
Factors Associated with the Occurrence, Potential Distribution and Conservation of Anoxia orientalis (Coleoptera, Scarabaeidae) at Different Spatial Scales


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
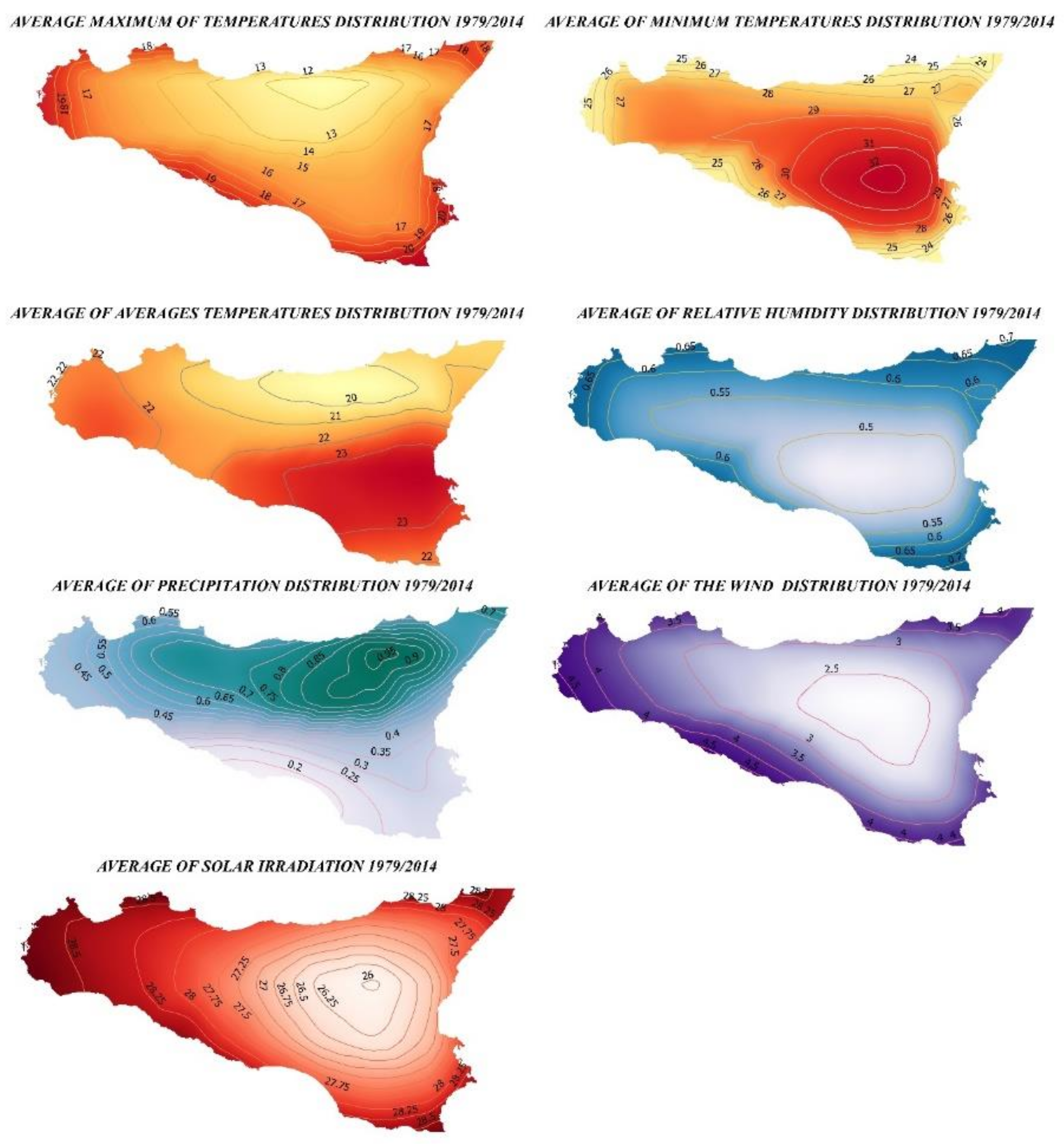
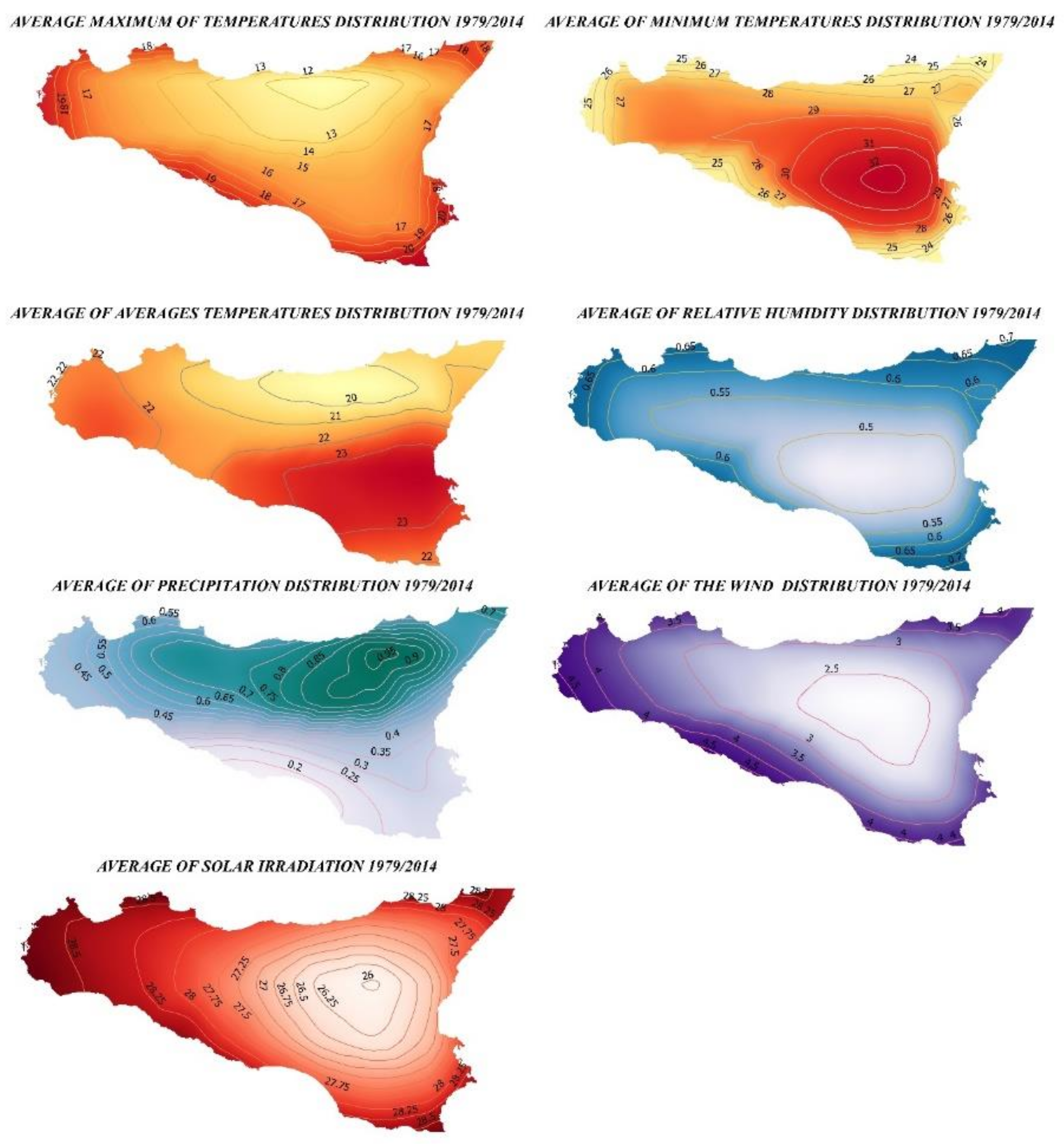
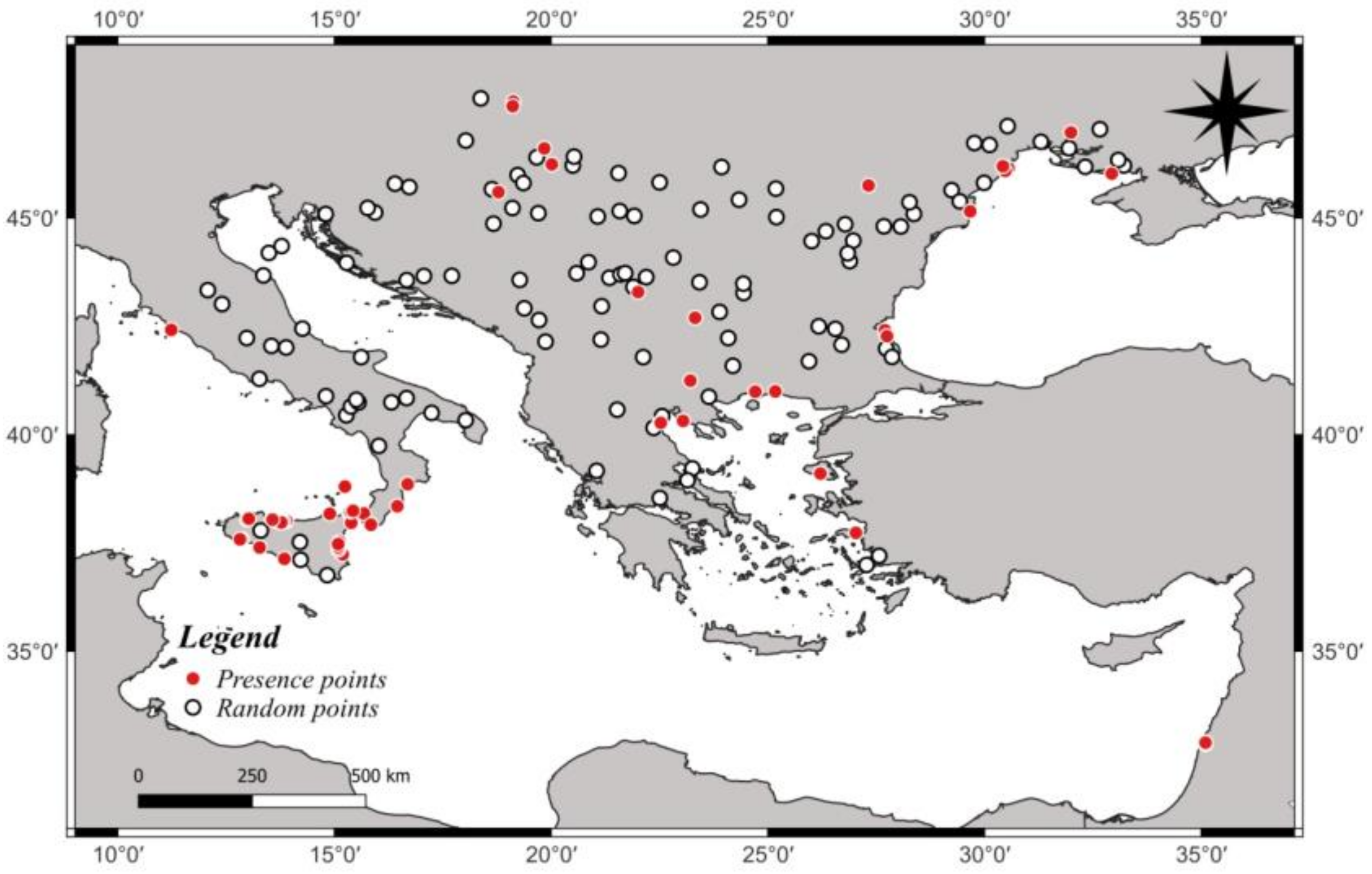
2.1. Study Area and Protocol
2.2. Statistical Analyses
3. Results
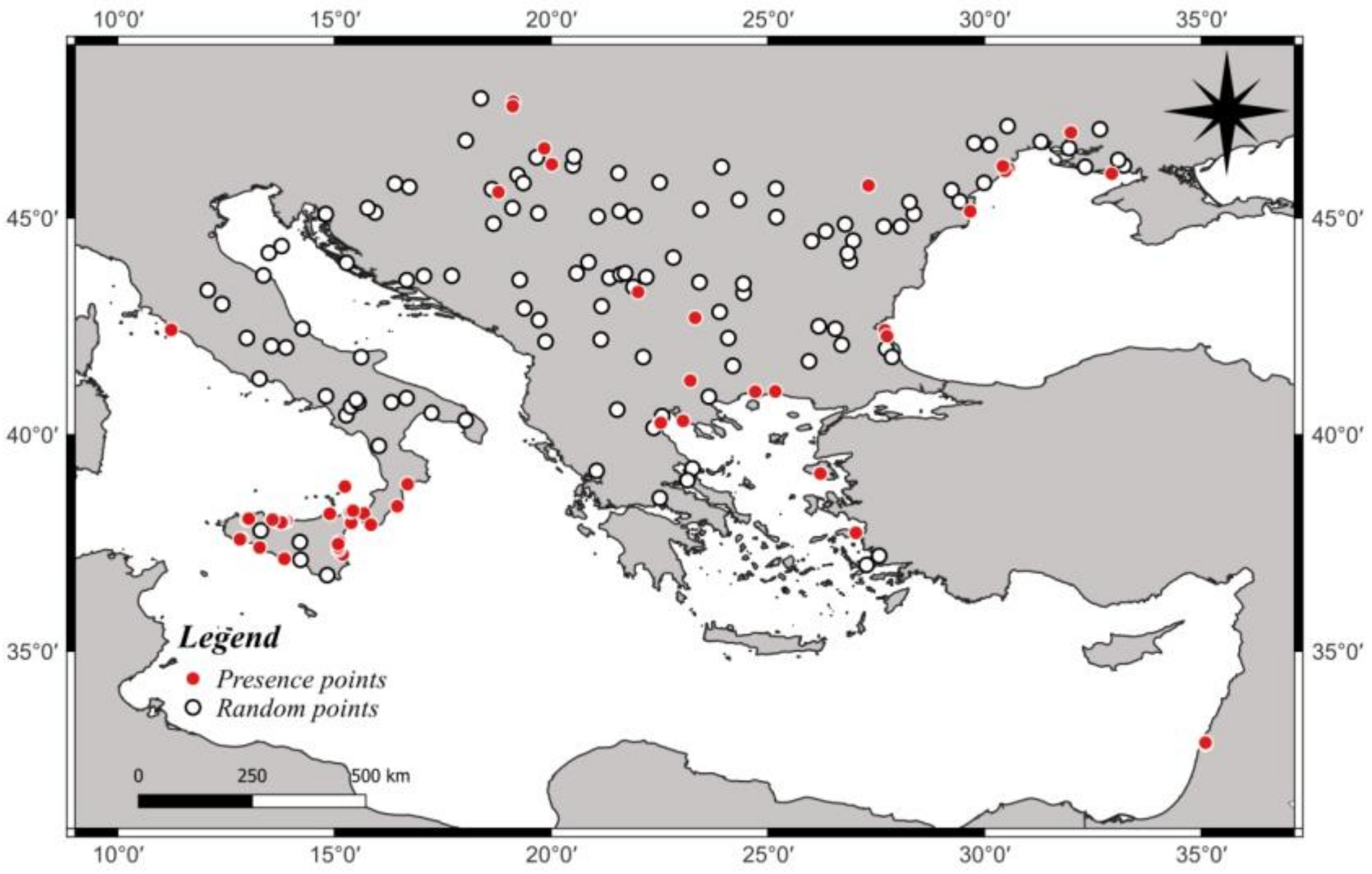
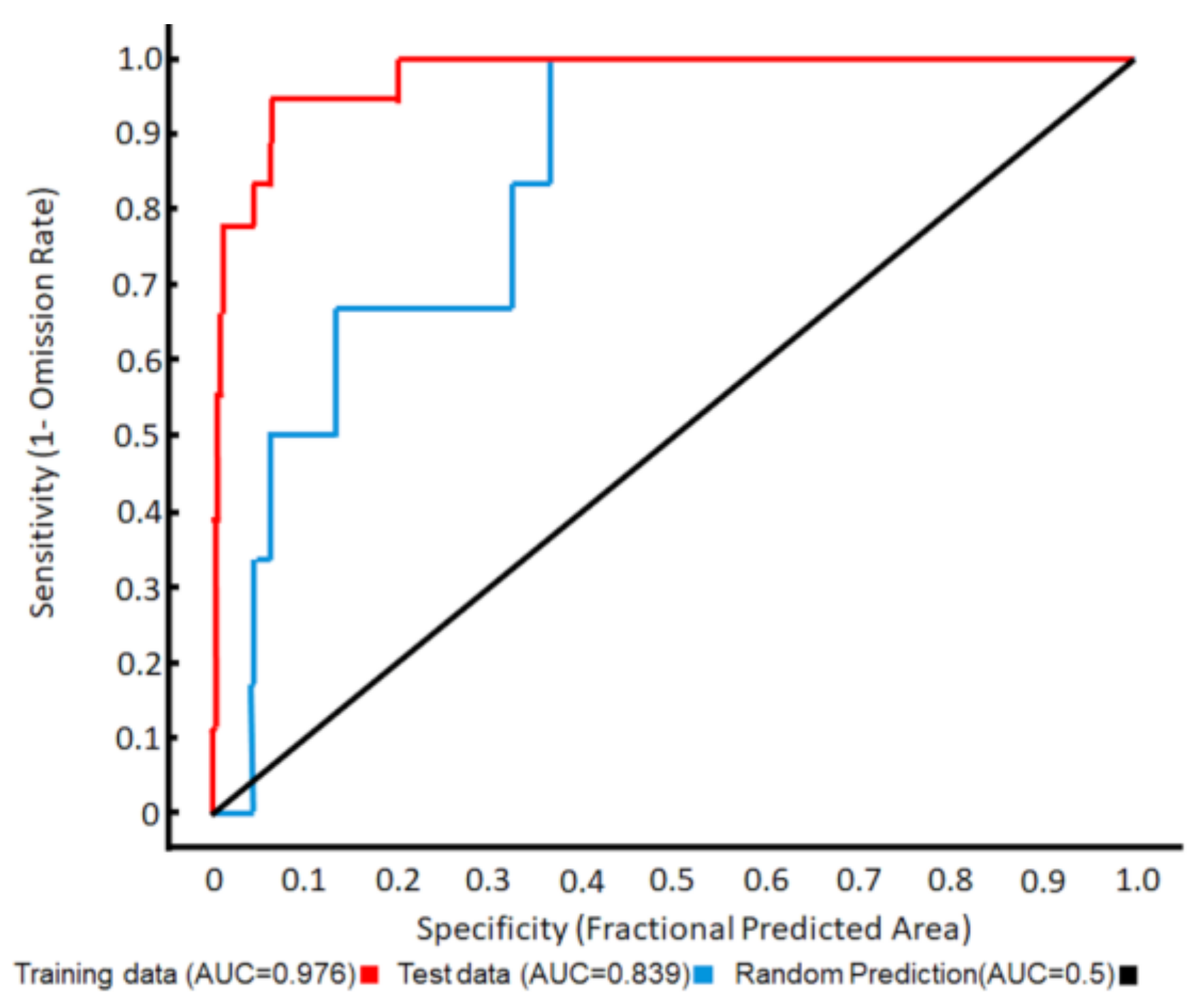
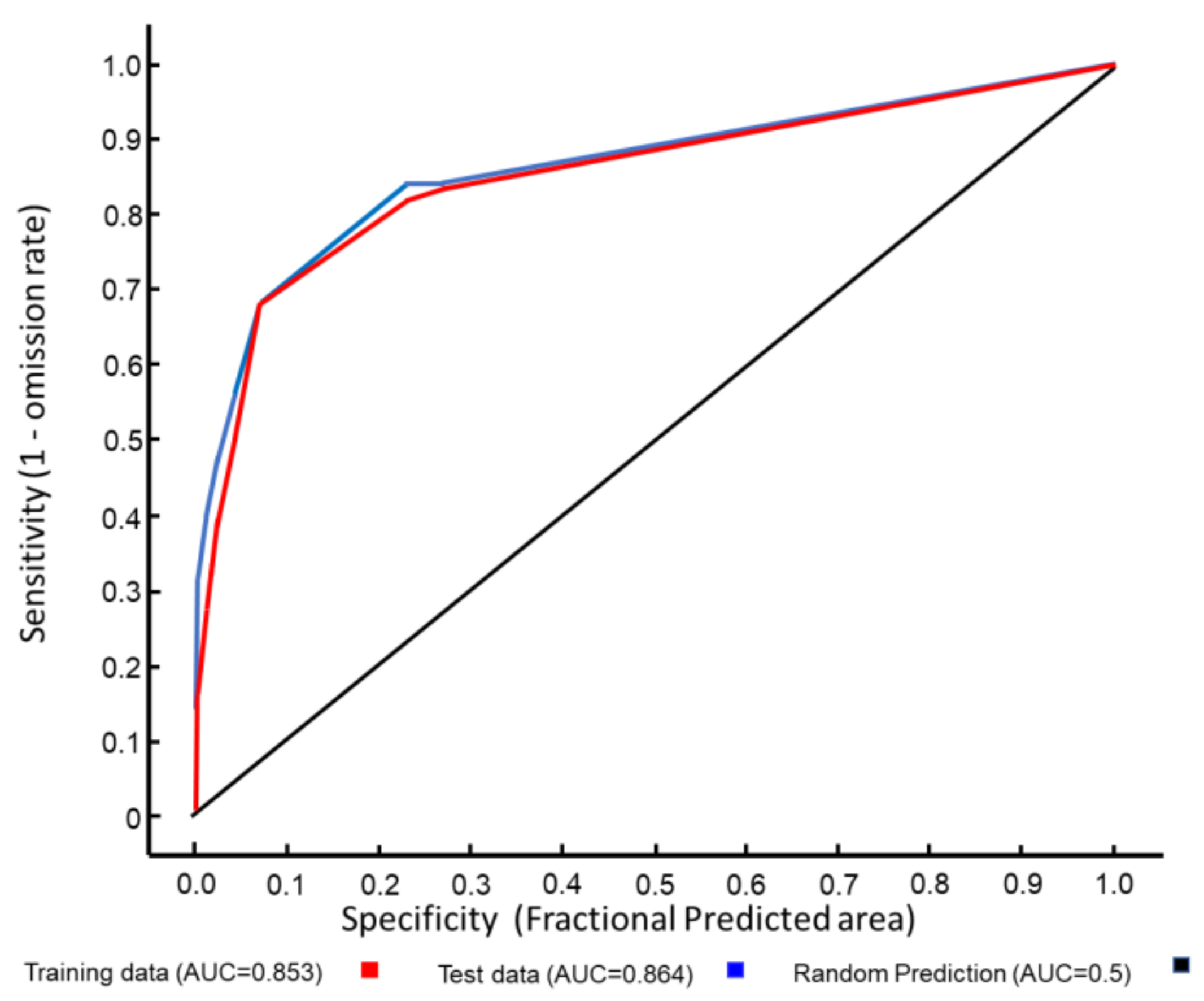
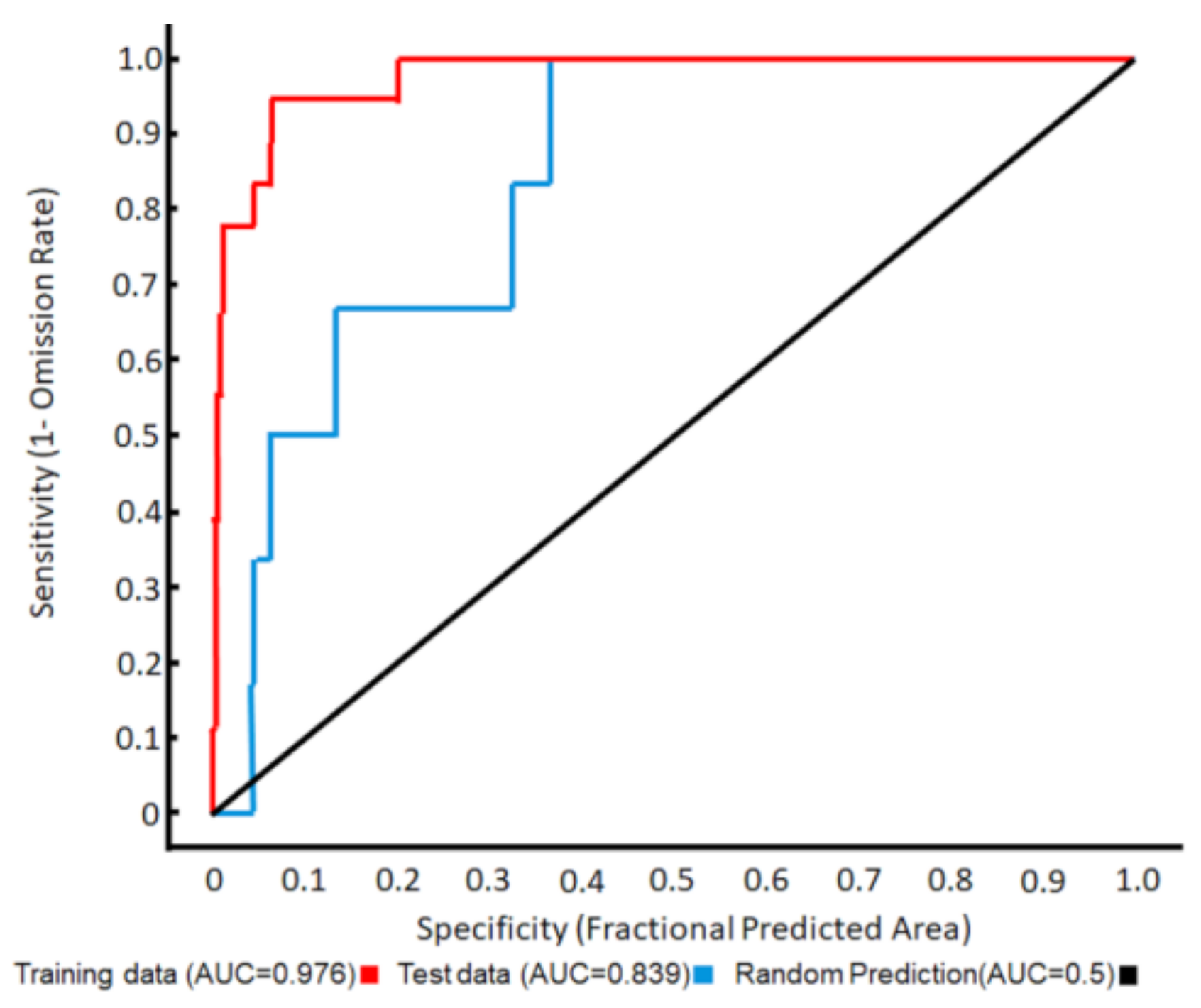
3.1. Ecological Modelling at Two Spatial Scales
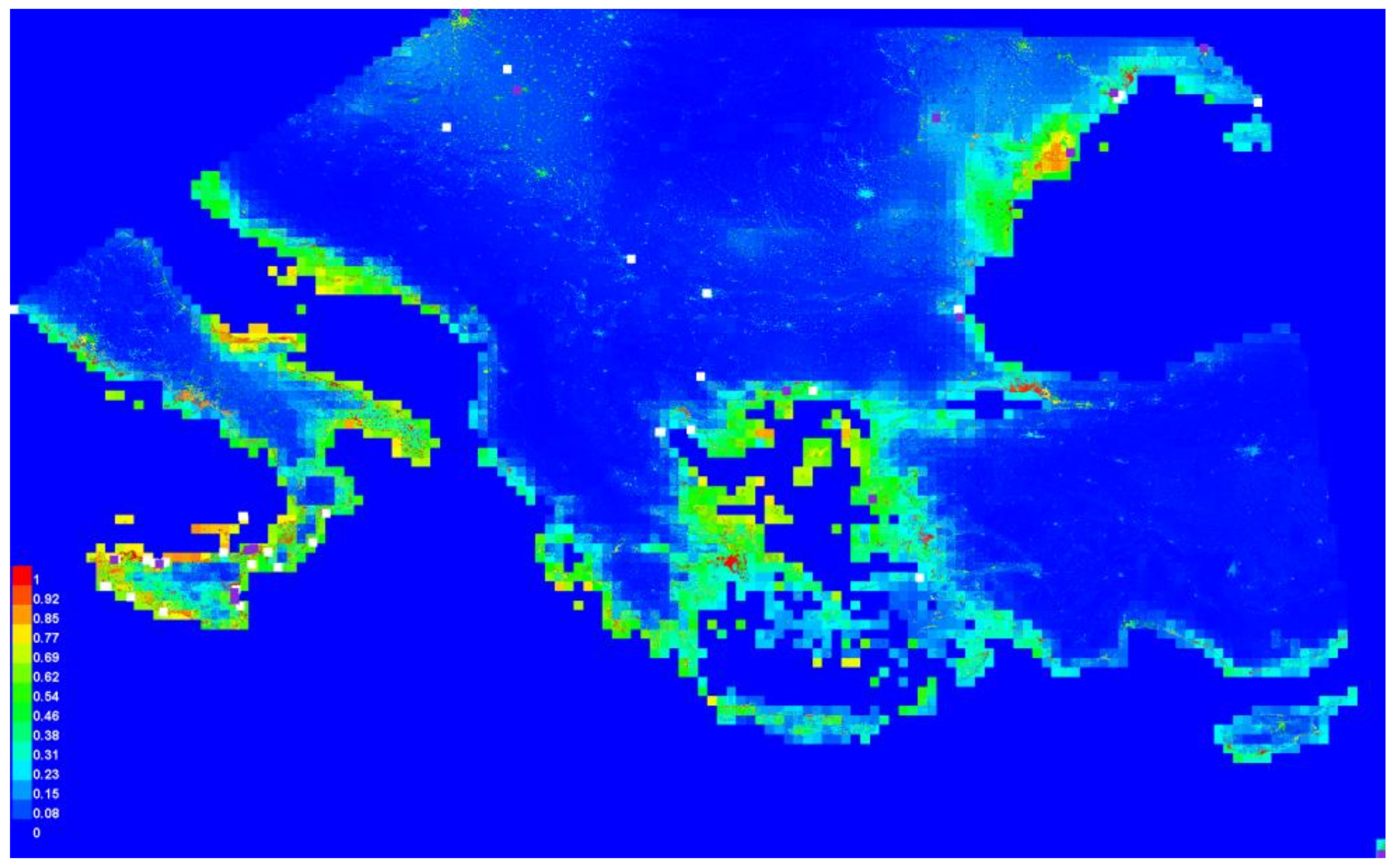
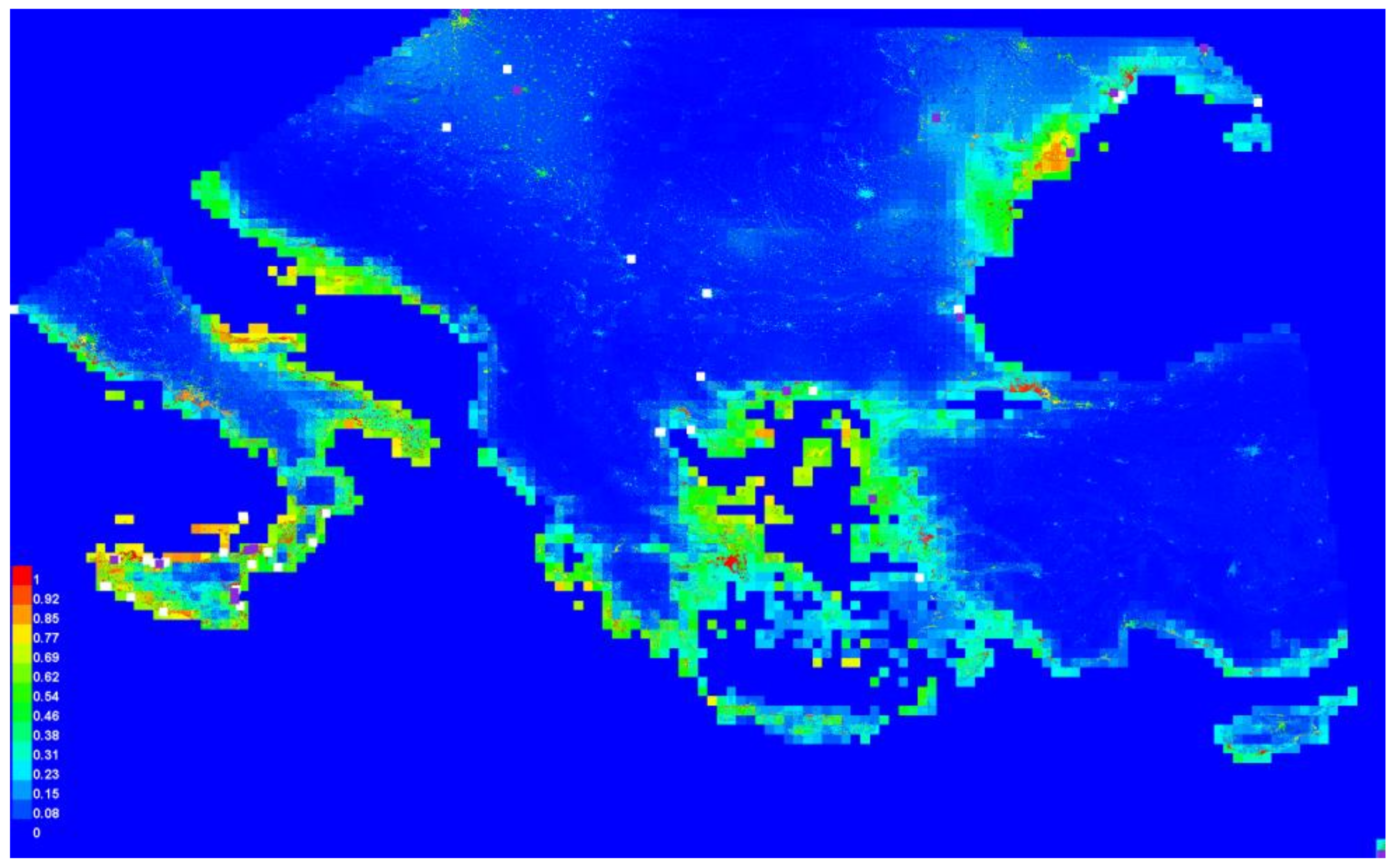
3.2. Graphical Model Map for Anoxia Orientalis in Europe
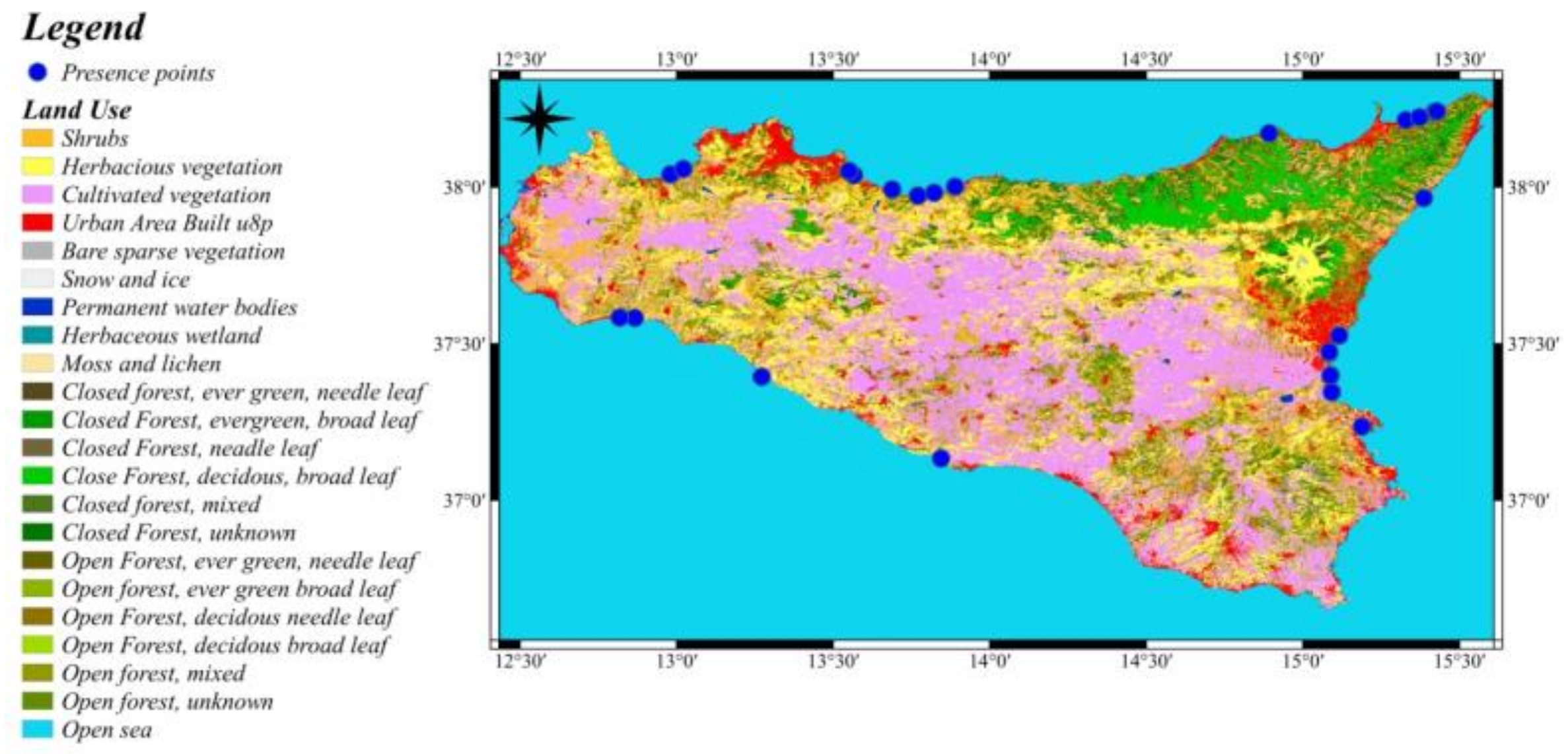
3.3. Graphical Model Map for Anoxia Orientalis in Sicily
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Appendix B

Appendix C
References
- Hengeveld, R. Dynamic Biogeography; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- Franklin, J. Moving beyond static species distribution models in support of conservation biogeography. Div. Distrib. 2010, 16, 321–330. [Google Scholar] [CrossRef]
- Vigna Taglianti, A.; Audisio, P.; Belfiore, C.M.; Biondi, M.; Bologna, M.A.; Carpaneto, G.M.; De Biase, A.; De Felici, S.; Piattella, E.; Racheli, T.; et al. Riflessioni di gruppo sui corotipi fondamentali della fauna W-paleartica ed in particolare italiana. Biogeographia 1992, 16, 159–179. [Google Scholar] [CrossRef] [Green Version]
- Carpaneto, G.M.; Piattella, E.; Pittino, R. The scarab beetles of Turkey: An updated checklist and chorotype analysis (Coleoptera, Scarabaeoidea). Biogeographia 2000, 21, 218–240. [Google Scholar] [CrossRef] [Green Version]
- Çobanoğlu, S.; Çakmak, I.; Başpinar, H. Hypoaspis krameri (Canestrini, 1881) (Mesostigmata: Laelapidae) an ectoparasitic mite associated with Anoxia orientalis Kryn. (Col., Scarabaeidae) from Turkey. Entomol. Mon. Mag. 2003, 139, 97–101. [Google Scholar]
- Baraud, J. Révision des Anoxia Castelnau d’Europe et d’Asie 1re note: Le sous-genre Protanoxia Medvedev (Coleoptera, Melolonthidae). Bull. Soc. Entomol. Fr. 1989, 93, 273–284. [Google Scholar] [CrossRef]
- Bezdek, A. Scarabaeidae: Melolonthinae: Melolonthini. In Catalogue of Palaearctic Coleoptera, Vol. 3. Scarabaeoidea–Scirtoidea–Dascilloidea–Buprestoidea–Byrrhoidea; Löbl, I., Löbl, D., Eds.; Brill: Leiden, The Netherlands, 2016; pp. 226–236. [Google Scholar]
- Rittner, O. Synopsis of the Melolonthini (Scarabaeidae: Melolonthinae) of Israel, with a first description of the female of Anoxia (Protanoxia) laevimacula Petrovitz, 1973. Israel J. Entomol. 2016, 46, 99–108. [Google Scholar]
- Perazzini, G. Osservazioni morfologiche e geonemiche su alcuni Scarabaeidae floricoli italiani (Coleoptera, Scarabaeidae). Acta Coleopt. 1987, 3, 33–40. [Google Scholar]
- Luigioni, P. I Coleotteri d’Italia. Catalogo sinonimico-topografico-bibliografico. Mem. Pontif. Accad. Sci. Nuovi Lincei (Ser. II) 1912, 13, 1–1159. [Google Scholar]
- Mariani, G. Ricerche coleotterologiche sul litorale ionico di Puglia, Lucania e Calabria. Campagne 1956-1957-1958. II. Coleoptera Lamellicornia. Mem. Soc. Entomol. Ital. Suppl. 1959, 38, 143–184. [Google Scholar]
- Mikšic, R. Beitrag zur kenntnis der Lamellicornia-Fauna der Apenninen. IV. Aspromonte. Mem. Mus. Civ. St. Nat. Verona 1961, 9, 5–25. [Google Scholar]
- Tassi, F. Reperti 5. Anoxia orientalis Kryn. (Col. Scarabaeidae). Boll. Assoc. Romana Entomol. 1967, 22, 46. [Google Scholar]
- Arnone, M. Quinto contributo alla revisione della collezione coleotterologica di Enrico Ragusa: Scarabaeoidea. Nat. Sicil. 2010, 34, 61–172. [Google Scholar]
- Sabella, G.; Sparacio, I. Il ruolo dei Parchi siciliani nella conservazione di taxa di Insetti di particolare interesse naturalistico (Insecta Coleoptera et Lepidoptera Rhopalocera). Nat. Sicil. 2004, 28, 447–508. [Google Scholar]
- Lapiana, F.; Sparacio, I. Coleotteri Lamellicorni delle Madonie (Sicilia) (Insecta Coleoptera Lucanoidea et Scarabaeoidea). Naturalista Sicil. (Ser. IV) 2006, 30, 227–292. [Google Scholar]
- Schatzmayr, A. Appunti coleotterologici. 12. Natura 1944, 35, 54–59. [Google Scholar]
- Homonnay, F. Data on the biology and development of soil-inhabiting pests in Hungary. Növényvédelem 1989, 25, 492–499. [Google Scholar]
- Dile, Y.T.; Srinivasan, R. Evaluation of CFSR climate data for hydrologic prediction in data-scarce watersheds: An application in the Blue Nile River Basin. J. Am. Water Resour. Assoc. 2014, 1, 1226–1241. [Google Scholar] [CrossRef]
- Fuka, D.R.; MacAllister, C.A.; Degaetano, A.T.; Easton, Z.M. Using the Climate Forecast System Reanalysis dataset to improve weather input data for watershed models. Hydrol. Proc. 2013, 30, 5613–5623. [Google Scholar] [CrossRef]
- Buchhorn, M.; Smets, B.; Bertels, L.; Lesiv, M.; Tsendbazar, N.E.; Herold, M.; Fritz, S. Copernicus Global Land Service: Land Cover 100m: Epoch 2015: Globe; (Version V2.0.2); Zenodo: Geneva, Switzerland, 2019. [Google Scholar]
- Hosmer, D.W.; Lemeshow, S. Applied Logistic Regression; Wiley: New York, NY, USA, 2000. [Google Scholar]
- Teixeira, J.; Ferrand, N.; Arntzen, J.W. Biogeography of the golden-striped salamander Chioglossa lusitanica: A field survey and spatial modelling approach. Ecography 2001, 24, 618–624. [Google Scholar] [CrossRef]
- Arntzen, J.W.; Alexandrino, J. Ecological modelling of genetically differentiated forms of the Iberian endemic golden-striped salamander, Chioglossa lusitanica. Herpetol. J. 2004, 14, 137–141. [Google Scholar]
- Holm, S. A simple sequential rejective multiple test procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Muscarella, C.; Luiselli, L.; Di Vittorio, M.; Sparacio, I.; Amori, G.; Dendi, D. Factors associated with occurrence, potential distribution and conservation of Polyphylla ragusae, an endemic Scarabaeidae (Melolonthinae) from Sicily. J. Insect. Conserv. 2022, in press. [Google Scholar]
- Aliquò, V.; Massa, B. Contributo allo studio di Anoxia scutellaris s. l. e descrizione di Anoxia scutellaris argentea n. ssp. di Sicilia. Boll. Soc. Ent. Ital. Genova 1976, 108, 151–157. [Google Scholar]
- Sabatinelli, G. Note su alcuni Scarabeoidea floricoli dell’ Italia meridionale e descrizione di Amphimallon pseudomajale n.sp. (Coleoptera). Boll. Ass. Romana Entomol. 1976, 31, 35–46. [Google Scholar]
- Tóth, J. Damage caused by cockchafers (Melolontha spp.) in Hungary during the last 30 years. In Proceedings of the Population Dynamics, Impacts, and Integrated Management of Forest Defoliating Insects, Banska Stiavnica, Slovakia, 18–23 August 1996; McManus, M.L., Liebhold, A.M., Eds.; Department of Agriculture, Forest Service, Northeastern Research Station: Radnor, PA, USA, 1998; p. 341. [Google Scholar]
- Lapiana, F.; Sparacio, I. Lo studio degli Insetti nella valutazione della naturalità degli ambienti dunali costieri in Sicilia: Coleoptera e Orthoptera. Nat. Sicil. 2008, 32, 411–434. [Google Scholar]
- Aliquo, V.; Massa, B.; Mignani, R. Brevi note sulla fauna coleotterologica di un particolare biotopo costiero del Palermitano (Coleoptera). Boll. Soc. Entomol. Ital. 1973, 105, 59–68. [Google Scholar]


| Definition According UN LCCS | MAP COCDE | Land Cover Class |
|---|---|---|
| These are woody perennial plants with persistent and woody stems and without any defined main stem being less than 5 m tall. The shrub foliage can be either evergreen or deciduous. | HISTO_20 | Shrubs |
| Plants without persistent stem or shoots above ground and lacking definite firm structure. Tree and shrub cover is less than 10%. | HISTO_30 | Herbaceous vegetation |
| Lands covered with temporary crops followed by harvest and a bare soil period (e.g., single and multiple cropping systems). Note that perennial woody crops will be classified as the appropriate forest or shrub land cover type. | HISTO_40 | Cultivated and managed vegetation/agriculture (cropland) |
| Land covered by buildings and other man-made structures | HISTO_50 | Urban/built up |
| Lands with exposed soil, sand or rocks and never has more than 10% vegetated cover during any time of the year | HISTO_60 | Bare/sparse vegetation |
| Lakes, reservoirs and rivers. Can be either fresh or salt-water bodies. | HISTO_80 | Permanent freshwater bodies |
| Land with a permanent mixture of water and herbaceous or woody vegetation. The vegetation can be present in either salt, brackish or fresh water. | HISTO_90 | Herbaceous wetland |
| Tree canopy > 70%, almost all needle leaf trees remain green all year. Canopy is never without green foliage. | HISTO_111 | Closed forest, evergreen, needle leaf |
| Tree canopy > 70%, consists of seasonal broadleaf tree communities with an annual cycle of leaf-on and leaf-off periods. | HISTO_114 | Closed forest, deciduous broad leaf |
| Closed forest, mix of types | HISTO_115 | Closed forest, mix |
| Closed forest, not matching any of the other definitions | HISTO_116 | Closed forest, unknown |
| Top layer trees 15–70% and second layer-mixed of shrubs and grassland, almost all needle leaf trees remain green all year. Canopy is never without green foliage. | HISTO_121 | Open forest, evergreen needle leaf |
| Top layer trees 15–70% and second layer mixed of shrubs and grassland, consists of seasonal broadleaf tree communities with an annual cycle of leaf-on and leaf-off periods. | HISTO_124 | Open forest, deciduous broad leaf |
| Open forest, not matching any of the other definitions | HISTO_126 | Open forest, unknown |
| Permanent freshwater bodies, lagoons, estuaries | HISTO_200 | Freshwater systems |
| Variable | All Data | Europe without Sicily | Sicily | |||
|---|---|---|---|---|---|---|
| Estimate | p-Value | Estimate | p-Value | Estimate | p-Value | |
| HISTO_20 | 3.88 | 0.049 | 0.036 | 0.849 | 8.406 | 0.004 |
| HISTO_30 | 0.127 | 0.721 | 0.614 | 0.433 | 5.776 | 0.016 |
| HISTO_40 | 30.951 | 0.0001 | 7.551 | 0.006 | 11.421 | 0.001 |
| HISTO_50 | 48.735 | 0.0001 | 49.044 | 0.0001 | 22.166 | 0.00001 |
| HISTO_80 | 16.985 | 0.0001 | 0.978 | 0.323 | 45.693 | 0.00001 |
| HISTO_90 | 3.291 | 0.07 | 4.947 | 0.026 | 10.266 | 0.001 |
| HISTO_110 | 4.113 | 0.043 | 1.738 | 0.187 | 0.822 | 0.365 |
| HISTO_114 | 29.139 | 0.0001 | 13.39 | 0.0001 | 3.911 | 0.048 |
| HISTO_115 | 1.945 | 0.163 | 0.64 | 0.424 | 8.215 | 0.004 |
| HISTO_116 | 1.125 | 0.289 | 2.964 | 0.085 | 3.963 | 0.047 |
| HISTO_121 | 0.547 | 0.459 | 1.614 | 0.204 | 18.176 | 0.00001 |
| HISTO_124 | 12.828 | 0.0001 | 5.478 | 0.019 | 8.406 | 0.004 |
| HISTO_126 | 23.615 | 0.0001 | 12.485 | 0.0001 | 5.776 | 0.016 |
| HISTO_200 | 3.223 | 0.0781 | 3.576 | 0.059 | 11.421 | 0.001 |
| Variable | % Contribution | Permutation Importance |
|---|---|---|
| Minimum Temperature | 30.7 | 44.9 |
| Land Use | 22.9 | 11.6 |
| Average Temperature | 19 | 0 |
| Maximum Temperature | 18.3 | 4 |
| Wind | 7.9 | 17.7 |
| Precipitation | 1.2 | 21.8 |
| Variable | % Contribution | Permutation Importance |
|---|---|---|
| Land Use | 58.1 | 23.0 |
| Minimum Temperature | 12.3 | 2.2 |
| Relative Humidity | 10.2 | 30.8 |
| Precipitation | 6.8 | 19.5 |
| Average Temperature | 6.1 | 19.4 |
| Wind | 4.7 | 2.7 |
| Solar Irradiation | 1.7 | 2.3 |
| Maximum Temperature | 0.0 | 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muscarella, C.; Luiselli, L.; Di Vittorio, M.; Sparacio, I.; Dendi, D. Factors Associated with the Occurrence, Potential Distribution and Conservation of Anoxia orientalis (Coleoptera, Scarabaeidae) at Different Spatial Scales. Diversity 2022, 14, 397. https://doi.org/10.3390/d14050397
Muscarella C, Luiselli L, Di Vittorio M, Sparacio I, Dendi D. Factors Associated with the Occurrence, Potential Distribution and Conservation of Anoxia orientalis (Coleoptera, Scarabaeidae) at Different Spatial Scales. Diversity. 2022; 14(5):397. https://doi.org/10.3390/d14050397
Chicago/Turabian StyleMuscarella, Calogero, Luca Luiselli, Massimiliano Di Vittorio, Ignazio Sparacio, and Daniele Dendi. 2022. "Factors Associated with the Occurrence, Potential Distribution and Conservation of Anoxia orientalis (Coleoptera, Scarabaeidae) at Different Spatial Scales" Diversity 14, no. 5: 397. https://doi.org/10.3390/d14050397
APA StyleMuscarella, C., Luiselli, L., Di Vittorio, M., Sparacio, I., & Dendi, D. (2022). Factors Associated with the Occurrence, Potential Distribution and Conservation of Anoxia orientalis (Coleoptera, Scarabaeidae) at Different Spatial Scales. Diversity, 14(5), 397. https://doi.org/10.3390/d14050397