Abstract
We designed and tested species-specific PCR primers to detect Trichobilharzia species via environmental DNA (eDNA) barcoding in selected Austrian water bodies. Tests were performed with eDNA samples from the field as well as with artificial samples from the lab, where snails releasing cercariae were kept in aquariums. From two localities, Trichobilharzia was documented based on the release of cercariae from snails, enabling morphological species identification. In both cases, the corresponding species were detected via eDNA: Trichobilharzia szidati and Trichobilharzia physellae. Nonetheless, the stochasticity was high in the replicates. PCR tests with aquarium water into which the cercariae had been released allowed eDNA detection even after 44 days. As in the PCRs with eDNA samples from the field, positive results of these experiments were not obtained for all samples and replicates. PCR sensitivity tests with dilution series of T. szidati genomic DNA as well as of PCR amplification products yielded successful amplification down to concentrations of 0.83 pg/µL and 0.008 pg/µL, respectively. Our results indicate that the presumed species specificity of PCR primers may not be guaranteed, even if primers were designed for specific species. This entails misidentification risks, particularly in areas with incomplete species inventories.
1. Introduction
Living organisms naturally release DNA into their environment. This may occur via cells separated from the body surface or by organs communicating with the external environment (e.g., mucus cells and intestinal cells). DNA release to the environment may take place on a large scale even after the death of the organism. Such DNA is termed environmental DNA or “eDNA” and can be detected and analysed using molecular genetic methods such as DNA barcoding [1]. In addition to this intracellular eDNA, free (extracellular) DNA is present in the environment, in the substrate and in water bodies, mostly bound to particles. Extracellular eDNA is generally more short-lived than intracellular eDNA and the speed of degradation differs between free-flowing vs. particle-bound eDNA [2].
Analyses of eDNA using environmental samples as the basis for species detection have gained increasing importance and have been discussed as alternatives or complementary approaches to classical species monitoring, which requires direct observation and/or collection of organisms or their traces. Samples of eDNA from water bodies offer methodological benefits compared to other environmental samples but are also associated with certain difficulties. Unlike in terrestrial systems, where DNA remains predominantly detectable locally, DNA or cells released into water may spread and thus are potentially detectable in wider parts of the water body at places where the organisms themselves do not occur. While in flowing waters the transport of eDNA plays a major role, this is of lesser importance in stagnant waters. Even in the latter, however, factors such as wind direction, water currents, and stratification may influence DNA distribution [3]. A second difference of eDNA in aquatic compared to terrestrial systems is its faster degradation in the former. The degradation speed can vary greatly and depends on factors including water temperature, pH, UV exposure, dissolved ions, as well as the organic activity of macro- and microorganisms [4,5,6,7].
The present study focused on eDNA detection by means of DNA barcode sequencing of avian schistosomes (Trematoda, Digenea, Schistosomatidae), obligate parasites of animals that use birds as final hosts. Eggs of the avian schistosomes are released into the water with the faeces of the final host. Swimming, free-living larvae—miracidia—hatch from the eggs and swim in search of the first intermediate host to infect, namely aquatic gastropods. Miracidia penetrate the gastropod host, then transform into mother sporocysts that produce several daughter sporocysts asexually, which finally release multiple infective larvae (cercariae). Once these cercariae are released from the snails, they actively penetrate the final host.
Among avian schistosomes, several species are of medical relevance because humans (and other mammals) may serve as accidental hosts. After penetration, the cercariae (schistosomula) show only limited development in humans and cannot mature [8,9,10,11], but they may cause dermatitis symptoms (“Swimmer’s itch”, “cercarial dermatitis”) with medical consequences such as intensively itching maculo–papulovesicular eruptions with possible secondary infections and local oedema. In serious cases, generalized signs such as fever, cough, diarrhoea, and local lymph node swelling have been observed [11,12]. The occurrence of cercariae and cases of cercarial dermatitis concern bathers as well as certain occupation groups such as those working in fishery and aquaculture, biologists working in water habitats, or lifeguards in natural swimming areas. This makes cercariae relevant for public health and the local economy (especially when tourists are concerned) because cercarial dermatitis may lead to the closure of near-natural bathing facilities [11,13]. The causative agents of cercarial dermatitis in Europe are mostly species of the genus Trichobilharzia with approximately 35 species worldwide [14]. Six of these species are known in Europe [15,16], namely Trichobilharzia szidati Neuhaus 1952 [17], Trichobilharzia regenti Horák, Kolářová, & Dvořák 1998 [18], Trichobilharzia franki Müller & Kimmig 1994 [19], Trichobilharzia salmanticensis Simon-Vicente & Simon-Martin 1999 [20], Trichobilharzia anseri Jouet et al. 2015 [15], and Trichobilharzia mergi Kolářová, Skírnisson, Ferté, & Jouet 2013 [21]. Recently, an additional species, Trichobilharzia physellae (Talbot, 1936) [22], originating from North America, was recorded in a lake in Austria [23]. Besides Trichobilharzia, five other genera of avian schistosomes have been reported to infect aquatic birds in Europe: Allobilharzia, Bilharziella, Dendritobilharzia, Gigantobilharzia, and Ornithobilharzia [13].
Detecting cercariae by direct sampling with nets is often hampered because of their unpredictable distribution in the water body combined with their small size and fragility. The traditional approach is to collect snails of potential host species, put them into separate glass jars filled with water, and incubate them under artificial or natural light to stimulate the shedding of the cercariae; these are subsequently further investigated (see Section 2.2). Due to the low prevalence of parasites in intermediate mollusc hosts in the patent period, however, the detection by such cercarial release experiments usually requires sampling large numbers of gastropods (which might be under nature protection). Such interventions in snail populations are therefore not desirable and may pose legal hurdles as well.
Monitoring by sampling and analysing environmental water would avoid the time-consuming processes of sampling gastropods, conducting releasing experiments, and mounting cercariae for species identification. Thus, eDNA analyses have several advantages for the detection of dermatitis-causing cercariae: they are less labour intensive, have only a minor impact on the environment (compared with collecting hundreds of snails), and are highly informative considering that cercariae of many species are morphologically not distinguishable [24]. Several studies based on eDNA combined with a qPCR approach successfully detected cercariae down to the species level [25,26,27,28,29,30,31]. Compared to the qPCR approach, where the presence of a species is indicated by PCR success, the barcoding approach includes sequencing of the obtained PCR product and thus has the potential to reveal false positives.
Several factors must be considered in evaluating the potential of detecting cercarial DNA in freshwater samples. The quantity and duration of cercarial shedding as well as the length of cercarial survival under natural conditions may influence the sensitivity of the analyses as well as the process of DNA degradation after cercarial death. Concerning the quantity and circadian rhythm of shedding, studies on Trichobilharzia szidati yielded high numbers (many thousands per day) and a long duration of shedding (several weeks), with maximum shedding depending of the time of day, temperature, and light [32]. Field studies with T. szidati revealed maximum shedding in late summer [33]. In laboratory experiments, those authors determined that cercarial life spans are inversely correlated to temperature and that cercaria remained active for up to 60 h at 20 °C. Accordingly, the success of DNA water analyses may depend on different parameters such as cercarial production, temperature, and light conditions, along with other external influences such as water current conditions, water physics/chemistry, and ecological interferences, which are poorly studied [33].
The initial aim of the present pilot study was (1) to design and test species-specific PCR primers for eDNA detection of Trichobilharzia species occurring in Austria. (2) We then compared the eDNA results with the results from cercarial release experiments (as described above and in Section 2.2). (3) We also report observations on the release of cercariae in the laboratory.
2. Materials and Methods
2.1. Field Work and eDNA Sampling
During field excursions in the framework of an overarching project in 2019 and 2020, freshwater snails as potential hosts were collected in six water bodies in Upper and Lower Austria, at altogether seven sampling localities (Table 1). The present study focused on species of the family Lymnaeidae because the Trichobilharzia species in Europe are almost exclusively found in lymnaeid snail hosts [34]. The water bodies in Table 1 include two tributaries of river systems (locality 1, 2), three lakes (locality 3, 5, 9) and three ponds (locality 4, 7, 8). They were selected based on the occurrence of Lymnaeidae as well as waterfowl. Moreover, swimmer’s itch had been recorded in some of them in the past (locality 4, 5, 8, 9).

Table 1.
List of locations where host snails and eDNA samples were collected for the present study.
Snails were examined for cercariae using conventional illumination methods (cercarial release experiments; see Section 2.2). In addition, eDNA water samples were taken at these water bodies (1, 2, 3, 4, 5, 8, 9). The aim was to compare the results of the release experiments (Section 2.3) with those from the eDNA analyses. The eDNA samples were taken near aquatic vegetation in shallow water (down to 0.5 m depth, not stirring up the sediment) as close as possible to where the host snails were collected. Sampling was conducted, with one exception (eDNA-9, locality 4), close to the shoreline. At every location visited in 2020, at least two samples were taken. For eDNA sampling, water was squeezed by hand with sterile, DNA-free plastic syringes through Sterivex™ filters with a pore size of 0.45 µm (Merck Millipore, Darmstadt, Germany). Sampling was conducted with fresh sterile gloves for each sampling, taking care that no sampling equipment came into contact with collected snails or transfer bottles. Filters were immediately stored in cool containers and subsequently transported to the laboratory and stored at −80 °C. The initial aim to filter a standardized volume (2 L) of water could not be accomplished. The maximum water volume that could be squeezed through the filters varied considerably among water bodies depending on the number of suspended solids: in rather turbid water, e.g., at locality 4, only 100 mL could be used compared to a maximum of 1200 mL at locality 8. Overall, 18 eDNA samples of water taken from seven localities were analysed: 13 of these were collected directly in the field, four were obtained in the laboratory from water samples (without snails) brought from the field in bottles (eDNA samples taken in the lab). One eDNA sample was taken from transport water containing a living Lymnaea stagnalis (which later released T. szidati cercariae). In addition, 15 eDNA samples were taken from aquariums in which snails were kept for some time (up to 24 h), three of them when the snails were still present and 12 of them after having removed the snails.
2.2. Release of Cercariae in the Laboratory
Collected snails were placed individually in jars and positioned on the windowsill (exposed to indirect sunlight during day) for up to three days, as light stimulates the release of cercariae. Released cercariae were fixed in 80–96% ethanol. For morphological identification, the cercariae isolated during the hatching experiments were transferred to a glycerol-ethanol (80%) mixture (50:50) including 1 mL borax-carmine solution. After evaporation of the ethanol in a thermo-incubator (approx. after 48 h at 40 °C), cercariae were placed in a drop of glycerol on a micro slide, covered with a cover glass, and sealed. The glycerol increases the lucency, whereas the slight staining with borax-carmine renders the anatomy more visible. Mounted cercariae were morphologically examined under a microscope (Nikon Eclipse Ni-U, Nikon Instruments Inc., New York, NY, USA) and microphotographs were made using the mounted Nikon DSRi2 microscope camera unit and the corresponding application NIS-Elements BR v.5.02.00. Subsequently, the microphotographs were processed with Gimp 2.10.24 (https://www.gimp.org, accessed on 28 May 2021). The specimens were stored in the NHMW collection Evertebrata Varia (inventory numbers for the permanent mounts NHMW-ZOO-EV-Micro 5860-5862 and for the fixed cercariae NHMW-ZOO-EV 21447-21449).
An additional experiment was performed to investigate how long cercariae are shed by an adult Lymnaea stagnalis (from Rekawinkel, Lower Austria) infected with T. szidati. This specimen had been collected and brought to the laboratory on 20 August 2020 and was kept in a glass jar with water from the pond. On 23 August 2020, the snail was placed into an aquarium for the experiments. On 24 August 2020, the snail was placed into another aquarium and eDNA longevity tests were conducted with the water from the first aquarium (see Section 2.9). All aquariums (25 L) were filled before the experiments with tap water and left to stand for two days before the snail was introduced in order not to stress the snail further. On three days, 2 and 16 September as well as 6 October 2020, the animal was incubated in a jar in 250 mL of tap water at 22 °C for 12 h in daylight to check for shedding of cercariae. The snail from the aquarium was transferred into the jar to observe and record cercarial release and to count numbers of cercariae, which was not possible in the aquarium.
2.3. Cercariae Analysed Genetically
Cercariae of Trichobilharzia species released from snails collected in the field were fixated. Some were mounted for morphological examination, and some were genetically analysed (see Section 2.4 following pages.) to obtain DNA barcode sequences of the mitochondrial cytochrome c oxidase subunit 1 gene (CO1). Their CO1 sequences were used for the procedures of primer design, which also included sequences of T. franki and T. physellae, which we published earlier [23,35], and of other species (from GenBank). Moreover, for primer design, the Department of Parasitology of Charles University (Prague, Czech Republic) provided further cercariae (T. franki, T. szidati, T. regenti, Table 1). This addition was important to include another species probably also present in Austria (T. regenti) and to cover a wider range of genetic variability. The five specimens analysed for the primer design in the present study were: T. franki (2 specimens; Ra2-4-014, Ra13-16-002), T. szidati (2 specimens; Ls17-15-003, Ls36-13-002), and T. regenti (1 specimen; Rlag1-15-002) (Table 2). All DNA barcodes of cercariae generated in the present study complemented the collection of reference sequences at the NHMW database (ABOL, Austrian Barcode of Life Initiative) and likewise were uploaded to the NCBI (National Center for Biotechnology Information) gene database GenBank, and BOLD (Barcode of Life Database; see Table 2).
Table 2.
List of cercariae analysed genetically and their snail host species. The locality numbers correspond to those in Table 1.
2.4. DNA Extraction
DNA was extracted in the clean room of the NHM DNA laboratory, following standard routines against contaminations, e.g., regular overnight UV irradiation of the clean room, treatment of work surfaces with DNA Exitus (AppliChem, Darmstadt, Germany) and/or 10% sodium hypochlorite, UV irradiation of reaction tubes. All post-PCR work was performed in a separate laboratory. All DNA extractions included control extractions without samples to screen for contaminated reagents. Likewise, all PCRs included negative control reactions without template DNA. PCRs with good-quality DNA (e.g., from cercariae) were made independently from those with eDNA. For the DNA analysis of cercariae, we used the QIAamp DNA Micro Kit (Qiagen, Hilden, Germany), which is optimised for small tissue samples. As described in Helmer et al. [23], the cercariae were isolated individually with stainless insect needles under a stereomicroscope (Wild-Leica Heerbrugg M420 macroscope, Leica, Wetzlar, Germany), dried on the needle for about 10 s to remove the ethanol, and then transferred to the lysis buffer. The extraction was performed according to the manufacturer’s protocol. In the last step, the DNA was eluted with 25 μL AE buffer.
For eDNA analysis, the collected filter samples were extracted using the dNeasy PowerWater Sterivex Kit (Qiagen, Hilden, Germany). The extraction was performed according to the manufacturer’s protocol except omitting the incubation of the Sterivex filter at 90 °C (step 7 of the protocol) because less DNA shearing was desired. In the last step, the DNA was eluted with 50 µL of EB solution.
2.5. PCR Amplification of Marker Sequences
Partial sequences of the following genes were studied: (1) the mitochondrial cytochrome c oxidase subunit 1 gene (CO1), (2) the nuclear-encoded 18S rRNA gene (18S). The latter were used to test eDNA samples for any potential trematode DNA (the 18S primers are universal primers in contrast to the CO1 primers, which were aimed to bind Trichobilharzia spp.). CO1 sequences were analysed in single cercariae as well as in eDNA samples (see Section 2.6). First, a partial sequence of the CO1 gene was amplified by PCR from five cercariae (Table 2) with the primer pair Cox1_schist_5_trich/Tricho_rev_20, which yields a fragment of 1216 bp in length. This section of the CO1 gene is considerably longer than the usually amplified DNA barcoding region (the 634 bp long, so-called “Folmer region“; [36]). Since sequencing of some sections of this fragment was difficult due to DNA homopolymers as well as due to limitations of Sanger sequencing, additional overlapping shorter fragments were amplified to achieve overall high-quality sequences and to achieve barcode quality according to the requirements of the BOLD database. For this task, the following primer pairs were used for PCR as well as sequencing: Tricho_tRNA_fw/Tricho_tRNA_rv_2 (amplicon length of 396 bp), Cox1_schist_5_trich/CO1560R_modif (amplicon length of 612 bp), Tricho_Fw2/Tricho_Rv2_2 (amplicon length of 491 bp), and ZDOE-COI-fw/Tricho_rev_20 (amplicon length of 486 bp). All primers are listed in Table 3. For an overview of the primer positions see Figure 1.
Table 3.
List of CO1 and 18S PCR primers used. Primer Tricho_tRNA_fw binds in the adjacent tRNA Ser gene. Fwd = forward primer, Rev = reverse primer. Asterisks: primers designed for particular species.
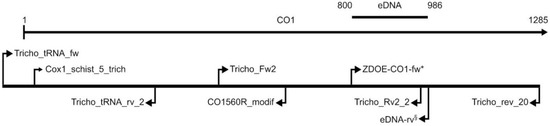
Figure 1.
Positions of primers used relative to CO1 of Trichobilharzia szidati (mt genome, GenBank accession number MG570047). The orientations of primers are marked by arrows. The long line on top shows part of the CO1 gene from the gene start (position 1) until the position of the 3′-end of primer Tricho_rev_20 (position 1285). The short line on top illustrates the position of the 186 bp-amplicon used in the eDNA analyses. The asterisk (*) indicates that this is the position of ZDOE-COI-fw_szidati as well as ZDOE-COI-fw_physellae; eDNA-rv§ indicates the position of the four primers eDNA-franki-rv, eDNA-regenti-rv, eDNA-szidati-rv, and eDNA-physellae-rv.
Polymerase chain reaction (PCR) was carried out in a 25 µL reaction volume with TopTaq or (later, when TopTaq was no longer available) Taq DNA polymerase (both from Qiagen, Hilden, Germany) as described in Helmer et al. [23]. For some experiments, i.e., for the tests of primer specificity (see Section 3.1 and Section 3.2), another polymerase was used (Qiagen Multiplex PCR Kit). PCR ingredients were, besides reaction buffer provided by the manufacturer, 0.25 units DNA polymerase, 0.5 µM of each primer, and 0.2 mM of each dNTP (Invitrogen, Carlsbad, CA, USA). For the amplifications with the Multiplex PCR Kit, 12.5 µL Multiplex PCR Master Mix was used. For amplification of the CO1 fragments from cercariae, 1.5 µL of DNA template was used, for eDNA (CO1 and 18S) 2 µL of DNA template was used. Thermal cycling conditions: 94 °C for 3 min; 45 cycles of 94 °C for 30 s, annealing temperature for 30 s, and 72 °C for 30 s; final extension at 72 °C for 10 min. Annealing temperatures of primers are listed in Table 4.
Table 4.
Conditions of PCR reactions as well as amplicon lengths for the different primer pairs. Tann = annealing temperature.
PCR products were extracted from agarose gels with the QIAquick Gel Extraction Kit (Qiagen, Inc.) and sequenced (both directions) at Microsynth Austria (Vienna, Austria) using the PCR primers.
2.6. Establishment of the eDNA PCR Primers
For eDNA analyses, primers for short fragments were designed. A partial sequence of the 18S gene was amplified with the primer pair Trem-18S-f/Trem-18S-r (Table 3; amplicon length 413–428 bp). Based on available mitochondrial genomes of Trichobilharzia species downloaded from GenBank and CO1 sequences generated in our own lab, specific PCR primers were developed for the species that could be plausibly expected for Upper Austria—T. franki, T. szidati, and T. regenti—allowing to amplify short fragments from these species to detect even highly degraded DNA in eDNA water samples. For each species, the primer pair consisted of ZDOE-COI-fw as a forward primer and a species-specific reverse-oriented eDNA primer (eDNA-franki-rv, eDNA-szidati-rv, eDNA-regenti-rv) (Table 3). These primer pairs yield a PCR product of 141 bp in length that lies outside the established CO1 barcoding region. This region of the CO1 gene was selected after preliminary tests of other primers (data not shown) because its high variation enabled differentiation and construction of species-specific primers, each amplifying the section from only one of the Trichobilharzia species. The primers were each tested with the extracted cercariae specimens of T. franki (Ra2-4-014), T. regenti (Rlag1-15-002), and T. szidati (Ls36-13-002) to ensure that they yield a PCR product only for the target species and not for the two remaining species. Later during the present study, after the finding of Trichobilharzia physellae in Upper Austria [23], a further species-specific reverse primer was designed for T. physellae. Subsequently, this primer was tested with the other three Trichobilharzia species, and the other species-specific primers were tested in T. physellae. After initial PCRs showed that the physellae primer also delivered results with T. szidati, we designed additional forward primers specific for T. physellae and for T. szidati (see Results). Furthermore, the eDNA primers were tested not only in PCRs with each of the available Trichobilharzia species but also with other trematode species, yielding no PCR products (data not shown).
2.7. Sequence Analysis
PCR products were purified using the QIAquick PCR Purification Kit (Qiagen, Hilden, Germany) according to the manufacturer’s protocol and sequenced in both directions at Microsynth Austria using the PCR primers plus additional sequencing primers (Table 3). The obtained electropherograms were checked with FinchTV 1.4.0 (Geospiza Inc.) and edited using GeneDoc 2.7.0 [38]. The edited sequences were subjected to a plausibility assessment by comparison with already published sequences using the BLAST search in the international database of the barcoding initiative BOLD (https://v4.boldsystems.org/ accessed on 6 February 2022) and the NCBI gene database GenBank (https://www.ncbi.nlm.nih.gov/genbank/ accessed on 6 February 2022).
Sequences generated from cercariae are registered in the BOLD database under the accession numbers NHBP008-21, NHBP010-21, NHBP014-21, NHBP016-21, and NHBP018-21, and in NCBI GenBank under the accession numbers OP347089–OP347093 (Table 2).
2.8. Detection of Trematode DNA in eDNA Samples from the Field
The DNA of each water sample was analysed three times by PCR with the species-specific primer pairs for T. franki, T. regenti, and T. szidati as well as once with the species-specific primers for T. physellae. The 3-fold repetition was conducted because false-negative results can occur in PCRs of eDNA samples if the amount of target DNA present is very small. If all three runs were negative, the eDNA extracts (from water samples taken in the field or lab) were also tested with the 18S primers to find possible evidence of other trematode species present. For all positive eDNA samples from the field, PCR products were sequenced (Table 5).
Table 5.
List of eDNA samples analysed. Ind. ID = laboratory number of the sample or individual; Locality numbers in parentheses, for eDNA samples from aquariums, indicate the water body from which the snails originated. For all aquarium samples, the column “sample date” gives the day of eDNA sampling (in the lab). “Volume” indicates the amount of water sampled. The column “CO1 Test T. szidati” indicates the number of times the species-specific eDNA primer produced a positive PCR result for the species of concern. Genetic identification based on NCBI-BLAST results (all above 96% identity).
2.9. eDNA from Aquarium Water
To answer the question how long cercariae are detectable via eDNA after their release from snails (and after the death of cercariae), water samples were taken with 0.45 µm Sterivex™ filter and syringes (as described above) from the middle of an aquarium (25 L) in which an individual of Lymnaea stagnalis had released T. szidati cercariae (not quantified) for one day (for setup of the aquariums see Section 2.2). Sampling of aquarium water (at room temperature, on average 23 °C) was repeated 14 times between 24 August 2020 (day 0) and 7 October 2020 (day 44). Two of these eDNA samples were taken before removal of the host snail from the aquarium on the 24 August (eDNA-5, eDNA-6; “aquarium sample” in Table 5) and thus served as positive controls in this experiment.
As with the eDNA samples taken from the field, several (3–6) PCR runs were conducted with the aquarium samples to detect false-negative results. PCR reactions with the primer pair specific for T. szidati (ZDOE-COI-fw/eDNA-szidati-rv) were performed with all the extracts, as this was the species released from the host snail. Additionally, each extract was tested once with the species-specific eDNA primers of T. franki and T. regenti to further test the specificity of the primers.
2.10. Sensitivity Tests
In another experiment performed to determine the smallest detectable amount of DNA in a water sample, dilution series of DNA were prepared. The concentration was measured by a Qubit fluorometer (Invitrogen). For this purpose, a PCR product (ZDOE-CO1-Fw/eDNA-szidati-rv and ZDOE-CO1-Fw/eDNA-franki-rv, respectively; 141 bp length) was prepared from two individuals: T. szidati Ls17-15-003 (concentration of PCR product: 7.98 ng/µL) and T. franki Ra13-16-002 (8.93 ng/µL) (Table 2). Furthermore, genomic DNA from these two individuals was used (Ls17-15-003, 0.0850 ng/µL; Ra13-16-002, 0.0830 ng/µL). For each template (both experiments: PCR products as well as genomic DNA), the dilution series consisted of six dilution steps (1/10–1/1,000,000), yielding seven DNA solutions each. For PCR, 1 µL of these DNA solutions was used as template. Species-specific primers were employed: ZDOE-CO1-Fw/eDNA-szidati-rv, ZDOE-CO1-Fw/eDNA-franki-rv). PCR with the dilution series of the PCR products was performed with 30 PCR cycles. PCR with genomic DNA (7 reactions each) was performed 1× with 30 cycles and 1× with 40 cycles.
3. Results
At seven localities in Lower and Upper Austria, a total of 13 eDNA samples were taken in 2019 and 2020 (Table 5). Our aim to compare the eDNA results with records of Trichobilharzia spp. in the field was hampered by the fact that only a few reports were available; this also corresponded with the lack of official reports on cercarial dermatitis in these two years.
The following four cases list snails collected in 2019 and 2020 that released Trichobilharzia cercariae (Table 6). (1) Cercariae tentatively assigned to T. szidati were released by Lymnaea stagnalis collected at locality 2 (Hagenauer Bucht 19 August 2019). Due to insufficient fixation, the cercariae were in poor condition. (2) Cercaria identified as T. franki were released by Radix auricularia collected at locality 1 (Reichersberger Au; 27 May 2019). (3) Cercariae identified as T. szidati were released by two Lymnaea stagnalis snails collected at locality 4 (Rekawinkel; 20 and 24 August 2020). (4) Cercariae identified as T. physellae were released by Physella acuta collected at locality 9 (Lake Pleschinger-See; 16 September 2020 and 11 November 2020). For details of the corresponding eDNA results, see Section 3.2.1.
Table 6.
List of Trichobilharzia species released from snails collected at four different sampling sites in the field during 2019 and 2020. The locality numbers correspond to the sampling sites in Table 1. * Trichobilharzia szidati cercaria that were only tentatively assigned due to insufficient fixation.
3.1. Establishment of the eDNA-PCR Primers
CO1 sequences (length of 1216 or 1263 bp) of five cercariae specimens were generated in the present study (Table 2). They formed the basis for eDNA primer design together with published sequences of seven species that were available at the time of the study: Trichobilharzia anseri, T. franki, T. ocellata (currently considered a synonym of T. szidati), T. physellae, T. regenti, T. stagnicolae, T. szidati).
Specific eDNA primers developed in the first phase of the present study for amplifying a 186 bp fragment of the mt CO1 gene (Table 3) were tested with DNA extracted from T. franki, T. szidati, and T. regenti. The cross tests performed with these primers showed clear species specificity. No PCR products could be obtained in the other Trichobilharzia species tested. Note that after the detection of T. physellae in Upper Austria, when test series were performed with eDNA primers designed for that species (primer pair ZDOE-COI-fw/eDNA-physellae-rv), certain problems arose. This indicates that a mismatch at the 3′–end of a primer is not sufficient to exclude species from amplification. Repeated tests with the other three species resulted in PCR products also in T. szidati despite a mismatch in the 3′-position of the primer eDNA-physellae-rv as well as additional internal mismatches (in combination with forward primer ZDOE-COI-fw). Thus, this primer pair could not prevent amplification in the non-target species. Subsequently, new forward primers with additional mismatches at the 3′–end were designed to further make the primers more specific (ZDOE-COI-fw_szidati, ZDOE-COI-fw_physellae). Tests with the new forward primers also yielded cross-species results (albeit with lower PCR success compared to the target species): ZDOE-COI-fw_physellae + eDNA-physellae-rv showed results also in T. szidati and T. regenti, ZDOE-COI-fw_szidati + eDNA-szidati-rv showed results also in T. physellae. Sequencing of the PCR products confirmed the authenticity of the sequence case (T. szidati or T. regenti, respectively). Moreover, it also showed that the non-matching 3′–base was present in both the forward as well as reverse primers while the sequence between the two primers was identical with the template tested in each case (T. szidati, T. physellae, or T. regenti). Thus, the amplification was not due to shorter primers (e.g., failure in primer production). This demonstrated that the T. physellae primers indeed amplified in those two other species as well as the T. szidati primer amplified in T. physellae. Interestingly, when we repeated these experiments with another DNA polymerase (Multiplex PCR Kit, Qiagen) there was no unspecific amplification. To summarize, there was a reproducible non-intentional amplification in some non-target species, which depended on the DNA polymerase used. It occurred with some (but not all) primer combinations, despite considerable mismatches (including the 3′–position) with the primer binding sites.
3.2. eDNA Experiments
Eighteen eDNA samples of water were taken from seven localities (Table 5). Thirteen eDNA samples were collected directly in the field (“water samples from field” in Table 5), four were obtained in the laboratory from water samples (without snails) brought from the field in bottles in which eDNA samples were taken in the lab (“water sample lab” in Table 5). One eDNA sample was taken from transport water ("transport water sample") containing a living Lymnaea stagnalis (which later released T. szidati cercariae). Furthermore, 15 eDNA samples were taken from aquariums in which snails were kept for some time (up to 24 hours), three of them when the snails were still present (“aquarium sample”) and 12 samples after having removed the snails (“aquarium test samples”).
3.2.1. eDNA Samples from the Field
Among all water samples from the field, four (eDNA-5, e-DNA-6, eDNA-7, eDNA-9) were positive for T. szidati using the species-specific CO1 primers (the maximum similarity values with BLAST search on 6 February 2022 were 100% for all four samples). They all were from sample sites of one water body (locality 4, Rekawinkel, Lower Austria). Moreover, the transport water sample from this locality delivered a positive result for T. szidati. Besides T. szidati, a second species—T. physellae—was demonstrated with eDNA analyses (similarity values 100%). In September 2020, after completion of the initially planned analyses, Trichobilharzia physellae was detected by hatching experiments in a snail collected at locality 9 (Lake Pleschinger-See). Since this was the first record of the species in Europe [23], it had not been covered by our experimental design. The subsequent construction of eDNA primers specific for T. physellae yielded a positive result from the eDNA sample taken from Lake Pleschinger-See, yet not from all samples. The eDNA samples from the field taken at the first day of record of T. physellae proved negative. From the four water samples transported to the lab on 16 October 2021, only one gave a positive result (eDNA-27).
The detection of an unexpected new species resulted in problems and reconsideration of our experimental design described above. Moreover, testing the physellae-specific reverse primer (Primer-Pair: ZDOE-COI-fw/eDNA-physellae-rv) with the eDNA samples yielded a PCR product in one sample that had been positive for T. szidati (sample eDNA-5). This confirmed the problems with species specificity mentioned above. Sequencing of the PCR products showed that the physellae-specific reverse primer had amplified a sequence of T. szidati (the species observed in that water body).
The PCR tests with the primers specific for T. regenti and T. franki were negative in all eDNA samples from the field. Importantly, even the eDNA sample from 2019 (4-W-8) from the bay Hagenauer Bucht at the lower Inn River (locality code 2) did not yield a PCR product with the Trichobilharzia-specific CO1 primers. At this locality, cercariae, presumably Trichobilharzia szidati, had been detected in 2019 by hatching experiments. Unfortunately, they could not be identified unambiguously due to insufficient fixation of the material. Nonetheless, despite of the evidence for the presence of Trichobilharzia sp. at that locality at that time, the corresponding eDNA sample did not provide evidence of any of the four species.
In summary (with the exception of sample 4-W-8 from 2019), the results of these eDNA analyses were mostly consistent with the rare detection of Trichobilharzia cercariae in the various water bodies.
Besides using CO1 as a marker sequence, we also tested universal 18S primers. They indicated the presence of trematodes in 8 of 16 water samples. The 18S sequences thus obtained were blasted in GenBank and BOLD. Some sequences yielded a single species as best hit based on 18S, i.e., Diplostomum pseudospathaceum (100% similarity), Echinostoma revolutum (99.72%), Opisthioglyphe ranae (98.68%), and Parastrigea robusta (100%). In sample eDNA-11, the identification to T. szidati based on CO1 was confirmed at genus level by 18S results. The BLAST similarity scores of 100% for various species of Trichobilharzia indicated for this genus a lack of taxonomic resolution at species level with the 18S marker sequence. The 18S sequences of sample 3-W-14 gave similarity values of up to 96% for species of the superorder Plagiorchiida. In addition, three samples yielded a closest hit with Sminthurides aquaticus (98.92–100%), a springtail species (Table 5; all similarity values from GenBank on 6 February 2022).
3.2.2. eDNA from Aquarium Water
Three samples were considered as positive controls because the host snail L. stagnalis was still present in the aquarium when the eDNA sample was taken (eDNA-5, eDNA-6, eDNA-11). All PCR reactions with the primers for T. szidati were positive in these samples, whereas the primers for T. franki and T. regenti yielded no PCR products, confirming that these primers were species-specific (these experiments were performed before the first record of T. physellae in Austria).
To test how long cercariae are detectable in aquarium water after removal of the host snail (from which the cercariae were released), 12 eDNA samples were taken at different times (Table 5). The first seven samples (first week after snail removal) tested positively for T. szidati in all three replicates. Of the samples taken thereafter, only the last one (eDNA-24, from 7 October 2020; day 44) was positive in one of the three PCR tests. The four negative ones (eDNA-20–eDNA-23) were additionally tested three times more with the CO1 primers for T. szidati. None of these subsequent PCR reactions resulted in PCR products. All controls with the eDNA primers for T. franki and T. regenti were performed with all water samples from the field and proved negative.
3.3. Sensitivity Tests
Dilution series of genomic DNA of two individuals (T. szidati Ls17-15-003; T. franki Ra13-16-002) were tested in a PCR with 30 thermal cycles. Amplification of the targeted PCR product was accomplished up to a dilution of 10−2, corresponding to a minimum concentration of 0.85 pg/µL for Ls17-15-003 and 0.83 pg/µL for Ra13-16-002. PCR with 40 cycles enabled amplification in dilutions up to 10−3 (corresponding to 0.085 pg/µL for Ls17-15-003 and 0.083 pg/µL for Ra13-16-002).
The second kind of sensitivity test was performed using the PCR fragments generated from the two individuals as template, i.e., it was a re-amplification (ZDOE-CO1-Fw/eDNA-szidati-rv and ZDOE-CO1-Fw/eDNA-franki-rv, respectively; 141 bp length). PCR with 40 cycles enabled amplification in the entire dilution series, i.e., gave positive results up to a dilution of 10−6 (corresponding to a minimum detectable concentration of 0.008 pg/µL for Ls17-15-003 and of 0.009 pg/µL for Ra13-16-002).
3.4. Cercariae Release Experiments
Cercarial release experiments with a Lymnaea stagnalis (from Rekawinkel, Lower Austria) infected with T. szidati showed that the snail released cercariae for six weeks. The snail expelled cercariae each time, which were fixed in 80% EtOH. A rough estimate of the released number was made by counting cercariae in small volumes and extrapolating. The estimates of released cercariae within 12 h were 14,000 (2 September 2020), 5000 (16 September 2020), and 900 (6 October 2020). By 16 October 2020, the host snail had died.
4. Discussion
4.1. Detection of Trichobilharzia Species—Correspondence between Observations and eDNA Tests
The results of the PCR detection of Trichobilharzia species in eDNA samples from the seven water bodies investigated yielded a positive result in 4 out of 18 samples. Trichobilharzia szidati was detected in water of a small pond (Rekawinkel, Lower Austria), whereas T. physellae was found in a water sample from Lake Pleschinger-See in Upper Austria (Table 5). This result should be considered in the context of the presumably scarce occurrence of Trichobilharzia species, as shown by the lack of reports on cercarial dermatitis in the years 2019–2020. Assuming similar transmission cycles of Trichobilharzia across Europe, previous studies on trematode diversity seem to confirm a low prevalence [39,40,41,42,43]. Another case occurred in the bay Hagenauer Bucht of the lower Inn River. Trichobilharzia cercariae had been detected there by hatching experiments with L. stagnalis collected in locality 2 in 2019, but could not be identified (morphologically) due to insufficient fixation. The analysis of the corresponding eDNA sample (from 2019; conducted in 2020) provided no evidence for the presence of Trichobilharzia. The presumably limited stability of eDNA samples after long-term storage as a possible explanation [44,45,46], however, is contradicted by a positive PCR product obtained with 18S primers in four samples from 2019. This indicates the presence of DNA, but not (or not enough) DNA from one of the Trichobilharzia target species. Another explanation might be that the corresponding eDNA sampling point in the water body (in the riparian area) was several meters away from where the host snail was collected. Two more eDNA samples taken in 2020 at the same position also failed to give a result for trematode DNA (eDNA-1 and eDNA-2). This implies that, generally, a considerable number of sampling sites would be needed to find eDNA of the target species with a high probability. For example, an eDNA study on carp showed the necessity of a high sampling effort if only little environmental DNA is present [47]. The mostly negative results for the eDNA samples from Lake Pleschinger-See, the water body where T. physellae could be detected via hatching tests, further confirmed the interpretation of a certain stochasticity of eDNA detection [23]. Only two of the samples proved positive, of which one contained T. physellae and the other Diplostomum pseudospathaceum.
In summary, eDNA assays based on CO1 were always negative in localities where no cercariae of Trichobilharzia species were found. However, using 18S primers, several of the samples delivered eDNA detection of other trematode taxa even though the 18S fragment was considerably longer than the CO1 fragments. Identification to species level is unreliable based on the 18S marker sequence due both to insufficient reference data and lack of taxonomic resolution.
The finding that the PCRs with species-specific eDNA primers designed for T. franki and T. regenti were negative was interpreted as confirmation of their specificity. Nonetheless, the design and testing of species-specific primers for T. physellae, after the species had been discovered in Austria, should be treated with caution. Although we still cannot explain why the primers for T. physellae amplified (despite several mismatches) in other species, too, we conclude that presumed species-specificity of primers may not be guaranteed. A detection system based on species-specific primers developed on a presumably known species inventory may be compromised when new (or formerly not detected) species appear. We therefore recommend checking PCR results by sequencing PCR products, or including other control procedures such as nested PCR, ensuring that species identification is reliable. Future experiments (e.g., mixing two species into one PCR reaction) should test in more detail how the properties of various DNA polymerases effect primer specificity.
4.2. Stochasticity of eDNA Detection
In general, our results underline the potential of the eDNA approach. However, we observed considerable stochasticity in detection success. This is in accordance with reports by Jothikumar et al. 2015 [30] about eDNA samples taken from lakes with suspected cases of swimmer’s itch. That analysis yielded only 32 % positive results for the presence of avian schistosomes. Similarly, in the study by Sato et al. 2018 [25] on Schistosoma mansoni, only one out of 14 environmental water samples was positive. In another study, however, S. mansoni was detected in water samples from four of five sites in central Kenya with known ongoing transmission [28]. Wind may be an important factor for the distribution of cercariae in a water body [48]. Those authors found that wind direction, wind speed, and time of day were the best predictors for the risk of contracting swimmer’s itch. Thus, even sampling at the same site over time might yield varying results. Accordingly, the location (and perhaps the circumstances) of sampling may be relevant, even if the water body is relatively small as in the case of site 11 (pond in Rekawinkel, about 300 m²). Of the four sampling sites in this water body, only two were positive (and here only 1 and 2 of each of the three PCR replicates, respectively). Interestingly, one (eDNA-7) of the positive samples was collected from the shore, the other one (eDNA-9) from the middle of the pond. In contrast, all PCR tests (six replicates each) from the other two sample collection sites were negative (one, eDNA-8, also from the shore, the other, eDNA-10, in a marshy area of the pond). The sample in the marshy area was possibly affected by the large amount of suspended sediment (only 100 mL water filtered). This was not the case, however, for the other negative sampling site (200 mL filtered water, compared to 120 and 170 mL for the two positive samples). This demonstrates, albeit with a small sample, that the sampling regime (position in the lake), the turbidity of the water, as well as the number of PCR repetitions can affect the results.
The fact that—among all the PCR reactions carried out with the five samples from Lake Pleschinger-See (where T. physellae cercariae were found, see [23])—only one yielded a positive PCR result for T. physellae demonstrates once again that PCR detection has a high stochasticity. One way to improve detection success would be to increase the amount of target eDNA by sampling larger volumes of water by pooling multiple eDNA filters [49]. However, besides a possibly low concentration of Trichobilharzia in the lake, our results reflect recent experiences of other studies reporting a heterogeneous distribution of eDNA in natural waters [6,50]. This somewhat relativizes a major expected advantage of eDNA detection compared to conventional screening of host snails by releasing experiments. Without knowledge of the specific zones that the host snails inhabit, their detection might be missed in still waters.
4.3. DNA Degradation and Sensitivity of PCR Detection
Our laboratory experiments with aquarium water showed that cercariae could still be detected with high reliability after one week. It remains open whether whole cercariae, their remains, or suspended DNA were sucked into the filters during sampling. Note, however, that this issue also arises with every eDNA sampling from waters in the field. The results were consistent because the samples of the first week were positive and the samples taken at a later time were, with one exception, negative. This result is consistent with a previous study that detected cercariae that had been shed by snails 8 days earlier [28]. Nonetheless, it is somewhat surprising that the very last sample provided a positive result (1 out of 3 PCRs). Here, a contamination cannot be ruled out completely. Another possible explanation is that the water had cleared up in the last week of sampling, enabling more water to be sampled through the filter. The negative results of the aquarium samples after 2 weeks and the single positive result of the last sample taken (after 6 weeks) suggest that a PCR response at very low concentrations is subject to a strong random factor. This result is interesting with regard to the reported degradation times for eDNA from different species, which can vary between 72 h and 25 days [6,51,52,53]. Since a positive result was obtained at extremely low concentrations in the PCR experiments with the dilution series, similar to those found by Kane et al. 2013 [29], a major factor potentially negatively influencing the results of eDNA analyses might be inhibitory substances and DNA from other organisms (e.g., algae, bacteria). These could be co-extracted from the eDNA filter (in the present case from the eDNA from aquarium water) and exert a competitive or inhibitory effect in the PCR. In this sense, the PCR experiments with the dilution series (which represent an ideal system) cannot be transferred directly to natural aquatic samples. In recent years, many factors have been shown to influence the rate of eDNA degradation in both limnic and marine habitats. For example, evidence is available that high water temperature boosts the activity of the microbiome and thus increases the degradation rate of both mitochondrial (e.g., CO1) and nuclear DNA (e.g., 18S) [4,54,55]. Accordingly, some of our negative results might be explained by the relatively warm water temperature of around 23 °C in our experiment. There also appears to be a relationship between degradation rate and the overall biomass of the water body studied. The more biomass a water body contains, the higher the degradation rate [4,54]. This could also have influenced our samples because the pond near Rekawinkel and the bay Hagenauer Bucht in particular contained abundant suspended matter, which also meant that less water was pressed through the filter while sampling. The pH as well as UV-B radiation also have an effect because degradation is faster in acidic environments and at high UV-B levels [5,56].
4.4. eDNA Barcoding for the Detection of Trichobilharzia in Practice
In recent years, studies in a variety of animal groups demonstrated the effective use of eDNA to detect animals/species groups that are otherwise difficult to detect or to monitor [57,58,59]. Our results confirmed the possibility to detect cercariae in water samples as well as the stability of DNA in aquarium water over several days, but also demonstrated the uncertainty of detection regarding the location of sampling. This could also be true for waters in the field, which raises certain caveats regarding the applicability for monitoring and assessment of current situations. The single observation of a laboratory-kept L. stagnalis individual shedding cercariae in large quantities over six weeks suggests that even when access to a water body is approved after a negative eDNA test, high levels of cercariae can reappear very quickly. For example, warm weather conditions could once again boost cercariae output. Therefore—even if our conclusions are currently based on few samples and data—both a warning system as well as an all-clear system based on eDNA do not seem reasonable (at least without an immense sampling effort). This, however, does not mean that eDNA tools cannot be usefully applied or expanded. Environmental DNA as a tool for identifying the causative agent in water bodies (where cercarial dermatitis was reported) is no doubt extremely helpful instead of collecting and examining (possibly protected) snails. In future experiments, we aim to optimize our species-specific primers and test them with the qPCR method.
5. Conclusions
We demonstrated that it is possible to detect trematodes in eDNA water samples, and in combination with matching primers, a barcoding approach is also plausible. Nonetheless, uncritical detection with supposedly species-specific primers based solely on a PCR product of the desired size is short-sighted and risky. This is largely because species specificity may not be guaranteed, especially if the species inventory of an area is incompletely known. This makes sequencing of every positive result still advisable. The collection of eDNA samples is additionally hampered in the case of potentially locally restricted tiny animals such as cercariae, as this creates stochasticity in the results. At the same time, however, the relatively long detectability of at least one week would enable recording the species population of trematodes of a water body through regular but not excessive sample collection. This calls for further studies, especially for improving the localisation of sampling.
Author Contributions
Conceptualisation: N.H., J.B., E.H., C.H., S.R., H.S. and N.U.S.; Field work: N.H., E.H., C.H., S.R., H.S. and N.U.S.; Morphological investigations: N.H., J.B., C.H. and H.S.; Laboratory procedures: N.H. and S.R.; Data analysis, N.H., C.H., H.S. and E.H.; contributed intellectually to the interpretation and discussion of results: N.H., J.B., E.H, C.H., S.R., H.S. and N.U.S.; Writing—Original draft preparation: N.H. and E.H.; Writing—Review & Editing, N.H., J.B., E.H, C.H., S.R., H.S. and N.U.S.; Project administration, E.H. and N.U.S.; Funding acquisition, E.H., C.H. and N.U.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received partial funding by the Institutional Grant Charles University Cooperation Biology 2022–2026 and by the Grant of Ministry of Education, Youth and Sports of the Czech Republic (8J21AT011). Furthermore, the study was supported by WTZ Programme (Scientific & Technological Cooperation) (CZ18/2021), by the Upper Austrian Provincial Government (Reference number: WW-583711-2019/KR), and the “Freunde des Naturhistorischen Museums Wien”.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Raw and processed data will be shared on personal request directly from the corresponding author.
Acknowledgments
We thank Wolfgang Heinisch and Hubert Blatterer (Department of Water Management, Upper Austria Provincial Government) for initiating and promoting the Upper Austrian cercaria project in the course of which the first field trips took place and the eDNA study was initiated. We are grateful to the Upper Austrian Provincial Government and the “Freunde des Naturhistorischen Museums Wien” for financial support. Thanks are also due to Julia Schindelar, Marcia Sittenthaler, and Alexandra Wanka for technical assistance in the lab. Finally, we thank Michael Stachowitsch for scientific English copyediting.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pawlowski, J.; Apothéloz-Perret-Gentil, L.; Mächler, E.; Altermatt, F. Anwendung von eDNA-Methoden in Biologischen Untersuchungen und bei der Biologischen Bewertung von Aquatischen Ökosystemen: Richtlinien; Bundesamt für Umwelt: Bern, Switzerland, 2020. [Google Scholar]
- Taberlet, P.; Bonin, A.; Zinger, L.; Coissac, E. Environmental DNA: For Biodiversity Research and Monitoring; Online Edi.; Oxford University Press: Oxford, UK, 2018; ISBN 9780198767220. [Google Scholar]
- Harrison, J.B.; Sunday, J.M.; Rogers, S.M. Predicting the fate of eDNA in the environment and implications for studying biodiversity. Proc. R. Soc. B Biol. Sci. 2019, 286, 20191409. [Google Scholar] [CrossRef] [PubMed]
- Jo, T.; Arimoto, M.; Murakami, H.; Masuda, R.; Minamoto, T. Estimating shedding and decay rates of environmental nuclear DNA with relation to water temperature and biomass. Environ. DNA 2020, 2, 140–151. [Google Scholar] [CrossRef]
- Strickler, K.M.; Fremier, A.K.; Goldberg, C.S. Quantifying effects of UV-B, temperature, and pH on eDNA degradation in aquatic microcosms. Biol. Conserv. 2015, 183, 85–92. [Google Scholar] [CrossRef]
- Troth, C.R.; Sweet, M.J.; Nightingale, J.; Burian, A. Seasonality, DNA degradation and spatial heterogeneity as drivers of eDNA detection dynamics. Sci. Total Environ. 2021, 768, 144466. [Google Scholar] [CrossRef]
- Bochove, K.; Bakker, F.T.; Beentjes, K.K.; Hemerik, L.; Vos, R.A.; Gravendeel, B. Organic matter reduces the amount of detectable environmental DNA in freshwater. Ecol. Evol. 2020, 10, 3647–3654. [Google Scholar] [CrossRef]
- Loker, E.S.; DeJong, R.J.; Brant, S.V. Scratching the Itch: Updated Perspectives on the Schistosomes Responsible for Swimmer’s Itch around the World. Pathogens 2022, 11, 587. [Google Scholar] [CrossRef]
- Auer, H.; Aspöck, H. “Vogelbilharzien” als Erreger einer Hautkrankheit: Die Zerkarien-Dermatitis. Denisia 2002, 6, 321–331. [Google Scholar]
- Sattmann, H.; Hörweg, C.; Konecny, R. Zerkariendermatitis in Österreich—Rückblick und Perspektiven. Denisia 2004, 13, 457–461. [Google Scholar]
- Horák, P.; Mikeš, L.; Lichtenbergová, L.; Skála, V.; Soldánová, M.; Brant, S.V. Avian Schistosomes and Outbreaks of Cercarial Dermatitis. Clin. Microbiol. Rev. 2015, 28, 165–190. [Google Scholar] [CrossRef]
- Kolářová, L.; Horák, P.; Skírnisson, K.; Marečková, H.; Doenhoff, M. Cercarial Dermatitis, a Neglected Allergic Disease. Clin. Rev. Allergy Immunol. 2013, 45, 63–74. [Google Scholar] [CrossRef]
- Soldánová, M.; Selbach, C.; Kalbe, M.; Kostadinova, A.; Sures, B. Swimmer’s itch: Etiology, impact, and risk factors in Europe. Trends Parasitol. 2013, 29, 65–74. [Google Scholar] [CrossRef]
- Brant, S.V.; Loker, E.S. Discovery-based studies of schistosome diversity stimulate new hypotheses about parasite biology. Trends Parasitol. 2013, 29, 449–459. [Google Scholar] [CrossRef]
- Jouet, D.; Kolářová, L.; Patrelle, C.; Ferté, H.; Skírnisson, K. Trichobilharzia anseri n. sp. (Schistosomatidae: Digenea), a new visceral species of avian schistosomes isolated from greylag goose (Anser anser L.) in Iceland and France. Infect. Genet. Evol. 2015, 34, 298–306. [Google Scholar] [CrossRef]
- Christiansen, A.Ø.; Olsen, A.; Buchmann, K.; Kania, P.W.; Nejsum, P.; Vennervald, B.J. Molecular diversity of avian schistosomes in Danish freshwater snails. Parasitol. Res. 2016, 115, 1027–1037. [Google Scholar] [CrossRef]
- Neuhaus, W. Biologie und Entwicklung von Trichobilharzia szidati n. sp. (Trematoda, Schistosomatidae), einem Erreger von Dermatitis beim Menschen. Z. Parasitenkd. 1952, 15, 203–266. [Google Scholar] [CrossRef]
- Horák, P.; Kolářová, L.; Dvořák, J. Trichobilharzia regenti n. sp. (Schistosomatidae, Bilharziellinae), a new nasal schistosome from Europe. Parasite 1998, 5, 349–357. [Google Scholar] [CrossRef]
- Müller, V.; Kimmig, P. Trichobilharzia franki n. sp.—Die Ursache für Badedermatitiden in südwestdeutschen Baggerseen [Trichobilharzia franki n. sp.—A causative agent of swimmer’s itch in south-western Germany]. Appl. Parasitol. 1994, 35, 12–31. [Google Scholar]
- Simon-Martin, F.; Simon-Vicente, F. The life cycle of Trichobilharzia salmanticensis n. sp. (Digenea: Schistosomatidae), related to cases of human dermatitis. Res. Rev. Parasitol. 1999, 59, 13–18. [Google Scholar]
- Kolářová, L.; Skírnisson, K.; Ferté, H.; Jouet, D. Trichobilharzia mergi sp. nov. (Trematoda: Digenea: Schistosomatidae), a visceral schistosome of Mergus serrator (L.) (Aves: Anatidae). Parasitol. Int. 2013, 62, 300–308. [Google Scholar] [CrossRef]
- Talbot, S.B. Studies on schistosome dermatitis: II. Morphological and life history studies on three dermatitis-producing schistosome cercariae, C. elvae Miller, 1923, C. stagnicolae n. sp., and C. physellae n. sp. Am. J. Epidemiol. 1936, 23, 372–384. [Google Scholar] [CrossRef]
- Helmer, N.; Blatterer, H.; Hörweg, C.; Reier, S.; Sattmann, H.; Schindelar, J.; Szucsich, N.U.; Haring, E. First Record of Trichobilharzia physellae (Talbot, 1936) in Europe, a Possible Causative Agent of Cercarial Dermatitis. Pathogens 2021, 10, 1473. [Google Scholar] [CrossRef] [PubMed]
- McPhail, B.A.; Froelich, K.; Reimink, R.L.; Hanington, P.C. Simplifying Schistosome Surveillance: Using Molecular Cercariometry to Detect and Quantify Cercariae in Water. Pathogens 2022, 11, 565. [Google Scholar] [CrossRef]
- Sato, M.O.; Rafalimanantsoa, A.; Ramarokoto, C.; Rahetilahy, A.M.; Ravoniarimbinina, P.; Kawai, S.; Minamoto, T.; Sato, M.; Kirinoki, M.; Rasolofo, V.; et al. Usefulness of environmental DNA for detecting Schistosoma mansoni occurrence sites in Madagascar. Int. J. Infect. Dis. 2018, 76, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Rudko, S.P.; Turnbull, A.; Reimink, R.L.; Froelich, K.; Hanington, P.C. Species-specific qPCR assays allow for high-resolution population assessment of four species avian schistosome that cause swimmer’s itch in recreational lakes. Int. J. Parasitol. Parasites Wildl. 2019, 9, 122–129. [Google Scholar] [CrossRef]
- Alzaylaee, H.; Collins, R.A.; Shechonge, A.; Ngatunga, B.P.; Morgan, E.R.; Genner, M.J. Environmental DNA-based xenomonitoring for determining Schistosoma presence in tropical freshwaters. Parasit. Vectors 2020, 13, 63. [Google Scholar] [CrossRef]
- Sengupta, M.E.; Hellström, M.; Kariuki, H.C.; Olsen, A.; Thomsen, P.F.; Mejer, H.; Willerslev, E.; Mwanje, M.T.; Madsen, H.; Kristensen, T.K.; et al. Environmental DNA for improved detection and environmental surveillance of schistosomiasis. Proc. Natl. Acad. Sci. USA 2019, 116, 8931–8940. [Google Scholar] [CrossRef] [PubMed]
- Kane, R.A.; Stothard, J.R.; Rollinson, D.; Leclipteux, T.; Evraerts, J.; Standley, C.J.; Allan, F.; Betson, M.; Kaba, R.; Mertens, P.; et al. Detection and quantification of schistosome DNA in freshwater snails using either fluorescent probes in real-time PCR or oligochromatographic dipstick assays targeting the ribosomal intergenic spacer. Acta Trop. 2013, 128, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Jothikumar, N.; Mull, B.J.; Brant, S.V.; Loker, E.S.; Collinson, J.; Secor, W.E.; Hill, V.R. Real-Time PCR and Sequencing Assays for Rapid Detection and Identification of Avian Schistosomes in Environmental Samples. Appl. Environ. Microbiol. 2015, 81, 4207–4215. [Google Scholar] [CrossRef] [PubMed]
- Rudko, S.P.; Reimink, R.L.; Froelich, K.; Gordy, M.A.; Blankespoor, C.L.; Hanington, P.C. Use of qPCR-Based Cercariometry to Assess Swimmer’s Itch in Recreational Lakes. Ecohealth 2018, 15, 827–839. [Google Scholar] [CrossRef]
- Soldánová, M.; Selbach, C.; Sures, B. The Early Worm Catches the Bird? Productivity and Patterns of Trichobilharzia szidati Cercarial Emission from Lymnaea stagnalis. PLoS ONE 2016, 11, e0149678. [Google Scholar] [CrossRef]
- Al-Jubury, A.; Kania, P.W.; Bygum, A.; Buchmann, K. Temperature and light effects on Trichobilharzia szidati cercariae with implications for a risk analysis. Acta Vet. Scand. 2020, 62, 54. [Google Scholar] [CrossRef] [PubMed]
- Horák, P.; Kolářová, L.; Adema, C.M. Biology of the schistosome genus Trichobilharzia. Adv. Parasitol. 2002, 52, 155–233. [Google Scholar] [CrossRef] [PubMed]
- Reier, S.; Haring, E.; Billinger, F.; Blatterer, H.; Duda, M.; Gorofsky, C.; Grasser, H.-P.; Heinisch, W.; Hörweg, C.; Kruckenhauser, L.; et al. First confirmed record of Trichobilharzia franki Müller & Kimmig, 1994, from Radix auricularia (Linnaeus, 1758) for Austria. Parasitol. Res. 2020, 119, 4135–4141. [Google Scholar] [CrossRef] [PubMed]
- Folmer, O.; Black, M.B.; Hoeh, W.R.; Lutz, R.A.; Vrijenhoek, R.C. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Hohensee, N. Digenean Trematodes in Freshwater Snails in the Surroundings of Vienna with a Focus on Species Involved in Human Infections; University of Vienna: Vienna, Austria, 2016. [Google Scholar]
- Nicholas, K.B.; Nicholas, H.B.J. GeneDoc: A tool for editing and annotating multiple sequence alignments. Embnet. News 1997, 4, 1–4. [Google Scholar]
- Al-Jubury, A.; Duan, Y.; Kania, P.W.; Tracz, E.S.; Bygum, A.; von Jørgensen, L.G.; Horák, P.; Buchmann, K. Avian schistosome species in Danish freshwater lakes: Relation to biotic and abiotic factors. J. Helminthol. 2021, 95, e22. [Google Scholar] [CrossRef]
- Gaub, L.; Sattmann, H.; Hörweg, C.; Walochnik, J. Molecular validation of species determination of larval trematodes from freshwater snail hosts in Austria, with special emphasis on the genus Trichobilharzia Skrjabin & Zakharow, 1920. Arianta 2020, 8, 13–19. [Google Scholar]
- Duan, Y.; Al-Jubury, A.; Kania, P.W.; Buchmann, K. Trematode diversity reflecting the community structure of Danish freshwater systems: Molecular clues. Parasit. Vectors 2021, 14, 43. [Google Scholar] [CrossRef]
- Gordy, M.A.; Hanington, P.C. A fine-scale phylogenetic assessment of digenean trematodes in central Alberta reveals we have yet to uncover their total diversity. Ecol. Evol. 2019, 9, 3153–3238. [Google Scholar] [CrossRef]
- Selbach, C.; Soldánová, M.; Feld, C.K.; Kostadinova, A.; Sures, B. Hidden parasite diversity in a European freshwater system. Sci. Rep. 2020, 10, 2694. [Google Scholar] [CrossRef]
- Mauvisseau, Q.; Halfmaerten, D.; Neyrinck, S.; Burian, A.; Brys, R. Effects of preservation strategies on environmental DNA detection and quantification using ddPCR. Environ. DNA 2021, 3, 815–822. [Google Scholar] [CrossRef]
- Allison, M.J.; Round, J.M.; Bergman, L.C.; Mirabzadeh, A.; Allen, H.; Weir, A.; Helbing, C.C. The effect of silica desiccation under different storage conditions on filter-immobilized environmental DNA. BMC Res. Notes 2021, 14, 106. [Google Scholar] [CrossRef] [PubMed]
- Spens, J.; Evans, A.R.; Halfmaerten, D.; Knudsen, S.W.; Sengupta, M.E.; Mak, S.S.T.; Sigsgaard, E.E.; Hellström, M. Comparison of capture and storage methods for aqueous macrobial eDNA using an optimized extraction protocol: Advantage of enclosed filter. Methods Ecol. Evol. 2017, 8, 635–645. [Google Scholar] [CrossRef]
- Furlan, E.M.; Gleeson, D.; Wisniewski, C.; Yick, J.; Duncan, R.P. eDNA surveys to detect species at very low densities: A case study of European carp eradication in Tasmania, Australia. J. Appl. Ecol. 2019, 56, 2505–2517. [Google Scholar] [CrossRef]
- Sckrabulis, J.P.; Flory, A.R.; Raffel, T.R. Direct onshore wind predicts daily swimmer’s itch (avian schistosome) incidence at a Michigan beach. Parasitology 2020, 147, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Hunter, M.E.; Ferrante, J.A.; Meigs-Friend, G.; Ulmer, A. Improving eDNA yield and inhibitor reduction through increased water volumes and multi-filter isolation techniques. Sci. Rep. 2019, 9, 5259. [Google Scholar] [CrossRef]
- Beentjes, K.K.; Speksnijder, A.G.C.L.; Schilthuizen, M.; Hoogeveen, M.; van der Hoorn, B.B. The effects of spatial and temporal replicate sampling on eDNA metabarcoding. PeerJ 2019, 7, e7335. [Google Scholar] [CrossRef]
- Barnes, M.A.; Turner, C.R.; Jerde, C.L.; Renshaw, M.A.; Chadderton, W.L.; Lodge, D.M. Environmental Conditions Influence eDNA Persistence in Aquatic Systems. Environ. Sci. Technol. 2014, 48, 1819–1827. [Google Scholar] [CrossRef]
- Dejean, T.; Valentini, A.; Duparc, A.; Pellier-Cuit, S.; Pompanon, F.; Taberlet, P.; Miaud, C. Persistence of Environmental DNA in Freshwater Ecosystems. PLoS ONE 2011, 6, e23398. [Google Scholar] [CrossRef]
- Pilliod, D.S.; Goldberg, C.S.; Arkle, R.S.; Waits, L.P. Factors influencing detection of eDNA from a stream-dwelling amphibian. Mol. Ecol. Resour. 2014, 14, 109–116. [Google Scholar] [CrossRef]
- Jo, T.; Murakami, H.; Yamamoto, S.; Masuda, R.; Minamoto, T. Effect of water temperature and fish biomass on environmental DNA shedding, degradation, and size distribution. Ecol. Evol. 2019, 9, 1135–1146. [Google Scholar] [CrossRef]
- Goldberg, C.S.; Strickler, K.M.; Fremier, A.K. Degradation and dispersion limit environmental DNA detection of rare amphibians in wetlands: Increasing efficacy of sampling designs. Sci. Total Environ. 2018, 633, 695–703. [Google Scholar] [CrossRef]
- Seymour, M.; Durance, I.; Cosby, B.J.; Ransom-Jones, E.; Deiner, K.; Ormerod, S.J.; Colbourne, J.K.; Wilgar, G.; Carvalho, G.R.; de Bruyn, M.; et al. Acidity promotes degradation of multi-species environmental DNA in lotic mesocosms. Commun. Biol. 2018, 1, 4. [Google Scholar] [CrossRef] [PubMed]
- Balasingham, K.D.; Walter, R.P.; Mandrak, N.E.; Heath, D.D. Environmental DNA detection of rare and invasive fish species in two Great Lakes tributaries. Mol. Ecol. 2018, 27, 112–127. [Google Scholar] [CrossRef]
- Dougherty, M.M.; Larson, E.R.; Renshaw, M.A.; Gantz, C.A.; Egan, S.P.; Erickson, D.M.; Lodge, D.M. Environmental DNA (eDNA) detects the invasive rusty crayfish Orconectes rusticus at low abundances. J. Appl. Ecol. 2016, 53, 722–732. [Google Scholar] [CrossRef] [PubMed]
- Ushio, M.; Murata, K.; Sado, T.; Nishiumi, I.; Takeshita, M.; Iwasaki, W.; Miya, M. Demonstration of the potential of environmental DNA as a tool for the detection of avian species. Sci. Rep. 2018, 8, 4493. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).