
Importance of Mesohabitat for Nest-Site Selection in Breeding Eagle Owls (Bubo bubo): A Multi-Scale Model




Abstract
:1. Introduction
2. Materials and Methods
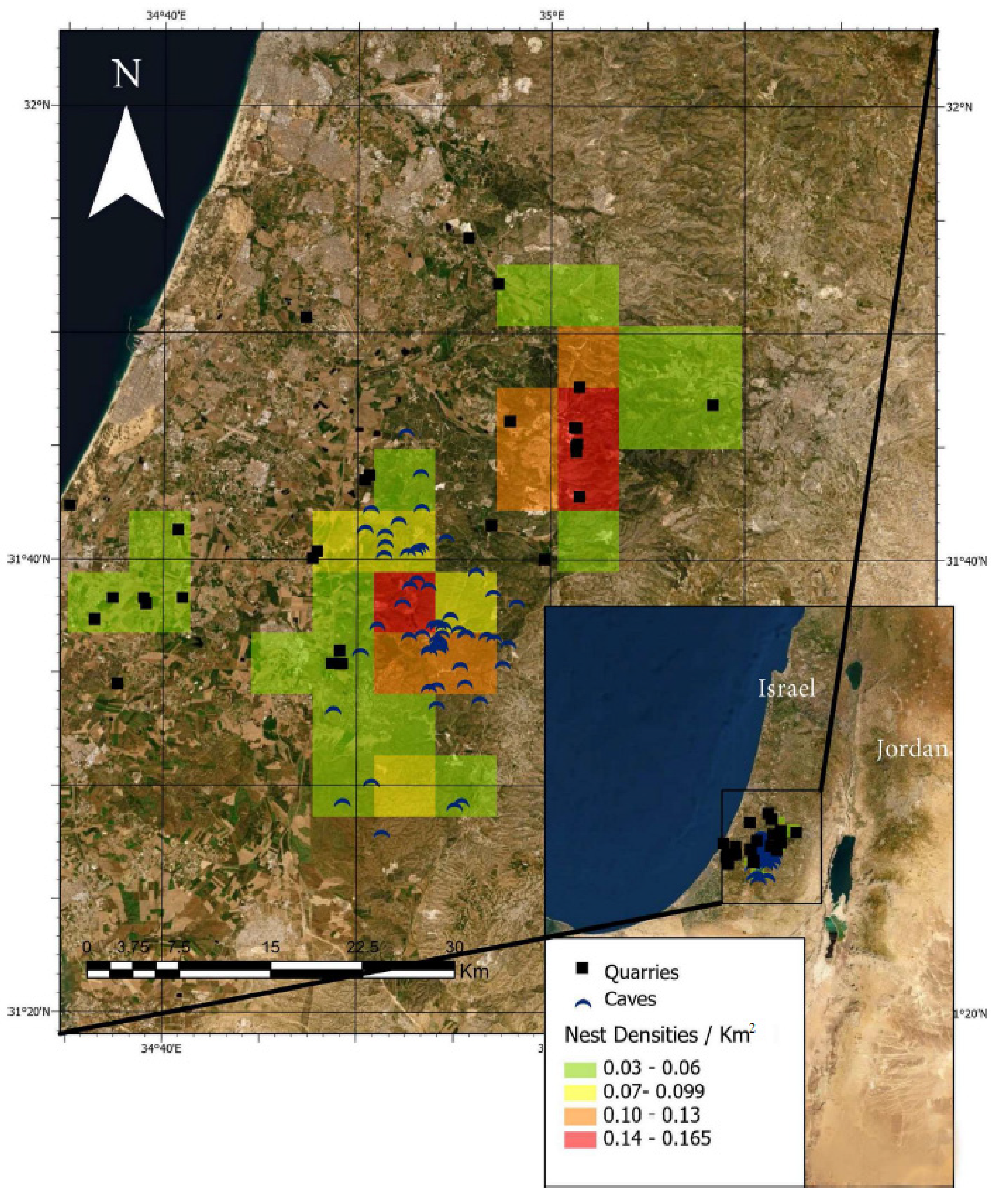
2.1. Study Area
2.2. Statistical Analysis
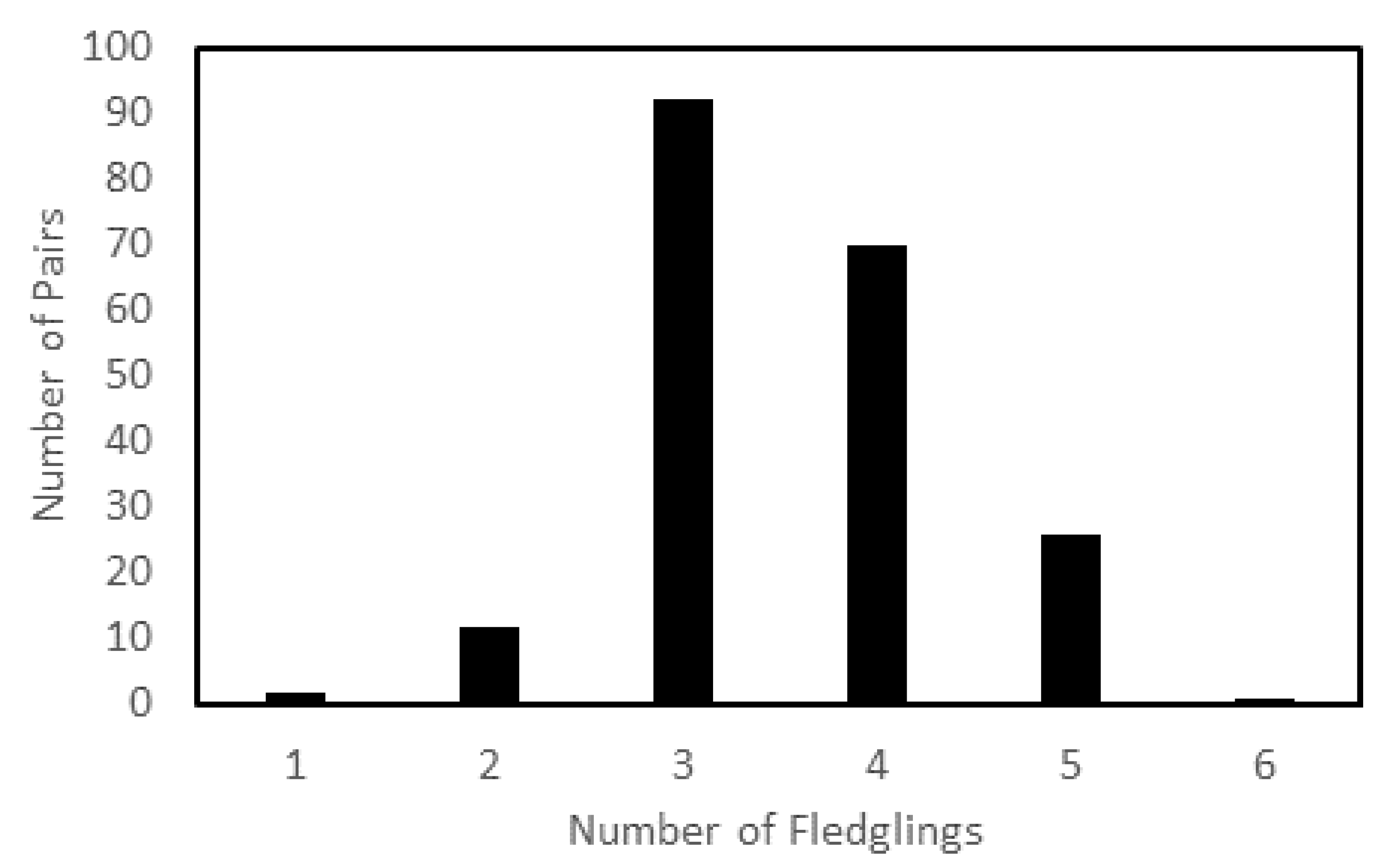
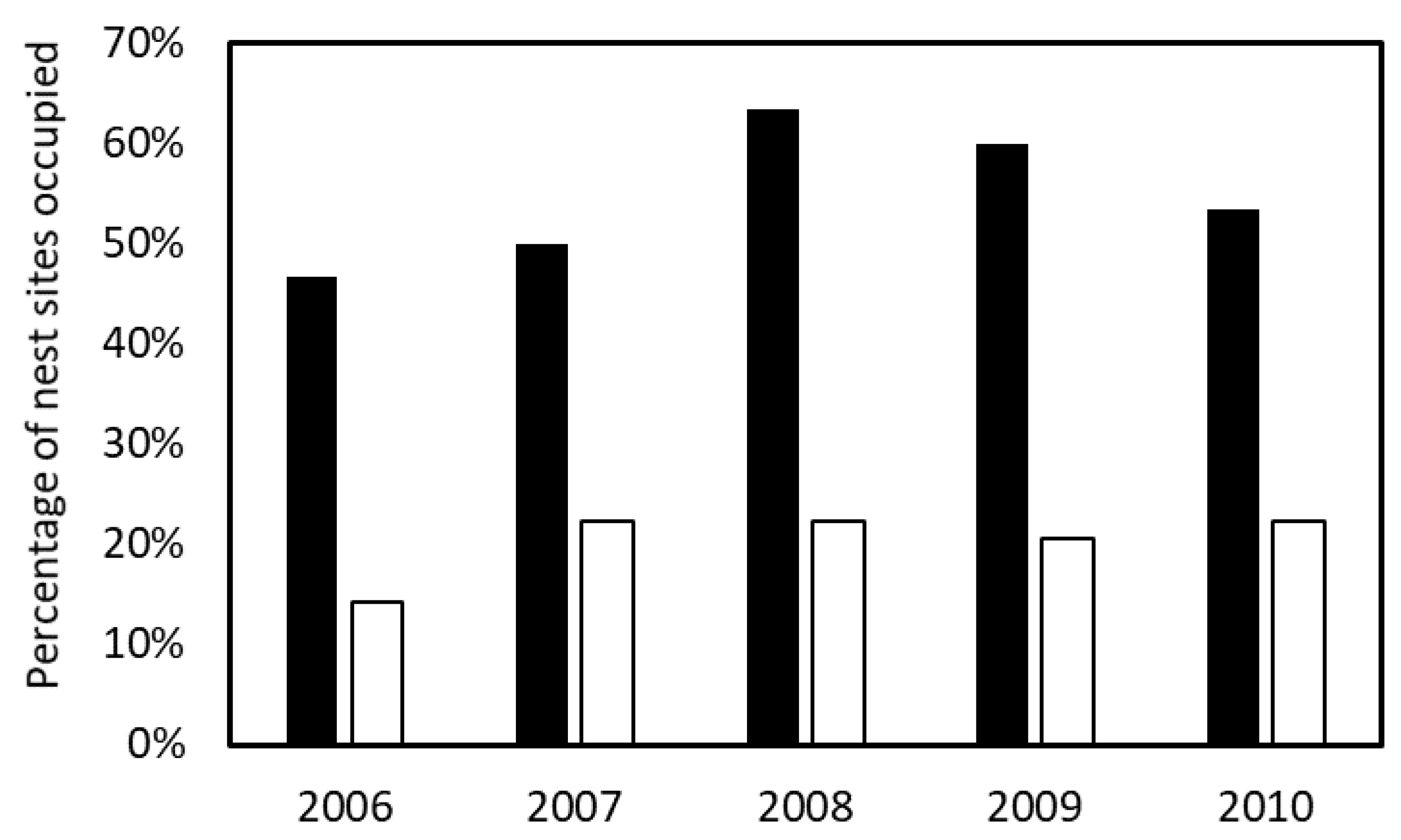
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brambilla, M.; Rubolini, D.; Guidali, F. Eagle Owl Bubo bubo proximity can lower productivity of cliff-nesting Peregrines Falco peregrinus. Ornis Fenn. 2006, 83, 20–26. [Google Scholar]
- Fahrig, L. Non-optimal animal movement in human-altered landscapes. Funct. Ecol. 2007, 21, 1003–1015. [Google Scholar] [CrossRef]
- McConkey, K.R.; O’Farrill, G. Cryptic function loss in animal populations. Trends Ecol. Evol. 2015, 30, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Newton, I. Population Limitation in Birds; Elsevier: Amsterdam, The Netherlands, 1998. [Google Scholar]
- Newton, I. Population limitation in birds: The last 100 years. Br. Birds 2007, 100, 518–539. [Google Scholar]
- Penteriani, V.; Gallardo, M.; Roche, P. Landscape structure and food supply affect eagle owl (Bubo bubo) density and breeding performance: A case of intra-population heterogeneity. J. Zool. 2002, 257, 365–372. [Google Scholar] [CrossRef] [Green Version]
- Martínez, J.A.; Serrano, D.; Zuberogoitia, I. Predictive models of habitat preferences for the Eurasian eagle owl Bubo bubo: A multiscale approach. Ecography 2003, 26, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Ortego, J.; Díaz, M. Habitat preference models for nesting eagle owls Bubo bubo: How much can be interred from changes with spatial scale? Ardeloa 2004, 51, 385–394. [Google Scholar]
- Donázar, J.A. Seleccion del habitat de nidificacion por el buho real (Bubo bubo) en Navarra. Ardeola 1988, 35, 233–245, (In Spanish, English Abstract). [Google Scholar]
- Lourenço, R.; Santos, S.; Rabaça, J.E.; Penteriani, V. Superpredation patterns in four large European raptors. Popul. Ecol. 2011, 53, 175–185. [Google Scholar] [CrossRef] [Green Version]
- Sergio, F.; Marchesi, L.; Pedrini, P. Integrating individual habitat choices and regional distribution of a biodiversity indicator and top predator. J. Biogeogr. 2004, 31, 619–628. [Google Scholar] [CrossRef]
- Bionda, R.; Brambilla, M. Rainfall and landscape features affect productivity in an alpine population of Eagle Owl Bubo bubo. J. Ornithol. 2012, 153, 167–171. [Google Scholar] [CrossRef]
- Donázar, J.A. Geographic variation in clutch and brood size of the Eagle Owl Bubo bubo in the Western Palearctic. J. Ornithol. 1990, 131, 439–443. [Google Scholar] [CrossRef]
- Dalbeck, L.; Heg, D. Reproductive success of a reintroduced population of Eagle Owls Bubo bubo in relation to habitat characteristics in the Eifel, Germany. Ardea 2006, 94, 3–21. [Google Scholar]
- Sergio, F.; Newton, I.P.; Marchesi, L. Top predators and biodiversity. Nature 2005, 436, 192. [Google Scholar] [CrossRef]
- Sergio, F.; Marchesi, L.; Pedrini, P.; Penteriani, V. Coexistence of a generalist owl with its intraguild predator: Distance-sensitive or habitat-mediated avoidance? Anim. Behav. 2007, 74, 1607–1616. [Google Scholar] [CrossRef] [Green Version]
- Tumurbat, J.; Sundev, G.; Yosef, R. Nest Site and Food Composition of the Eagle Owl Bubo bubo in Mongolia. Ardea 2009, 97, 519–523. [Google Scholar] [CrossRef]
- Pande, S.; Pawashe, A.; Mahajan, M.; Mahabal, A.; Joglekar, C.; Yosef, R. Breeding Biology, Nesting Habitat, and Diet of the Rock Eagle-Owl (Bubo bengalensis). J. Raptor Res. 2011, 45, 211–219. [Google Scholar] [CrossRef]
- Penteriani, V.; del Mar Delgado, M. The Eagle Owl; T & AD Poyser Ltd.: London, UK, 2019. [Google Scholar]
- Clucas, B.; McHugh, K.; Caro, T. Flagship species on covers of US conservation and nature magazines. Biodivers. Conserv. 2008, 17, 1517–1528. [Google Scholar] [CrossRef]
- Fischhendler, I.; Frumkin, A. Distribution, evolution, and morphology of caves in southwestern Samaria, Israel. Isr. J. Earth Sci. 2008, 57, 311–322. [Google Scholar] [CrossRef]
- Zissu, B.; Kloner, A. The bell-shaped quarries of the Judean foothills, Israel. Opera Ipogea 2014, 2, 47–60. [Google Scholar]
- Ratti, J.T.; Garton, E.O. Research and experimental design. In Research and Management Techniques for Wildlife and Habitats; Bookhout, T.A., Ed.; The Wildlife Society: Lawrence, Kansas, 1994; pp. 1–23. [Google Scholar]
- Kozak, M. Optimal stratification using random search methods in agricultural surveys. Stat. Transit. 2004, 6, 797–806. [Google Scholar]
- Andersen, D.E. Survey techniques. In Raptor Research and Management Techniques; Bird, D.M., Bildstein, K.L., Eds.; Raptor Research Foundation: Washington, DC, USA, 2007. [Google Scholar]
- Penteriani, V.; Delgado, M.D.M.; Campioni, L. Quantifying space use of breeders and floaters of a long-lived species using individual movement data. Sci. Nat. 2015, 102, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleming, C.H.; Fagan, W.F.; Mueller, T.; Olson, K.A.; Leimgruber, P.; Calabrese, J.M. Rigorous home range estimation with movement data: A new autocorrelated kernel density estimator. Ecology 2015, 96, 1182–1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swindle, K.A.; Ripple, W.J.; Meslow, E.C.; Schäfer, D. Old-Forest Distribution around Spotted Owl Nests in the Central Cascade Mountains, Oregon. J. Wildl. Manag. 1999, 63, 1212–1221. [Google Scholar] [CrossRef]
- Von Christoph, L. Telemetric study in the Eagl-Owl (Bubo bubo) in the foreland of the Alps in Lower Austria—Methods and first results. Egretta 1992, 35, 69–71. [Google Scholar]
- Pande, S.A.; Yosef, R.; Deshpande, P.; Pawar, R.; Mahajan, M.N. Breeding biology of the mottled wood owl (Strix ocellata) in west-central inida. J. Raptor Res. 2018, 52, 240–244. [Google Scholar] [CrossRef]
- Dalbeck, L.; Bergerhausen, W.; Krischer, O. Telemtriestudie zur Orts- und Partnertreue bei Uhu Bubo bubo. Vogelwelt 1998, 119, 337–344. [Google Scholar]
- Raudenbush, S.W.; Bryk, A.S. Hierarchical Linear Models: Applications and Data Analysis Methods; Sage: Thousand Oaks, CA, USA, 2002; Volume 1. [Google Scholar]
- Martínez, J.E.; Calvo, J.F. Seleccion de habitat de nidificacion por el buho real Bubo bubo en ambientes mediterraneos semiaridos. Ardeola 2000, 47, 215–220, (In Spanish, English Abstract). [Google Scholar]
- Milgrom, T. Environmental aspects of rehabilitating abandoned quarries: Israel as a case study. Landsc. Urban Plan. 2008, 87, 172–179. [Google Scholar] [CrossRef]
- Penteriani, V.; Gallardo, M.; Ferrer, M. Spatial heterogeneity and structure of bird populations: A case example with the eagle owl. Popul. Ecol. 2004, 46, 185–192. [Google Scholar] [CrossRef] [Green Version]
- León-Ortega, M.; Martínez, J.E.; Pérez, E.; LaCalle, J.A.; Calvo, J.F. The contribution of non-protected areas to the conservation of Eurasian Eagle-owls in Mediterranean ecosystems. Ecosphere 2017, 8, e01952. [Google Scholar] [CrossRef]
- Ortego, J. Consequences of Eagle Owl nest-site habitat preference for breeding performance and territory stability. Ornis Fenn. 2007, 84, 78–90. [Google Scholar]
- Symes, A.; Taylor, J.; Mallon, D.; Porter, R.; Simms, C.; Budd, K. The Conservation Status and Distribution of the Breeding Birds of the Arabian Peninsula; IUCN: Cambridge, UK; Gland, Switzerland; Environment and Protected Areas Authority: Sharjah, United Arab Emirates, 2015. [CrossRef]
- Orenstein, D.E.; Hamburg, S.P. Population and pavement: Population growth and land development in Israel. Popul. Environ. 2010, 31, 223–254. [Google Scholar] [CrossRef]
- Levin, N.; Lahav, H.; Ramon, U.; Heller, A.; Nizry, G.; Tsoar, A.; Sagi, Y. Landscape continuity analysis: A new approach to conservation planning in Israel. Landsc. Urban Plan. 2007, 79, 53–64. [Google Scholar] [CrossRef]
- Tipling, D. Bird photography—A new code of practice. Br. Birds 2011, 104, 223–224. [Google Scholar]
- Radulski, B.G. Understanding Human Disturbance to Birds at the Intersection of Birding and Bird Photography. Ph.D. Thesis, Purdue University Graduate School, West Lafayette, IN, USA, 2020. [Google Scholar]
- Martínez, J.E.; Martínez, J.A.; Zuberogoitia, I.; Zabala, J.; Redpath, S.M.; Calvo, J.F. The effect of intra- and interspecific interactions on the large-scale distribution of cliff-nesting raptors. Ornis Fenn. 2008, 85, 13–21. [Google Scholar]
- Iezekiel, S.; Yosef, R.; Bakaloudis, D.E.; Papakosta, M.A.; Vlachos, C.G.; Antoniou, A.; Zduniak, P. The endemic Cyprus Wheatear (Oenanthe cypriaca) adapts readily to artificial nest sites. Biol. Conserv. 2017, 213, 1–4. [Google Scholar] [CrossRef]
- Iezekiel, S.; Yosef, R.; Themistokleus, C.; Bakaloudis, D.; Vlachos, C.; Antoniou, A.; Iezekiel, E.; Papakosta, M.; Kosicki, J. Endemic Cyprus Scops Owl Otus cyprius Readily Breeds in Artificial Nest Boxes. Animals 2021, 11, 1775. [Google Scholar] [CrossRef]
- Crooks, K.R.; Soulé, M.E. Mesopredator release and avifaunal extinctions in a fragmented system. Nature 1999, 400, 563–566. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Scale (km) | 1000 | 2000 | 3000 | 4000 | 5000 |
|---|---|---|---|---|---|
| Built | 8.4 | 9.1 | 10.4 | 11.4 | 12.1 |
| Disturbed | 2.3 | 1.5 | 1.1 | 1.1 | 1.1 |
| Grasslands | 0.8 | 1 | 1.1 | 1.2 | 1.3 |
| Maquis | 10.5 | 9.8 | 9.2 | 8.6 | 8.2 |
| Shrub | 12.4 | 10.9 | 10.2 | 9.9 | 9.6 |
| Grove | 22.4 | 21.7 | 22.1 | 22.2 | 21.9 |
| Woods | 19.4 | 18 | 16.6 | 15.9 | 15.5 |
| Agriculture | 18.3 | 21.5 | 22.5 | 23 | 23 |
| Orchards | 5.4 | 6.5 | 6.7 | 6.6 | 7.1 |
| Water | 0.1 | 0.1 | 0.1 | 0.2 | 0.2 |
| Radius | AIC | Observed Mean | Simulated Mean | p-Value Model | p-Value Variable | |
|---|---|---|---|---|---|---|
| 1000 | Cave | 191.664 | 0.73 | 0.069 | 0.0001 | 0.0120 |
| Quarry | 176.457 | 0.60 | 0.04 | <0.0001 | 0.0004 | |
| C + Q | 153.781 | <0.0001 | <0.0001 | |||
| 2000 | Cave | 192.633 | 1.67 | 0.26 | 0.0002 | 0.0130 |
| Quarry | 178.548 | 1.05 | 0.12 | <0.0001 | 0.0005 | |
| C + Q | 159.405 | <0.0001 | <0.0001 | |||
| 3000 | Cave | 194.437 | 2.77 | 0.60 | 0.0005 | 0.0137 |
| Quarry | 190.323 | 1.29 | 0.36 | <0.0001 | 0.0006 | |
| C + Q | 175.112 | <0.0001 | <0.0001 | |||
| 4000 | Cave | 201.343 | 3.81 | 1.66 | 0.0234 | 0.0373 |
| Quarry | 191.845 | 1.73 | 0.59 | 0.0001 | 0.0006 | |
| C + Q | 192.764 | 0.0002 | 0.0022 | |||
| 5000 | Cave | 200.991 | 5.45 | 2.42 | 0.0191 | 0.0290 |
| Quarry | 190.916 | 2.18 | 0.85 | <0.0001 | 0.0003 | |
| C + Q | 193.841 | 0.0004 | 0.0022 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hadad, E.; Malkinson, D.; Yosef, R.; Weil, G.; Charter, M. Importance of Mesohabitat for Nest-Site Selection in Breeding Eagle Owls (Bubo bubo): A Multi-Scale Model. Diversity 2022, 14, 438. https://doi.org/10.3390/d14060438
Hadad E, Malkinson D, Yosef R, Weil G, Charter M. Importance of Mesohabitat for Nest-Site Selection in Breeding Eagle Owls (Bubo bubo): A Multi-Scale Model. Diversity. 2022; 14(6):438. https://doi.org/10.3390/d14060438
Chicago/Turabian StyleHadad, Ezra, Dan Malkinson, Reuven Yosef, Gilad Weil, and Motti Charter. 2022. "Importance of Mesohabitat for Nest-Site Selection in Breeding Eagle Owls (Bubo bubo): A Multi-Scale Model" Diversity 14, no. 6: 438. https://doi.org/10.3390/d14060438
APA StyleHadad, E., Malkinson, D., Yosef, R., Weil, G., & Charter, M. (2022). Importance of Mesohabitat for Nest-Site Selection in Breeding Eagle Owls (Bubo bubo): A Multi-Scale Model. Diversity, 14(6), 438. https://doi.org/10.3390/d14060438