
Prey Selectivity in Juvenile Red King Crabs from the Coastal Barents Sea



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling
2.3. Sample Analysis
2.4. Statistical Analysis
3. Results
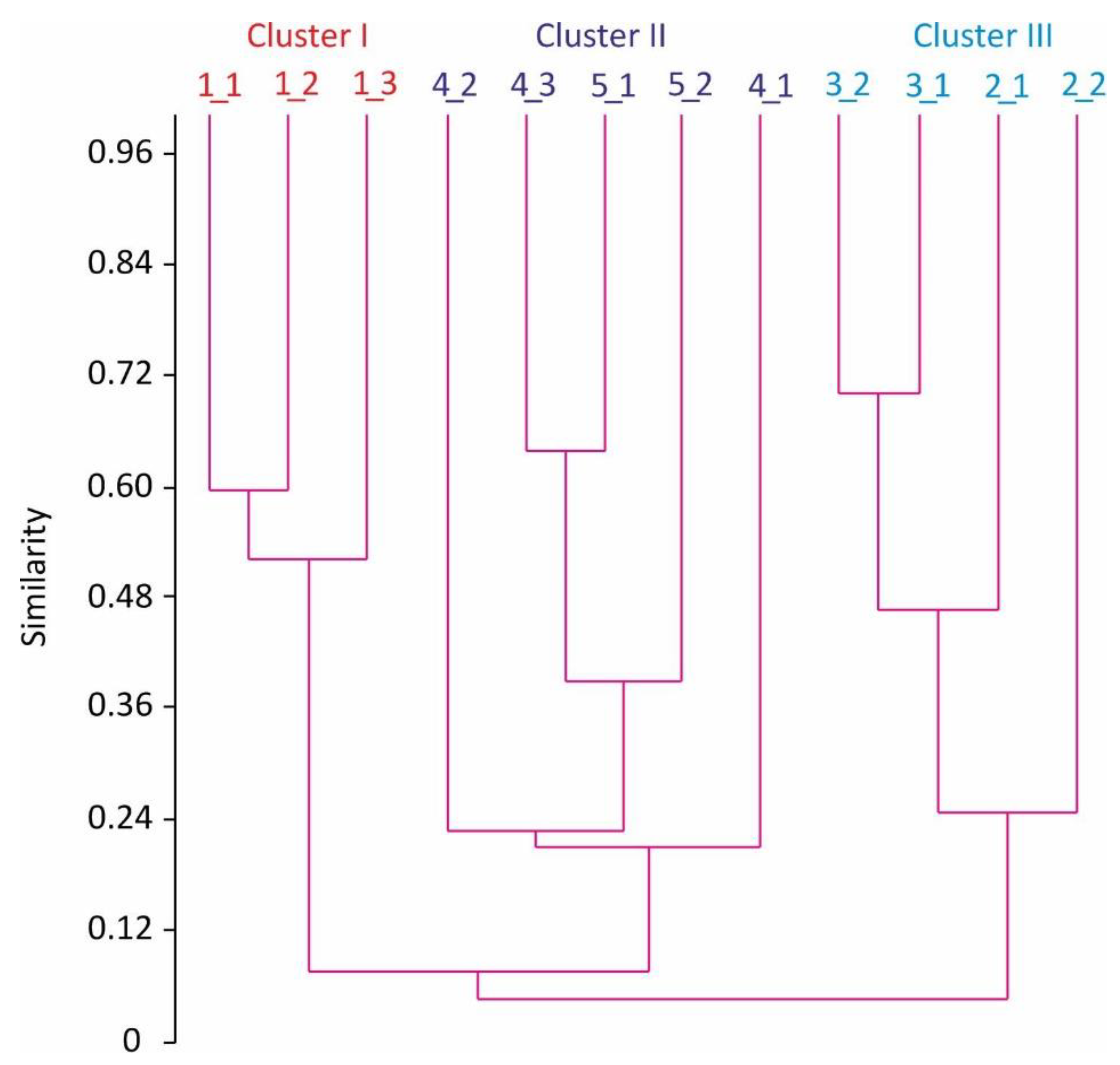
3.1. Red King Crabs
3.2. Benthic Communities
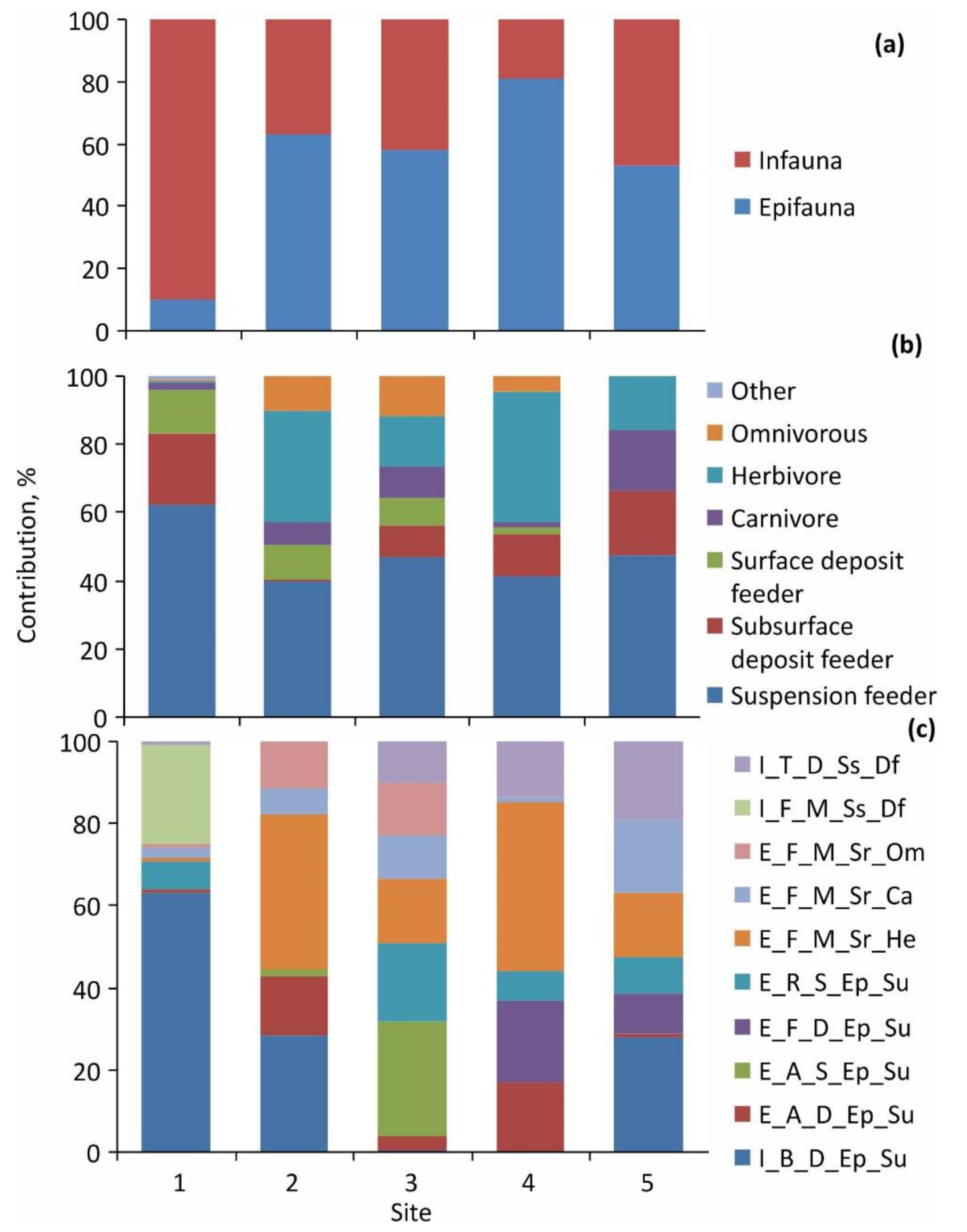
3.3. Diet Composition
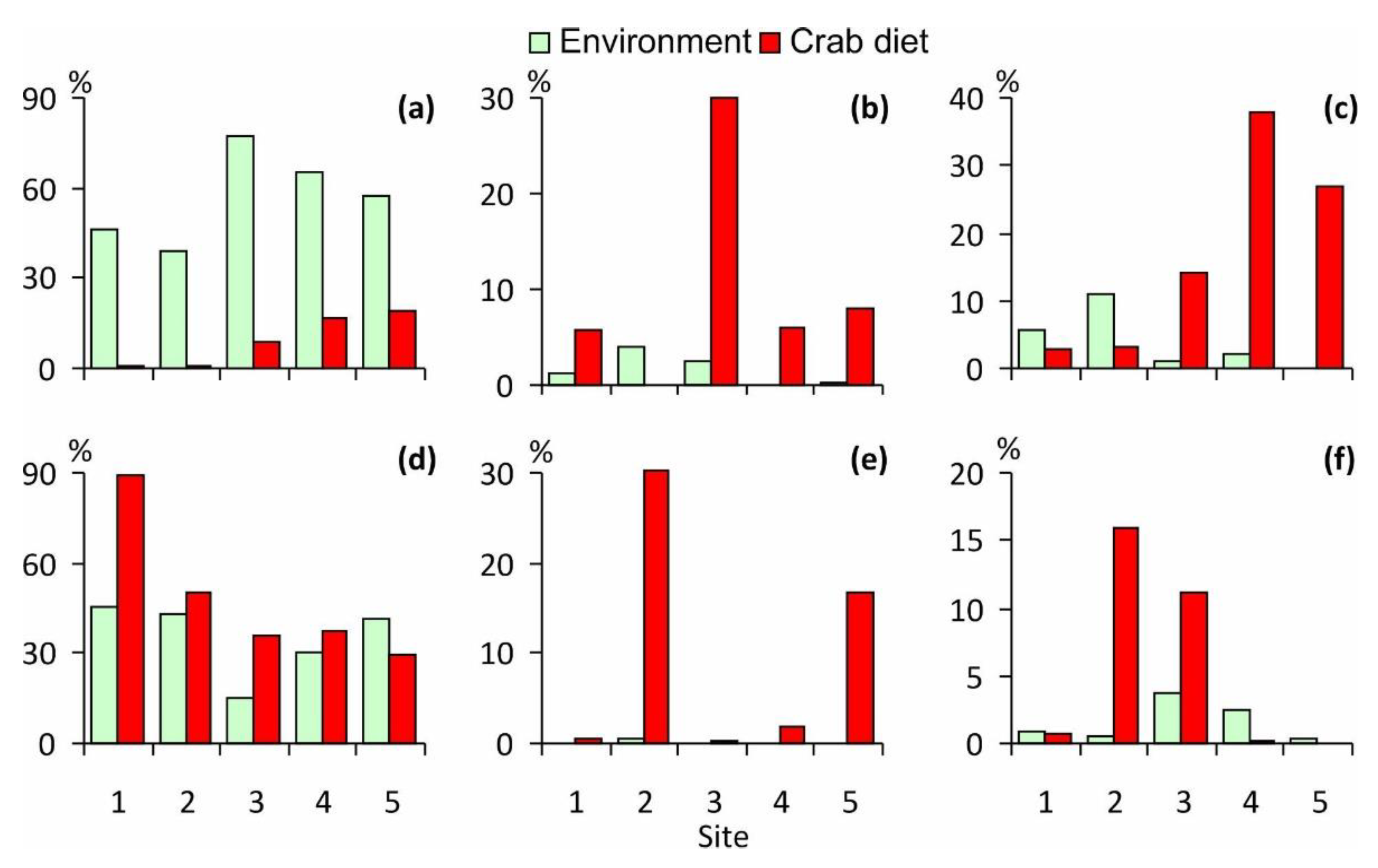
3.4. Comparison of the Crab Diet with Benthic Fauna
3.5. Prey Selectivity
4. Discussion
4.1. Red King Crabs
4.2. Benthic Communities
4.3. Diet Composition
4.4. Prey Selectivity
4.5. Impact of Red King Crabs on Benthic Communities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chan, F.T.; Briski, E. An overview of recent research in marine biological invasions. Mar. Biol. 2017, 164, 121. [Google Scholar] [CrossRef] [PubMed]
- Giangrande, A.; Pierri, C.; Del Pasqua, M.; Gravili, C.; Gambi, M.C.; Gravina, M.F. The Mediterranean in check: Biological invasions in a changing sea. Mar. Ecol. 2020, 41, e12583. [Google Scholar] [CrossRef]
- Simberloff, D. Biological invasions: What’s worth fighting and what can be won? Ecol. Eng. 2014, 65, 112–121. [Google Scholar] [CrossRef]
- Zieritz, A.; Gallardo, B.; Baker, S.J.; Britton, J.R.; Van Valkenburg, J.L.; Verreycken, H.; Aldridge, D.C. Changes in pathways and vectors of biological invasions in Northwest Europe. Biol. Invas. 2017, 19, 269–282. [Google Scholar] [CrossRef]
- Carlton, J.T. The global dispersal of marine and estuarine crustaceans. In In the Wrong Place: Alien Marine Crustaceans—Distribution, Biology and Impacts; Galil, B.S., Clark, P.F., Carlton, J.T., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 3–23. [Google Scholar]
- Chan, F.T.; Stanislawczyk, K.; Sneekes, A.C.; Dvoretsky, A.; Gollasch, S.; Minchin, D.; David, M.; Jelmert, A.; Albretsen, J.; Bailey, S.A. Climate change opens new frontiers for marine species in the Arctic: Current trends and future invasion risks. Glob. Change Biol. 2019, 25, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Dvoretsky, A.G.; Dvoretsky, V.G. Red king crab (Paralithodes camtschaticus) fisheries in Russian waters: Historical review and present status. Rev. Fish Biol. Fish. 2018, 28, 331–353. [Google Scholar] [CrossRef]
- Kuzmin, S.A.; Gudimova, E.N. Introduction of the Kamchatka (Red King) Crab in the Barents Sea. Peculiarities of Biology, Perspectives of Fishery; KSC RAS Press: Apatity, Russia, 2002. (In Russian) [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Commercial fish and shellfish in the Barents Sea: Have introduced crab species affected the population trajectories of commercial fish? Rev. Fish Biol. Fish. 2015, 25, 297–322. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Inter-annual dynamics of the Barents Sea red king crab (Paralithodes camtschaticus) stock indices in relation to environmental factors. Polar Sci. 2016, 10, 541–552. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. New echinoderm-crab epibiotic associations from the coastal Barents Sea. Animals 2021, 11, 917. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Epibiotic communities of common crab species in the coastal Barents Sea: Biodiversity and infestation patterns. Diversity 2022, 14, 6. [Google Scholar] [CrossRef]
- Bakanev, S.V. Red king crab (Paralithodes camtschaticus) of the Barents Sea. In Materials of Total Allowable Catches of Water Biological Resources in Fishing Areas in Inland Seas of the Russian Federation, on the Continental Shelf of the Russian Federation, in the Exclusive Economical Zone of the Russian Federation, in the Azov and Caspian Seas in 2023; Sologub, D.O., Ed.; FGBUN VNIRO (Polar branch): Murmansk, Russia, 2022; pp. 10–30. (In Russian) [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Renewal of the recreational red king crab fishery in Russian waters of the Barents Sea: Potential benefits and costs. Mar. Policy 2022, 136, 104916. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Bichkaeva, F.A.; Baranova, N.F.; Dvoretsky, V.G. Fatty acid composition of the Barents Sea red king crab (Paralithodes camtschaticus) leg meat. J. Food Compos. Anal. 2021, 98, 103826. [Google Scholar] [CrossRef]
- Ponomareva, T.; Timchenko, M.; Filippov, M.; Lapaev, S.; Sogorin, E. Prospects of red king crab hepatopancreas processing: Fundamental and applied biochemistry. Recycling 2021, 6, 3. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Bichkaeva, F.A.; Baranova, N.F.; Dvoretsky, V.G. Fatty acid composition in the hepatopancreas of the Barents Sea red king crab. Biol. Bull. 2020, 47, 332–338. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Bichkaeva, F.A.; Baranova, N.F.; Dvoretsky, V.G. Fatty acids in the circulatory system of an invasive king crab from the Barents Sea. J. Food Compos. Anal. 2022, 110, 104528. [Google Scholar] [CrossRef]
- Pavlova, L.V. Effect of juvenile red king crabs on zoobenthos in Kola Bay (Barents Sea). Dokl. Biol. Sci. 2008, 422, 312–315. [Google Scholar] [CrossRef]
- Pavlova, L.V. Red king crab trophic relations and its influence on bottom biocenoses. In Biology and Physiology of the Red King Crab from the Coastal Zone of the Barents Sea; Matishov, G.G., Ed.; KSC RAS Press: Apatity, Russia, 2008; pp. 77–104. (In Russian) [Google Scholar]
- Britayev, T.A.; Rzhavsky, A.V.; Pavlova, L.V.; Dvoretskij, A.G. Studies on impact of the alien Red King Crab (Paralithodes camtschaticus) on the shallow water benthic communities of the Barents Sea. J. Appl. Ichthyol. 2010, 26, 66–73. [Google Scholar] [CrossRef]
- Oug, E.; Cochrane, S.K.J.; Sundet, J.H.; Norling, K.; Nilsson, H.C. Effects of the invasive red king crab (Paralithodes camtschaticus) on soft-bottom fauna in Varangerfjorden, northern Norway. Mar. Biodiv. 2011, 41, 467–479. [Google Scholar] [CrossRef]
- Oug, E.; Sundet, J.H.; Cochrane, S.K.J. Structural and functional changes of soft-bottom ecosystems in northern fjords invaded by the red king crab (Paralithodes camtschaticus). J. Mar. Syst. 2018, 180, 255–264. [Google Scholar] [CrossRef]
- Fuhrmann, M.M.; Pedersen, T.; Ramasco, V.; Nilssen, E.M. Macrobenthic biomass and production in a heterogenic subarctic fjord after invasion by the red king crab. J. Sea Res. 2015, 106, 1–13. [Google Scholar] [CrossRef]
- Falk-Petersen, J.; Renaud, P.; Anisimova, N. Establishment and ecosystem effects of the alien invasive red king crab (Paralithodes camtschaticus) in the Barents Sea—A review. ICES J. Mar. Sci. 2011, 68, 479–488. [Google Scholar] [CrossRef]
- Pavlova, L.V.; Britayev, T.A.; Rzhavsky, A.V. Benthos elimination by juvenile red king crabs Paralithodes camtschaticus (Tilesius, 1815) in the Barents Sea coastal zone: Experimental data. Dokl. Biol. Sci. 2007, 414, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Pavlova, L.V. Ration of the red king crab on coastal shoals of the Barents Sea. Dokl. Biol. Sci. 2015, 463, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Fuhrmann, M.M.; Pedersen, T.; Nilssen, E.M. Trophic niche of the invasive red king crab Paralithodes camtschaticus in a benthic food web. Mar. Ecol. Prog. Ser. 2017, 565, 113–129. [Google Scholar] [CrossRef]
- Pavlova, L.V. Estimation of foraging on the sea urchin Strongylocentrotus droebachiensis (Echinoidea: Echinoida) by the red king crab Paralithodes camtschaticus (Malacostraca: Decapoda) in coastal waters of the Barents Sea. Russ. J. Mar. Biol. 2009, 35, 288–295. [Google Scholar] [CrossRef]
- Pavlova, L.V. The red king crab Paralithodes camchaticus (Tilesius, 1815) (Decapoda: Anomura): The use of species equality indicators to assess the influence on the benthos of the Barents Sea. Russ. J. Mar. Biol. 2021, 47, 508–514. [Google Scholar] [CrossRef]
- Micheli, F. Behavioral plasticity in prey-size selectivity of the blue crab Callinectes sapidus feeding on bivalve prey. J. Anim. Ecol. 1995, 64, 63–74. [Google Scholar] [CrossRef]
- Romero, M.C.; Lovrich, G.A.; Tapella, F.; Thatje, S. Feeding ecology of the crab Munida subrugosa (Decapoda: Anomura: Galatheidae) in the Beagle Channel, Argentina. J. Mar. Biol. Ass. UK 2004, 84, 359–365. [Google Scholar] [CrossRef]
- Baeta, A.; Cabral, H.N.; Marques, J.C.; Pardal, M.A. Feeding ecology of the green crab, Carcinus maenas (L., 1758) in temperate estuary, Portugal. Crustaceana 2007, 79, 1181–1193. [Google Scholar]
- Seitz, R.D.; Knick, K.E.; Westphal, M. Diet selectivity of juvenile blue crabs (Callinectes sapidus) in Chesapeake Bay. Integr. Compar. Biol. 2011, 51, 598–607. [Google Scholar] [CrossRef]
- Jørgensen, L.L.; Primicerio, R. Impact scenario for the invasive red king crab Paralithodes camtschaticus (Tilesius, 1815) (Reptantia, Lithodidae) on Norwegian, native, epibenthic prey. Hydrobiologia 2007, 590, 47–54. [Google Scholar] [CrossRef]
- Michelsen, H.K. Prey Selection of the Red King Crab (Paralithodes camtschaticus) Preying on Lumpfish Eggs (Cyclopterus lumpus), Sea Urchins (Strongylocentrotus droebachiensis) and Scallops (Chlamys islandica). Master’s Thesis, University of Tromsø, Tromsø, Norway, 2011. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Population dynamics of the invasive lithodid crab, Paralithodes camtschaticus, in a typical bay of the Barents Sea. ICES J. Mar. Sci. 2013, 70, 1255–1262. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Effects of environmental factors on the abundance, biomass, and individual weight of juve-nile red king crabs in the Barents Sea. Front. Mar. Sci. 2020, 7, 726. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Life cycle of Oithona similis (Copepoda, Cyclopoida) in Kola Bay (Barents Sea). Mar. Biol. 2009, 156, 1433–1446. [Google Scholar] [CrossRef]
- Deryugin, K.M. Fauna of the Kola Fjord and the conditions of its existence. Trans. Imp. Acad. Sci. St. Petersburg 1915, 34, 1–929. (In Russian) [Google Scholar]
- Dzhenyuk, S.L.; Savelieva, S.P. Hydrological characteristics. In The Kola Bay: Oceanography, Biology, Ecosystems, Pollutants; Matishov, G.G., Ed.; KSC RAS Press: Apatity, Russia, 1997; pp. 51–58. (In Russian) [Google Scholar]
- Zuyev, Y.A. The Upper Sublitoral Megabenthos of Kola Bay (Barents Sea). Ph.D. Thesis, MMBI RAS, Murmansk, Russia, 2012. (In Russian). [Google Scholar]
- Potanin, V.A.; Larin, B.V. Dynamics of waters of the southern part of the Kola Bay. In Nature and Economy of the North; Anonymous, Ed.; Book Press: Murmansk, Russia, 1989; pp. 66–71. (In Russian) [Google Scholar]
- Pavlova, L.V.; Nekhaev, I.O.; Panteleeva, N.N.; Akhmetchina, O.Y.; Garbul, E.A.; Dikaeva, D.R.; Zimina, O.L.; Lyubina, O.C.; Frolov, A.A.; Frolova, E.A. Shallow benthos of Kola Bay (Barents Sea): Biodiversity and assessment of the current state of communities. Transact. Kola Sci. Cent. RAS. Ser. 5 2018, 4, 61–92. (In Russian) [Google Scholar]
- Pavlova, L.V.; Zuyev, Y.A. The red king crab (Paralithodes camtschaticus) in Kola bay: Distribution, population and size/age structure changes from 2006 to 2008. Rybn. Khoz. 2010, 6, 66–69. (In Russian) [Google Scholar]
- Blinova, E.I.; Vilkova, O.Y.; Milyutin, D.M.; Pronina, O.A.; Shtrik, B.A. Methods of Landscape Research and Assessment of Stocks of Benthic Invertebrates and Algae in the Marine Coastal Zone. Study of Ecosystems of Unsustainable Water Bodies, Collection and Processing of Data on Aquatic Biological Resources, Equipment and Technology for their Extraction and Processing; VNIRO: Moscow, Russia, 2005. (In Russian) [Google Scholar]
- Donaldson, W.E.; Byersdorfer, S.E. Biological Field Techniques for Lithodid Crabs; University of Alaska: Fairbanks, AK, USA, 2005. [Google Scholar]
- Brotskaya, V.A.; Zenkevich, L.A. Counting of the Barents Sea bottom-living fauna. Tr. VNIRO 1939, 4, 3–150. (In Russian) [Google Scholar]
- McDonald, P.S.; Holsman, K.K.; Beauchamp, D.A.; Dumbauld, B.R.; Armstrong, D.A. Bioenergetics modeling to investigate habitat use by the non-indigenous crab, Carcinus maenas, in Willapa Bay, Washington, USA. Estuar. Coasts 2006, 29, 1132–1149. [Google Scholar] [CrossRef]
- Jumars, P.A.; Dorgan, K.M.; Lindsay, S.M. Diet of worms emended: An update of polychaete feeding guilds. Annu. Rev. Mar. Sci. 2015, 7, 497–520. [Google Scholar] [CrossRef]
- Rzhavkiy, A.V.; Pereladov, M.V. Feeding of king crab (Paralithodes camtschaticus) on the Varanger-fjord shoalness (Barents Sea): Studies of the alimentary canal content and visual observations. Tr. VNIRO 2003, 51, 120–130. (In Russian) [Google Scholar]
- Tarverdieva, M.I. On the feeding of immature king crab (Paralithodes camtschaticus) in the Teriberka Bay of the Barents Sea. Tr. VNIRO 2003, 142, 92–102. (In Russian) [Google Scholar]
- Ivlev, V.S. Experimental Ecology of the Feeding of Fishes; Yale University Press: New Haven, CT, USA, 1961. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Size-at-age of juvenile red king crab (Paralithodes camtschaticus) in the coastal Barents Sea. Cah. Biol. Mar. 2014, 55, 43–48. [Google Scholar]
- Powell, G.C.; Nickerson, R.B. Aggregations among juvenile king crab (Paralithodes camtschaticus), Kodiak, Alaska. Anim. Behav. 1965, 13, 374–380. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Tipisova, E.V.; Elfimova, A.E.; Alikina, V.A.; Dvoretsky, V.G. Sex hormones in hemolymph of red king crabs from the Barents Sea. Animals 2021, 11, 2149. [Google Scholar] [CrossRef] [PubMed]
- Dvoretsky, A.G.; Dvoretsky, V.G. Size at maturity of female red king crab, Paralithodes camtschaticus, from the costal zone of Kola Peninsula (southern Barents Sea). Cah. Biol. Mar. 2015, 56, 49–54. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Hemolymph molting hormone concentrations in red king crabs from the Barents Sea. Polar Biol. 2010, 33, 1293–1298. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Does spine removal affect molting process in the king red crab (Paralithodes camtschaticus) in the Barents Sea? Aquaculture 2012, 326–329, 173–177. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Tipisova, E.V.; Alikina, V.A.; Elfimova, A.E. Thyroid hormones in hemolymph of red king crabs from the Barents Sea. Animals 2022, 12, 379. [Google Scholar] [CrossRef]
- Deart, Y.V.; Antokhina, T.I.; Spiridonov, V.A.; Britayev, T.A. Seasonal distribution of red king crab in Zelenaya Inlet (Murmansk coast, Barents Sea). Tr. VNIRO 2018, 172, 149–159. (In Russian) [Google Scholar] [CrossRef]
- Spiridonov, V.A.; Zalota, A.K.; Pereladov, M.V.; Deart, Y.V.; Tiunov, A.V.; Britayev, T.A. Fjordic lagoons of the Barents Sea as models for study of the dynamics of coastal communities with the introduced red king crab, Paralithodes camtschaticus (Decapoda, Lithodidae). Zool. Zhurn. 2020, 99, 801–818. (In Russian) [Google Scholar]
- Dew, C.B. Behavioral ecology of podding red king crab Paralithodes camtschatica. Can. J. Fish. Aquat. Sci. 1990, 47, 1944–1958. [Google Scholar] [CrossRef]
- Stevens, B.G. Biology and ecology of juvenile king crabs. In King Crabs of the World: Biology and Fisheries Management; Stevens, B.G., Ed.; CRC Press (Taylor and Francis): Boca Raton, FL, USA, 2014; pp. 261–284. [Google Scholar]
- Pardo, L.M.; Andrade, C.; Zenteno-Devaud, L.; Garrido, B.; Rivera, C. Trophic ecology of juvenile southern king crab associated with kelp forest: Evidence of cannibalism. Diversity 2021, 13, 556. [Google Scholar] [CrossRef]
- Britayev, T.A.; Rzhavsky, A.V.; Pavlova, L.V.; Kuzmin, S.A.; Dvoretsky, A.G. Modem state of bottom communities and settlements of rnacrozoobenthos in shal1ow-waters of the Barents Sea and the role of anthropogenic factor in their dynamics. In Dynamics of Marine Ecosystems and Modern Problems of Conservation of Biological Resources of the Russian Seas; Tarasov, V.G., Ed.; Dalnauka: Vladivostok, Russia, 2007; pp. 314–356. (In Russian) [Google Scholar]
- Lyubina, O.S.; Zimina, O.L.; Frolova, E.A.; Frolov, A.A.; Dikaeva, D.R.; Panteleeva, N.N.; Nekhaev, I.O.; Garbul, E.A. Distribution of zoobenthos on soft bottom in the Ivanovskaya and Drozdovka bays of the eastern Murman coast. Dokl. Biol. Sci. 2012, 447, 353–356. [Google Scholar] [CrossRef]
- Deart, Y.V.; Britayev, T.A. “New” benthic community dominated by Oweniidae (Polychaeta, Oweniidae) at the Murman coast: Structure and causes of appearance. Dokl. Biol. Sci. 2014, 454, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Anisimova, N.A.; Frolova, E.A. Benthos of the Dolgaya Bay in the Eastern Murman. Composition, quantitative distribution. In Hydrobiological Essays in Gulfs and Bays of the Russian Arctic Seas; Matishov, G.G., Ed.; KSC Press: Apatity, Russia, 1994; pp. 43–91. (In Russian) [Google Scholar]
- Kennish, M.J.; Haag, S.M.; Sakowicz, G.P.; Durand, J.B. Benthic macrofaunal community structure along a well-defined salinity gradient in the Mullica River-Great Bay estuary. J. Coast. Res. 2004, 45, 209–226. [Google Scholar] [CrossRef]
- Antonio, E.S.; Kasai, A.; Ueno, M.; Ishihi, Y.; Yokoyama, H.; Yamashita, Y. Spatial-temporal feeding dynamics of benthic communities in an estuary-marine gradient. Estuar. Coast. Shelf Sci. 2012, 112, 86–97. [Google Scholar] [CrossRef]
- Kim, S.-L.; Yu, O.-H. Understanding the spatial and temporal distribution and environmental characteristics of polychaete assemblages in the coastal waters of Ulleungdo, East Sea of Korea. J. Mar. Sci. Eng. 2021, 9, 1310. [Google Scholar] [CrossRef]
- Quiroz-Martínez, B.; Hernández-Alcántara, P.; Salas-de-León, D.A.; Solís-Weiss, V. Polychaete (Annelida) diversity patterns in Southern Gulf of Mexico: The influence of spatial structure and environmental variables. Diversity 2021, 13, 425. [Google Scholar] [CrossRef]
- Gerasimova, O.V. Analysis of king crab (Paralithodes camtschatica) trophic links in the Barents Sea. ICES CM 1997, GG:03, 1–21. [Google Scholar]
- Sundet, J.H.; Rafter, E.E.; Nilssen, E. Sex and seasonal variation in the stomach content of the red king crab, Paralithodes camtschaticus in the southern Barents Sea. In The Biodiversity Crisis and Crustacea; Klein, C.V.V., Schram, F.R., Eds.; Balkema Publishers: Rotterdam, The Netherlands, 2000; pp. 193–200. [Google Scholar]
- Manushin, I.; Anisimova, N. Selectivity in the red king crab feeding in the Barents Sea. In Research on the Red King Crab (Paralithodes camtschaticus) from the Barents Sea in 2005–2007; Sundet, J.H., Berenboim, B., Eds.; IMR/PINRO: Bergen/Murmansk, Norway/Russia, 2008; pp. 24–28. [Google Scholar]
- Eggleston, D.B.; Lipcius, R.N.; Hines, A.H. Density-dependent predation by blue crabs upon infaunal clam species with contrasting distribution and abundance patterns. Mar. Ecol. Prog. Ser. 1992, 85, 55–68. [Google Scholar] [CrossRef]
- Hegele-Drywa, J.; Normant, M. Feeding ecology of the American crab Rhithropanopeus harrisii (Crustacea, Decapoda) in the coastal waters of the Baltic Sea. Oceanologia 2009, 51, 361–375. [Google Scholar] [CrossRef][Green Version]
- Rosewarne, P.J.; Mortimer, R.J.; Newton, R.J.; Grocock, C.; Wing, C.D.; Dunn, A.M. Feeding behaviour, predatory functional responses and trophic interactions of the invasive Chinese mitten crab (Eriocheir sinensis) and signal crayfish (Pacifastacus leniusculus). Freshw. Biol. 2016, 61, 426–443. [Google Scholar] [CrossRef]
- Abelló, P. Note on the diet of Lithodes ferox (Anomura: Lithodidae) off Namibia. S. Afr. J. Mar. Sci. 1995, 15, 273–277. [Google Scholar] [CrossRef]
- Divine, L.M.; Bluhm, B.A.; Mueter, F.J.; Iken, K. Diet analysis of Alaska Arctic snow crabs (Chionoecetes opilio) using stomach contents and δ13C and δ15N stable isotopes. Deep-Sea Res. II 2017, 135, 124–136. [Google Scholar] [CrossRef]
- Macdonald, T.A.; Burd, B.J.; Macdonald, V.I.; van Roodselaar, A. Taxonomic and Feeding Guild Classification for the Marine Benthic Macroinvertebrates of the Strait of Georgia, British Columbia; Canadian Technical Report of Fisheries and Aquatic Sciences: Sidney, BC, Canada, 2010; Volume 2874, pp. 1–63. [Google Scholar]
- Logvinovich, D.N. Aquarium observations on the feeding of the Kamchatka crab. Izv. TINRO 1945, 19, 79–97. (In Russian) [Google Scholar]
- Zubkova, N.A. An attempt of king crab keeping in aquaria. Tr. Murmansk Mar. Biol. Inst. 1964, 5, 161–169. (In Russian) [Google Scholar]
- Greenaway, P. Calcium balance and moulting in the Crustacea. Biol. Rev. 1985, 60, 425–454. [Google Scholar] [CrossRef]
- Weber, M.J.; Dettmers, J.M.; Wahl, D.H.; Czesny, S.J. Effects of predator–prey interactions and benthic habitat complexity on selectivity of a foraging generalist. Trans. Am. Fish. Soc. 2010, 139, 1004–1013. [Google Scholar] [CrossRef]
- Elner, R.W.; Hughes, R.N. Energy maximization in the diet of the shore crab, Carcinus maenas. J. Anim. Ecol. 1978, 47, 103–116. [Google Scholar] [CrossRef]
- Lovrich, G.A.; Tapella, F. Southern king crabs. In King Crabs of the World: Biology and Fisheries Management; Stevens, B.G., Ed.; CRC Press (Taylor and Francis): Boca Raton, FL, USA, 2014; pp. 449–484. [Google Scholar]
- Stevens, B.G.; Jewett, S.C. Growth, molting, and feeding of king crabs. In King Crabs of the World: Biology and Fisheries Management; Stevens, B.G., Ed.; CRC Press (Taylor and Francis): Boca Raton, FL, USA, 2014; pp. 315–362. [Google Scholar]
- Andrade, C.; Rivera, C.; Daza, E.; Almonacid, E.; Ovando, F.; Morello, F.; Pardo, L.M. Trophic niche dynamics and diet partitioning of king crab Lithodes santolla in Chile’s Sub-Antarctic water. Diversity 2022, 14, 56. [Google Scholar] [CrossRef]
- Rzhavsky, A.V.; Pavlova, L.V.; Kuzmin, S.A.; Kulikova, V.I. On the distribution of some species of macrobenthos in the Dal’nezelenetskaya Bay (Barents Sea) after red king crab introduction. In Shelf Zoobenthos Investigations. Information Support of the Ecosystem Investigations; Matishov, G.G., Ed.; KSC Press: Apatity, Russia, 2004; pp. 105–116. (In Russian) [Google Scholar]
- Britaev, T.A.; Rzhavsky, A.V.; Pavlova, L.V. 2006. The state of the hard bottom communities of the Barents Sea shallow after the red king crab introduction. In Current State of Crabs’ Populations in the Barents Sea and their Interaction with Bottom Biocenoses; Matishov, G.G., Kuzmin, S.A., Zenzerov, V.S., Eds.; Sever Press: Murmansk, Russia, 2006; pp. 15–18. (In Russian) [Google Scholar]
- Pavlova, L.V. Dynamics of the benthos biomass in the hard bottom of the Dalnezelenetskaya Bay (Barents Sea) in connection with red king crab introduction. In Proceedings of the Materials of the 22nd Conference of Young Scientists of the Murmansk Marine Biological Institute, Murmansk, Russia, May 2014; Matishov, G.G., Ed.; MMBI Press: Murmansk, Russia, 2004; pp. 83–89. (In Russian). [Google Scholar]
- McNair, J.N.; Newbold, J.D.; Hart, D.D. Turbulent transport of suspended particles and dispersing benthic organisms: How long to hit bottom? J. Theor. Biol. 1997, 188, 29–52. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Site | ||||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |
| Average carapace length (min–max), mm | 82 ± 10 (62–98) | 62 ± 4 (57–67) | 27 ± 5 (21–37) | 30 ± 6 (20–39) | 38 ± 5 (30–45) |
| Average weight (min–max), g | 410.8 ± 140.8 (183.7–644.0) | 165.4 ± 40.0 (78.6–214.2) | 13.7 ± 8.4 (5.5–33.1) | 20.0 ± 9.6 (6.0–41.1) | 42.2 ± 20.0 (18.4–54.3) |
| Density, ind. 1000 m–2 | 38 | 63 | 34 | 3 | 2 |
| Sample size | 18 | 13 | 15 | 11 | 10 |
| Parameter | Site | ||||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |
| Number of species | 97 | 104 | 79 | 55 | 62 |
| α-diversity | 59 ± 11 | 71 ± 2 | 44 ± 19 | 36 ± 3 | 46 ± 6 |
| Mean biomass, g m–2 | 21 ± 5 | 73 ± 26 | 80 ± 34 | 58 ± 20 | 41 ± 10 |
| Dominant by biomass | Macoma calcarea Cistenides granulata Crenella decussata | Spirobranchus triqueter Astarte borealis | Spirobranchus triqueter | Alitta virens Laonice cirrata Mytilus edulis | Laonice cirrata Arctica islandica |
| Average abundance (ind. m–2) | 6220 ± 900 | 4420 ± 1800 | 5690 ± 2530 | 5990 ± 1120 | 6470 ± 660 |
| Average weight (min–max), g | 0.003 ± 0.001 (0.001–0.006) | 0.017 ± 0.008 (0.007–0.063) | 0.014 ± 0.006 (0.002–0.035) | 0.010 ± 0.005 (0.001–0.030) | 0.006 ± 0.001 (0.002–0.010) |
| Dominant by abundance | Macoma calcarea | Onoba semicostata | Heteranomia squamula Mytilus edulis | Micronephthys neotena | Chaetozone setosa |
| Epifauna:infauna ratio by biomass | 1:5 | 1.3:1 | 15:1 | 1:2.3 | 1:22 |
| Total number of trophic groups | 6 | 6 | 7 | 6 | 6 |
| Dominant trophic group | Subsurface deposit feeder (36%) | Suspension feeder (78%) | Suspension feeder (85%) | Suspension feeder (34%) | Suspension feeder (52%) |
| Total number of functional groups | 26 | 28 | 23 | 18 | 19 |
| Dominant functional groups | I_F_D_Ss_Df (33%) | I_B_D_Ep_Su (37%) E_T_S_Ep_Su (30%) | E_T_S_Ep_Su (70%) | I_B_M_Sr_Om (30%) E_A_S_Ep_Su (26%) | I_B_D_Ep_Su (41%) I_T_D_Sr_Df (36%) |
| Sediments | Muddy sand, sand with shell debris | Mud, pebbles, boulders | Rock, boulders, mud with pebbles | Muddy sand | Muddy sand |
| Temperature, °C | 3–4 | 3.8 | 4 | 4 | 4 |
| Depth, m | 5–20 | 10–20 | 5–20 | 5–20 | 5–10 |
| Stations | 1_1, 1_2, 1_3 | 2_1, 2_2 | 3_1, 3_2 | 4_1, 4_2, 4_3 | 5_1, 5_2 |
| Cluster I | Cluster II | Cluster I | Cluster III | Cluster II | Cluster III |
|---|---|---|---|---|---|
| Av. Diss = 85.44 | Av. Diss = 88.64 | Av. Diss = 79.23 | |||
| Taxa | Con, % | Taxa | Con, % | Taxa | Con, % |
| Spirobranchus triqueter | 10.20 | Spirobranchus triqueter | 12.63 | Laonice cirrata | 7.95 |
| Macoma calcarea | 5.96 | Laonice cirrata | 8.15 | Macoma calcarea | 6.81 |
| Leitoscoloplos acutus | 5.72 | Heteranomia squamula | 4.49 | Leitoscoloplos acutus | 5.97 |
| Crenella decussata | 4.66 | Alitta virens | 4.46 | Cistenides granulata | 5.76 |
| Heteranomia squamula | 3.65 | Chaetozone setosa | 4.28 | Alitta virens | 4.67 |
| Cistenides granulata | 3.28 | Arctica islandica | 3.65 | Arctica islandica | 4.20 |
| Astarte borealis | 2.90 | Astarte borealis | 3.38 | Onoba semicostata | 3.11 |
| Parvicardium pinnulatum | 2.65 | Balanus balanus | 2.74 | Chaetozone setosa | 3.07 |
| Eteone flava | 2.45 | Mytilus edulis | 2.47 | Parvicardium pinnulatum | 3.03 |
| Onoba semicostata | 2.40 | Hydroides norvegica | 2.35 | Pholoe assimilis | 2.82 |
| Balanus | 2.29 | Scoletoma fragilis | 2.31 | Leitoscoloplos acutus | 2.64 |
| Prey Groups | Site | ||||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |
| Benthic taxon | |||||
| Polychaeta | −1.0 | −1.0 | −0.8 | −0.6 | −0.5 |
| Crustacea | 0.6 | −1.0 | 0.8 | 1.0 | 0.9 |
| Gastropoda | −0.4 | −0.6 | 0.9 | 0.9 | 1.0 |
| Bivalvia | 0.3 | 0.1 | 0.4 | 0.1 | −0.2 |
| Echinodermata | 1.0 | 1.0 | 0.5 | 1.0 | 1.0 |
| Trophic group | |||||
| Suspension-feeder | 0.4 | −0.3 | −0.3 | 0.1 | −0.1 |
| Herbivore | −0.8 | 0.5 | 0.6 | 0.9 | 1.0 |
| Surface deposit-feeder | −0.2 | 0.6 | 0.6 | −0.8 | −1.0 |
| Subsurface deposit-feeder | −0.3 | −0.7 | 0.4 | 0.2 | 0.9 |
| Functional group | |||||
| E_A_D_Ep_Su | 1.0 | 0.9 | −0.2 | −0.2 | 1.0 |
| E_A_S_Ep_Su | – | 0.6 | 1.0 | −1.0 | – |
| E_T_S_Ep_Su | – | −1.0 | −1.0 | – | – |
| E_R_S_Ep_Su | 1.0 | −0.7 | 0.3 | 1.0 | 1.0 |
| E_F_D_Ep_Su | – | 0.1 | −1.0 | 1.0 | 1.0 |
| I_B_D_Ep_Su | 0.7 | −0.2 | 0.0 | −1.0 | −0.2 |
| E_F_M_Sr_He | −0.8 | 0.5 | 0.6 | 0.9 | 1.0 |
| E_F_M_Sr_Ca | −0.2 | 1.0 | 0.6 | −0.4 | 0.8 |
| E_F_M_Sr_Om | 0.9 | 0.9 | 1.0 | −1.0 | −1.0 |
| I_B_D_Sr_Df | −0.2 | −1.0 | −1.0 | −1.0 | −1.0 |
| I_F_M_Ss_Df | 0.1 | −1.0 | −1.0 | −1.0 | −1.0 |
| I_T_D_Ss_Df | −0.9 | −1.0 | 0.6 | 0.6 | 1.0 |
| I_T_D_Sr_Df | −1.0 | −1.0 | −1.0 | −1.0 | −1.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavlova, L.V.; Dvoretsky, A.G. Prey Selectivity in Juvenile Red King Crabs from the Coastal Barents Sea. Diversity 2022, 14, 568. https://doi.org/10.3390/d14070568
Pavlova LV, Dvoretsky AG. Prey Selectivity in Juvenile Red King Crabs from the Coastal Barents Sea. Diversity. 2022; 14(7):568. https://doi.org/10.3390/d14070568
Chicago/Turabian StylePavlova, Lyudmila V., and Alexander G. Dvoretsky. 2022. "Prey Selectivity in Juvenile Red King Crabs from the Coastal Barents Sea" Diversity 14, no. 7: 568. https://doi.org/10.3390/d14070568
APA StylePavlova, L. V., & Dvoretsky, A. G. (2022). Prey Selectivity in Juvenile Red King Crabs from the Coastal Barents Sea. Diversity, 14(7), 568. https://doi.org/10.3390/d14070568