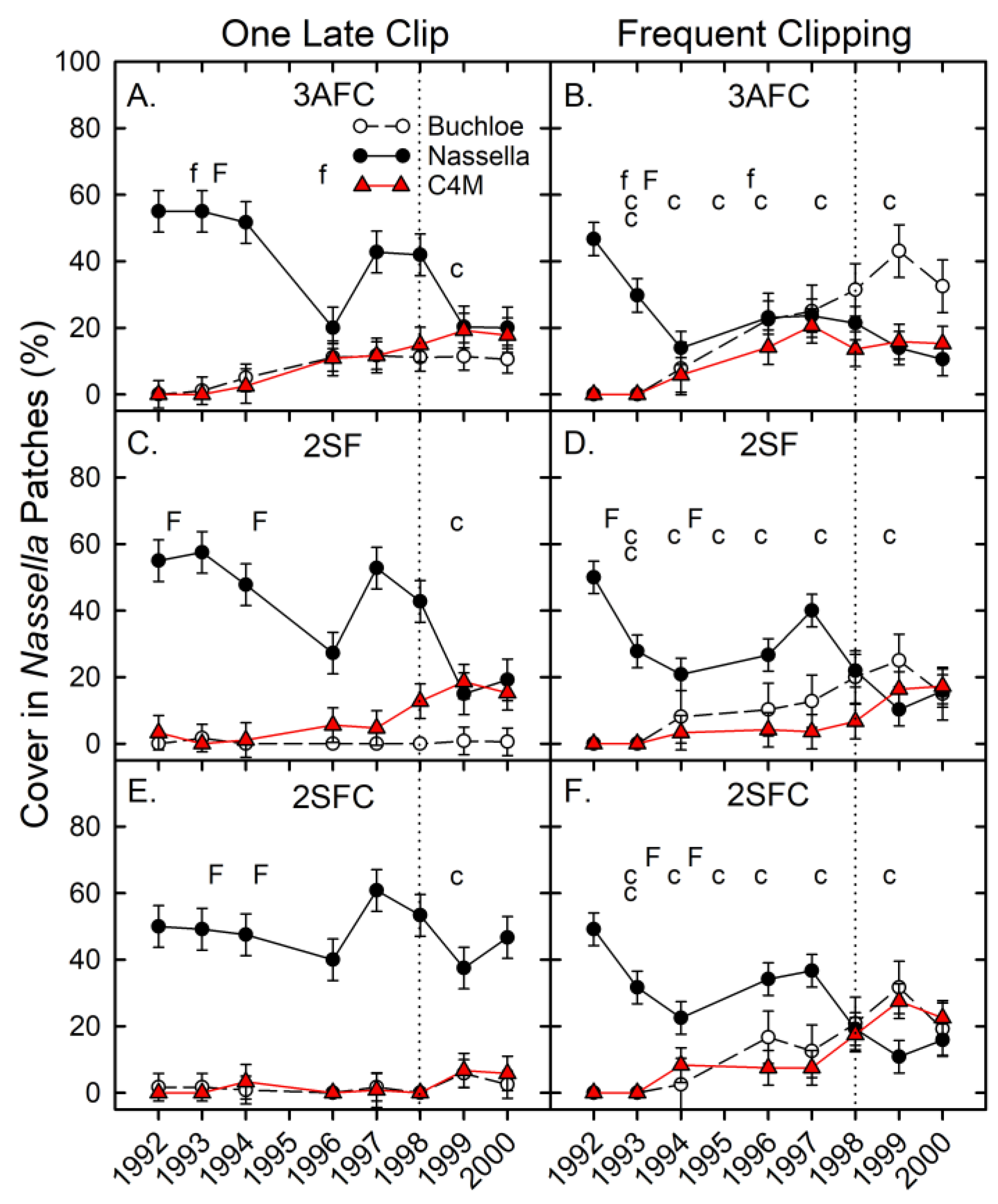
3.2. Phase 1—Herbaceous Cover Responses
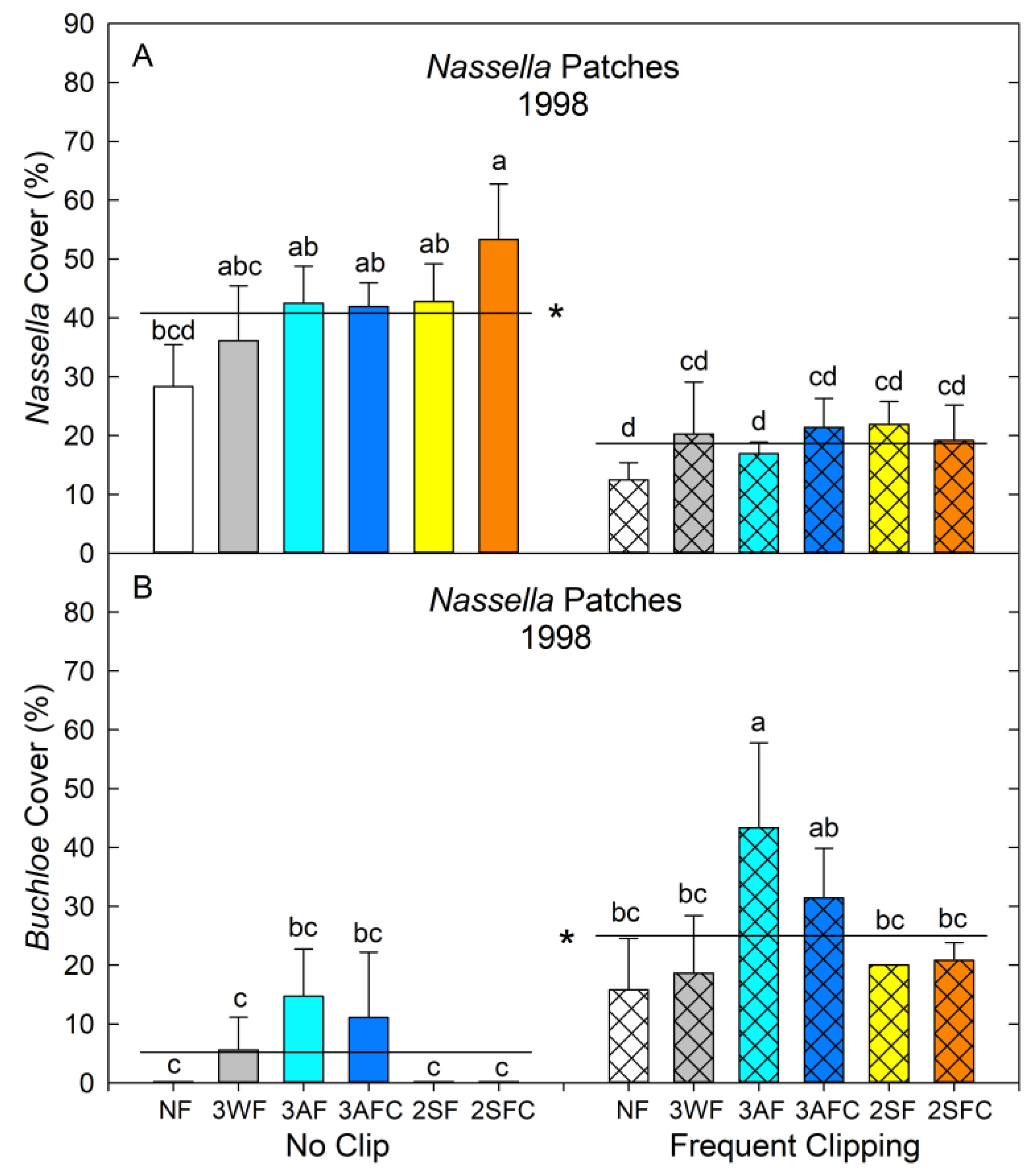
3.3. End of Phase 1 Herbaceous Cover (1998)
3.4. Phase 2—Herbaceous Cover Responses
4. Discussion
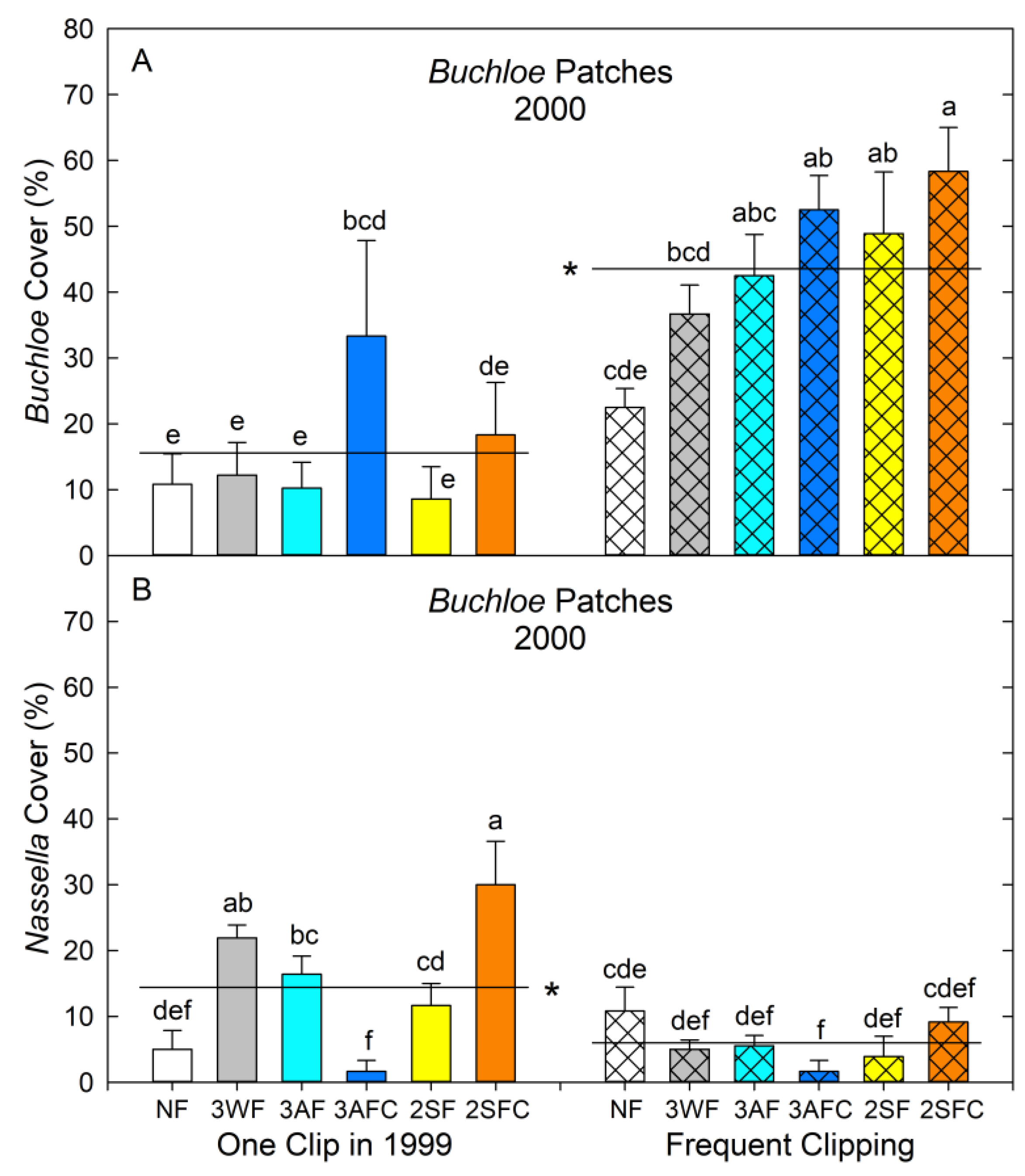
4.1. Responses in Buchloe Patches
4.2. Responses in Nassella Patches
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Scientific Name and Author(s) | Common Name |
|---|---|---|
| C3 annual grass | Bromus japonicus Houtt. | Japanese brome |
| Hordeum pusillum Nutt. | Little barley | |
| C3 mid-grass | Nassella leucotricha (Trin. and Rupr.) R.W. Pohl. | Texas wintergrass |
| Elymus smithii (Rydb.) Gould 1 | Western wheatgrass | |
| Poa arachnifera Torr. | Texas bluegrass | |
| C4 short-grass | Buchloe dactyloides (Nutt.) Engelm. 2 | Buffalograss |
| Bouteloua hirsuta Lag. | Hairy grama | |
| Bouteloua rigidiseta (Steud.) Hitchc. | Texas grama | |
| Chloris verticillata Nutt. | Tumble windmillgrass | |
| Schedonnardus paniculatus (Nutt.) Trel. | Tumblegrass | |
| C4 mid-grass bunchgrass | Aristida purpurea Nutt. | Purple threeawn |
| Bothriochloa laguroides (DC.) Herter | Silver bluestem | |
| Bouteloua curtipendula (Michx.) Torr. | Sideoats grama | |
| Bouteloua gracilis (Kunth) Lag. ex Griffiths 3 | Blue grama | |
| Digitaria californica [Benth.] Henrard | Arizona cottontop | |
| Eragrostis intermedia Hitchc. | Plains lovegrass | |
| Eragrostis secundiflora J. Presl | Red lovegrass | |
| Eriochloa sericea (Scheele) Munro ex Vasey | Texas cupgrass | |
| Leptochloa dubia (Kunth) Nees | Green sprangletop | |
| Panicum virgatum L. | Switchgrass | |
| Sporobolus giganteus Nash | Giant dropseed | |
| Sporobolus. | Tall dropseed | |
| Sporobolus compositus (Poir.) Merrcryptandrus (Torr.) A. Gray | Sand dropseed | |
| Tridens albescens (Vasey) Wooton and Standl. | White tridens | |
| Tripsacum dactyloides L. | Eastern gamagrass | |
| C4 mid-grass rhizomatous | Hilaria mutica (Buckley) Benth. 4 | Tobosagrass |
| Panicum obtusum Kunth | Vinemesquite |
References
- Lauenroth, W.K.; Burke, I.C.; Gutmann, M.P. The structure and function of ecosystems in the Central North American grassland region. Great Plains Res. 1999, 9, 223–259. Available online: https://digitalcommons.unl.edu/cgi/viewcontent.cgi?article=1452&context=greatplainsresearch (accessed on 20 September 2023).
- Omernik, J.M.; Griffith, G.E. Ecoregions of the conterminous United States: Evolution of a hierarchical spatial framework. Environ. Manag. 2014, 54, 1249–1266. [Google Scholar] [CrossRef]
- Knapp, A.K.; Chen, A.; Griffin-Nolan, R.J.; Baur, L.E.; Carroll, C.J.W.; Gray, J.E.; Hoffman, A.M.; Li, X.; Post, A.K.; Slette, I.J.; et al. Resolving the Dust Bowl paradox of grassland responses to extreme drought. Proc. Natl. Acad. Sci. USA 2020, 117, 22249–22255. Available online: https://www.pnas.org/cgi/doi/10.1073/pnas.1922030117 (accessed on 20 September 2023). [CrossRef] [PubMed]
- Wood, M.K.; Blackburn, W.H. Vegetation and soil responses to cattle grazing systems in the Texas Rolling Plains. J. Range Manag. 1984, 37, 303–308. [Google Scholar] [CrossRef]
- Heitschmidt, R.K.; Schultz, R.D.; Scifres, C.J. Herbaceous biomass dynamics and net primary production following chemical control of honey mesquite. J. Range Manag. 1986, 39, 67–71. [Google Scholar] [CrossRef]
- Archer, S. Tree-grass dynamics in a Prosopis-thornscrub savanna parkland: Reconstructing the past and predicting the future. EcoScience 1995, 2, 83–99. [Google Scholar] [CrossRef]
- Archer, S.R.; Andersen, E.M.; Predick, K.I.; Schwinning, S.; Steidl, R.J.; Woods, S.R. Woody plant encroachment: Causes and consequences. In Rangeland Systems—Processes, Management and Challenges; Briske, D.D., Ed.; Springer Series on Environmental Management; Springer: Berlin/Heidelberg, Germany, 2017; pp. 25–84. [Google Scholar] [CrossRef]
- Laxson, J.D.; Schacht, W.H.; Owens, M.K. Above-ground biomass yields at different densities of honey mesquite. J. Range Manag. 1997, 50, 550–554. [Google Scholar] [CrossRef]
- Van Auken, O.W. Shrub invasions of North American semiarid grasslands. Ecology 2000, 31, 197–215. [Google Scholar] [CrossRef]
- Ratajczak, Z.; Nippert, J.B.; Collins, S.L. Woody encroachment decreases diversity across North American grasslands and savannas. Ecology 2012, 93, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Archer, S.R.; Predick, K.I. An ecosystem services perspective on brush management: Research priorities for competing land-use objectives. J. Ecol. 2014, 102, 1394–1407. [Google Scholar] [CrossRef]
- Hughes, R.F.; Archer, S.R.; Asner, G.P.; Wessman, C.A.; McMurtry, C.; Nelson, J.; Ansley, R.J. Changes in aboveground primary production and carbon and nitrogen pools accompanying woody encroachment in a temperate savanna. Glob. Chang. Biol. 2006, 12, 1733–1747. [Google Scholar] [CrossRef]
- Barger, N.N.; Archer, S.R.; Campbell, J.L.; Huang, C.; Morton, J.A.; Knapp, A.K. Woody plant proliferation in North American drylands: A synthesis of impacts on ecosystem carbon balance. J. Geophys. Res. 2011, 116, G00K07. [Google Scholar] [CrossRef]
- Archer, S. Development and stability of grass/woody mosaics in a subtropical savanna parkland, Texas, U.S.A. J. Biogeogr. 1990, 17, 453–462. [Google Scholar] [CrossRef]
- Ansley, R.J.; Mirik, M.; Wu, X.B.; Heaton, C.B. Woody cover and grass production in a mesquite savanna: Geospatial relationships and precipitation. Rangel. Ecol. Manag. 2013, 66, 621–633. [Google Scholar] [CrossRef]
- Bedunah, D.J.; Sosebee, R.E. Forage response of a mesquite-buffalograss community following range rehabilitation. J. Range Manag. 1984, 37, 483–487. [Google Scholar] [CrossRef]
- Hicks, R.A.; Briske, D.D.; Call, C.A.; Ansley, R.J. Co-existence of a perennial C3 bunchgrass in a C4 dominated grassland: An evaluation of gas exchange characteristics. Photosynthetica 1990, 24, 63–74. [Google Scholar] [CrossRef]
- Murray, D.B.; Muir, J.P.; Miller, M.S.; Erxleben, D.R.; Mote, K.D. Effective management practices for increasing native plant diversity on mesquite savanna-Texas wintergrass-dominated rangelands. Rangel. Ecol. Manag. 2021, 75, 161–169. [Google Scholar] [CrossRef]
- USDA-NRCS (United States Department of Agriculture—Natural Resource Conservation Service). Nassella leucotricha (Trin. & Rupr.) Pohl; Texas Wintergrass. 2023. Available online: https://plants.usda.gov/home/plantProfile?symbol=NALE3 (accessed on 9 May 2023).
- Whisenant, S.G.; Ueckert, D.N.; Scifres, C.J. Effects of fire on Texas wintergrass communities. J. Range Manag. 1984, 37, 387–391. [Google Scholar] [CrossRef]
- Ansley, R.J.; Cooper, C.; Zhang, T. Invasion promotes invasion: Facilitation of C3 perennial grass dominance in mixed C3/C4 grassland by an invasive C3 woody sprouter (Prosopis glandulosa). Ecol. Evol. 2019, 9, 13438–13449. [Google Scholar] [CrossRef]
- Tiedemann, A.R.; Klemmedson, J.O. Effect of mesquite trees on vegetation and soils in the desert grassland. J. Range Manag. 1977, 30, 361–367. [Google Scholar] [CrossRef]
- Geesing, D.; Felker, P.; Bingham, R.L. Influence of mesquite (Prosopis glandulosa) on soil nitrogen and carbon development: Implications for global carbon sequestration. J. Arid Environ. 2000, 46, 157–180. [Google Scholar] [CrossRef]
- Wedin, D.A.; Tilman, D. Competition among grasses along a nitrogen gradient: Initial conditions and mechanisms of competition. Ecol. Monogr. 1993, 63, 199–229. [Google Scholar] [CrossRef]
- Stuart-Hill, G.C.; Tainton, N.M. The competitive interaction between Acacia karoo and the herbaceous layer and how this is influenced by defoliation. J. Appl. Ecol. 1989, 26, 285–298. [Google Scholar] [CrossRef]
- Rossi, B.E.; Villagra, P.E. Effects of Prosopis flexuosa on soil properties and the spatial pattern of understorey species in arid Argentina. J. Veg. Sci. 2003, 14, 543–550. [Google Scholar] [CrossRef]
- Prober, S.M.; Thiele, K.R.; Lunt, I.D.; Koen, T.B. Restoring ecological function in temperate grassy woodlands: Manipulating soil nutrients, exotic annuals and native perennial grasses through carbon supplements and spring burns. J. Appl. Ecol. 2005, 42, 1073–1085. [Google Scholar] [CrossRef]
- Nordenstahl, M.; Gundel, P.E.; Clavijo, M.P.; Jobbagy, E.G. Forage production in natural and afforested grasslands of the Pampas: Ecological complementarity and management opportunities. Agrofor. Syst. 2011, 83, 201–211. [Google Scholar] [CrossRef]
- Rauber, R.; Steinaker, D.; Demaria, M.; Arroyo, D. Factors associated to invasion by non-palatable grasses in Argentinean semi-arid forest. Ecol. Austral 2014, 234, 320–326. Available online: https://bibliotecadigital.exactas.uba.ar/download/ecologiaaustral/ecologiaaustral_v024_n03_p320.pdf (accessed on 29 August 2022). [CrossRef]
- USDA-NRCS (United States Department of Agriculture—Natural Resource Conservation Service). Bouteloua dactyloides (Nutt.) J.T. Columbus; Buffalograss. 2023. Available online: https://plants.usda.gov/home/plantProfile?symbol=BODA2 (accessed on 9 May 2023).
- Ansley, R.J.; Moeller, A.; Fuhlendorf, S.D. Pyric-based restoration of C4 grasses in woody encroached (Prosopis glandulosa) grassland is best with an alternating seasonal fire regime. Restor. Ecol. 2022, 30, e13644. [Google Scholar] [CrossRef]
- Frost, C.C. Presettlement fire frequency regimes of the United States: A first approximation. In Fire in Ecosystem Management: Shifting the Paradigm from Suppression to Prescription; Pruden, T.L., Brennan, L.A., Eds.; Tall Timbers Fire Ecology Conference No. 20; Tall Timbers Research Station: Tallahassee, FL, USA, 1998; pp. 70–81. [Google Scholar]
- Guyette, R.P.; Stambaugh, M.C.; Dey, D.C.; Muzika, R. Predicting fire frequency with chemistry and climate. Ecosystems 2012, 15, 322–335. [Google Scholar] [CrossRef]
- Dai, X.; Boutton, T.W.; Hailemichael, M.; Ansley, R.J.; Jessup, K.E. Soil carbon and nitrogen storage in response to fire in a temperate mixed-grass savanna. J. Environ. Qual. 2006, 35, 1620–1628. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.D.; Boutton, T.W.; Jastrow, J.D. Organic matter turnover in soil physical fractions following woody plant invasion of grassland: Evidence from natural 13C and 15N. Soil Biol. Biochem. 2006, 38, 3197–3210. [Google Scholar] [CrossRef]
- Linex, R.J. Range Plants of North Central Texas—A Land User’s Guide to Their Identification, Value and Management; USDA-NRCS Publication: Weatherford, TX, USA, 2014; pp. 345p. [Google Scholar]
- Tomecek, J.M.; Pierce, B.L.; Reyna, K.S.; Peterson, M.J. Inadequate thermal refuge constrains landscape habitability for a grassland bird species. PeerJ 2017, 5, e3709. [Google Scholar] [CrossRef] [PubMed]
- Bernath-Plaisted, J.S.; Correll, M.D.; Somershoe, S.G.; Dwyer, A.M.; Bankert, A.; Beh, A.; Berlanga, H.; Boyle, W.A.; Cruz-Romo, J.L.; George, T.L.; et al. Review of conservation challenges and possible solutions for grassland birds of the North American Great Plains. Rangel. Ecol. Manag. 2023, 90, 165–185. [Google Scholar] [CrossRef]
- Albertson, F.W.; Tomanek, G.W.; Riegel, A. Ecology of drought cycles and grazing intensity on grasslands of Central Great Plains. Ecol. Monogr. 1957, 27, 27–44. [Google Scholar] [CrossRef]
- Hatch, S.L.; Pluhar, J. Texas Range Plants; Texas A&M University Press: College Station, TX, USA, 1993; 326p. [Google Scholar]
- Wright, H.A.; Bailey, A.W. Fire Ecology; Wiley-Interscience, John Wiley & Sons: New York, NY, USA, 1982; 501p. [Google Scholar]
- Engle, D.M.; Bidwell, T.G. Viewpoint: The response of central North American prairies to seasonal fire. J. Range Manag. 2001, 54, 2–10. [Google Scholar] [CrossRef]
- Ansley, R.J.; Boutton, T.W.; Hollister, E.B. Can prescribed fires restore C4 grasslands invaded by a C3 woody species and a co-dominant C3 grass species? Ecosphere 2021, 12, e03885. [Google Scholar] [CrossRef]
- Drewa, P.B. Effects of fire season and intensity on Prosopis glandulosa Torr. var. glandulosa. Int. J. Wildland Fire 2003, 12, 147–157. [Google Scholar] [CrossRef]
- Ansley, R.J.; Pinchak, W.E.; Jones, D.L. Mesquite, tobosagrass and common broomweed responses to summer and winter season fires. Rangel. Ecol. Manag. 2008, 61, 588–597. [Google Scholar] [CrossRef]
- Twidwell, D.; Rogers, W.E.; Wonkka, C.L.; Taylor, C.A., Jr.; Kreuter, U.P. Extreme prescribed fire during drought reduces survival and density of woody sprouters. J. Appl. Ecol. 2016, 53, 1585–1596. [Google Scholar] [CrossRef]
- Ansley, R.J.; Zhang, T.; Cooper, C. Soil moisture, grass production and mesquite resprout architecture following mesquite above-ground mortality. Water 2018, 10, 1243. [Google Scholar] [CrossRef]
- USDA-NRCS (United States Department of Agriculture—Natural Resource Conservation Service). Chapter H—Central Great Plains Winter Wheat and Range Region. In Land Resource Regions and Major Land Resource Areas of the United States, the Caribbean, and the Pacific Basin; USDA Agricultural Handbook 296; USDA-NRCS Publication: Washington, DC, USA, 2006; pp. 195–228. [Google Scholar]
- Bartush, B.; Kloesel, K.; Banner, J.; Brown, D.; Lemery, J.; Lin, X.; Loeffler, C.; McManus, G.; Mullens, E.; Nielsen-Gammon, J.; et al. Chapter 23-Southern Great Plains. In U.S. Global Change Research Program, Fourth National Climate Assessment; Nasser, E., Ed.; Steering Committee; U.S. Global Change Research Program: Washington, DC, USA, 2018; pp. 987–1035. Available online: https://nca2018.globalchange.gov/chapter/23/ (accessed on 29 August 2022).
- NFWF (National Fish and Wildlife Foundation). Southern Great Plains Map. 2023. Available online: https://www.nfwf.org/sites/default/files/2022-01/cpp-jan2022-southern-great-plains-map.pdf (accessed on 29 August 2022).
- NOAA-NCEI (National Oceanic and Atmospheric Administration—National Centers for Environmental Information). Wilbarger County, Texas Precipitation. 2022. Available online: https://www.ncei.noaa.gov/cag/county/time-series/TX-487/pcp/all/12/2010-2018 (accessed on 29 August 2022).
- USDA-NRCS (United States Department of Agriculture—Natural Resource Conservation Service). Web Soil Survey. 2022. Available online: http://websoilsurvey.nrcs.usda.gov/app/WebSoilSurvey.aspx (accessed on 13 July 2022).
- Stubbendieck, J.; Hatch, S.L.; Butterfield, C.H. North American Range Plants; University of Nebraska Press: Lincoln, NE, USA, 1992; 493p. [Google Scholar]
- Stubbendieck, J.; Hatch, S.L.; Dunn, C.D. Grasses of the Great Plains; Texas A&M University Press: College Station, TX, USA, 2017; 722p. [Google Scholar]
- Tyrl, R.J.; Bidwell, T.G.; Masters, R.E.; Elmore, R.D. Field Guide to Oklahoma Plants—Commonly Encountered Prairie, Shrubland and Forest Species; Department of Natural Resource Ecology and Management, Oklahoma State University: Stillwater, OK, USA, 2008; pp. 138–139. [Google Scholar]
- Asner, G.P.; Archer, S.R.; Hughes, R.F.; Ansley, R.J.; Wessman, C.A. Net changes in regional woody vegetation cover and carbon storage in Texas drylands, 1937–1999. Glob. Chang. Biol. 2003, 9, 316–335. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS/STAT Version 14.1 User’s Guide, Proc Mixed Procedure; SAS Institute Inc.: Cary, NC, USA, 2015; pp. 6048–6234. Available online: https://support.sas.com/documentation/onlinedoc/stat/141/mixed.pdf (accessed on 29 August 2022).
- Marcum, K.B.; Engelke, M.C.; Morton, S.J. Rooting characteristics of buffalograsses grown in flexible plastic tubes. HortScience 1995, 30, 1390–1392. [Google Scholar] [CrossRef]
- Qian, Y.; Fry, J.D. Water relations and drought tolerance of four turfgrasses. J. Am. Soc. Hort. Sci. 1997, 122, 129–133. [Google Scholar] [CrossRef]
- Huang, B. Water relations and root activities of Buchloe dactyloides and Zoysia japonica in response to localized soil drying. Plant Soil 1999, 208, 179–186. [Google Scholar] [CrossRef]
- Ford, P.L. Response of buffalograss (Buchloe dactyloides) and blue grama (Bouteloua gracilis) to fire. Great Plains Res. 1999, 9, 261–276. [Google Scholar]
- Porensky, L.M.; Derner, J.D.; Augustine, D.J.; Milchunas, D.G. Plant community composition after 75 yr of sustained grazing intensity treatments in Shortgrass Steppe. Rangel. Ecol. Manag. 2017, 70, 456–464. [Google Scholar] [CrossRef]
- Ansley, R.J.; Zhang, T.; Murray, B.D. Sensitivity of different grass functional groups to honey mesquite encroachment: Toward developing a multi-year model. Rangel. Ecol. Manag. 2023, 90, 279–289. [Google Scholar] [CrossRef]
- Ansley, R.J.; Boutton, T.W.; Jacoby, P.W. Root biomass and distribution patterns in a semi-arid mesquite savanna: Responses to long-term rainfall manipulation. Rangel. Ecol. Manag. 2014, 67, 206–218. [Google Scholar] [CrossRef]
- Walter, H. Chapter 6: Natural savannahs as a transition to the arid zone. In Ecology of Tropical and Subtropical Vegetation; Walter, H., Ed.; Oliver and Boyd: Edinburgh, Scotland, 1971; pp. 238–265. [Google Scholar]
- Walker, B.H.; Ludwig, D.; Holling, C.S.; Peterman, R.M. Stability of semi-arid savanna grazing systems. J. Ecol. 1981, 69, 473–498. [Google Scholar] [CrossRef]
- Heitschmidt, R.K.; Ansley, R.J.; Dowhower, S.L.; Jacoby, P.W.; Price, D.L. Some observations from the excavation of honey mesquite root systems. J. Range Manag. 1988, 41, 227–231. [Google Scholar] [CrossRef]
- Anderson, R.C. The historic role of fire in the North American grassland. In Fire in North American Grasslands; Collins, S.L., Wallace, L.L., Eds.; University of Oklahoma Press: Norman, OK, USA, 1990; pp. 8–18. [Google Scholar]
- Wright, H.A. Effect of fire on southern mixed prairie grasses. J. Range Manag. 1974, 27, 417–419. [Google Scholar] [CrossRef]
- Ansley, R.J.; Castellano, M.J.; Pinchak, W.E. Sideoats grama growth responses to seasonal fires and clipping. Rangel. Ecol. Manag. 2006, 59, 258–266. [Google Scholar] [CrossRef]
- Ford, P.L.; Johnson, G.V. Effects of dormant- vs. growing-season fire in shortgrass steppe: Biological soil crust and perennial grass responses. J. Arid Environ. 2006, 67, 1–14. [Google Scholar] [CrossRef]
- Cialdella, A.M.; Salariato, D.L.; Aagesen, L.; Giussani, L.M.; Zuloaga, F.O.; Morrone, O. Phylogeny of New World Stipeae (Poaceae): An evaluation of the monophyly of Aciachne and Amelichloa. Cladistics 2010, 26, 563–578. [Google Scholar] [CrossRef] [PubMed]
- Romaschenko, K.; Peterson, P.M.; Soreng, R.J.; Garcia-Jacas, N.; Futorna, O.; Susanna, A. Systematics and evolution of the needle grasses (Poaceae: Pooideae: Stipeae) based on analysis of multiple chloroplast loci, ITS, and lemma micromorphology. Taxon 2012, 161, 18–44. [Google Scholar] [CrossRef]
- Bessega, C.; Vilardi, J.C.; Saidman, B.O. Genetic relationships among American species of the genus Prosopis (Mimosoideae, Leguminosae) inferred from ITS sequences: Evidence for long-distance dispersal. J. Biogeogr. 2006, 33, 1905–1915. [Google Scholar] [CrossRef]
- Ansley, R.J.; Castellano, M.J. Texas wintergrass and buffalograss response to seasonal fires and clipping. Rangel. Ecol. Manag. 2007, 60, 154–164. [Google Scholar] [CrossRef]
- Hulbert, L.C. Fire and litter effects in undisturbed bluestem prairie in Kansas. Ecology 1969, 50, 874–877. [Google Scholar] [CrossRef]
- Knapp, A.K.; Seastedt, T.R. Detritus accumulation limits productivity of tallgrass prairie. Bioscience 1986, 36, 622–668. [Google Scholar] [CrossRef]
- Facelli, J.M.; Pickett, S.T.A. Plant litter: Light interception and effects on an oldfield plant community. Ecology 1991, 72, 1024–1031. [Google Scholar] [CrossRef]



| Treatment Short Name | Treatment Long Name | Steps in Each Fire Treatment | Time Burned |
|---|---|---|---|
| No fire | No Fire | None | None |
| 3WF | Repeated winter fires | w91 + w93 + w95 | Late January to early March |
| 3AF | Alternate season fires | w91 + s92 + w94 | Winter—late January to early March; summer—early September |
| 3AFC | Alternate season fires compressed sequence | w93 + s93 + w96 | Winter—late January to early March; summer—early September |
| 2SF | Repeated summer fires | s92 + s94 | Early September |
| 2SFC | Repeated summer fires compressed sequence | s93 + s94 | August; early September |
| Effect | df | Buchloe | Nassella | C4M | C3A | Forbs | Litter | Bare |
|---|---|---|---|---|---|---|---|---|
| ----- Buchloe patches ----- | ||||||||
| Clip | 1 | 0.1027 | <0.0001 | 0.0003 | <0.0001 | 0.2480 | 0.0001 | <0.0001 |
| Fire | 5 | <0.0001 | <0.0001 | 0.0303 | 0.0026 | 0.0079 | <0.0001 | 0.0517 |
| Year | 5 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Clip × Fire | 5 | 0.0240 | 0.3054 | 0.9274 | 0.7287 | 0.6684 | 0.0420 | 0.4078 |
| Clip × Year | 5 | <0.0001 | 0.0293 | 0.0984 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Fire × Year | 24 | <0.0001 | 0.0063 | 0.2178 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Clip × Fire × Year | 24 | 0.3689 | 0.2224 | 0.9932 | 0.0237 | 0.0142 | 0.0736 | 0.4612 |
| ----- Nassella patches ----- | ||||||||
| Clip | 1 | <0.0001 | <0.0001 | 0.9347 | <0.0001 | 0.4579 | <0.0001 | <0.0001 |
| Fire | 5 | <0.0001 | 0.0024 | 0.0017 | <0.0001 | 0.0009 | <0.0001 | <0.0001 |
| Year | 5 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Clip × Fire | 5 | 0.8585 | 0.0217 | 0.0275 | 0.5851 | 0.3135 | 0.5106 | 0.0007 |
| Clip × Year | 5 | <0.0001 | <0.0001 | 0.9018 | <0.0001 | 0.0005 | <0.0001 | <0.0001 |
| Fire × Year | 24 | 0.9137 | 0.0355 | 0.7052 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Clip × Fire × Year | 24 | 1.0000 | 0.9253 | 0.9900 | 0.1693 | 0.7040 | 0.0059 | 0.0149 |
| Fire Effect | Buchloe Patches | Nassella Patches | Year Effect | Buchloe Patches | Nassella Patches |
|---|---|---|---|---|---|
| No fire | 4.33 ± 1.35 a | 3.50 ± 1.07 b | 1992 | 0.50 ± 0.30 cd | 0.33 ± 0.33 c |
| 3WF | 1.18 ± 0.42 c | 6.97 ± 1.22 ab | 1993 | 0.01 ± 0.01 d | 0.46 ± 0.21 c |
| 3AF | 2.81 ± 0.66 ab | 7.85 ± 1.99 a | 1994 | 2.53 ± 0.63 ab | 3.91 ± 0.91 b |
| 3AFC | 1.99 ± 0.56 abc | 7.85 ± 1.71 a | 1996 | 1.64 ± 0.55 bc | 8.19 ± 1.44 a |
| 2SF | 1.25 ± 0.45 c | 3.77 ± 0.87 b | 1997 | 3.84 ± 0.91 a | 8.68 ± 1.45 a |
| 2SFC | 1.74 ± 0.61 bc | 3.75 ± 1.17 b | 1998 | 4.14 ± 0.96 a | 11.57 ± 1.96 a |
| Effect | df | Buchloe | Nassella | C4M | C3A | Forbs | Litter | Bare |
|---|---|---|---|---|---|---|---|---|
| ----- Buchloe patches ----- | ||||||||
| Clip | 1 | 0.7030 | 0.0001 | 0.0457 | 0.2393 | 0.0046 | 0.5847 | 0.0006 |
| Fire | 5 | 0.1077 | 0.0008 | 0.3641 | 0.0160 | 0.0608 | 0.0035 | 0.0860 |
| Clip × Fire | 5 | 0.7764 | 0.0574 | 0.9599 | 0.7388 | 0.2640 | 0.2271 | 0.7662 |
| ----- Nassella patches ----- | ||||||||
| Clip | 1 | <0.0001 | <0.0001 | 0.7323 | 0.4156 | 0.0115 | 0.0019 | <0.0001 |
| Fire | 5 | 0.0979 | 0.2687 | 0.2958 | 0.0054 | 0.0018 | <0.0001 | 0.0113 |
| Clip × Fire | 5 | 0.9345 | 0.7893 | 0.2229 | 0.5115 | 0.8175 | 0.3634 | 0.4142 |
| Treatment | C3A | Forbs | Litter | Bare Ground |
|---|---|---|---|---|
| ----- Buchloe patches ----- | ||||
| No clip | 5.63 ± 0.94 x | 2.04 ± 0.42 y | 16.68 ± 3.15 x | 1.21 ± 0.61 y |
| Clip | 6.85 ± 0.85 x | 4.31 ± 0.54 x | 17.12 ± 1.75 x | 7.22 ± 1.88 x |
| No fire | 8.17 ± 1.46 ab | 1.67 ± 1.05 b | 26.92 ± 6.52 a | 10.00 ± 4.87 a |
| 3WF | 9.22 ± 1.57 a | 4.45 ± 0.85 a | 21.50 ± 3.68 ab | 4.17 ± 1.90 ab |
| 3AF | 2.55 ± 0.56 c | 3.33 ± 0.69 ab | 7.75 ± 1.35 d | 0.83 ± 0.53 b |
| 3AFC | 7.38 ± 1.80 ab | 2.78 ± 1.09 ab | 19.72 ± 3.39 abc | 5.15 ± 1.70 ab |
| 2SF | 5.60 ± 0.96 abc | 3.90 ± 0.95 ab | 15.00 ± 1.61 bcd | 1.80 ± 1.26 b |
| 2SFC | 4.53 ± 1.28 bc | 2.92 ± 1.00 ab | 10.53 ± 3.00 cd | 3.33 ± 2.86 ab |
| ----- Nassella patches ----- | ||||
| No clip | 7.68 ± 1.01 x | 2.08 ± 0.55 y | 31.11 ± 3.74 x | 0.79 ± 0.46 y |
| Clip | 8.76 ± 1.11 x | 4.58 ± 0.85 x | 22.09 ± 2.89 y | 8.10 ± 1.25 x |
| No fire | 11.88 ± 2.08 a | 0.42 ± 0.42 d | 48.55 ± 5.21 a | 5.83 ± 2.39 ab |
| 3WF | 10.28 ± 1.13 a | 2.37 ± 0.92 bcd | 28.53 ± 2.62 b | 4.03 ± 2.00 ab |
| 3AF | 4.42 ± 0.97 b | 1.38 ± 0.63 cd | 11.98 ± 1.84 d | 1.67 ± 1.67 b |
| 3AFC | 4.38 ± 1.20 b | 4.45 ± 1.30 abc | 15.63 ± 4.04 cd | 8.33 ± 2.69 a |
| 2SF | 9.85 ± 1.31 a | 6.38 ± 1.34 a | 28.80 ± 2.52 b | 1.80 ± 1.26 b |
| 2SFC | 8.52 ± 1.94 ab | 5.00 ± 1.44 ab | 26.10 ± 5.28 bc | 5.00 ± 2.50 ab |
| Effect | df | Buchloe | Nassella | C4M | C3A | Forbs | Litter | Bare |
|---|---|---|---|---|---|---|---|---|
| Buchloe patches | ||||||||
| Clip | 1 | <0.0001 | <0.0001 | 0.0001 | 0.6078 | 0.0026 | <0.0001 | 0.0356 |
| Fire | 5 | <0.0001 | <0.0001 | 0.0018 | 0.0006 | 0.0008 | <0.0001 | 0.0003 |
| Year | 2 | <0.0001 | 0.0001 | 0.3656 | <0.0001 | 0.2668 | <0.0001 | <0.0001 |
| Clip × Fire | 5 | 0.1797 | <0.0001 | 0.7053 | 0.0898 | 0.8432 | 0.4419 | 0.0667 |
| Clip × Year | 2 | <0.0001 | 0.6876 | 0.8054 | 0.5829 | 0.4732 | <0.0001 | 0.0046 |
| Fire × Year | 10 | 0.5771 | 0.0882 | 0.9998 | 0.0070 | 0.3309 | 0.6958 | 0.2671 |
| Clip × Fire × Year | 10 | 0.8645 | 0.5736 | 0.9998 | 0.6033 | 0.5339 | 0.9226 | 0.9906 |
| Nassella patches | ||||||||
| Clip | 1 | <0.0001 | <0.0001 | 0.2418 | 0.3770 | 0.3450 | <0.0001 | 0.0003 |
| Fire | 5 | 0.0004 | 0.0091 | 0.0642 | 0.0001 | 0.0004 | <0.0001 | 0.0336 |
| Year | 2 | 0.1352 | <0.0001 | 0.0527 | <0.0001 | 0.1256 | <0.0001 | <0.0001 |
| Clip × Fire | 5 | 0.4860 | 0.0137 | 0.0235 | 0.1033 | 0.5333 | 0.3347 | 0.0027 |
| Clip × Year | 2 | 0.5787 | 0.0730 | 0.9030 | 0.8978 | 0.1057 | 0.2293 | 0.0004 |
| Fire × Year | 10 | 0.6542 | 0.0149 | 0.9921 | 0.1157 | 0.0084 | 0.0193 | 0.0329 |
| Clip × Fire × Year | 10 | 0.9967 | 0.9172 | 0.9975 | 0.8987 | 0.7603 | 0.7377 | 0.6975 |
| Effect | df | Buchloe | Nassella | C4M | C3A | Forbs | Litter | Bare |
|---|---|---|---|---|---|---|---|---|
| Buchloe patches | ||||||||
| Clip | 1 | <0.0001 | 0.0018 | 0.0332 | 0.8677 | 0.4107 | 0.0001 | 0.8856 |
| Fire | 5 | 0.0230 | 0.0002 | 0.1364 | 0.0172 | 0.0316 | 0.1089 | 0.3914 |
| Clip × Fire | 5 | 0.3407 | 0.0175 | 0.9251 | 0.2276 | 0.7323 | 0.8595 | 0.6728 |
| Nassella patches | ||||||||
| Clip | 1 | <0.0001 | 0.0015 | 0.2944 | 0.6208 | 0.9149 | <0.0001 | 0.9937 |
| Fire | 5 | 0.0651 | 0.0094 | 0.6559 | 0.0751 | 0.0196 | 0.0103 | 0.1662 |
| Clip × Fire | 5 | 0.8676 | 0.1872 | 0.6687 | 0.5132 | 0.9397 | 0.3971 | 0.1787 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ansley, R.J.; Pinchak, W.E. Stability of C3 and C4 Grass Patches in Woody Encroached Rangeland after Fire and Simulated Grazing. Diversity 2023, 15, 1069. https://doi.org/10.3390/d15101069
Ansley RJ, Pinchak WE. Stability of C3 and C4 Grass Patches in Woody Encroached Rangeland after Fire and Simulated Grazing. Diversity. 2023; 15(10):1069. https://doi.org/10.3390/d15101069
Chicago/Turabian StyleAnsley, R. James, and William E. Pinchak. 2023. "Stability of C3 and C4 Grass Patches in Woody Encroached Rangeland after Fire and Simulated Grazing" Diversity 15, no. 10: 1069. https://doi.org/10.3390/d15101069
APA StyleAnsley, R. J., & Pinchak, W. E. (2023). Stability of C3 and C4 Grass Patches in Woody Encroached Rangeland after Fire and Simulated Grazing. Diversity, 15(10), 1069. https://doi.org/10.3390/d15101069