
Warmer and Poorer: The Fate of Alpine Calcareous Grasslands in Central Apennines (Italy)

 ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
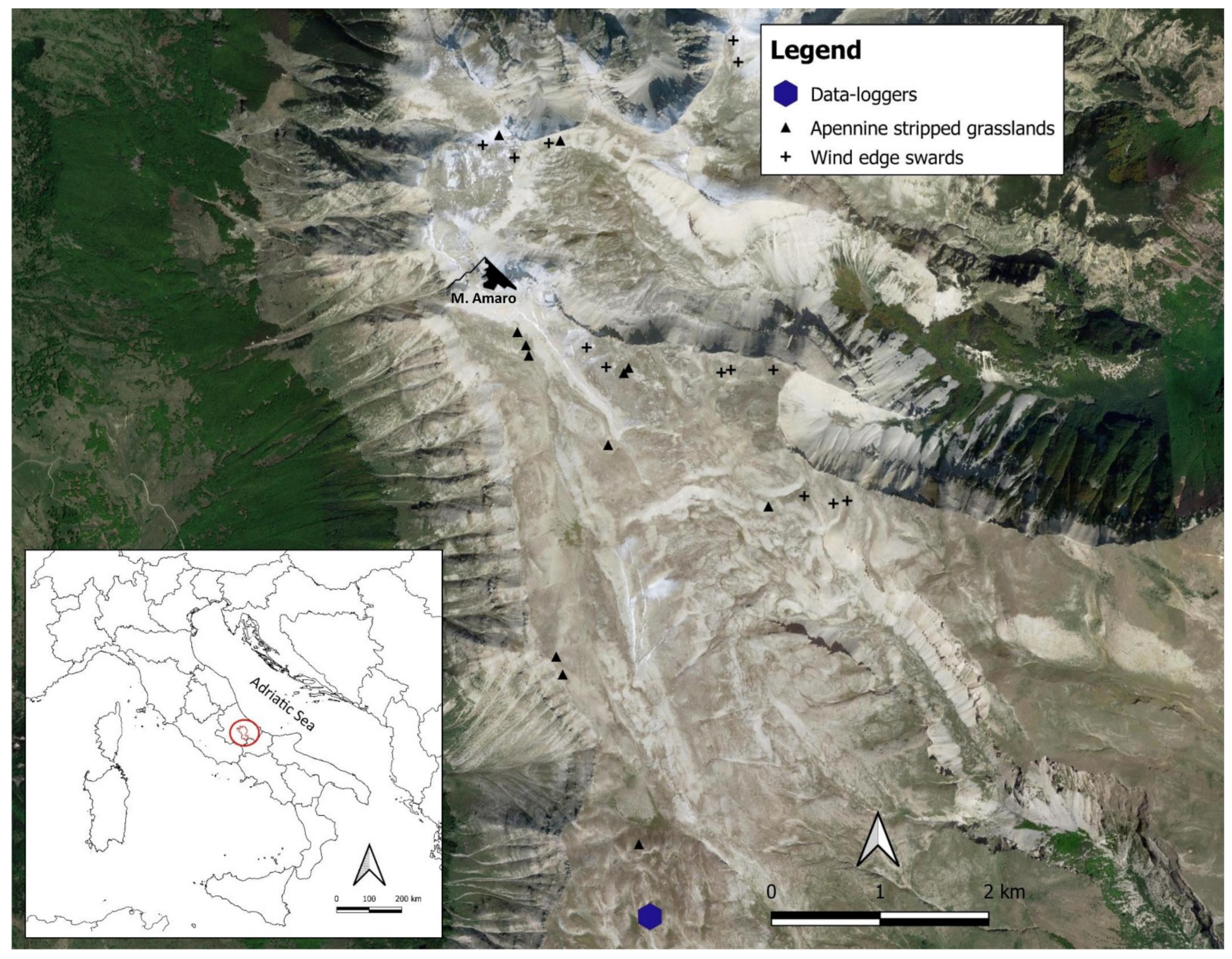
2.1. Study Area
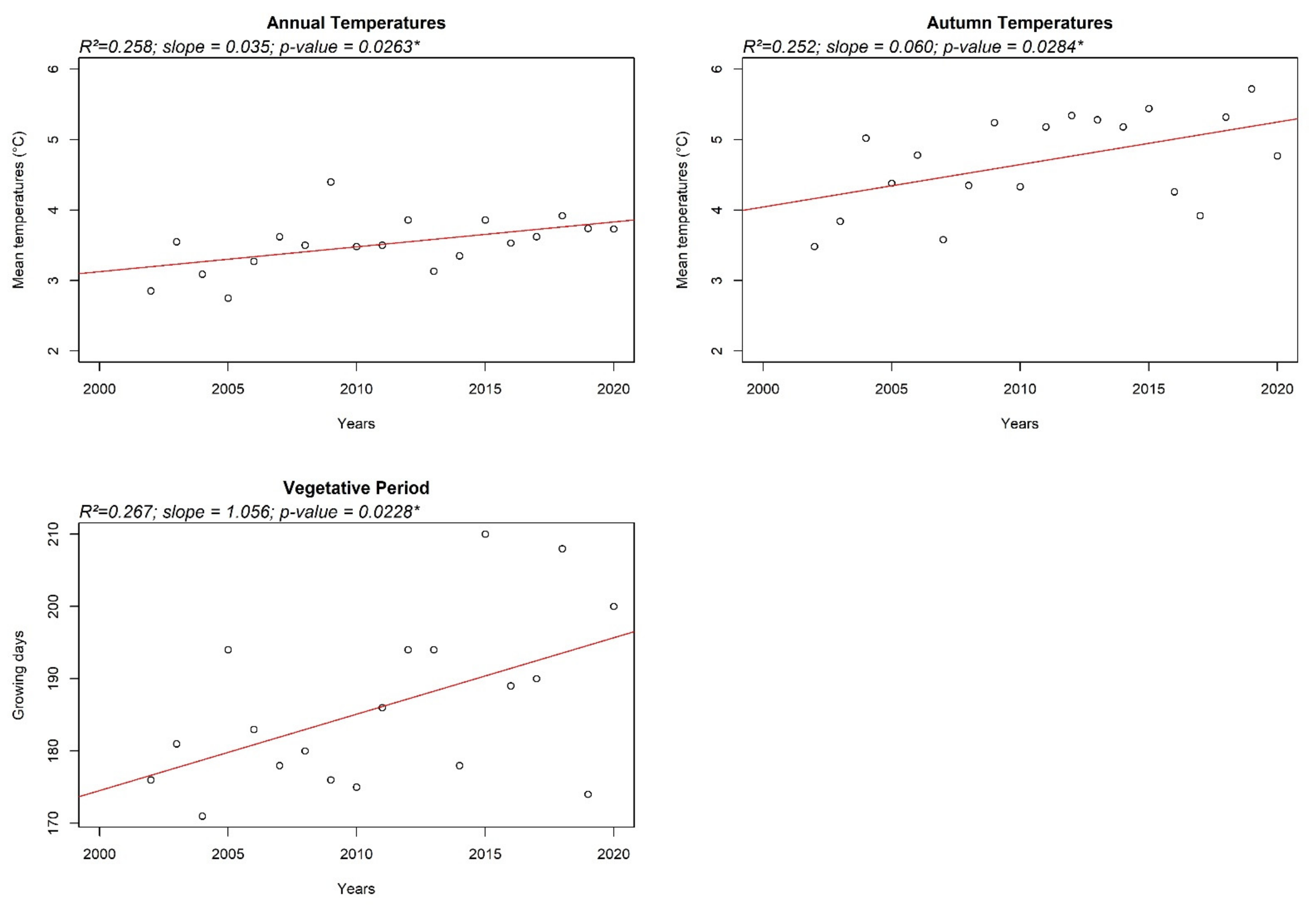
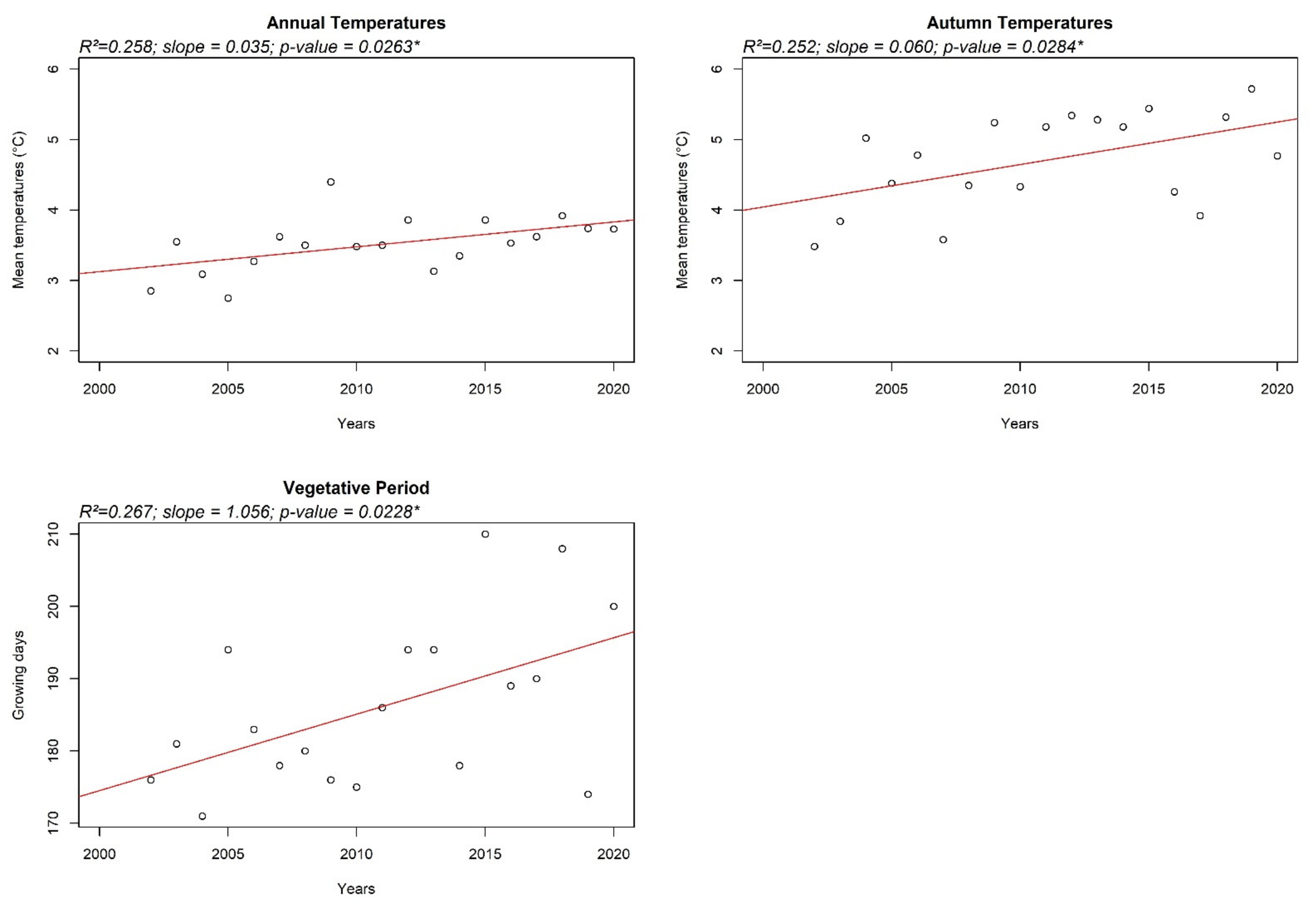
2.2. Climate Features
2.3. Plant Communities
2.4. Vegetation Data
2.5. Statistical Analysis
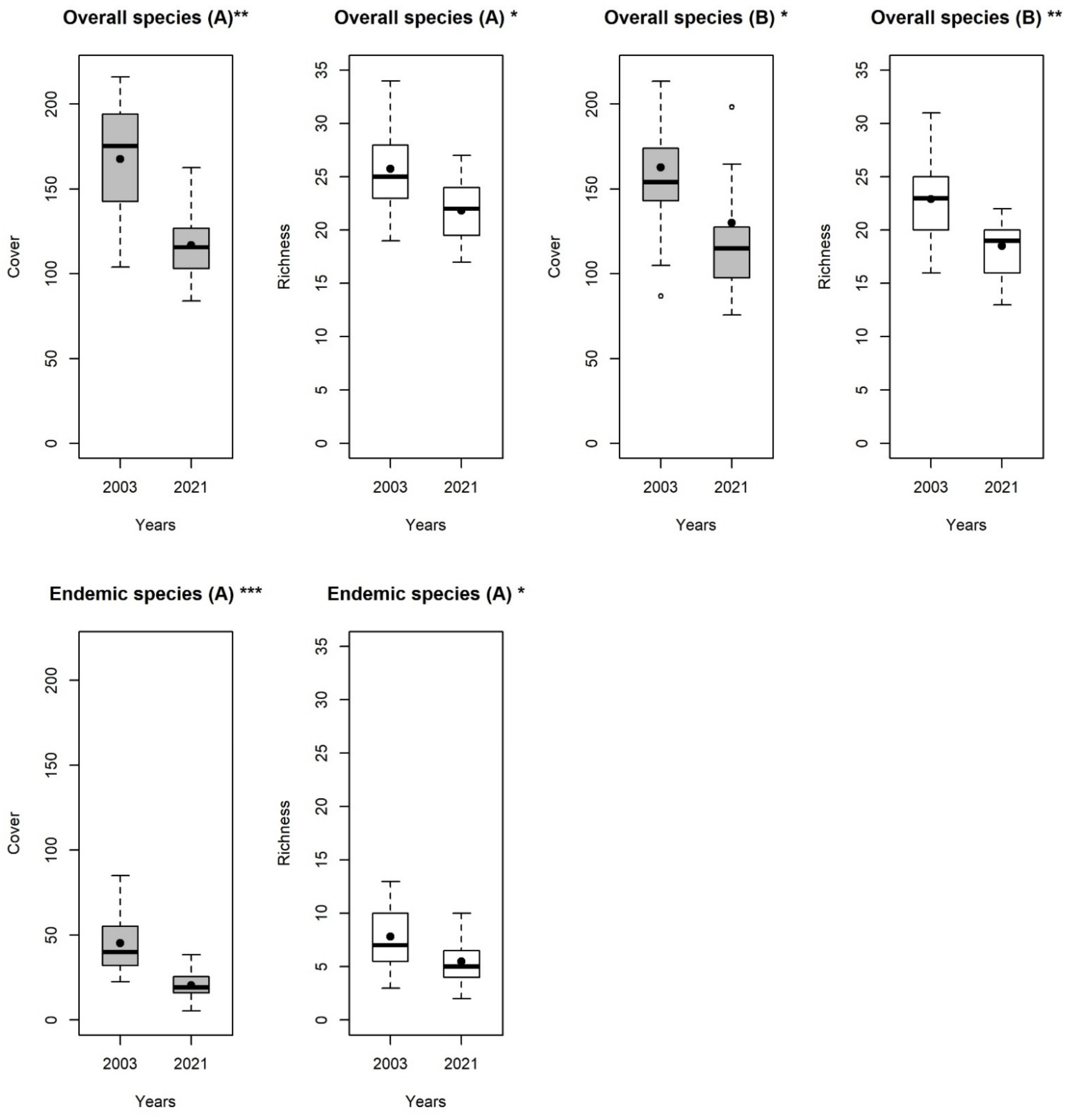
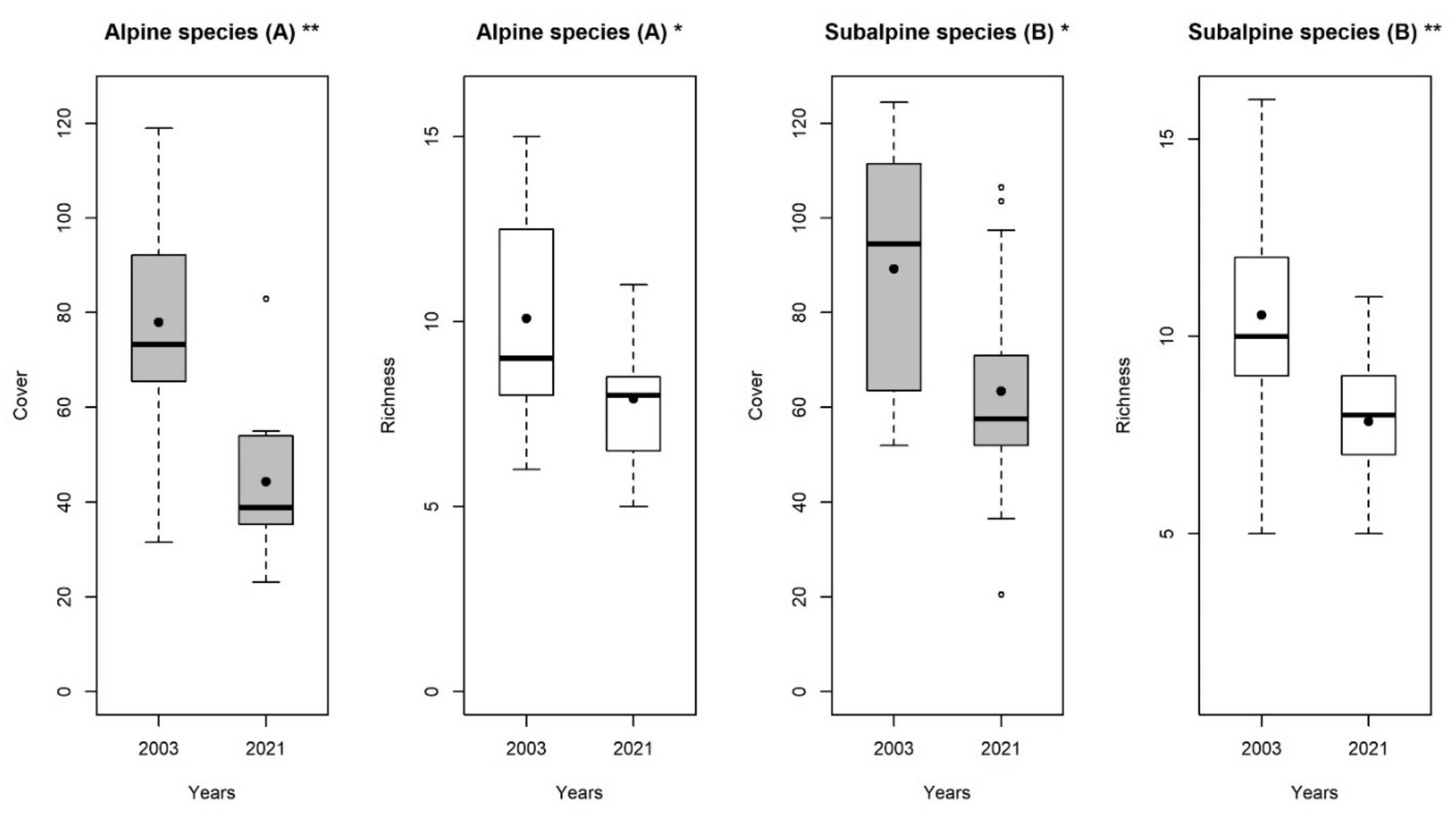
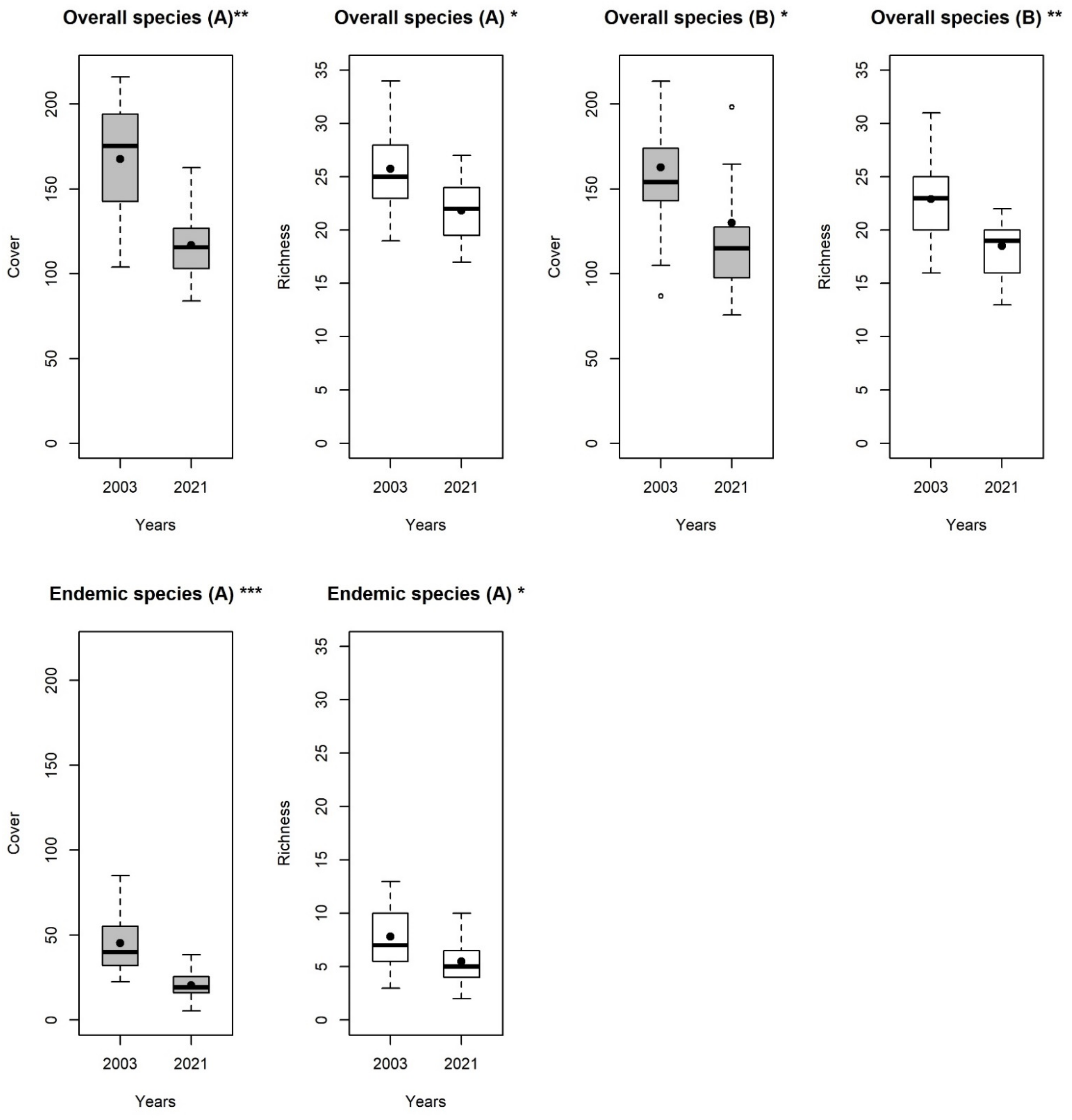
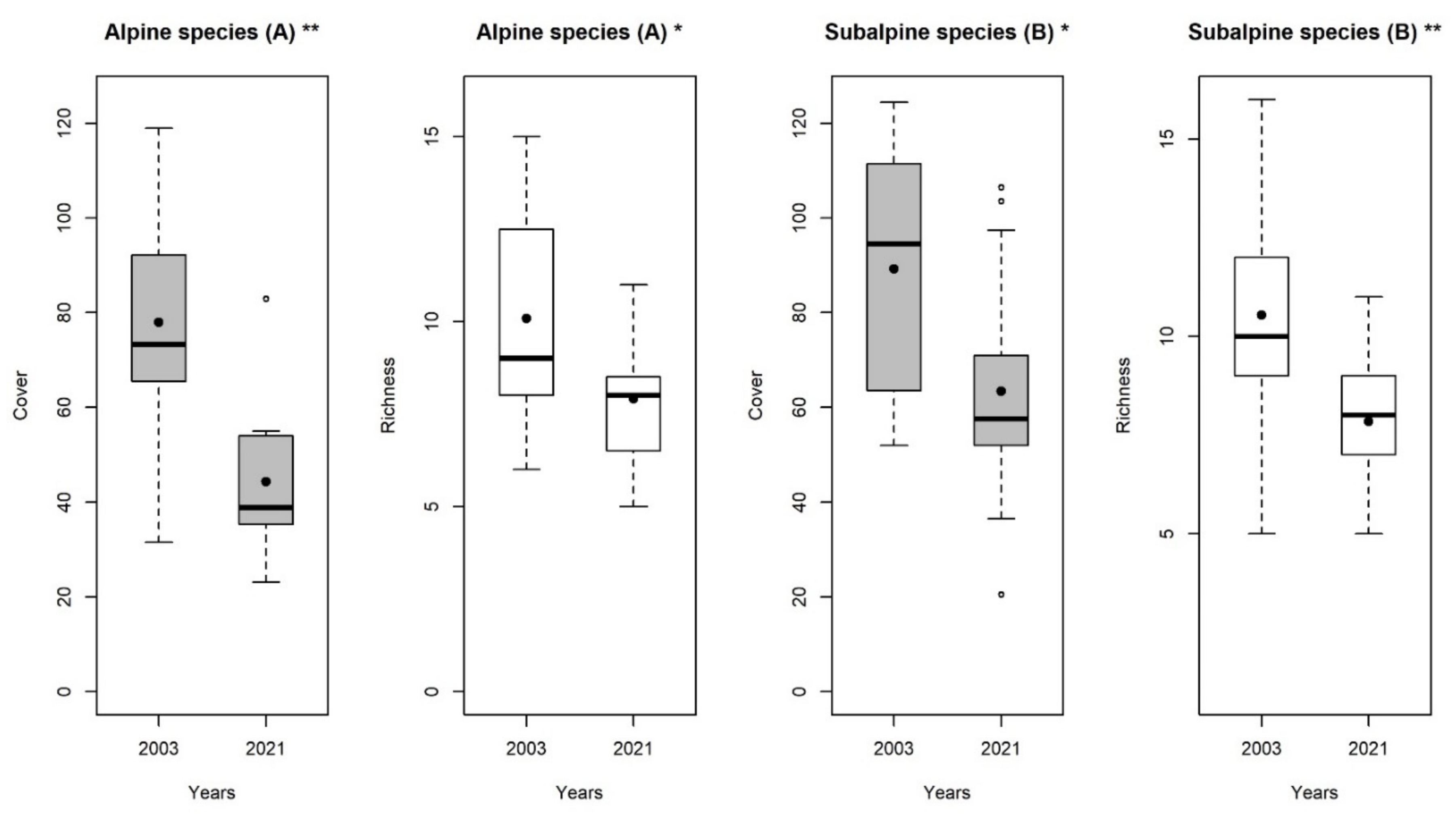
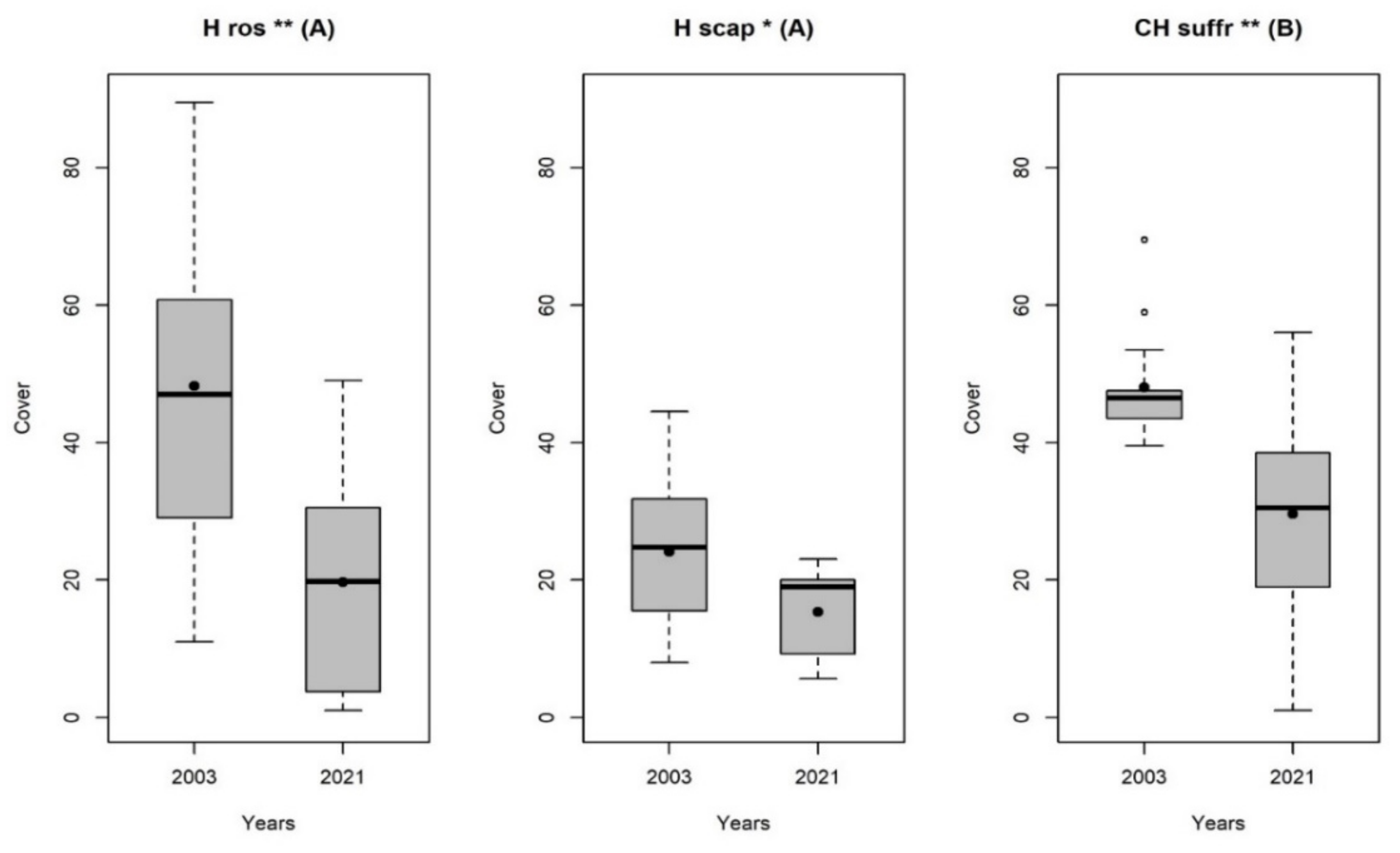
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Community | Relevé Number | Relevé Area (m2) | Altitude (m a.s.l.) | Aspect | Slope (Degree) |
|---|---|---|---|---|---|
| Apennine stripped grasslands | 1 | 16 | 2555 | E | 15 |
| 2 | 16 | 2563 | E | 18 | |
| 3 | 16 | 2601 | NW | 15 | |
| 4 | 16 | 2483 | S | 13 | |
| 5 | 16 | 2558 | SW | 5 | |
| 6 | 16 | 2614 | S | 20 | |
| 7 | 16 | 2539 | S | 15 | |
| 8 | 16 | 2572 | S | 22 | |
| 9 | 16 | 2501 | S | 34 | |
| 10 | 16 | 2400 | N | 6 | |
| 11 | 16 | 2554 | NE | 7 | |
| 12 | 16 | 2567 | E | 7 | |
| Wind edge swards | 1 | 16 | 2658 | 7 | |
| 2 | 16 | 2524 | NW | 10 | |
| 3 | 16 | 2529 | SW | 10 | |
| 4 | 16 | 2525 | NW | 7 | |
| 5 | 16 | 2479 | S | 5 | |
| 6 | 16 | 2481 | S | 7 | |
| 7 | 16 | 2481 | E | 4 | |
| 8 | 16 | 2610 | SW | 22 | |
| 9 | 16 | 2617 | E | 10 | |
| 10 | 16 | 2665 | SE | 16 | |
| 11 | 16 | 2597 | NE | 18 | |
| 12 | 16 | 2598 | E | 12 | |
| 13 | 16 | 2536 | SE | 20 |
References
- Chelli, S.; Wellstein, C.; Campetella, G.; Canullo, R.; Tonin, R.; Zerbe, S.; Gerdol, R. Climate change response of vegetation across climatic zones in Italy. Clim. Res. 2017, 71, 249–262. [Google Scholar] [CrossRef]
- Rogora, M.; Buzzi, F.; Dresti, C.; Leoni, B.; Lepori, F.; Mosello, R.; Patelli, M.; Salmaso, N. Climatic effects on vertical mixing and deep-water oxygen content in the subalpine lakes in Italy. Hydrobiologia 2018, 824, 33–50. [Google Scholar] [CrossRef]
- Verrall, B.; Pickering, C.M. Alpine vegetation in the context of climate change: A global review of past research and future directions. Sci. Total Environ. 2020, 748, 141344. [Google Scholar] [CrossRef]
- Körner, C. Alpine Plant Life, 2nd ed.; Springer: Berlin, Germany, 2003. [Google Scholar]
- Carlson, B.Z.; Choler, P.; Renaud, J.; Dedieu, J.P.; Thuiller, W. Modelling snow cover duration improves predictions of functional and taxonomic diversity for alpine plant communities. Ann. Bot. 2015, 116, 1023–1034. [Google Scholar] [CrossRef] [PubMed]
- Green, K.; Pickering, C. Vegetation, microclimate and soils associated with the latestlying snowpatches in Australia. Plant Ecol. Divers. 2009, 2, 289–300. [Google Scholar] [CrossRef]
- Fisk, M.C.; Schmidt, S.K.; Seastedt, T.R. Topographic patterns of above-and belowground production and nitrogen cycling in alpine tundra. Ecology 1998, 79, 2253–2266. [Google Scholar] [CrossRef]
- Hua, X.; Ohlemüller, R.; Sirguey, P. Differential effects of topography on the timing of the growing season in mountainous grassland ecosystems. Environ. Adv. 2022, 8, 100234. [Google Scholar] [CrossRef]
- Heywood, V.H. Global Biodiversity Assessment; Cambridge Press: Cambridge, UK, 1995. [Google Scholar]
- Körner, C. The grand challenges in functional plant ecology. Front. Plant Sci. 2011, 2, 1–3. [Google Scholar] [CrossRef]
- Médail, F.; Quézel, P. Biodiversity hotspots in the Mediterranean Basin: Setting global conservation priorities. Conserv. Biol. 1999, 13, 1510–1514. [Google Scholar] [CrossRef]
- Väre, H.; Lampinen, R.; Humphries, C.; Williams, P. Taxonomic diversity of vascular plants in the European alpine areas. In Alpine Biodiversity in Europe; Nagy, L., Grabherr, G., Körner, C., Thompson, D.B.A., Eds.; Springer: Berlin, Germany, 2003; pp. 133–148. [Google Scholar]
- Aeschimann, D.; Rasolofo, N.; Theurillat, J.-P. Analyse de la flore des Alpes. 5: Milieux et phytosociologie. Candollea 2013, 68, 5–27. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Körner, C. Impact of atmospheric changes on alpine vegetation: The ecophysiological perspective. In Potential Ecological Impacts of Climate Change in the Alps and Fennoscandian Mountains; Guisan, A., Holten, J.I., Spichiger, R., Tessier, L., Eds.; Conservatoire et Jardin Botaniques de Genève: Geneva, Switzerland, 1995; pp. 113–120. [Google Scholar]
- Testolin, R.; Attorre, F.; Borchardt, P.; Brand, R.F.; Bruelheide, H.; Chytrý, M.; De Sanctis, M.; Dolezal, J.; Finckh, M.; Haider, S.; et al. Global patterns and drivers of alpine plant species richness. Global Ecol. Biogeogr. 2021, 30, 1218–1231. [Google Scholar] [CrossRef]
- Favarger, C. Endemism in the Montane Floras of Europe. In Taxonomy Phytogeography and Evolution; Valentine, D.H., Ed.; Academic Press: London, UK; New York, NY, USA, 1972; pp. 191–204. [Google Scholar]
- Gils, H.V.; Conti, F.; Ciaschetti, G.; Westinga, E. Fine resolution distribution modelling of endemics in Majella National Park, Central Italy. Plant Biosyst. 2012, 146, 276–287. [Google Scholar] [CrossRef]
- Hughes, P.D.; Woodward, J.C. Timing of glaciation in the Mediterranean mountains during the last cold stage. J. Quat. Sci. 2008, 23, 575–588. [Google Scholar] [CrossRef]
- Catonica, C.; Manzi, A. L’influenza della storia climatica e geologica recente sulla Flora d’alta quota dei gruppi montuosi del Gran Sasso e della Majella (Appennino Centrale). Riv. Piemont. Stor. Nat. 2002, 23, 19–29. [Google Scholar]
- Stocker, T.F.; Qin, D.; Plattner, G.-K.; Alexander, L.V.; Allen, S.K.; Bindoff, N.L.; Bréon, F.-M.; Church, J.A.; Cubasch, U.; Xie, S.-P. Technical Summary. In Climate Change 2013: The Physical Science Basis; Contribution of working group to the fifth assessment report of the intergovernmental panel on climate change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; pp. 33–109. [Google Scholar]
- Klein, G.; Rebetez, M.; Rixen, C.; Vitasse, Y. Unchanged risk of frost exposure for subalpine and alpine plants after snowmelt in Switzerland despite climate warming. Int. J. Biometeorol. 2018, 62, 1755–1762. [Google Scholar] [CrossRef]
- Pauli, H.; Gottfried, M.; Dullinger, S.; Abdaladze, O.; Akhalkatsi, M.; Alonso, J.L.B.; Coldea, G.; Dick, J.; Erschbamer, B.; Calzado, R.F.; et al. Recent plant diversity changes on Europe’s mountain summits. Science 2012, 336, 353–355. [Google Scholar] [CrossRef]
- Matteodo, M.; Ammann, K.; Verrecchia, E.P.; Vittoz, P. Snowbeds are more affected than other subalpine–alpine plant communities by climate change in the Swiss Alps. Ecol. Evol. 2016, 6, 6969–6982. [Google Scholar] [CrossRef]
- Steinbauer, M.J.; Grytnes, J.-A.; Jurasinski, G.; Kulonen, A.; Lenoir, J.; Pauli, H.; Rixen, C.; Winkler, M.; Bardy-Durchhalter, M.; Barni, E.; et al. Accelerated increase in plant species richness on mountain summits is linked to warming. Nature 2018, 556, 231–234. [Google Scholar] [CrossRef]
- Cannone, N.; Sgorbati, S.; Guglielmin, M. Unexpected impacts of climate change on alpine vegetation. Front. Ecol. Environ. 2007, 5, 360–364. [Google Scholar] [CrossRef]
- Holzinger, B.; Hulber, K.; Camenisch, M.; Grabherr, G. Changes in plant species richness over the last century in the eastern Swiss Alps: Elevational gradient, bedrock effects and migration rates. Plant Ecol. 2008, 195, 179–196. [Google Scholar] [CrossRef]
- Parolo, G.; Rossi, G. Upward migration of vascular plants following a climate warming trend in the Alps. Basic Appl. Ecol. 2008, 9, 100–107. [Google Scholar] [CrossRef]
- Britton, A.J.; Beale, C.M.; Towers, W.; Hewison, R.L. Biodiversity gains and losses: Evidence for homogenisation of Scottish alpine vegetation. Biol. Conserv. 2009, 142, 1728–1739. [Google Scholar] [CrossRef]
- Engler, R.; Randin, C.F.; Thuiller, W.; Dullinger, S.; Zimmermann, N.E.; Araújo, M.B.; Pearman, P.B.; Le Lay, G.; Piedallu, C.; Albert, C.H.; et al. 21st Century Climate Change Threatens Mountain Flora Unequally Across Europe. Glob. Change Biol. 2011, 17, 2330–2341. [Google Scholar] [CrossRef]
- Palombo, C.; Chirici, G.; Marchetti, M.; Tognetti, R. Is land abandonment affecting forest dynamics at high elevation in Mediterranean mountains more than climate change? Plant Biosyst. 2013, 147, 1–11. [Google Scholar] [CrossRef]
- Erschbamer, B.; Unterluggauer, P.; Winkler, E.; Mallaun, M. Changes in plant species diversity revealed by long-term monitoring on mountain summits in the Dolomites (northern Italy). Preslia 2011, 83, 387–401. [Google Scholar]
- Gottfried, M.; Pauli, H.; Futschik, A.; Akhalkatsi, M.; Barančok, P.; Benito Alonso, J.L.; Coldea, G.; Dick, J.; Erschbamer, B.; Fernández Calzado, M.R.; et al. Continent-wide response of mountain vegetation to climate change. Nat. Clim. Chang. 2012, 2, 111–115. [Google Scholar] [CrossRef]
- Stanisci, A.; Frate, L.; Di Cella, M.U.; Pelino, G.; Petey, M.; Siniscalco, C.; Carranza, M.L. Short-Term Signals of Climate Change in Italian Summit Vegetation: Observations at Two GLORIA Sites. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2014, 150, 227–235. [Google Scholar] [CrossRef]
- Evangelista, A.; Frate, L.; Carranza, M.L.; Attorre, F.; Pelino, G.; Stanisci, A. Changes in composition, ecology and structure of high-mountain vegetation: A re-visitation study over 42 years. AoB Plants 2016, 8, 1–11. [Google Scholar] [CrossRef]
- Cannone, N.; Pignatti, S. Ecological responses of plant species and communities to climate warming: Upward shift or range filling processes? Clim. Chang. 2014, 123, 201–214. [Google Scholar] [CrossRef]
- Calabrese, V.; Carranza, M.L.; Evangelista, A.; Marchetti, M.; Stinca, A.; Stanisci, A. Long-term changes in the composition, ecology, and structure of Pinus mugo scrubs in the Apennines (Italy). Diversity 2018, 10, 70. [Google Scholar] [CrossRef]
- Erschbamer, B.; Kiebache, T.; Mallaun, M.; Unterluggauer, P. Short-term signals of climate change along an altitudinal gradient in the South Alps. Plant Ecol. 2009, 202, 79–89. [Google Scholar] [CrossRef]
- Schwartz, M.W.; Iverson, L.R.; Prasad, A.M.; Matthews, S.N.; O’Connor, R.J. Predicting extinctions as a result of climate change. Ecology 2006, 87, 1611–1615. [Google Scholar] [CrossRef]
- Dirnbock, T.; Essl, F.; Rabitsch, W. Disproportional risk for habitat loss of high-altitude endemic species under climate change. Glob. Chang. Biol. 2011, 17, 990–996. [Google Scholar] [CrossRef]
- Fernández-Calzado, R.; Molero, M.J.; Merzouki, A.; Casares, P. Vascular plant diversity and climate change in the upper zone of Sierra Nevada, Spain. Plant Biosyst. 2012, 146, 1044–1053. [Google Scholar] [CrossRef]
- Nogués-Bravo, D.; Araújo, M.B.; Lasanta, T.; Moreno, J.I.L. Climate Change in Mediterranean Mountains during the 21st Century. Ambio 2008, 37, 280–285. [Google Scholar] [CrossRef]
- Cherif, S.; Doblas-Miranda, E.; Lionello, P.; Borrego, C.; Giorgi, F.; Iglesias, A.; Jebari, S.; Mahmoudi, E.; Moriondo, M.; Pringault, O.; et al. Climate and Environmental Change in the Mediterranean Basin–Current Situation and Risks for the Future. In Drivers of Change; Cramer, W., Guiot, J., Marini, K., Eds.; Union for the Mediterranean, Plan Bleu, UNEP/MAP: Marseille, France, 2021; pp. 59–180. [Google Scholar]
- Lamprecht, A.; Pauli, H.; Fernández Calzado, M.R.; Lorite, J.; Molero, M.J.; Steinbauer, K.; Winkler, M. Changes in plant diversity in a water-limited and isolated high-mountain range (Sierra Nevada, Spain). Alp. Bot. 2021, 131, 17–39. [Google Scholar] [CrossRef]
- Kazakis, G.; Ghosn, D.; Remoundou, I.; Nyktas, P.; Talias, M.A.; Vogiatzakis, I.N. Altitudinal Vascular Plant Richness and Climate Change in the Alpine Zone of the Lefka Ori, Crete. Diversity 2021, 13, 22. [Google Scholar] [CrossRef]
- Porro, F.; Tomaselli, M.; Abeli, T.; Gandini, M.; Gualmini, M.; Orsenigo, S.; Petraglia, A.; Rossi, G.; Carbognani, M. Could plant diversity metrics explain climate-driven vegetation changes on mountain summits of the GLORIA network? Biodivers. Conserv. 2019, 28, 3575–3596. [Google Scholar] [CrossRef]
- Nicklas, L.; Walde, J.; Wipf, S.; Lamprecht, A.; Mallaun, M.; Rixen, C.; Steinbauer, K.; Theurillat, J.-P.; Unterluggauer, P.; Vittoz, P.; et al. Climate Change Affects Vegetation Differently on Siliceous and Calcareous Summits of the European Alps. Front. Ecol. Evol. 2021, 9, 642309. [Google Scholar] [CrossRef]
- Turco, M.; Vezzoli, R.; De Ronco, P.; Mercogliano, P. Variation in Discharge, Precipitation and Temperature in Po River and Tributaries Basins; no. 185; Centro Euro-Mediterraneo sui Cambiamenti Climatici (CMCC): Lecce, Italy, 2013. [Google Scholar]
- Sillmann, J.; Kharin, V.V.; Zwiers, F.W.; Zhang, X.; Bronaugh, D. Climate extremes indices in the CMIP5 multimodel ensemble: Part 2. Future climate projections. J. Geophys. Res. Atmos. 2013, 118, 2473–2493. [Google Scholar] [CrossRef]
- Barredo, J.I.; Mauri, A.; Caudullo, G.; Dosio, A. Assessing Shifts of Mediterranean and arid climates under RCP4.5 and RCP8.5 climate projections in Europe. Pure Appl. Geophys. 2018, 175, 3955–3971. [Google Scholar] [CrossRef]
- Di Nuzzo, L.; Vallese, C.; Benesperi, R.; Giordani, P.; Chiarucci, A.; Di Cecco, V.; Di Martino, L.; Di Musciano, M.; Gheza, G.; Lelli, C.; et al. Contrasting multitaxon responses to climate change in Mediterranean mountains. Sci. Rep. 2021, 11, 4438. [Google Scholar] [CrossRef]
- Biondi, E.; Blasi, C.; Burrascano, S.; Casavecchia, S.; Copiz, R.; Del Vico, E.; Galdenzi, D.; Gigante, D.; Lasen, C.; Spampinato, G.; et al. Manuale Italiano di Interpretazione Degli Habitat Della Direttiva 92/43/CEE.; Ministero dell’Ambiente e della Tutela del Territorio e del Mare, Direzione per la Protezione della Natura: Rome, Italy, 2009; pp. 1–16.
- European Commission DG Environment. Interpretation Manual of European Union Habitats. [Eur 28. Nature ENV B.3]. 2013. Available online: http://ec.europa.eu/environment/nature/legislation/habitatsdirective/docs/Int_Manual_EU28.pdf (accessed on 26 June 2019).
- Jaurand, E. Les Heritages Glaciaire de l’Apennin. Ph.D. Thesis, Éditions de la Sorbonne Paris, Paris, France, 1994. [Google Scholar]
- Giraudi, C. Nuovi dati sul glacialismo della montagna della Majella (Abruzzo, Italia centrale). Ital. J. Quat. Sci. 1998, 11, 265–271. [Google Scholar]
- Romeo, R.; Vita, A.; Manuelli, S.; Zanini, E.; Freppaz, M.; Stanchi, S. Understanding Mountain Soils: A contribution from Mountain Areas to the International Year of Soils 2015; FAO: Rome, Italy, 2015. [Google Scholar]
- Blasi, C.; Di Pietro, R.; Fortini, P.; Catonica, C. The main Plant community types of the alpine belt of the Apennine chain. Plant Biosyst. 2003, 137, 83–110. [Google Scholar] [CrossRef]
- Stanisci, A.; Carranza, M.L.; Pelino, G.; Chiarucci, A. Assessing the diversity pattern of cryophilous plant species in high elevation habitats. Plant Ecol. 2010, 212, 595–600. [Google Scholar] [CrossRef]
- Stanisci, A.; Pelino, G.; Blasi, C. Vascular plant diversity and climate change in the alpine belt of the central Apennines (Italy). Biodivers. Conserv. 2005, 14, 1301–1318. [Google Scholar] [CrossRef]
- Stanisci, A.; Evangelista, A.; Frate, L.; Stinca, A.; Carranza, M.L. VIOLA—Database of high mountain vegetation of central Apennines. Phytocoenologia 2016, 46, 231–232. [Google Scholar] [CrossRef]
- Cervellini, M.; Zannini, P.; Di Musciano, M.; Fattorini, S.; Jiménez-Alfaro, B.; Rocchini, D.; Field, R.; Vetaas, O.R.; Severin, D.H.I.; Beierkuhnlein, C.; et al. A grid-based map for the Biogeographical Regions of Europe. Biodivers. Data J. 2020, 8, e53720. [Google Scholar] [CrossRef] [PubMed]
- Petriccione, B.; Bricca, A. Thirty years of ecological research at the Gran Sasso d’Italia LTER site: Climate change in action. Nat. Cons. 2019, 34, 9–39. [Google Scholar] [CrossRef]
- Steinbauer, K.; Lamprecht, A.; Winkler, M.; Di Cecco, V.; Fasching, V.; Ghosn, D.; Maringer, A.; Remoundou, I.; Suen, M.; Stanisci, A.; et al. Recent changes in high-mountain plant community functional composition in contrasting climate regimes. Sci. Total Environ. 2022, 829, 154541. [Google Scholar] [CrossRef] [PubMed]
- Intergovernmental Panel on Climate Change. Sixth Assessment Report. Climate Change 2022: Impacts, Adaptation and Vulnerability; Cambridge University Press: Cambridge, UK, 2022. [Google Scholar]
- Pauli, H.; Gottfried, M.; Lamprecht, A.; Niessner, S.; Rumpf, S.; Winkler, M.; Steinbauer, K.; Grabherr, G. The GLORIA Field Manual; Global Observation Research Initiative in Alpine Environments: Vienna, Austria, 2015. [Google Scholar]
- Chytrý, M.; Tichý, L.; Hennekens, S.M.; Knollová, I.; Janssen, J.A.M.; Rodwell, J.S.; Peterka, T.; Marcenò, C.; Landucci, F.; Danihelka, J.; et al. EUNIS Habitat Classification: Expert system, characteristic species combinations and distribution maps of European habitats. Appl. Veg. Sci. 2020, 23, 648–675. [Google Scholar] [CrossRef]
- Biondi, E.; Blasi, C.; Allegrezza, M.; Anzellotti, I.; Azzella, M.M.; Carli, E.; Casavecchia, S.; Copiz, R.; Del Vico, E.; Facioni, L.; et al. Plant communities of Italy: The Vegetation Prodrome. Plant Biosyst. 2014, 148, 728–814. [Google Scholar] [CrossRef]
- Evangelista, A.; Frate, L.; Stinca, A.; Carranza, M.L.; Stanisci, A. VIOLA-The vegetation database of the central Apennines: Structure, current status and usefulness for monitoring Annex i EU habitats (92/43/EEC). Plant Sociol. 2016, 53, 47–58. [Google Scholar] [CrossRef]
- Vymazalová, M.; Axmanová, I.; Tichý, L. Effect of intra-seasonal variability on vegetation data. J. Veg. Sci. 2012, 23, 978–984. [Google Scholar] [CrossRef]
- Chytrý, M.; Tichý, L.; Hennekens, S.M.; Schaminée, J.H.J. Assessing vegetation change using vegetation-plot databases: A risky business. Appl. Veg. Sci. 2014, 17, 32–41. [Google Scholar] [CrossRef]
- Braun-Blanquet, J. Pflanzensoziologie: Grundzüge der Vegetationskunde; Springer: Berlin, Germany, 1964; ISBN 3709140781. [Google Scholar]
- Westhoff, V.; Van Der Maarel, E. The braun-blanquet approach. In Classification of Plant Communities; Springer: Berlin/Heidelberg, Germany, 1978; pp. 287–399. [Google Scholar]
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Albano, A.; Alessandrini, A.; Ardenghi, N.M.G.; Astuti, G.; Bacchetta, G.; Ballelli, S.; Banfi, E.; et al. An updated checklist of the vascular flora native to Italy. Plant Biosyst. 2018, 152, 179–303. [Google Scholar] [CrossRef]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora d’Italia; Edagricole: Rome, Italy, 2017; Volume 1, ISBN 8850652429. [Google Scholar]
- Landolt, E.; Bäumler, B.; Erhardt, A.H.O.; Klötzli, F.; Lämmler, W.; Nobis, M.; Rudmann-Maurer, K.; Schweingruber, F.H.; Theurillat, J.-P.; Urmi, E.; et al. Flora Indicativa. Ecological Indicator Values and Biological Attributes of the Flora of Switzerland and the Alps, 2nd ed.; Haupt Verlag: Berna, Switzerland, 2010. [Google Scholar]
- Frate, L.; Carranza, M.L.; Evangelista, A.; Stinca, A.; Schaminée, J.H.J.; Stanisci, A. Climate and land use change impacts on Mediterranean high-mountain vegetation in the Apennines since the 1950s. Plant Ecol. Divers. 2018, 11, 85–96. [Google Scholar] [CrossRef]
- Tichý, L.; Hennekens, S.M.; Novák, P.; Rodwell, J.S.; Schaminée, J.H.J.; Chytrý, M. Optimal transformation of species cover for vegetation classification. Appl. Veg. Sci. 2020, 23, 710–717. [Google Scholar] [CrossRef]
- Raunkiær, C.C. The Life Forms of Plants and Statistical Plant Geography; Oxford University Press: Oxford, UK, 1934. [Google Scholar]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: http://www.r-project.org/index.html (accessed on 7 February 2021).
- Oksanen, A.J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; Hara, R.B.O.; Simpson, G.L.; Solymos, P.; et al. Vegan Package 2020. Available online: https://github.com/vegandevs/vegan (accessed on 7 February 2021).
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past Palaeontological Statistics, Ver. 1.79. 2001, pp. 1–88. Available online: https://palaeo-electronica.org/2001_1/past/issue1_01.htm#:~{}:text=The%20program%2C%20called%20PAST%20(PAleontological,plotting%2C%20and%20simple%20phylogenetic%20analysis (accessed on 10 June 2021).
- Mariotti, A.; Zeng, N.; Yoon, J.-H.; Artale, V.; Navarra, A.; Alpert, P.; Li, L.Z.X. Mediterranean water cycle changes: Transition to drier 21st century conditions in observations and CMIP3 simulations. Environ. Res. Lett. 2008, 3, 044001. [Google Scholar] [CrossRef]
- Toreti, A.; Fioravanti, G.; Percontia, W.; Desiatoa, F. Annual and seasonal precipitation over Italy from 1961 to 2006. Int. J. Climatol. 2009, 29, 1976–1987. [Google Scholar] [CrossRef]
- Fernández Calzado, M.R.; Molero, J. Changes in the summit flora of a Mediterranean mountain (Sierra Nevada, Spain) as a possible effect of climate change. Lazaroa 2013, 34, 65–75. [Google Scholar] [CrossRef]
- Brunetti, M.; Buffoni, L.; Mangianti, F.; Maugeri, M.; Nanni, T. Temperature, precipitation and extreme events during the last century in Italy. Glob. Planet Chang. 2004, 40, 141–149. [Google Scholar] [CrossRef]
- Elguindi, N.; Rauscher, S.A.; Giorgi, F. Historical and future changes in maximum and minimum temperature records over Europe. Clim. Chang. 2013, 117, 415–431. [Google Scholar] [CrossRef]
- Simolo, C.; Brunetti, M.; Maugeri, M.; Nanni, T. Increasingly warm summers in the Euro-Mediterranean zone: Mean temperatures and extremes. Reg. Environ. Chang. 2014, 14, 1825–1832. [Google Scholar] [CrossRef]
- Christensen, J.H.; Hewiston, B.; Busuioc, A.; Chen, A.; Gao, X.; Held, I.; Jones, R.; Kolli, R.K.; Kwon, W.T.; Laprise, R.; et al. AR4 Climate Change 2007: The Physical Science Basis; Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge Univ. Press: Cambridge, UK, 2007; pp. 847–940. [Google Scholar]
- Benito, B.; Lorite, J.; Penas, J. Simulating potential effects of climatic warming on altitudinal patterns of key species in Mediterranean-alpine ecosystems. Clim. Chang. 2011, 108, 471–483. [Google Scholar] [CrossRef]
- Mondoni, A.; Rossi, G.; Orsenigo, S.; Probert, R.J. Climate warming could shift the timing of seed germination in alpine plants. Ann. Bot. 2012, 110, 155–164. [Google Scholar] [CrossRef]
- Giménez-Benavides, L.; Escudero, A.; Pérez-García, F. Seed germination of high mountain Mediterranean species: Altitudinal, interpopulation and interannual variability. Ecol. Res. 2005, 20, 433–444. [Google Scholar] [CrossRef]
- Orsenigo, S.; Abeli, T.; Rossi, G.; Bonasoni, P.; Pasquaretta, C.; Gandini, M.; Mondoni, A. Effects of autumn and spring heat waves on seed germination of high mountain plants. PLoS ONE 2015, 10, e0133626. [Google Scholar] [CrossRef]
- Varricchione, M.; Di Cecco, V.; Santoianni, L.A.; Stanisci, A.; Di Febbraro, M.; Di Martino, L.; Carranza, M.L. Diagnostic Species Diversity Pattern Can Provide Key Information on Vegetation Change: An Insight into High Mountain Habitats in Central Apennines. J. Zool. Bot. Gard. 2021, 2, 453–472. [Google Scholar] [CrossRef]
- Edwards, A.C.; Scalenghe, R.; Freppaz, M. Changes in the seasonal snow cover of alpine regions and its effect on soil processes: A review. Quat. Int. 2007, 162–163, 172–181. [Google Scholar] [CrossRef]
- Stanisci, A.; Bricca, A.; Calabrese, V.; Cutini, M.; Pauli, H.; Steinbauer, K.; Carranza, M.L. Functional composition and diversity of leaf traits in subalpine versus alpine vegetation in the Apennines. AoB Plants 2020, 12, 1–11. [Google Scholar] [CrossRef]
- Haslinger, K.; Schöner, W.; Anders, I. Future drought probabilities in the Greater Alpine Region based on COSMO-CLM experiments–Spatial patterns and driving forces. Meteorol. Z. 2016, 25, 137–148. [Google Scholar] [CrossRef]
- Cutini, M.; Marzialetti, F.; Giuliana, B.; Rianna, G.; Theurillat, J.-P. Bioclimatic pattern in a Mediterranean mountain area: Assessment from a classification approach on a regional scale. Int. J. Biometeorol. 2021, 65, 1085–1097. [Google Scholar] [CrossRef]
- Chen, I.-C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid Range Shifts of Species Associated with High Levels of Climate Warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef]
- Rehm, E.M.; Olivas, P.; Stroud, J.; Feeley, K.J. Losing your edge: Climate change and the conservation value of range-edge populations. Ecol. Evol. 2015, 5, 4315–4326. [Google Scholar] [CrossRef]
- Jump, A.S.; Peñuelas, J. Running to stand still: Adaptation and the response of plants to rapid climate change. Ecol. Lett. 2005, 8, 1010–1020. [Google Scholar] [CrossRef]
- Hoffmann, S.; Irl, S.D.H.; Beierkuhnlein, C. Predicted climate shifts within terrestrial protected areas worldwide. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef]
- Manes, S.; Costello, M.J.; Beckett, H.; Debnath, A.; Devenish-Nelson, E.; Grey, K.-A.; Jenkins, R.; Ming Khan, T.; Kiessling, W.; Krause, C.; et al. Endemism increases species’ climate change risk in areas of global biodiversity importance. Biol. Conserv. 2021, 25, 109270. [Google Scholar] [CrossRef]
- Watts, S.H.; Mardon, D.K.; Mercer, C.; Watson, D.; Cole, H.; Shaw, R.F.; Jump, A.S. Riding the elevator to extinction: Disjunct arctic-alpine plants of open habitats decline as their more competitive neighbors expand. Biol. Conserv. 2022, 272, 109620. [Google Scholar] [CrossRef]
- Nagy, L.; Grabherr, G.; Körner, C.; Thompson, D.B.A. Alpine Biodiversity in Europe; Springer: Berlin, Germany, 2003. [Google Scholar]
- Kougioumoutzis, K.; Kokkoris, I.P.; Panitsa, M.; Trigas, P.; Strid, A.; Dimopoulos, P. Plant Diversity Patterns and Conservation Implications under Climate-Change Scenarios in the Mediterranean: The Case of Crete (Aegean, Greece). Diversity 2020, 12, 270. [Google Scholar] [CrossRef]
- Di Biase, L.; Pace, L.; Mantoni, C.; Fattorini, S. Variations in Plant Richness, Biogeographical Composition, and Life Forms along an Elevational Gradient in a Mediterranean Mountain. Plants 2021, 10, 2090. [Google Scholar] [CrossRef] [PubMed]
- Gentili, R.; Baroni, C.; Caccianiga, M.; Armiraglio, S.; Ghiani, A.; Citterio, S. Potential warm-stage microrefugia for alpine plants: Feedback between geomorphological and biological processes. Ecol. Complex. 2015, 21, 87–99. [Google Scholar] [CrossRef]
- Veron, S.; Faith, D.P.; Pellens, R.; Pavoine, S. Priority areas for phylogenetic diversity: Maximising gains in the mediterranean basin. In Phylogenetic Diversity: Applications and Challenges in Biodiversity Science; Springer International Publishing: Cham, Switzerland, 2018; pp. 145–166. [Google Scholar]
- Haight, J.; Hammill, E. Protected areas as potential refugia for biodiversity under climatic change. Biol. Conserv. 2020, 241, 108258. [Google Scholar] [CrossRef]
- Di Cecco, V.; Di Santo, M.; Di Musciano, M.; Manzi, A.; Di Cecco, M.; Ciaschetti, G.; Marcantonio, G.; Di Martino, L. The Majella National Park: A case study for the conservation of plant biodiversity in the Italian Apennines. Ital. Bot. 2020, 10, 1–24. [Google Scholar] [CrossRef]
- Hawkes, J.G.; Maxted, N.; Ford-Lloyd, B.V. The Ex Situ Conservation of Plant Genetic Resources; Springer Science and Business Media: Berlin, Germany, 2012. [Google Scholar]

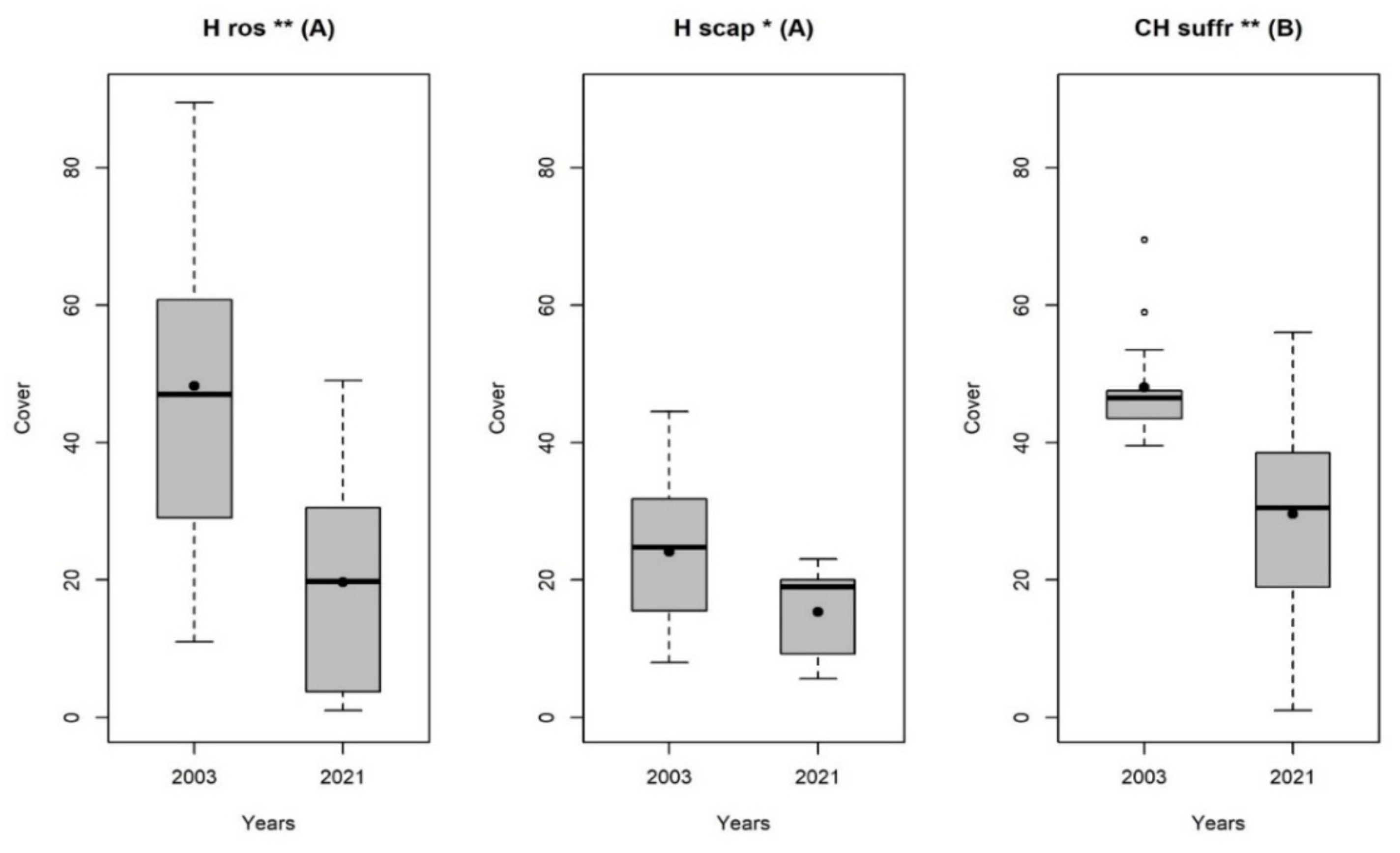
| Species | Growth Form | Average Altitudinal Distribution | EIVs T |
|---|---|---|---|
| Achillea barrelieri subsp. barrelieri | H scap | Alpine | 1.5 |
| Alyssum diffusum subsp. diffusum | CH suffr | Montane | 3 |
| Androsace vitaliana subsp. praetutiana | CH suffr | Alpine | 1.5 |
| Armeria gracilis subsp. majellensis | H ros | Subalpine | 2 |
| Carduus chrysacanthus | H scap | Subalpine | 2.5 |
| Cerastium thomasii | CH suffr | Alpine | 1.5 |
| Erysimum majellense | H scap | Subalpine | 2.5 |
| Festuca violacea subsp. italica | H caesp | Alpine | 1.5 |
| Galium magellense | H scap | Alpine | 1.5 |
| Helictochloa praetutiana subsp. praetutiana | H caesp | Subalpine | 2.5 |
| Leontopodium nivale | H scap | Alpine | 1.5 |
| Myosotis graui | H scap | Subalpine | 2.5 |
| Noccaea stylosa | CH suffr | Subalpine | 2 |
| Pedicularis elegans | H ros | Subalpine | 2.5 |
| Phyllolepidum rupestre | CH suffr | Alpine | 1.5 |
| Ranunculus pollinensis | H scap | Subalpine | 2.5 |
| Rhinanthus wettsteinii | T scap | Subalpine | 2 |
| Saxifraga italica | CH pulv | Alpine | 1 |
| Scorzoneroides montana subsp. breviscapa | H ros | Alpine | 1.5 |
| Trifolium pratense subsp. semipurpureum | H scap | Montane | 3.5 |
| Valeriana saliunca | H scap | Alpine | 1 |
| Viola eugeniae subsp. eugeniae | H scap | Subalpine | 2.5 |
| Viola magellensis | H scap | Alpine | 1 |
| Plant Community | Species | Growth Form | Range | EIVs | Species Contribution (%) | Cumulative Contribution (%) | Mean Cover | |
|---|---|---|---|---|---|---|---|---|
| T | T1 | T2 | ||||||
| Apennine stripped grasslands | Plantago atrata subsp. atrata | H ros | Alpine | 1.5 | 10 | 10 | 20.5 | 8.8 |
| Anthyllis vulneraria subsp. pulchella | H scap | Montane | 3 | 7.9 | 17.9 | 11.7 | 15.4 | |
| Carex kitaibeliana | H caesp | Subalpine | 2 | 7 | 25 | 18.5 | 20.6 | |
| ° Armeria gracilis subsp. majellensis | H ros | Subalpine | 2 | 6.7 | 31.7 | 14.8 | 3.1 | |
| ° Festuca violacea subsp. italica | H caesp | Alpine | 1.5 | 6.4 | 38 | 13.9 | 7.3 | |
| Poa alpina subsp. alpina | H caesp | Alpine | 1.5 | 5.6 | 43.6 | 12 | 2.6 | |
| Helianthemum oelandicum subsp. alpestre | CH suffr | Subalpine | 2.5 | 5 | 48.6 | 7 | 4.5 | |
| Silene acaulis subsp. bryoides | CH pulv | Alpine | 1 | 4.1 | 52.7 | 5.9 | 5.6 | |
| Wind edge swards | Carex kitaibeliana | H caesp | Subalpine | 2 | 12.4 | 12.4 | 25.1 | 20.3 |
| Helianthemum oelandicum subsp. alpestre | CH suffr | Subalpine | 2.5 | 11.1 | 23.5 | 39.4 | 23.3 | |
| Salix retusa | CH frut | Alpine | 1.5 | 7.9 | 31.4 | 5.9 | 13.3 | |
| ° Festuca violacea subsp. italica | H caesp | Alpine | 1.5 | 5.7 | 37.1 | 10.7 | 4.9 | |
| Carex myosuroides | H caesp | Alpine | 1.5 | 5.4 | 42.5 | 4.3 | 8.5 | |
| Oxytropis campestris | H scap | Alpine | 1.5 | 4.2 | 46.7 | 5.9 | 2.6 | |
| Anthyllis vulneraria subsp. pulchella | H scap | Montane | 3 | 4.2 | 50.9 | 14 | 14 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varricchione, M.; Carranza, M.L.; Di Cecco, V.; Di Martino, L.; Stanisci, A. Warmer and Poorer: The Fate of Alpine Calcareous Grasslands in Central Apennines (Italy). Diversity 2022, 14, 695. https://doi.org/10.3390/d14090695
Varricchione M, Carranza ML, Di Cecco V, Di Martino L, Stanisci A. Warmer and Poorer: The Fate of Alpine Calcareous Grasslands in Central Apennines (Italy). Diversity. 2022; 14(9):695. https://doi.org/10.3390/d14090695
Chicago/Turabian StyleVarricchione, Marco, Maria Laura Carranza, Valter Di Cecco, Luciano Di Martino, and Angela Stanisci. 2022. "Warmer and Poorer: The Fate of Alpine Calcareous Grasslands in Central Apennines (Italy)" Diversity 14, no. 9: 695. https://doi.org/10.3390/d14090695
APA StyleVarricchione, M., Carranza, M. L., Di Cecco, V., Di Martino, L., & Stanisci, A. (2022). Warmer and Poorer: The Fate of Alpine Calcareous Grasslands in Central Apennines (Italy). Diversity, 14(9), 695. https://doi.org/10.3390/d14090695