
Formica picea and F. candida (Hymenoptera: Formicidae): Synonyms or Two Species?


 ,
,
Abstract
1. Introduction
2. Materials and Methods
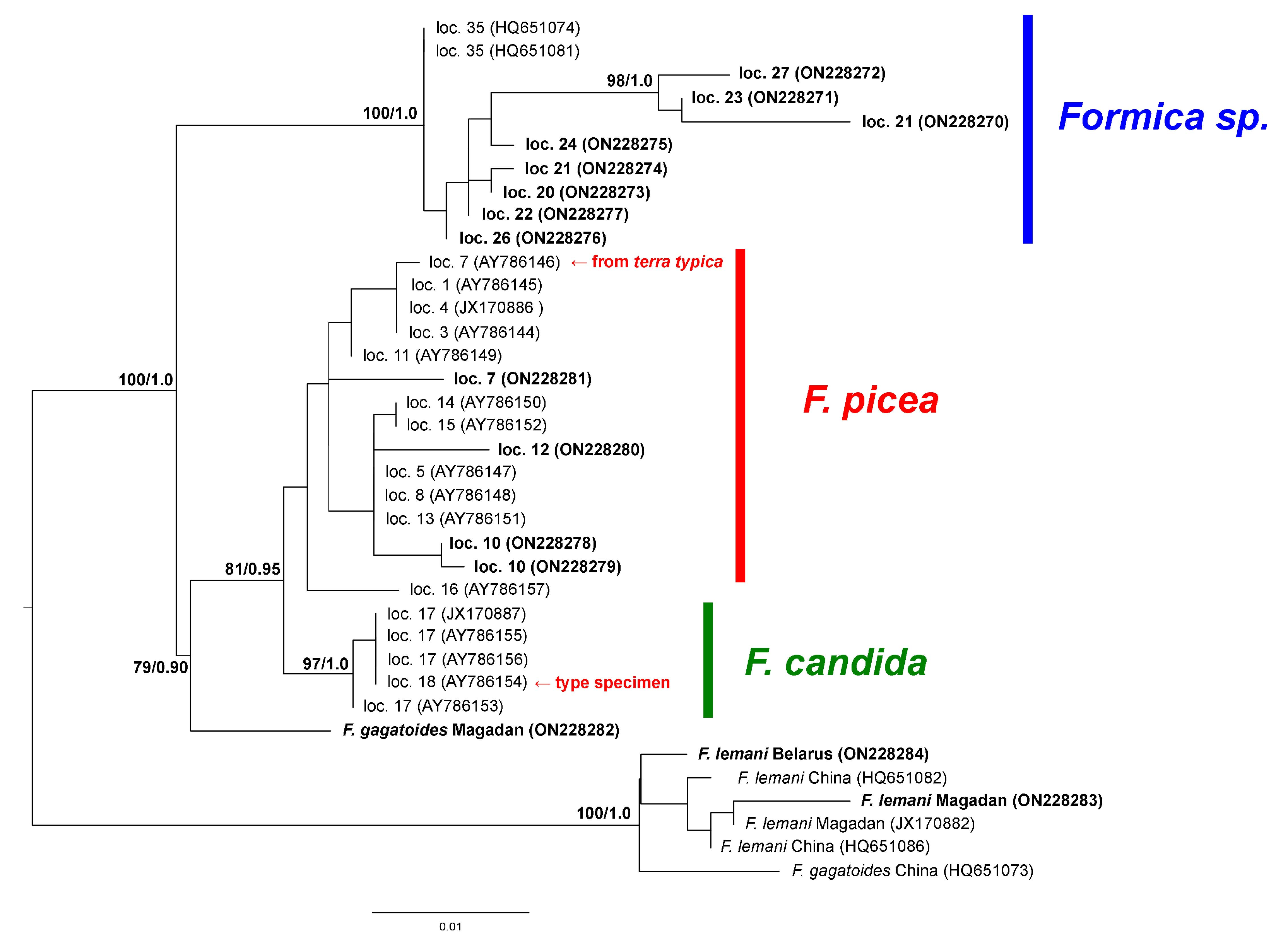
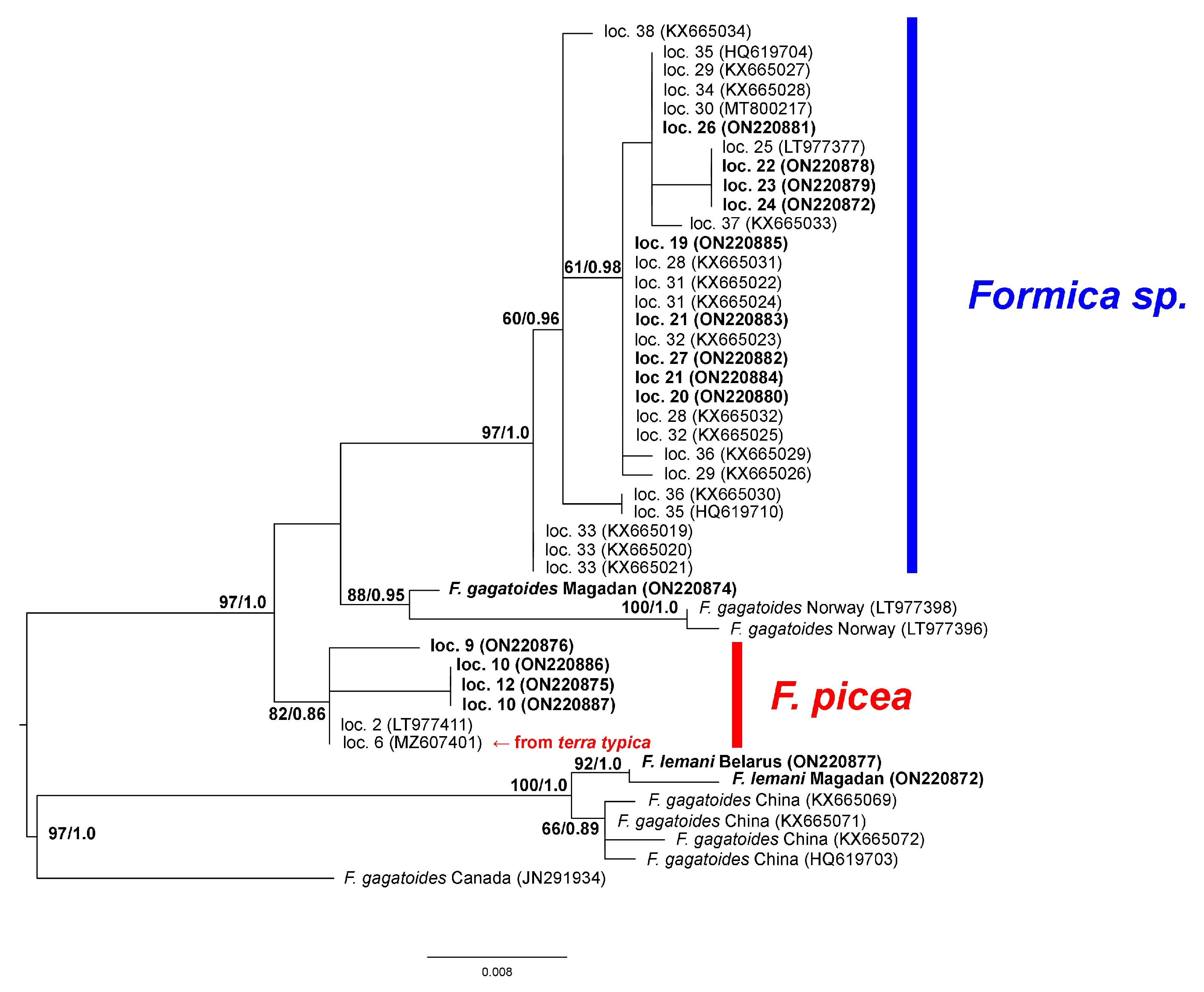
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Borowiec, M.L.; Moreau, C.S.; Rabeling, C. Ants: Phylogeny and Classification. In Encyclopedia of Social Insects; Springer International Publishing: Cham, Switzerland, 2020; pp. 1–18. [Google Scholar]
- Valenzuela-González, J.E.; Pérez-Toledo, G.R.; García-Martínez, M.A. Adelomyrmex dorae sp. nov. Garcia-Martinez (Hymenoptera: Formicidae): A New Species Supported by Parsimony Analysis of Morphological Characters. Trans. Am. Entomol. Soc. 2017, 143, 713–727. [Google Scholar] [CrossRef]
- Goropashnaya, A. Phylogeographic Structure and Genetic Variation in Formica Ants. Ph.D. Thesis, Uppsala University, Uppsala, Sweden, 20 December 2003. [Google Scholar]
- Schär, S.; Talavera, G.; Espadaler, X.; Rana, J.D.; Andersen, A.; Cover, S.P.; Vila, R. Do Holarctic ant species exist? Trans-Beringian dispersal and homoplasy in the Formicidae. J. Biogeogr. 2018, 45, 1917–1928. [Google Scholar] [CrossRef]
- Goropashnaya, A.V.; Fedorov, V.B.; Seifert, B.; Pamilo, P. Phylogenetic relationships of Palaearctic Formica species (Hymenoptera, Formicidae) based on mitochondrial cytochrome b sequences. PLoS ONE 2012, 7, e41697. [Google Scholar] [CrossRef] [PubMed]
- Roslin, T.; Somervuo, P.; Pentinsaari, M.; Hebert, P.D.N.; Agda, J.; Ahlroth, P.; Anttonen, P.; Aspi, J.; Blagoev, G.; Blanco, S.; et al. A molecular-based identification resource for the arthropods of Finland. Mol. Ecol. Resour. 2022, 22, 803–822. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, S. Phylogenetic relationships based on DNA barcoding among 16 species of the ant genus Formica (Hymenoptera: Formicidae) from China. J. Insect Sci. 2017, 17, 117. [Google Scholar] [CrossRef]
- Park, S.; Noh, P.; Kang, J.-Y. Endosymbionts and phage WO infections in Korean ant species (Hymenoptera: Formicidae). Proc. Natl. Inst. Ecol. Repub. Korea 2020, 1, 52–57. [Google Scholar] [CrossRef]
- Chen, Z.; Zhou, S.Y.; Ye, D.; Chen, Y.; Lu, C. Molecular phylogeny of the ant subfamily Formicinae (Hymenoptera, Formicidae) from China Based on mitochondrial genes. Sociobiology 2013, 60, 135–144. [Google Scholar] [CrossRef][Green Version]
- Shekhovtsov, S.V.; Berman, D.I.; Bulakhova, N.A.; Makarova, O.L.; Peltek, S.E. Phylogeography of earthworms from high latitudes of Eurasia. Acta Zool. Acad. Sci. Hung. 2018, 64, 369–382. [Google Scholar] [CrossRef]
- Dlussky, G.M. Ants of the Genus Formica (Hymenoptera: Formicidae g. Formica); Nauka: Moscow, Russia, 1967; p. 236. [Google Scholar]
- Bolton, B. A New General Catalogue of the Ants of the World; Harvard University Press: Cambridge, UK, 1995; p. 502. [Google Scholar]
- Seifert, B. The “Black Bog Ant” Formica picea Nylander, 1846—A species different from Formica candida Smith, 1878 (Hymenoptera: Formicidae). Myrmecolog. Nachr. 2004, 6, 29–38. [Google Scholar]
- Zakharov, A.A.; Dlussky, G.M.; Goryunov, D.N.; Gilyov, A.V.; Zryanin, V.A.; Fedoseeva, E.B.; Gorokhovskaya, E.A.; Radchenko, A.G. Monitoring of the Formica Ants; KMK Scientific Press: Moscow, Russia, 2013; p. 99. [Google Scholar]
- Ward, P.S.; Brady, S.G.; Fisher, B.L.; Schultz, T.R. The evolution of myrmicine ants: Phylogeny and biogeography of a hyperdiverse ant clade (Hymenoptera: Formicidae). Syst. Entomol. 2015, 40, 61–81. [Google Scholar] [CrossRef]
- Antonov, I.A.; Bukin, Y.S. Molecular phylogenetic analysis of the ant genus Formica L. (Hymenoptera: Formicidae) from Palearctic region. Russ. J. Genet. 2016, 52, 810–820. [Google Scholar] [CrossRef]
- Berman, D.I.; Alfimov, A.V.; Zhigulskaya, Z.A.; Leirikh, A.N. Overwintering and Cold-Hardiness of Ants in the Northeast of Asia; Pensoft Publishers: Sofia-Moscow, Russia, 2010; p. 294. [Google Scholar]
- Radchenko, A.G. Ants (Hymenoptera, Formicidae) of Ukraine; Institute of Zoology of Ukraine NAS: Kyiv, Ukraine, 2016; p. 495. [Google Scholar]
- Folmer, O.; Hoeh, W.R.; Black, M.B.; Vrijenhoek, R.C. Conserved primers for PCR amplification of mitochondrial DNA from different invertebrate phyla. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Shekhovtsov, S.V.; Shipova, A.A.; Poluboyarova, T.V.; Vasiliev, G.V.; Golovanova, E.V.; Geraskina, A.P.; Bulakhova, N.A.; Szederjesi, T.; Peltek, S.E. Species delimitation of the Eisenia nordenskioldi complex (Oligochaeta, Lumbricidae) using transcriptomic data. Front. Genet. 2020, 11, 1508. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModeltest Ver. 2; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Seifert, B. Cryptic species in ants (Hymenoptera: Formicidae) revisited: We need a change in the alpha-taxonomic approach. Myrmecol. News 2009, 12, 149–166. [Google Scholar]
- Bernasconi, C.; Cherix, D.; Seifert, B.; Pamilo, P. Molecular taxonomy of the Formica rufa group (red wood ants) (Hymenoptera: Formicidae): A new cryptic species in the Swiss Alps. Myrmecol. News 2011, 14, 37–47. [Google Scholar]
- Seifert, B. Inconvenient hyperdiversity—The traditional concept of “Pheidole pallidula” includes four cryptic species (Hymenoptera: Formicidae). SOIL Org. 2016, 88, 1–17. [Google Scholar]
- Fournier, D.; Tindo, M.; Kenne, M.; Mbenoun Masse, P.S.; Van Bossche, V.; De Coninck, E.; Aron, S. Genetic structure, nestmate recognition and behaviour of two cryptic species of the invasive big-headed ant Pheidole megacephala. PLoS ONE 2012, 7, e31480. [Google Scholar] [CrossRef]
- Seifert, B.; Yazdi, A.B.; Schultz, R. Myrmica martini sp. n.—A cryptic species of the Myrmica scabrinodis species complex (Hymenoptera: Formicidae) revealed by geometric morphometrics and nest-centroid clustering. Myrmecol. News 2014, 19, 171–183. [Google Scholar]
- Ebsen, J.R.; Boomsma, J.J.; Nash, D.R. Phylogeography and cryptic speciation in the Myrmica scabrinodis Nylander, 1846 species complex (Hymenoptera: Formicidae), and their conservation implications. Insect Conserv. Divers. 2019, 12, 467–480. [Google Scholar] [CrossRef]
- Seifert, B. A taxonomic revision of the Palaearctic members of the Formica rufa group (Hymenoptera: Formicidae)–The famous mound-building red wood ants. Myrmecol. News 2021, 31, 133–179. [Google Scholar] [CrossRef]
- Pamilo, P.; Kulmuni, J. Genetic identification of Formica rufa group species and their putative hybrids in northern Europe. Myrmecol. News 2022, 32. [Google Scholar] [CrossRef]
- Sanllorente, O.; Lorite, P.; Ruano, F.; Palomeque, T.; Tinaut, A. Phylogenetic relationships between the slave-making ants Rossomyrmex and their Proformica hosts in relation to other genera of the ant tribe Formicini (Hymenoptera: Formicidae). J. Zool. Syst. Evol. Res. 2018, 56, 48–60. [Google Scholar] [CrossRef]
- Smith, F. Scientific Results of the Second Yarkand Mission; Based upon the Collections and Notes of the Late Ferdinand Stoliczka, Ph.D.; Superintendent of Government Printing (Government of India): Calcutta, India, 1878; p. 22. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
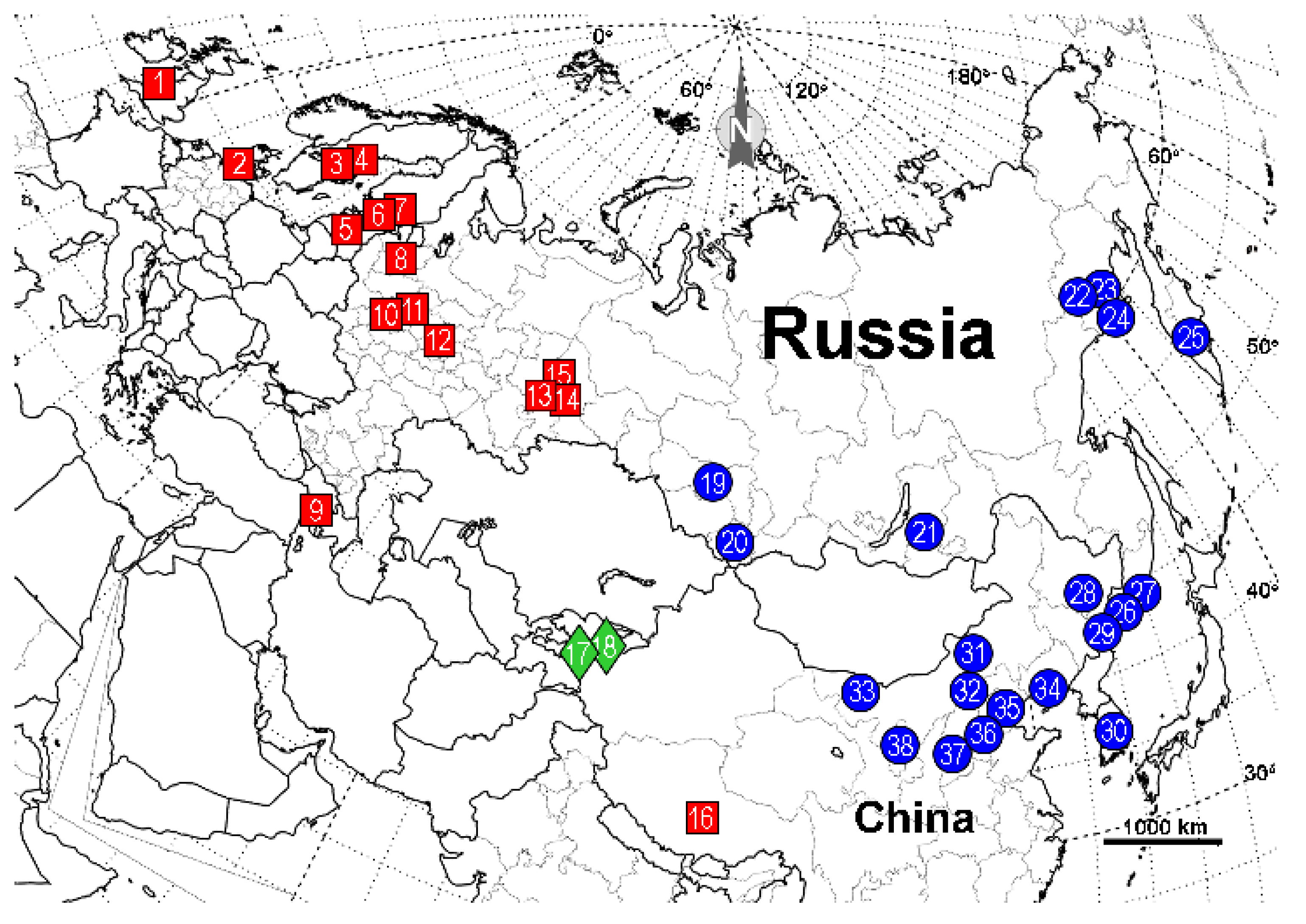
| Location | Species | cox1 | cytb |
|---|---|---|---|
| Formica picea | |||
| 1 | United Kingdom: Cors Goch National Nature Reserve, S. Wales | - | AY786145 (as F. candida) [3] |
| 2 | Denmark: N55.83 E12.56 | LT977411 [4] | - |
| 3 | Sweden: Trajmossen, Brattforsheden | - | AY786144 (as F. candida) [3] |
| 4 | Sweden: Varmland | - | JX170886 [5] |
| 5 | Estonia: Torma | - | AY786147 (as F. candida) [3] |
| 6 | Finland: Nylandia, Raasepori, Harpar Stortraesket | MZ607401 * [6] | - |
| 7 | Finland: Helsinki, Vihti | - | AY786146 * (as F. candida) [3] |
| 8 | Russia: Novgorod oblast | - | AY786148 (as F. candida) [3] |
| 9 | Armenia: Aragaz mtn, N40.43203 E44.23642 | ON220876 | ON228281 |
| 10 | Russia: Moscow oblast, Odintsovo district, raised bog, moss tussock, N55.733 E36.850 | ON220886-87 | ON228278-79 |
| 11 | Russia: Moscow oblast, collected by G. Dlussky in 2000 | - | AY786149 (as F. candida) [3] |
| 12 | Russia: Nizhniy, Novgorod oblast, Kerzhenskiy Natural Reserve, Vishenskoye swamp, N56.467 E44.500 | ON220875 | ON228280 |
| 13 | Russia: Sverdlovsk oblast, Revda | - | AY786151 (as F. candida) [3] |
| 14 | Russia: Sverdlovsk oblast, Yekaterinburg | - | AY786150 (as F. candida) [3] |
| 15 | Russia: Perm oblast, 30 km SSW of Kachkanar | - | AY786152 (as F. candida) [3] |
| 16 | China: Quinghai, Tibet | - | AY786157 (as F. candida) [3] |
| Formica candida | |||
| 17 | Kyrgyzstan: Alai Valley | - | AY786154 **, AY786153 [3], JX170887 [5] |
| 18 | Kyrgyzstan: Tian-Shan | - | AY786155-AY786156 [3] |
| Formica sp. | |||
| 19 | Russia: Novosibirsk oblast, Iskitim district, Listvyanskiy town, N54.450 E83.483 | ON220885 | - |
| 20 | Russia: Altai Republic, Ongudai town, N50.733 E86.133 | ON220880 | ON228273 |
| 21 | Russia: Buryatia Republic, Kizhinga town, N51.84639 E109.91278 | ON220883-84 | ON228270, ON228274 |
| 22 | Russia: Magadan oblast, 15 km from Yagodnoye town, bank of Sokhatina river, N62.517 E149.617 | ON220878 | ON228277 |
| 23 | Russia: Magadan oblast, 16 km from Yagodnoye town, bank of Detrin river, N62.750 E150.467 | ON220879 | ON228271 |
| 24 | Russia: Magadan oblast, Arman town, high floodplain, N59.667 E150.117 | ON220872 | ON228275 |
| 25 | Russia, Kamchatka oblast, N53.19 E158.48 | LT977377 (as F. candida) [4] | - |
| 26 | Russia: Primorye krai, Lazo town, Lazovka river, N43.38056 E133.89778 | ON220881 | ON228276 |
| 27 | Russia: Primorye krai, Sinii Utes town, N43.06222 E131.35472 | ON220882 | ON228272 |
| 28 | China: Harbin, HelongJiang | KX665031-32 (as F. candida) [7] | - |
| 29 | China: Yanbian, Jilin | KX665026-27 (as F. candida) [7] | - |
| 30 | South Korea | MT800217 (as F. candida) [8] | - |
| 31 | Xilin Gol League, Inner Mongolia | KX665022, KX665024 (as F. candida) [7] | - |
| 32 | Ulanqab, Inner Mongolia | KX665023, KX665025 (as F. candida) [7] | - |
| 33 | Helan Mountains, Inner Mongolia | KX665019-21 (as F. candida) [7] | - |
| 34 | Qinglong, Hebei | KX665028 (as F. candida) [7] | - |
| 35 | China: Hebei | HQ619704, HQ619710 (as F. candida) [9] | HQ651081, HQ651074 (as F. candida) [9] |
| 36 | Xiaowutai Mountain, Hebei | KX665029-30 (as F. candida) [7] | - |
| 37 | Zhong-Tiao Mountains, Shanxi | KX665033 (as F. candida) [7] | - |
| 38 | Liupan Mountains, Ningxia | KX665034 (as F. candida) [7] | |
| Outgroups | |||
| F. lemani Belarus: Vitebsk oblast, Braslav district, bank of Uklya lake, N55.60001 E27.333918 | ON220877 | ON228284 | |
| F. lemani Magadan, Arman town high river terrace, N59.867 E150.567 | ON220872 | ON228283 | |
| F. lemani China | - | HQ651082, HQ651086 [9] | |
| F. lemani Magadan | - | JX170882 [10] | |
| F. gagatoides China: Ningxia | - | HQ651073 [9] | |
| F. gagatoides China | KX665069, KX665071, KX665072 [7], HQ619703 | - | |
| F. gagatoides Canada | JN291934 | - | |
| F. gagatoides Norway | LT977396, LT977398 [4] | - | |
| F. gagatoides Russia: Magadan oblast, Arman town, N59.667 E150.117 | ON220874 | ON228282 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhigulskaya, Z.A.; Shekhovtsov, S.V.; Poluboyarova, T.V.; Berman, D.I. Formica picea and F. candida (Hymenoptera: Formicidae): Synonyms or Two Species? Diversity 2022, 14, 613. https://doi.org/10.3390/d14080613
Zhigulskaya ZA, Shekhovtsov SV, Poluboyarova TV, Berman DI. Formica picea and F. candida (Hymenoptera: Formicidae): Synonyms or Two Species? Diversity. 2022; 14(8):613. https://doi.org/10.3390/d14080613
Chicago/Turabian StyleZhigulskaya, Zoya A., Sergei V. Shekhovtsov, Tatiana V. Poluboyarova, and Daniil I. Berman. 2022. "Formica picea and F. candida (Hymenoptera: Formicidae): Synonyms or Two Species?" Diversity 14, no. 8: 613. https://doi.org/10.3390/d14080613
APA StyleZhigulskaya, Z. A., Shekhovtsov, S. V., Poluboyarova, T. V., & Berman, D. I. (2022). Formica picea and F. candida (Hymenoptera: Formicidae): Synonyms or Two Species? Diversity, 14(8), 613. https://doi.org/10.3390/d14080613