
Soil Arthropod Community in Spruce Forests (Picea abies) Affected by a Catastrophic Storm Event




Abstract
:1. Introduction
2. Materials and Methods
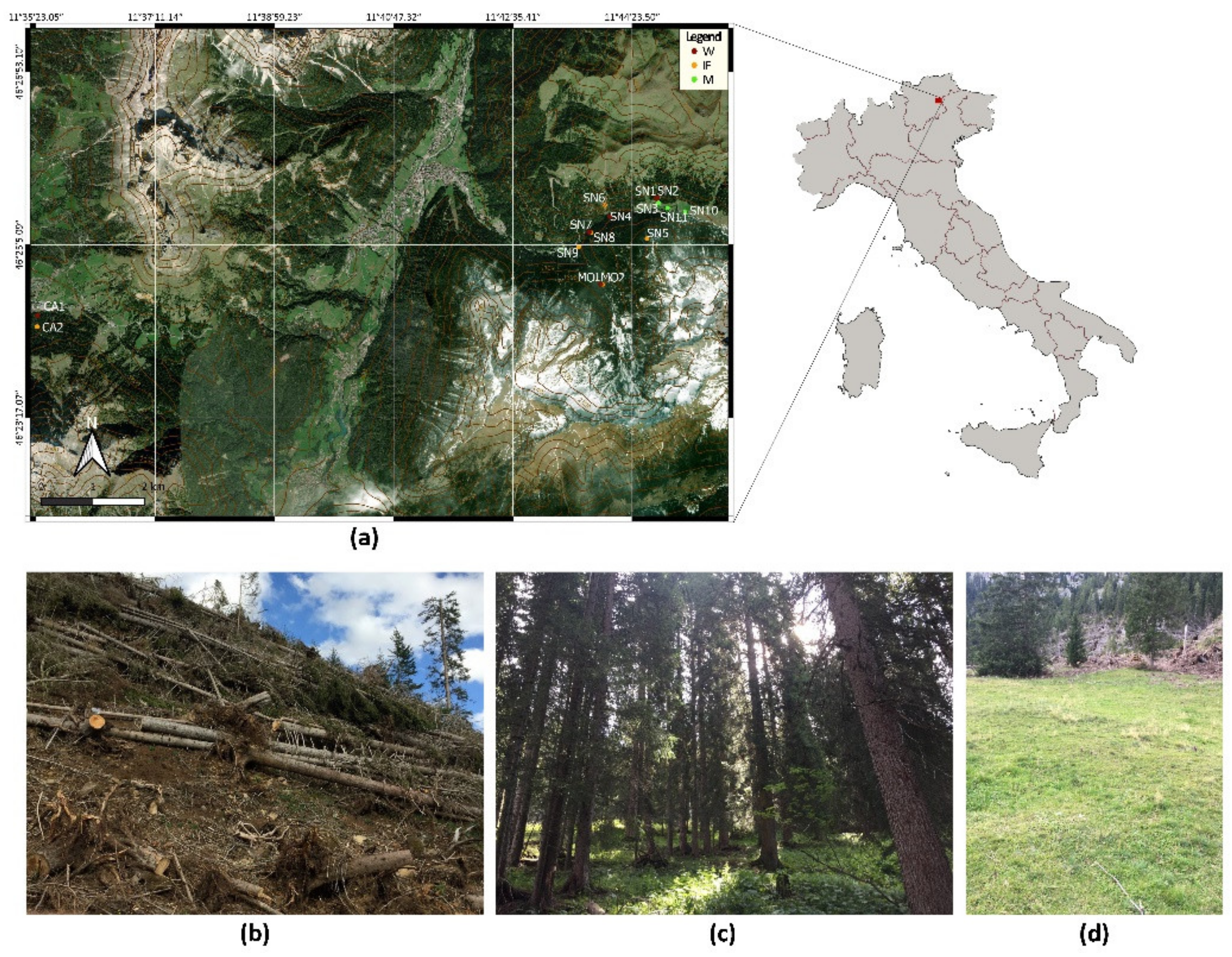
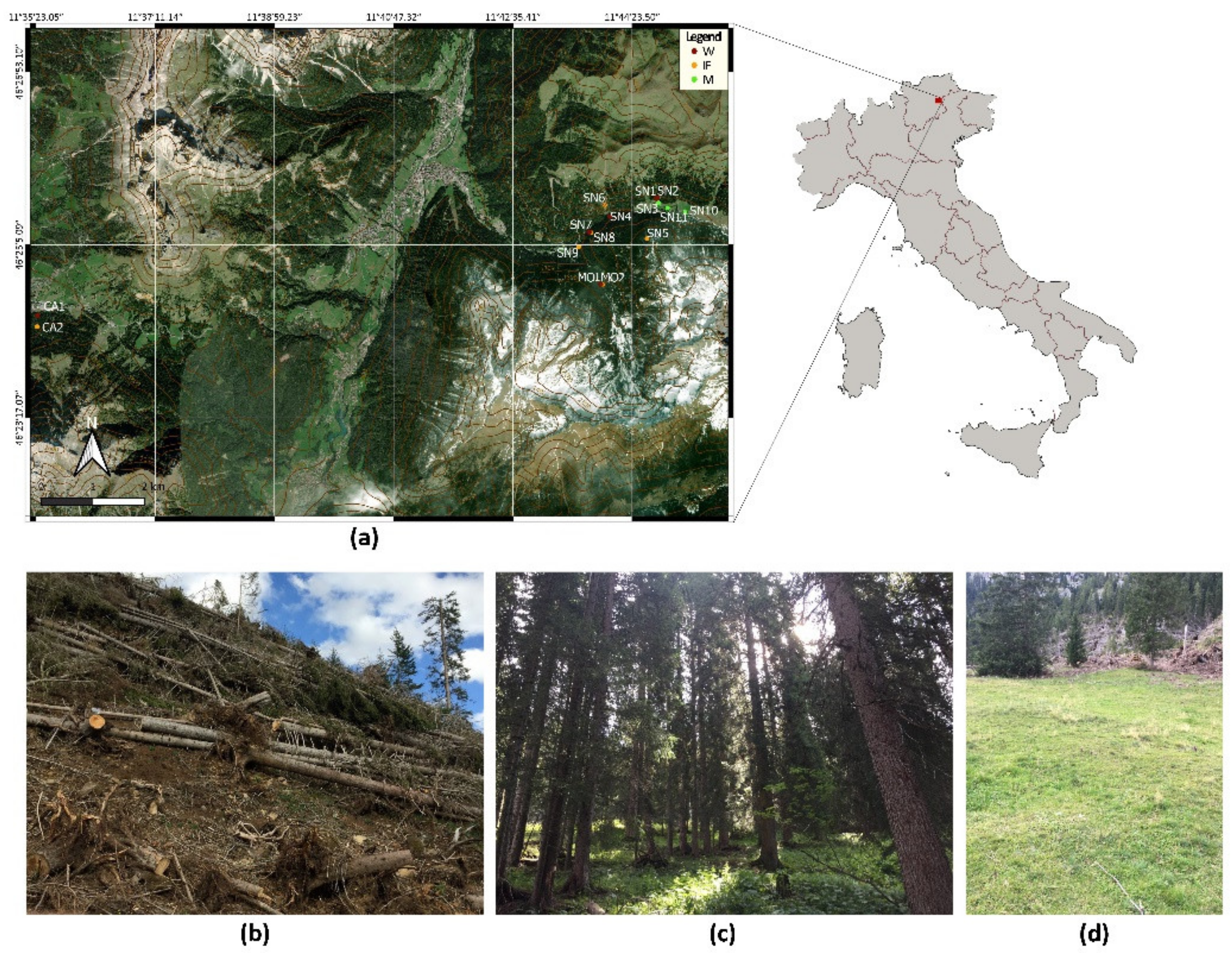
2.1. Study Area and Soil Collection
2.2. Chemical Analyses
2.3. Soil Arthropod Extraction
2.4. Statistical Analysis
3. Results
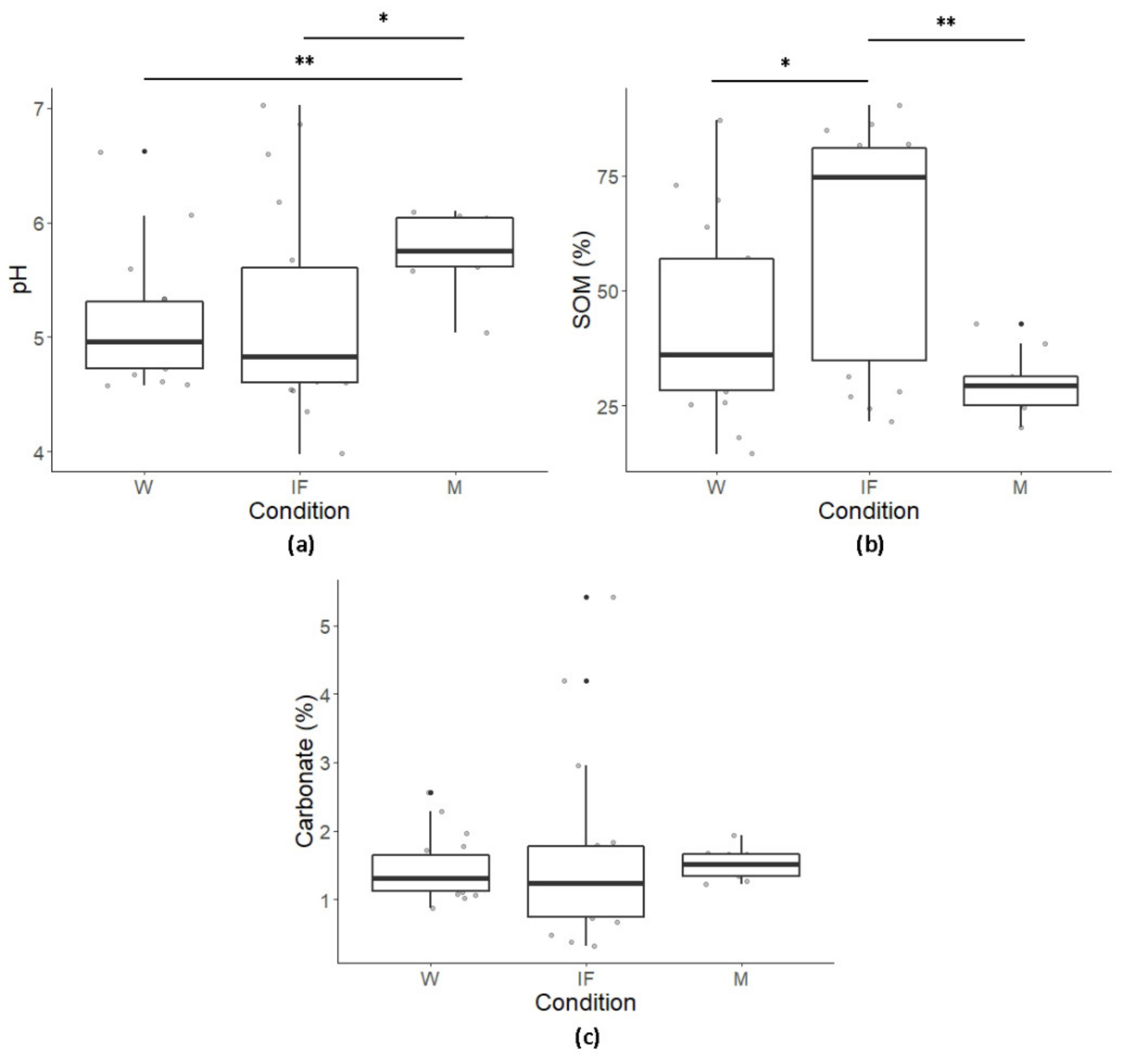
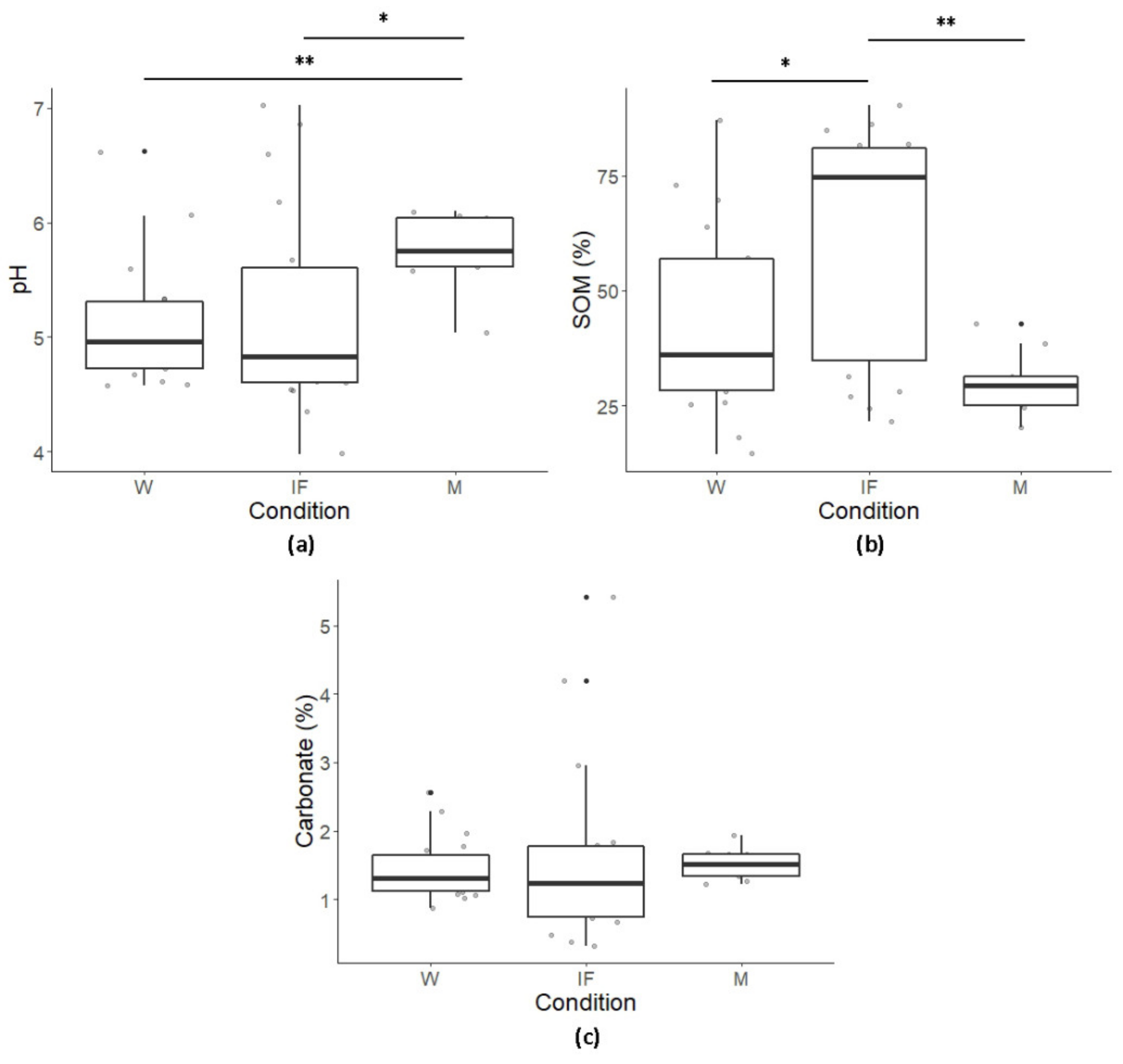
3.1. Chemical Parameters
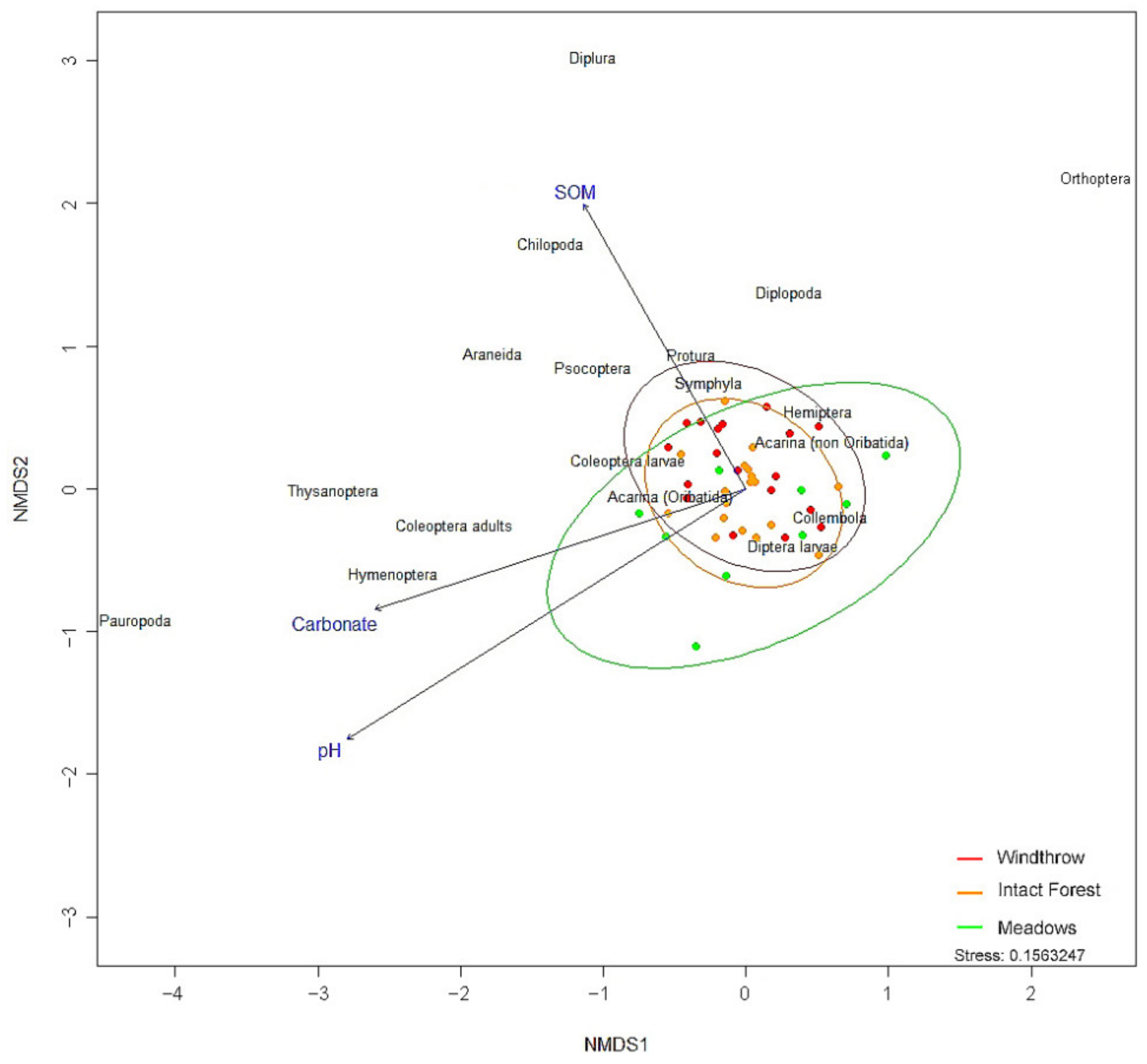
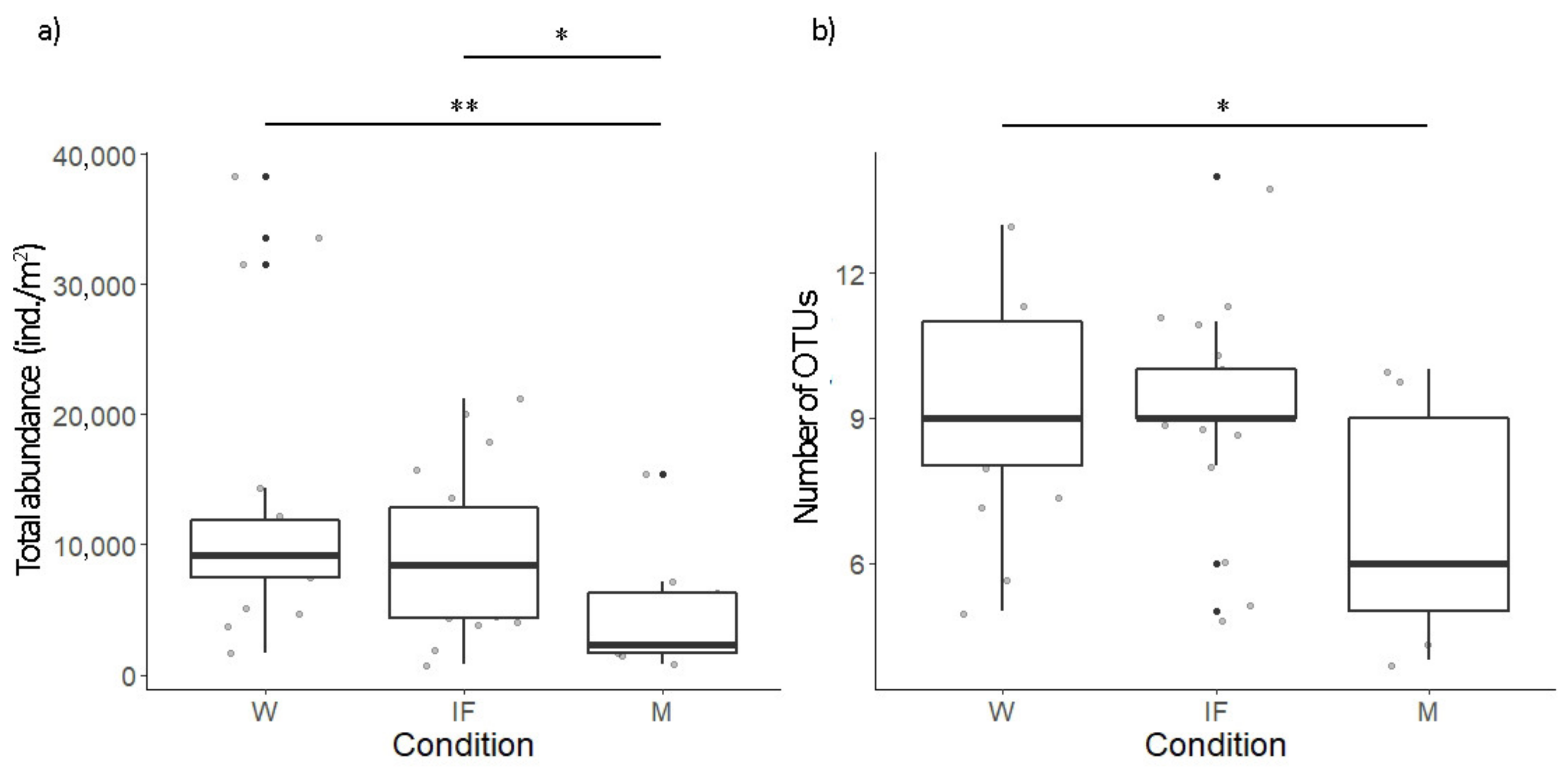
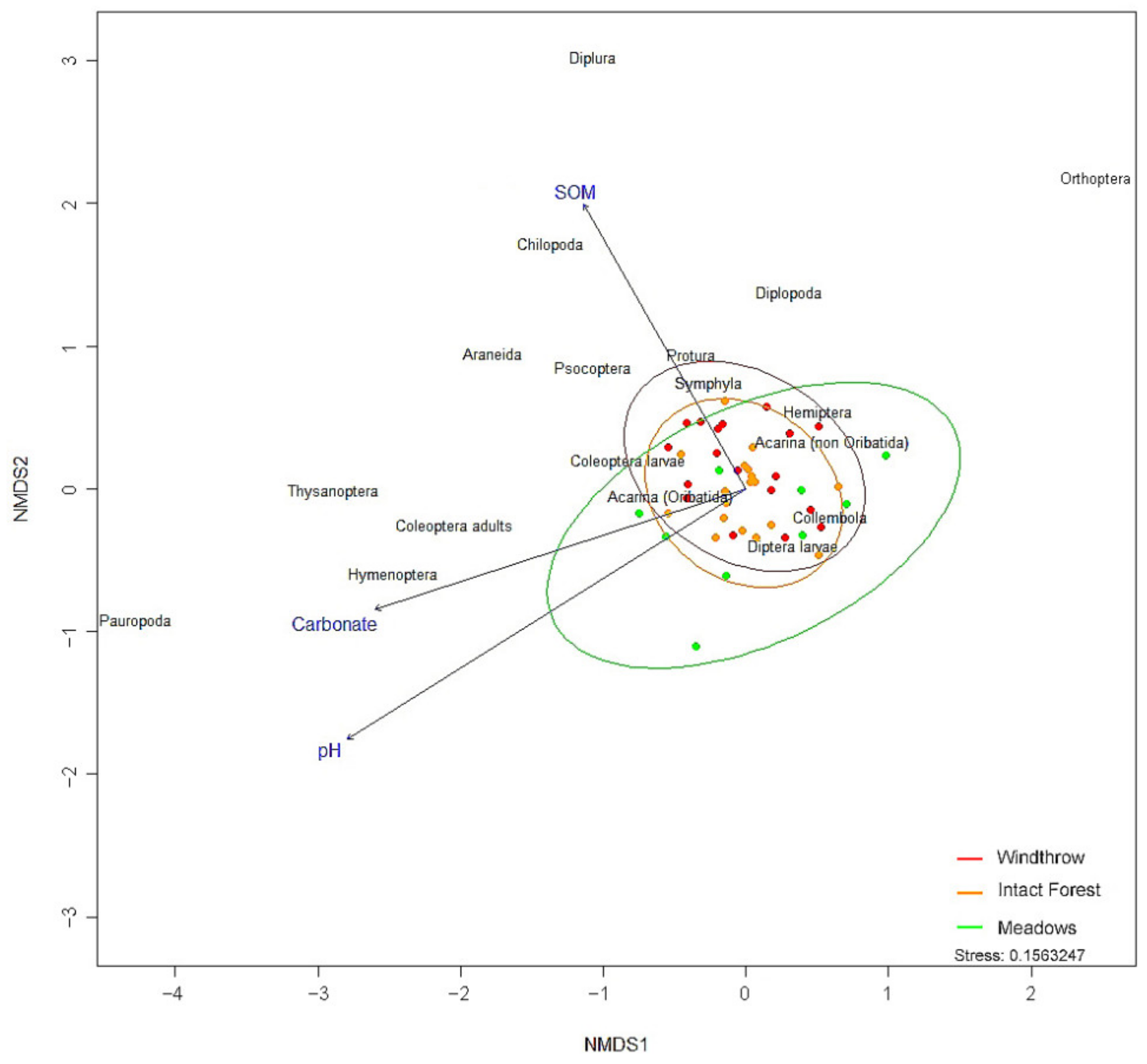
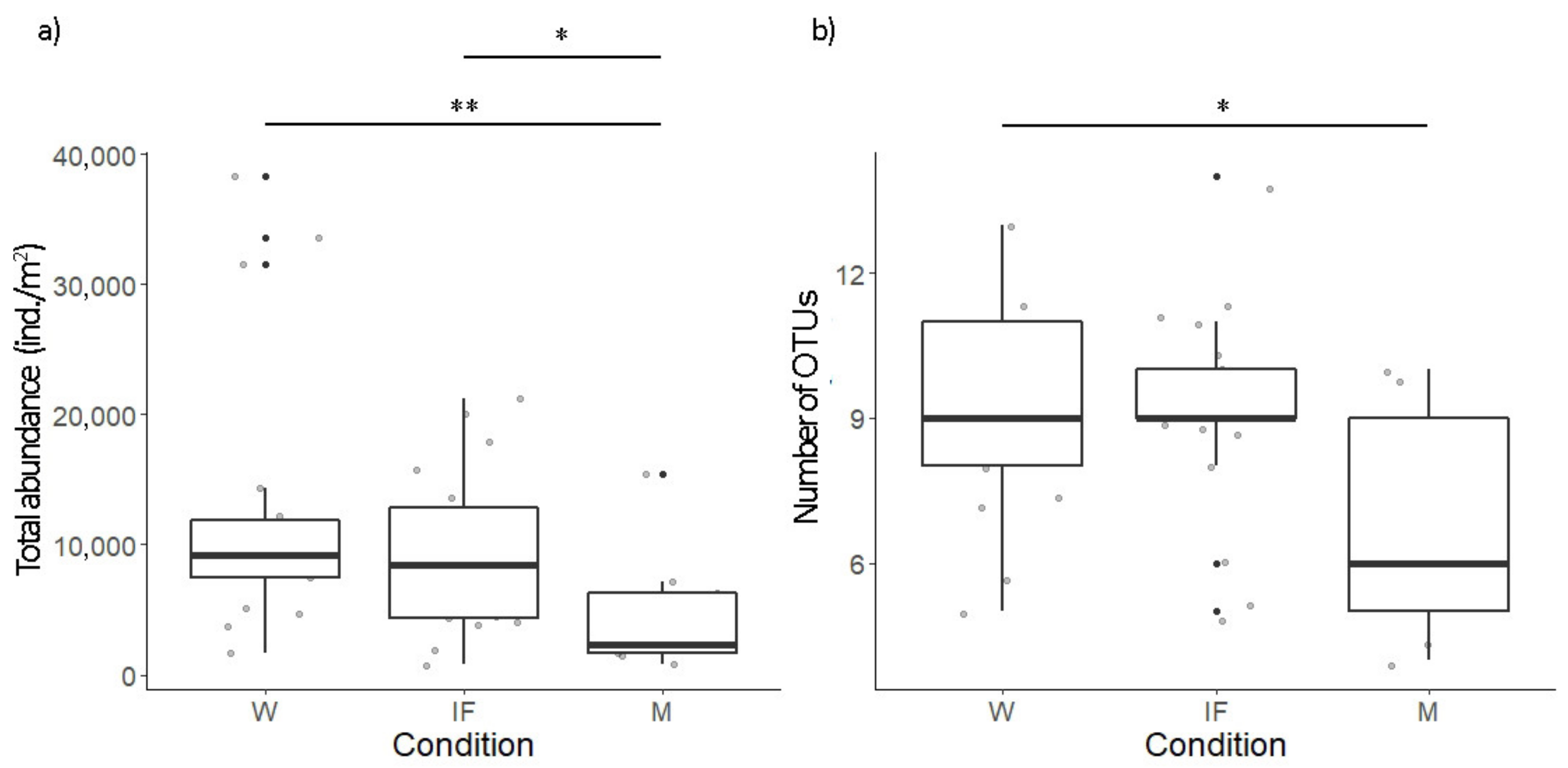
3.2. Soil Arthropods
3.2.1. Myriapoda
3.2.2. Chelicerata
3.2.3. Entognatha
3.2.4. Insecta
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coumou, D.; Rahmstorf, S. A decade of weather extremes. Nat. Clim. Chang. 2012, 2, 491–496. [Google Scholar] [CrossRef]
- IPCC. 2021 Summary for Policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2021. [Google Scholar]
- Matteucci, G.; Cammarano, M.; Dezi, S.; Mancini, M.; Mugnozza, G.S.; Magnani, F. Climate Change Impacts on Forests and Forest Products in the Mediterranean Area. In Advances in Global Change Research; Springer International Publishing: Cham, Switzerland, 2013; Volume 51, pp. 71–100. [Google Scholar] [CrossRef]
- Schelhaas, M.-J.; Nabuurs, G.-J.; Schuck, A. Natural disturbances in the European forests in the 19th and 20th centuries. Glob. Chang. Biol. 2003, 9, 1620–1633. [Google Scholar] [CrossRef]
- Seidl, R.; Schelhaas, M.-J.; Rammer, W.; Verkerk, P.J. Increasing forest disturbances in Europe and their impact on carbon storage. Nat. Clim. Chang. 2014, 4, 806–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franklin, J.F.; Spies, T.A.; Van Pelt, R.; Carey, A.B.; Thornburgh, D.A.; Berg, D.R.; Lindenmayer, D.B.; Harmon, M.E.; Keeton, W.S.; Shaw, D.C.; et al. Disturbances and structural development of natural forest ecosystems with silvicultural implications, using Douglas-fir forests as an example. For. Ecol. Manag. 2002, 155, 399–423. [Google Scholar] [CrossRef]
- Thom, D.; Seidl, R. Natural disturbance impacts on ecosystem services and biodiversity in temperate and boreal forests. Biol. Rev. 2016, 91, 760–781. [Google Scholar] [CrossRef]
- Graham, E.B.; Averill, C.; Bond-Lamberty, B.; Knelman, J.E.; Krause, S.; Peralta, A.L.; Shade, A.; Smith, A.P.; Cheng, S.J.; Fanin, N.; et al. Toward a Generalizable Framework of Disturbance Ecology through Crowdsourced Science. Front. Ecol. Evol. 2021, 9, 76. [Google Scholar] [CrossRef]
- IPCC. 2019 Summary for Policymakers. In Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; IPCC: Geneva, Switzerland, 2019. [Google Scholar]
- Lugo, A.E. Effects and outcomes of Caribbean hurricanes in a climate change scenario. Sci. Total Environ. 2000, 262, 243–251. [Google Scholar] [CrossRef]
- Shiels, A.B.; Gonzalez, G. Understanding the key mechanisms of tropical forest responses to canopy loss and biomass deposition from experimental hurricane effects. For. Ecol. Manag. 2014, 332, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kramer, M.G.; Sollins, P.; Sletten, R.S. Soil carbon dynamics across a windthrow disturbance sequence in southeast Alaska. Ecology 2004, 85, 2230–2244. [Google Scholar] [CrossRef] [Green Version]
- Coyle, D.R.; Nagendra, U.J.; Taylor, M.K.; Campbell, J.H.; Cunard, C.E.; Joslin, A.H.; Mundepi, A.; Phillips, C.A.; Callaham, M.A. Soil fauna responses to natural disturbances, invasive species, and global climate change: Current state of the science and a call to action. Soil Biol. Biochem. 2017, 110, 116–133. [Google Scholar] [CrossRef]
- Meehan, M.L.; Song, Z.; Lumley, L.; Cobb, T.P.; Proctor, H. Soil mites as bioindicators of disturbance in the boreal forest in northern Alberta, Canada: Testing taxonomic sufficiency at multiple taxonomic levels. Ecol. Indic. 2019, 102, 349–365. [Google Scholar] [CrossRef]
- Wermelinger, B.; Duelli, P.; Obrist, M.K. Windthrow Stimulates Arthropod Biodiversity in Forests. Dead wood: A key to biodiversity. In Proceedings of the International Symposium, Mantova, Italy, 29–31 May 2003; pp. 79–82. [Google Scholar]
- Duelli, P.; Obrist, M.K.; Wermelinger, B. Windthrow-Induced Changes in Faunistic Biodiversity in Alpine Spruce Forests. For. Snow Landsc. Res 2002, 77, 117–131. [Google Scholar]
- Doran, J.W.; Zeiss, M.R. Soil Health and Sustainability: Managing the Biotic Component of Soil Health and Sustainability: Managing the Biotic Component of Soil Quality Soil Quality Soil Health and Sustainability: Managing the Biotic Component of Soil Quality. Appl. Soil Ecol. 2000, 15, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Kibblewhite, M.G.; Ritz, K.; Swift, M.J. Soil Health in Agricultural Systems. Philos. Trans. R. Soc. Biol. Sci. 2008, 363, 685–701. [Google Scholar] [CrossRef] [Green Version]
- Wall, D.H.; Bradford, M.A.; John, M.G.S.; Trofymow, J.A.; Behan-Pelletier, V.; Bignell, D.E.; Dangerfield, J.M.; Parton, W.J.; Rusek, J.; Voigt, W.; et al. Global decomposition experiment shows soil animal impacts on decomposition are climate-dependent. Glob. Chang. Biol. 2008, 14, 2661–2677. [Google Scholar] [CrossRef] [Green Version]
- Soong, J.L.; Vandegehuchte, M.L.; Horton, A.J.; Nielsen, U.N.; Denef, K.; Shaw, E.A.; de Tomasel, C.M.; Parton, W.; Wall, D.H.; Cotrufo, M.F. Soil microarthropods support ecosystem productivity and soil C accrual: Evidence from a litter decomposition study in the tallgrass prairie. Soil Biol. Biochem. 2016, 92, 230–238. [Google Scholar] [CrossRef] [Green Version]
- Neher, D.A.; Barbercheck, M.E. Soil Microarthropods and Soil Health: Intersection of Decomposition and Pest Suppression in Agroecosystems. Insects 2019, 10, 414. [Google Scholar] [CrossRef] [Green Version]
- Lakshmi, G.; Beggi, F.; Menta, C.; Kumar, N.K.; Jayesh, P. Dynamics of soil microarthropod populations affected by a combination of extreme climatic events in tropical home gardens of Kerala, India. Pedobiologia 2021, 85–86, 150719. [Google Scholar] [CrossRef]
- Farská, J.; Prejzková, K.; Rusek, J. Management intensity affects traits of soil microarthropod community in montane spruce forest. Appl. Soil Ecol. 2014, 75, 71–79. [Google Scholar] [CrossRef]
- Johnstone, J.; Chapin, F.S. Effects of Soil Burn Severity on Post-Fire Tree Recruitment in Boreal Forest. Ecosystems 2006, 9, 14–31. [Google Scholar] [CrossRef]
- Čuchta, P.; Miklisová, D.; Kováč, Ľ. A three-year study of soil Collembola communities in spruce forest stands of the High Tatra Mts (Slovakia) after a catastrophic windthrow event. Eur. J. Soil Biol. 2012, 50, 151–158. [Google Scholar] [CrossRef]
- Zanella, A.; Ponge, J.-F.; Andreetta, A.; Aubert, M.; Bernier, N.; Bonifacio, E.; Bonneval, K.; Bolzonella, C.; Chertov, O.; Costantini, E.A.C.; et al. Combined forest and soil management after a catastrophic event. J. Mt. Sci. 2020, 17, 2459–2484. [Google Scholar] [CrossRef] [PubMed]
- Chirici, C.; Giannetti, G.; Travaglini, T.; Nocentini, N.; Francini, F.; D’Amico, D.; Calvo, C.; Fasolini, F.; Broll, B.; Maistrelli, M.; et al. Stima Dei Danni Della Tempesta “Vaia” Alle Foreste in Italia. For. J. Silvic. For. Ecol. 2019, 16, 3. [Google Scholar] [CrossRef] [Green Version]
- QGIS Development Team. QGIS Geographic Information System. Open Source Geospatial Foundation. Map Tiles by Google Satellite. Available online: http://qgis.org (accessed on 17 November 2021).
- Klironomos, J.N.; Rillig, M.; Allen, M.F. Designing belowground field experiments with the help of semi-variance and power analyses. Appl. Soil Ecol. 1999, 12, 227–238. [Google Scholar] [CrossRef]
- S.I.S.S. Metodi Normalizzati di Analisi del Suolo; Edagricole: Bologna, Italy, 1986. [Google Scholar]
- Heiri, O.; Lotter, A.F.; Lemcke, G. Loss on ignition as a method for estimating organic and carbonate content in sediments: Reproducibility and comparability of results. J. Paleolimnol. 2001, 25, 101–110. [Google Scholar] [CrossRef]
- John, F.; Weisberg, S.; Price, B. An {R} Companion to Applied Regression; Sage: New York, NY, USA, 2019; Volume 58. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package “vegan”. Title Community Ecol. Package Version 2.5-7 2020, 9, 295. [Google Scholar]
- Martinez Arbizu, P. PairwiseAdonis: Pairwise Multilevel Comparison Using Adonis. Available online: https://github.com/pmartinezarbizu/pairwiseAdonis/tree/master/pairwiseAdonis (accessed on 23 September 2020).
- De Cáceres, M.; Jansen, F.; Noah, D. Relationship between Species and Groups of Sites. Package ‘Indicspecies’. Available online: https://cran.r-project.org/web/packages/indicspecies/indicspecies.pdf (accessed on 6 March 2022).
- Pagès, J. Analyse Factorielle de Données Mixtes; Société Française De Statistique: Paris, France, 2004; Volume 52. [Google Scholar]
- Husson, F.; Josse, J.; Le, S.; Maintainer, J.M. Multivariate Exploratory Data Analysis and Data Mining. Available online: https://cran.r-project.org/web/packages/FactoMineR/FactoMineR.pdf (accessed on 12 December 2020).
- Kassambara, A.; Mundt, F. Extract and Visualize the Results of Multivariate Data Analyses. Available online: https://cran.r-project.org/web/packages/factoextra/factoextra.pdf (accessed on 13 April 2020).
- Wood, S.N. Generalized Additive Models: An Introduction with R, 2nd ed.; Chapman and Hall/CR: London, UK, 2017; ISBN 9781498728348. [Google Scholar]
- Marra, G.; Wood, S.N. Coverage Properties of Confidence Intervals for Generalized Additive Model Components. Scand. J. Stat. 2012, 39, 53–74. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing 2021; Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Jobbágy, E.G.; Jackson, R.B. Patterns and mechanisms of soil acidification in the conversion of grasslands to forests. Biogeochemistry 2003, 64, 205–229. [Google Scholar] [CrossRef]
- Brandtberg, P.-O.; Lundkvist, H.; Bengtsson, J. Changes in forest-floor chemistry caused by a birch admixture in Norway spruce stands. For. Ecol. Manag. 2000, 130, 253–264. [Google Scholar] [CrossRef]
- Binkley, D.; Valentine, D. Fifty-year biogeochemical effects of green ash, white pine, and Norway spruce in a replicated experiment. For. Ecol. Manag. 1991, 40, 13–25. [Google Scholar] [CrossRef]
- Binkley, D.; Giardina, C. Why do tree species affect soils? The Warp and Woof of tree-soil interactions. Biogeochemistry 1998, 42, 89–106. [Google Scholar] [CrossRef]
- Lóšková, J.; Ľuptáčik, P.; Miklisová, D.; Kováč, Ľ. Community structure of soil oribatida (acari) two years after windthrow in the high tatra mountains. Biologia 2013, 68, 932–940. [Google Scholar] [CrossRef]
- Mayer, M.; Sandén, H.; Rewald, B.; Godbold, D.L.; Katzensteiner, K. Increase in heterotrophic soil respiration by temperature drives decline in soil organic carbon stocks after forest windthrow in a mountainous ecosystem. Funct. Ecol. 2017, 31, 1163–1172. [Google Scholar] [CrossRef]
- Tonon, G.; Sohi, S.; Francioso, O.; Ferrari, E.; Montecchio, D.; Gioacchini, P.; Ciavatta, C.; Panzacchi, P.; Powlson, D. Effect of soil pH on the chemical composition of organic matter in physically separated soil fractions in two broadleaf woodland sites at Rothamsted, UK. Eur. J. Soil Sci. 2010, 61, 970–979. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Hobbs, P.J.; Frostegård, Å. Changes in soil fungal:bacterial biomass ratios following reductions in the intensity of management of an upland grassland. Biol. Fertil. Soils 1996, 22, 261–264. [Google Scholar] [CrossRef]
- Kramer, M.G. Maritime Windstorm Influence on Soil Process in a Temperate Rainforest; Oregon State University: Corvallis, OR, USA, 2001. [Google Scholar]
- Schaetzl, R.J. Complete soil profile inversion by tree uprooting. Phys. Geogr. 1986, 7, 181–189. [Google Scholar] [CrossRef]
- Tolimir, M.; Kresović, B.; Životić, L.; Dragović, S.; Dragović, R.; Sredojević, Z.; Gajić, B. The conversion of forestland into agricultural land without appropriate measures to conserve SOM leads to the degradation of physical and rheological soil properties. Sci. Rep. 2020, 10, 13668. [Google Scholar] [CrossRef]
- Batjes, N.H. Total carbon and nitrogen in the soils of the world. Eur. J. Soil Sci. 1996, 47, 151–163. [Google Scholar] [CrossRef]
- Wang, X.; Wang, J.; Xu, M.; Zhang, W.; Fan, T.; Zhang, J. Carbon accumulation in arid croplands of northwest China: Pedogenic carbonate exceeding organic carbon. Sci. Rep. 2015, 5, 11439. [Google Scholar] [CrossRef] [Green Version]
- Lal, R.; Kimble, J.M.; Stewart, B.A.; Eswaran, H. Global Climate Change and Pedogenic Carbonates; U.S. Department of Energy: Washington, DC, USA, 1999.
- Ananthakrishnan, T.N. Biosystematics of Thysanoptera. Annu. Rev. Èntomol. 1979, 24, 159–183. [Google Scholar] [CrossRef]
- Berthet, P. Mesure de La Consummation d’oxygene Des Oribates (Acariens) de La Litieredesforêts. In Soil Organisms; Doeksen, J., van der Drift, J., Eds.; North-Holland Publishing Co: Amsterdam, The Netherlands, 1963; pp. 18–31. [Google Scholar]
- Wehner, K.; Simons, N.K.; Blüthgen, N.; Heethoff, M. Drought, windthrow and forest operations strongly affect oribatid mite communities in different microhabitats. Glob. Ecol. Conserv. 2021, 30, e01757. [Google Scholar] [CrossRef]
- Kazeev, K.S.; Poltoratskaya, T.A.; Yakimova, A.S.; Odobashyan, M.Y.; Shkhapatsev, A.K.; Kolesnikov, S.I. Post-Fire Changes in the Biological Properties of the Brown Soils in the Utrish State Nature Reserve (Russia). Nat. Conserv. Res. 2019, 4, 93–104. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Egorov, L.V.; MacGowan, I.; Makarkin, V.N.; Antropov, A.V.; Gornostaev, N.G.; Khapugin, A.A.; Dvořák, L.; Esin, M.N. Post-fire insect fauna explored by crown fermental traps in forests of the European Russia. Sci. Rep. 2021, 11, 21334. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, K.J.; Spence, J.R.; Langor, D.W.; E Morgantini, L. Fire residuals as habitat reserves for epigaeic beetles (Coleoptera: Carabidae and Staphylinidae). Biol. Conserv. 2001, 102, 131–141. [Google Scholar] [CrossRef]
- Perry, K.I.; Herms, D.A. Dynamic Responses of Ground-Dwelling Invertebrate Communities to Disturbance in Forest Ecosystems. Insects 2016, 10, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atchison, R.A.; Hulcr, J.; Lucky, A. Managed Fire Frequency Significantly Influences the Litter Arthropod Community in Longleaf Pine Flatwoods. Environ. EÈntomol. 2018, 47, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Jabin, M.; Topp, W.; Kulfan, J.; Zach, P. The distribution pattern of centipedes in four primeval forests of central Slovakia. Biodivers. Conserv. 2007, 16, 3437–3445. [Google Scholar] [CrossRef]
- Topp, W. Seasonal time partitioning and polymorphism in the developmental cycles of sympatric Staphylinoidea (Coleoptera) living in an unstable environment. In Insect Life-Cycle Polymorph; Springer: Dordrecht, The Netherlands, 1994; pp. 277–312. [Google Scholar] [CrossRef]
- Ruchin, A.B.; “Smolny”, J.D.O.T.M.S.N.R.A.N.P.; Alekseev, S.K.; Khapugin, A.; Ecological club «Stenus». Tyumen State University Post-fire fauna of carabid beetles (Coleoptera, Carabidae) in forests of the Mordovia State Nature Reserve (Russia). Nat. Conserv. Res. 2019, 4, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Čuchta, P.; Kaňa, J.; Pouska, V. An important role of decomposing wood for soil environment with a reference to communities of springtails (Collembola). Environ. Monit. Assess. 2019, 191, 222. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| W | IF | M | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| OTUs | % | Mean | ± | St.err. | % | Mean | ± | St.err. | % | Mean | ± | St.err. |
| Hexapoda Entognatha | ||||||||||||
| Collembola | 36.81 | 4611.63 | ± | 1610.39a | 22.78 | 2097.05 | ± | 520.18a | 67.93 | 3078.35 | ± | 1532.93a |
| Protura | 0.14 | 17.69 | ± | 7.33a | 0.35 | 31.84 | ± | 18.18a | 0.10 | 4.72 | ± | 3.12a |
| Hexapoda Insecta | ||||||||||||
| Coleoptera adults | 0.31 | 38.92 | ± | 11.29a | 0.65 | 60.15 | ± | 10.47a | 1.04 | 47.18 | ± | 19.31a |
| Coleoptera larvae | 0.39 | 49.54 | ± | 9.56b | 1.56 | 143.89 | ± | 28.59a | 0.68 | 30.67 | ± | 11.79b |
| Diplura | - | - | a | 0.01 | 1.18 | ± | 1.18a | - | - | a | ||
| Diptera larvae | 0.26 | 33.02 | ± | 8.95a | 0.90 | 82.56 | ± | 19.33a | 0.57 | 25.95 | ± | 9.21a |
| Hemiptera | 0.83 | 103.79 | ± | 65.56a | 6.67 | 614.49 | ± | 450.13a | 0.62 | 28.31 | ± | 16.21a |
| Hymenoptera | 0.02 | 2.36 | ± | 2.36b | 0.26 | 23.59 | ± | 12.95a | 0.68 | 30.67 | ± | 30.66ab |
| Orthoptera | 0.01 | 1.18 | ± | 1.18a | - | - | a | - | - | a | ||
| Psocoptera | 0.07 | 9.44 | ± | 8.27a | 0.06 | 5.90 | ± | 3.35a | 0.05 | 2.36 | ± | 2.36a |
| Thysanoptera | 0.07 | 9.44 | ± | 4.92a | 0.04 | 3.54 | ± | 1.92a | 1.20 | 54.25 | ± | 31.87a |
| Arachnida | ||||||||||||
| Acarina (non Oribatida) | 27.88 | 3492.33 | ± | 1058.89a | 27.52 | 2533.45 | ± | 463.97a | 14.37 | 651.05 | ± | 314.51b |
| Acarina (Oribatida) | 31.77 | 3980.62 | ± | 739.83a | 38.23 | 3519.46 | ± | 705.94a | 11.76 | 533.11 | ± | 297.17b |
| Araneae | 0.42 | 53.07 | ± | 18.02a | 0.23 | 21.23 | ± | 12.96a | 0.47 | 21.23 | ± | 18.72a |
| Myriapoda | ||||||||||||
| Chilopoda | 0.40 | 50.72 | ± | 13.96a | 0.18 | 16.51 | ± | 6.55b | - | - | c | |
| Diplopoda | 0.44 | 55.44 | ± | 26.76a | 0.35 | 31.84 | ± | 14.07a | 0.31 | 14.15 | ± | 10.01a |
| Pauropoda | - | - | a | - | - | a | 0.05 | 2.36 | ± | 2.36a | ||
| Symphyla | 0.15 | 18.87 | ± | 6.40a | 0.22 | 20.05 | ± | 7.17a | 0.16 | 7.08 | ± | 3.54a |
| Diversity indices | ||||||||||||
| H | 1.19 | ± | 0.06a | 1.26 | ± | 0.06a | 0.97 | ± | 0.17a | |||
| 1-D | 0.62 | ± | 0.02a | 0.63 | ± | 0.02a | 0.48 | ± | 0.09a | |||
| Dependent Variable | Parametric Coefficients | Smooth Terms | R2 Adjusted | Deviance Explained | ||
|---|---|---|---|---|---|---|
| Terms | Estimate | Terms | edf | |||
| Abundance | Condition|W | 5.25 * | SOM:Condition|IF | 1.94 * | 0.47 | 64.5% |
| Carbonate | −3.08 | SOM:Condition|W | 4.64 ** | |||
| Condition|IF:pH | 0.06 | |||||
| Condition|W:pH | −0.90 * | |||||
| N° of OTUs | Condition|W | 0.05 | Carbonate | 7.37 | 0.31 | 51.4% |
| SOM | 0.95 ** | |||||
| pH | 0.08 | |||||
| Acarina (Oribatida) | Condition|W | −0.23 | SOM | 3.62 | 0.52 | 62.4% |
| Carbonate | −47.82 * | pH: Condition|IF | 2.35 * | |||
| pH: Condition|W | 3.42 | |||||
| Araneae | Condition|W | 3.58 ** | Carbonate | 2.38 | 0.68 | 71.8% |
| SOM | 8.57 | |||||
| pH | 3.94 | |||||
| Chilopoda | Condition|W | −2.59 | SOM:Condition|IF | 2.83 | 0.83 | 75.4% |
| Carbonate | −164.63 | SOM:Condition|W | 7.55 * | |||
| pH | 1.04 | |||||
| Condition|W:Carbonate | 337.81 ** | |||||
| Coleoptera larvae | Condition|W | −0.78 * | SOM | 7.39 | 0.66 | 71.7% |
| Carbonate | −27.98 | pH | 3.89 * | |||
| Diptera larvae | Condition|W | −1.11 * | Carbonate | 4.62 | 0.66 | 65.9% |
| SOM | 1.33 | |||||
| pH | 6.63 * | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menta, C.; Lozano Fondón, C.; Remelli, S. Soil Arthropod Community in Spruce Forests (Picea abies) Affected by a Catastrophic Storm Event. Diversity 2022, 14, 440. https://doi.org/10.3390/d14060440
Menta C, Lozano Fondón C, Remelli S. Soil Arthropod Community in Spruce Forests (Picea abies) Affected by a Catastrophic Storm Event. Diversity. 2022; 14(6):440. https://doi.org/10.3390/d14060440
Chicago/Turabian StyleMenta, Cristina, Carlos Lozano Fondón, and Sara Remelli. 2022. "Soil Arthropod Community in Spruce Forests (Picea abies) Affected by a Catastrophic Storm Event" Diversity 14, no. 6: 440. https://doi.org/10.3390/d14060440
APA StyleMenta, C., Lozano Fondón, C., & Remelli, S. (2022). Soil Arthropod Community in Spruce Forests (Picea abies) Affected by a Catastrophic Storm Event. Diversity, 14(6), 440. https://doi.org/10.3390/d14060440