
Qipengyuania pacifica sp. nov., a Novel Carotenoid-Producing Marine Bacterium of the Family Erythrobacteraceae, Isolated from Sponge (Demospongiae), and Antimicrobial Potential of Its Crude Extract



Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation, Cultivation, and Maintenance of Bacteria
2.2. Morphological, Cultural, and Physiological Characterization
2.3. 16S rRNA Gene Sequence and Phylogenetic Analysis
2.4. Chemotaxonomy
2.5. Secondary Metabolites Production and Minimum Inhibitory Concentration Assay
2.6. Genome Analysis
3. Results and Discussions
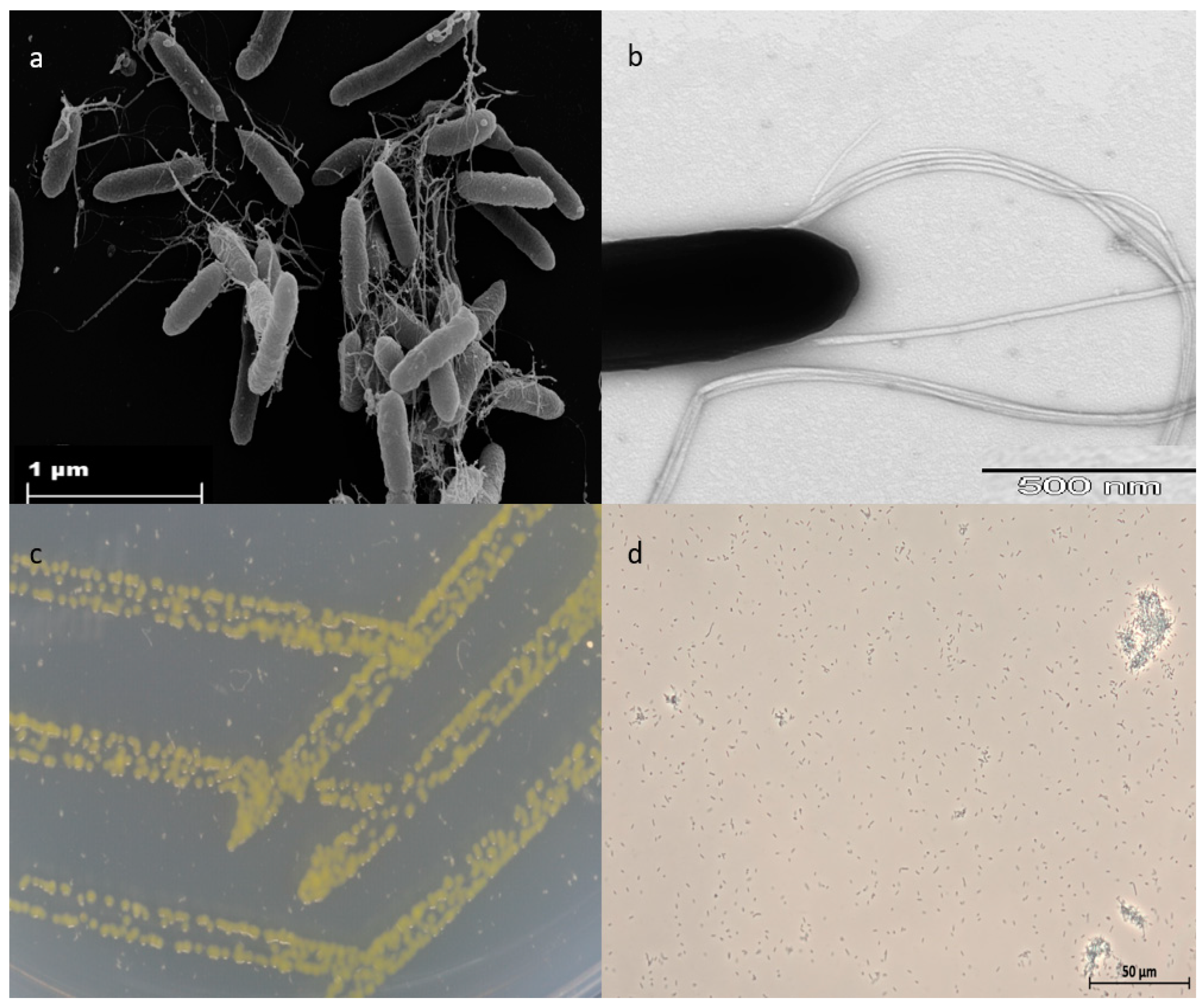
3.1. Morphological and Physiological Characteristics
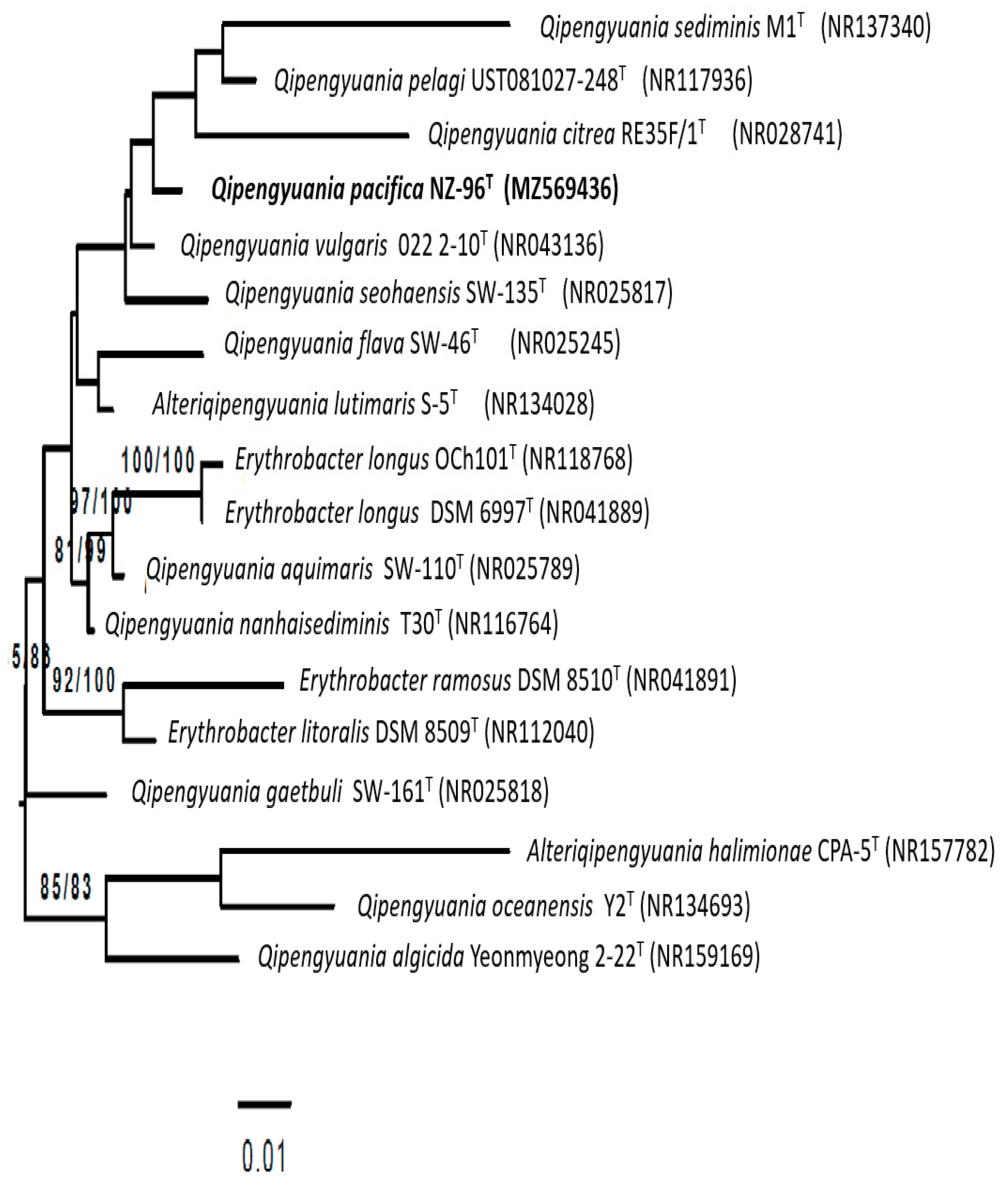
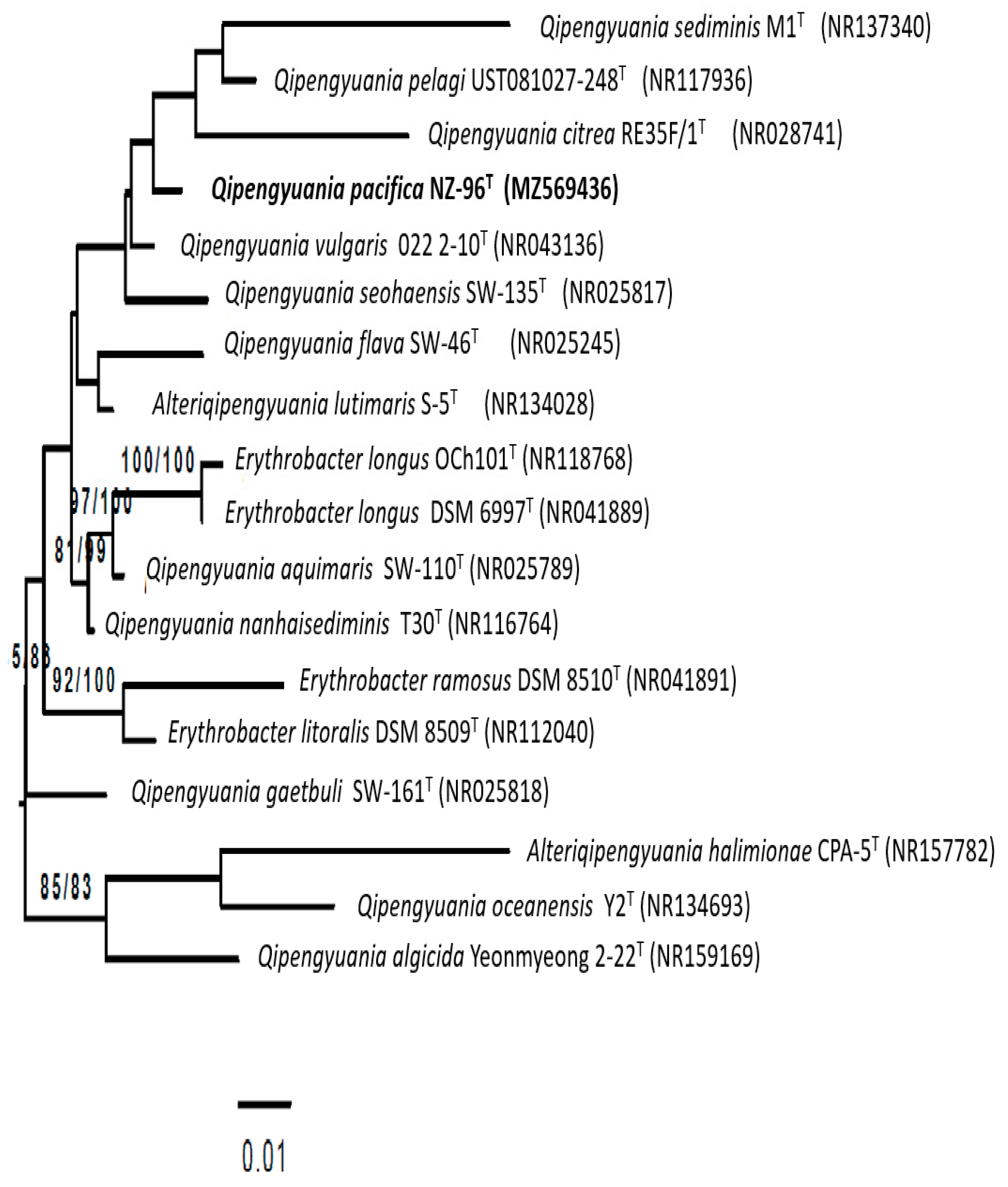
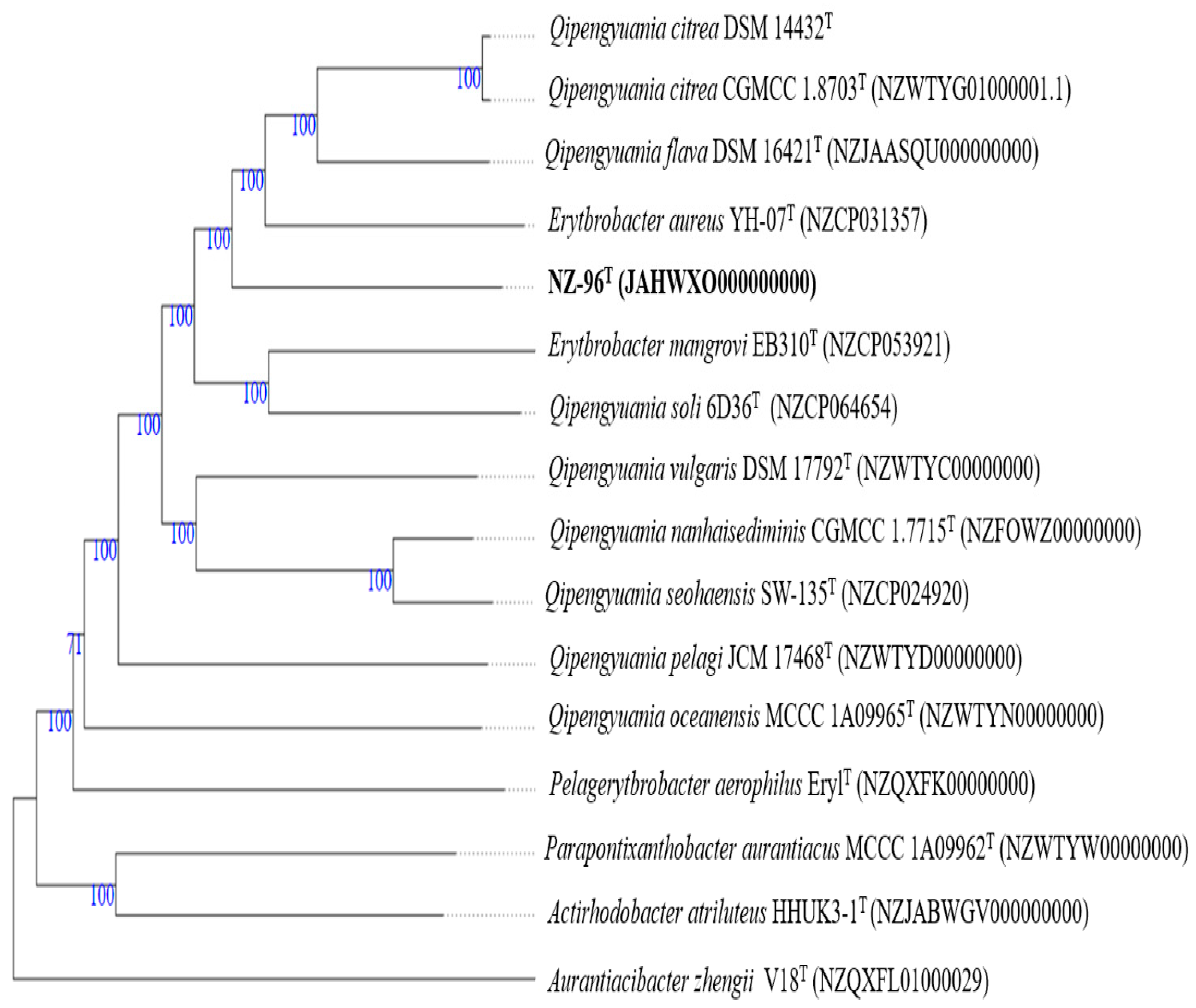
3.2. 16S rRNA Gene-Based Phylogenetic Analysis
3.3. Chemotaxonomy
3.4. Antimicrobial Potential
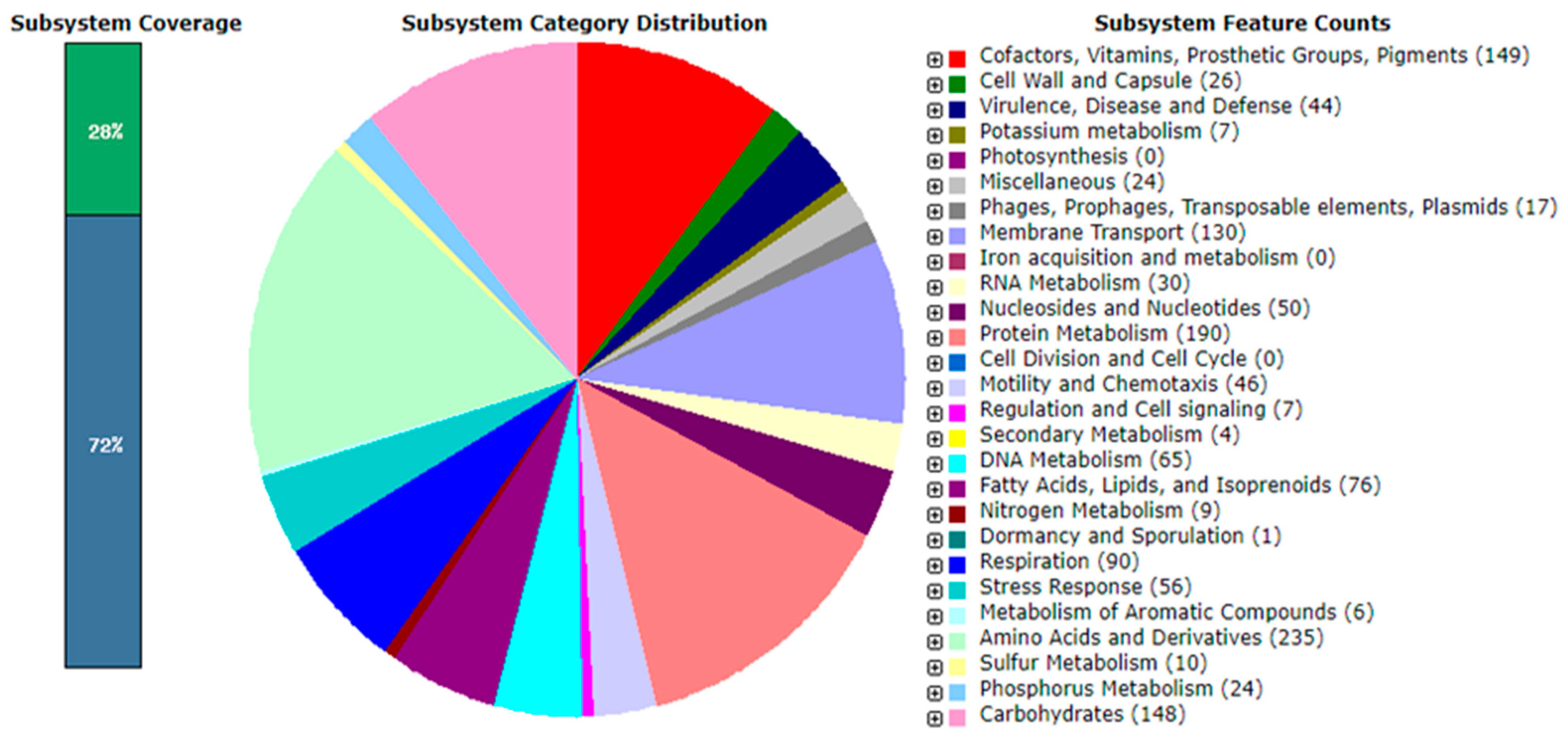
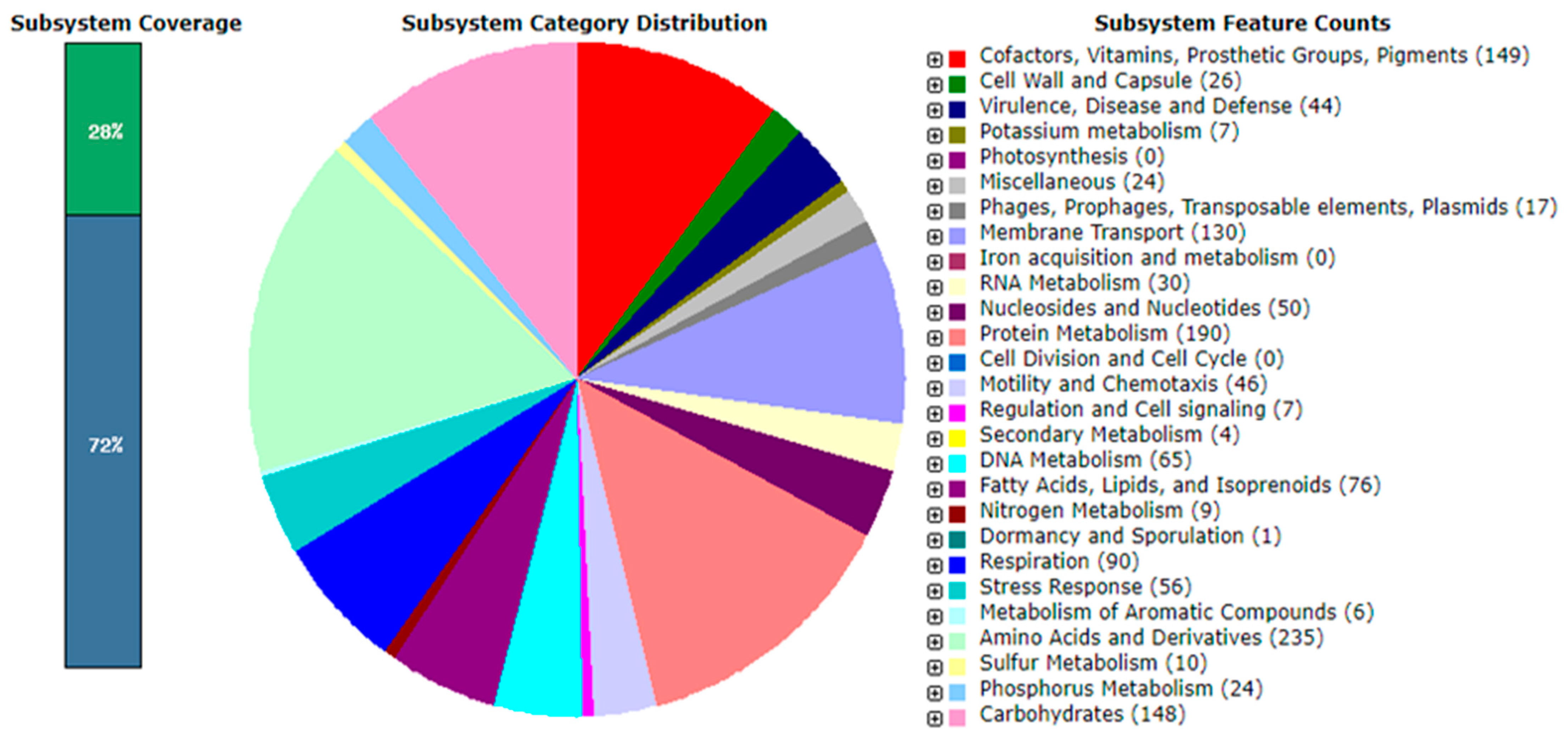
3.5. Genome Analysis
4. Description of Qipengyuania pacifica sp. nov.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Back, C.R.; Stennett, H.L.; Williams, S.E.; Wang, L.; Ojeda Gomez, J.; Abdulle, O.M.; Duffy, T.; Neal, C.; Mantell, J.; Jepson, M.A.; et al. A New Micromonospora Strain with Antibiotic Activity Isolated from the Microbiome of a Mid-Atlantic Deep-Sea Sponge. Mar. Drugs 2021, 19, 105. [Google Scholar] [CrossRef]
- Webster, N.S.; Taylor, M.W. Marine Sponges and Their Microbial Symbionts: Love and Other Relationships. Environ. Microbiol. 2012, 14, 335–346. [Google Scholar] [CrossRef]
- Schmitt, S.; Tsai, P.; Bell, J.; Fromont, J.; Ilan, M.; Lindquist, N.; Perez, T.; Rodrigo, A.; Schupp, P.J.; Vacelet, J.; et al. Assessing the Complex Sponge Microbiota: Core, Variable and Species-Specific Bacterial Communities in Marine Sponges. ISME J. 2012, 6, 564–576. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Sun, C.; Fang, C.; Oren, A.; Xu, X.W. Genomic-Based Taxonomic Classification of the Family Erythrobacteraceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 4470–4495. [Google Scholar] [CrossRef]
- Zhuang, L.; Lin, B.; Xu, L.; Li, G.; Wu, C.J.; Luo, L. Erythrobacter spongiae sp. nov., isolated from marine sponge. Int. J. Syst. Evol. Microbiol. 2019, 69, 1111–1116. [Google Scholar] [CrossRef]
- Fang, C.; Wu, Y.H.; Sun, C.; Wang, H.; Cheng, H.; Meng, F.X.; Wang, C.S.; Xu, X.W. Erythrobacter zhengii sp. nov., a bacterium isolated from deep-sea sediment. Int. J. Syst. Evol. Microbiol. 2019, 69, 241–248. [Google Scholar] [CrossRef]
- Denner, E.B.M.; Vybiral, D.; Koblízek, M.; Kämpfer, P.; Busse, H.-J.; Velimirov, B. Erythrobacter citreus sp. nov., a yellow-pigmented bacterium that lacks Bacteriochlorophyll a, isolated from the western mediterranean sea. Int. J. Syst. Evol. Microbiol. 2002, 52, 1655–1661. [Google Scholar] [CrossRef] [Green Version]
- Lei, X.; Zhang, H.; Chen, Y.; Li, Y.; Chen, Z.; Lai, Q.; Zhang, J.; Zheng, W.; Xu, H.; Zheng, T. Erythrobacter luteus sp. nov., isolated from mangrove sediment. Int. J. Syst. Evol. Microbiol. 2015, 65, 2472–2478. [Google Scholar] [CrossRef] [Green Version]
- Subhash, Y.; Tushar, L.; Sasikala, C.; Ramana, C.V.; Ch Ramana, C.V. Erythrobacter odishensis sp. nov. and Pontibacter odishensis sp. nov. isolated from dry soil of a solar saltern. Int. J. Syst. Evol. Microbiol. 2013, 63, 4524–4532. [Google Scholar] [CrossRef]
- Xing, T.; Liu, Y.; Wang, N.; Xu, B.; Liu, K.; Shen, L.; Gu, Z.; Guo, B.; Zhou, Y.; Liu, H. Erythrobacter arachoides sp. nov., isolated from ice core. Int. J. Syst. Evol. Microbiol. 2017, 67, 4235–4239. [Google Scholar] [CrossRef]
- Park, S.; Won, S.M.; Yoon, J.H. Erythrobacter marisflavi sp. nov., isolated from estuary water. Int. J. Syst. Evol. Microbiol. 2019, 69, 2696–2702. [Google Scholar] [CrossRef]
- Tonon, L.A.C.; Moreira, A.P.B.; Thompson, F. The Family Erythrobacteraceae. In The Prokaryotes: Alphaproteobacteria and Betaproteobacteria; Springer: Berlin/Heidelberg, Germany, 2014; Volume 9783642301, pp. 213–235. [Google Scholar] [CrossRef]
- Shiba, T.; Simidu, U. Erythrobacter longus gen. nov., sp. nov., an aerobic bacterium which contains Bacteriochlorophyll a. Int. J. Syst. Bacteriol. 1982, 32, 211–217. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.M.; Mo, Y.X.; Han, L.; Nogi, Y.; Zhu, Y.H.; Lv, J. Qipengyuania sediminis gen. nov., sp. nov., a Member of the Family Erythrobacteraceae Isolated from Subterrestrial Sediment. Int. J. Syst. Evol. Microbiol. 2015, 65, 3658–3665. [Google Scholar] [CrossRef]
- Parte, A.C. LPSN—List of Prokaryotic Names with Standing in Nomenclature (Bacterio.Net), 20 Years On. Int. J. Syst. Evol. Microbiol. 2018, 68, 1825–1829. [Google Scholar] [CrossRef]
- Hosoya, S.; Arunpairojana, V.; Suwannachart, C.; Kanjana-Opas, A.; Yokota, A. Aureispira marina gen. nov., sp. nov., a gliding, Arachidonic Acid-Containing Bacterium Isolated from the Southern Coastline of Thailand. Int. J. Syst. Evol. Microbiol. 2006, 56, 2931–2935. [Google Scholar] [CrossRef] [Green Version]
- Landwehr, W.; Kämpfer, P.; Glaeser, S.P.; Rückert, C.; Kalinowski, J.; Blom, J.; Goesmann, A.; Mack, M.; Schumann, P.; Atasayar, E.; et al. Taxonomic Analyses of Members of the Streptomyces cinnabarinus Cluster, Description of Streptomyces cinnabarigriseus sp. nov. and Streptomyces davaonensis sp. nov. Int. J. Syst. Evol. Microbiol. 2018, 68, 382–393. [Google Scholar] [CrossRef]
- Safaei, N.; Nouioui, I.; Mast, Y.; Zaburannyi, N.; Rohde, M.; Schumann, P.; Müller, R.; Wink, J. Kibdelosporangium persicum sp. nov., a new member of the Actinomycetes from a Hot Desert in Iran. Int. J. Syst. Evol. Microbiol. 2021, 71, 004625. [Google Scholar] [CrossRef]
- Wu, H.X.; Lai, P.Y.; Lee, O.O.; Zhou, X.J.; Miao, L.; Wang, H.; Qian, P.Y. Erythrobacter Pelagi sp. nov., a member of the Family Erythrobacteraceae isolated from the Red Sea. Int. J. Syst. Evol. Microbiol. 2012, 62, 1348–1353. [Google Scholar] [CrossRef]
- Tareen, S.; Risdian, C.; Müsken, M.; Wink, J. Alteriqipengyuania Abyssalis sp. nov., a Novel Member of the Class Alphaproteobacteria Isolated from Sponge, and Emended Description of the Genus Alteriqipengyuania. Diversity 2021, 13, 670. [Google Scholar] [CrossRef]
- Mohr, K.I.; Garcia, R.O.; Gerth, K.; Irschik, H.; Müller, R. Sandaracinus Amylolyticus gen. nov., sp. nov., a starch-degrading soil myxobacterium, and Description of Sandaracinaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2012, 62, 1191–1198. [Google Scholar] [CrossRef] [Green Version]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.P.; Göker, M. Genome Sequence-Based Species Delimitation with Confidence Intervals and Improved Distance Functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Goloboff, P.A.; Farris, J.S.; Nixon, K.C. TNT, a Free Program for Phylogenetic Analysis. Cladistics 2008, 24, 774–786. [Google Scholar] [CrossRef]
- Pattengale, N.D.; Alipour, M.; Bininda-Emonds, O.R.P.; Moret, B.M.E.; Stamatakis, A. How Many Bootstrap Replicates Are Necessary? J. Comput. Biol. 2010, 17, 337–354. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP Phylogenetic Analysis Using Parsimony (and Other Methods), Version 4.0 Beta 10. Sinauer Associates, Sunderland—References—Scientific Research Publishing. 2002. Available online: https://www.scirp.org/(S(lz5mqp453edsnp55rrgjct55))/reference/ReferencesPapers.aspx?ReferenceID=1958232 (accessed on 14 June 2021).
- Minnikin, D.E.; O’Donnell, A.G.; Goodfellow, M.; Alderson, G.; Athalye, M.; Schaal, A.; Parlett, J.H. An Integrated Procedure for the Extraction of Bacterial Isoprenoid Quinones and Polar Lipids. J. Microbiol. Methods 1984, 2, 233–241. [Google Scholar] [CrossRef]
- Goldberg, S.R.; Correa, H.; Haltli, B.A.; Kerr, R.G. Fulvivirga Aurantia sp. nov. and Xanthovirga Aplysinae gen. nov., sp. nov., Marine Bacteria Isolated from the Sponge Aplysina Fistularis, and Emended Description of the Genus Fulvivirga. Int. J. Syst. Evol. Microbiol. 2020, 70, 2766–2781. [Google Scholar] [CrossRef]
- Bóna-Lovász, J.; Bóna, A.; Ederer, M.; Sawodny, O.; Ghosh, R. A Rapid Method for the Extraction and Analysis of Carotenoids and Other Hydrophobic Substances Suitable for Systems Biology Studies with Photosynthetic Bacteria. Metabolites 2013, 3, 912–930. [Google Scholar] [CrossRef]
- Risdian, C.; Landwehr, W.; Rohde, M.; Schumann, P.; Hahnke, R.L.; Spröer, C.; Bunk, B.; Kämpfer, P.; Schupp, P.J.; Wink, J. Streptomyces Bathyalis sp. nov., an Actinobacterium Isolated from the Sponge in a Deep Sea. Antonie Van Leeuwenhoek 2021, 114, 425–435. [Google Scholar] [CrossRef]
- Sasser, M. Identification of Bacteria by Gas Chromatography of Cellular Fatty Acids. Technical Note # 101. 2001. Available online: https://www.researchgate.net/publication/284025789_Identification_of_bacteria_by_gas_chromatography_of_cellular_fatty_acids (accessed on 29 February 2022).
- Khosravi Babadi, Z.; Ebrahimipour, G.; Wink, J.; Narmani, A.; Risdian, C. Isolation and Identification of Streptomyces sp. Act4Zk, a Good Producer of Staurosporine and Some Derivatives. Lett. Appl. Microbiol. 2021, 72, 206–218. [Google Scholar] [CrossRef]
- Khodamoradi, S.; Hahnke, R.L.; Mast, Y.; Schumann, P.; Kämpfer, P.; Steinert, M.; Rückert, C.; Surup, F.; Rohde, M.; Wink, J. Streptomonospora litoralis sp. nov., a halophilic thiopeptides producer isolated from Sand Collected at Cuxhaven Beach. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2021, 114, 1483–1496. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving Bacterial Genome Assemblies from Short and Long Sequencing Reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Lee, I.; Chalita, M.; Ha, S.M.; Na, S.I.; Yoon, S.H.; Chun, J. ContEst16S: An Algorithm That Identifies Contaminated Prokaryotic Genomes Using 16S RNA Gene Sequences. Int. J. Syst. Evol. Microbiol. 2017, 67, 2053–2057. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid Annotations Using Subsystems Technology. BMC Genom. 2008, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. AntiSMASH 5.0: Updates to the Secondary Metabolite Genome Mining Pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef] [Green Version]
- Meier-Kolthoff, J.P.; Göker, M.; Spröer, C.; Klenk, H.P. When Should a DDH Experiment Be Mandatory in Microbial Taxonomy? Arch. Microbiol. 2013, 195, 413–418. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kosako, Y.; Yabuuchi, E.; Naka, T.; Fujiwara, N.; Kobayashi, K. Proposal of Sphingomonadaceae fam. Nov., Consisting of Sphingomonas Yabuuchi et Al. 1990, Erythrobacter Shiba and Shimidu 1982, Erythromicrobium Yurkov et Al. 1994, Porphyrobacter Fuerst et Al. 1993, Zymomonas Kluyver and van Niel 1936, and Sandaracinobac. Microbiol. Immunol. 2000, 44, 563–575. [Google Scholar] [CrossRef]
- Lefort, V.; Desper, R.; Gascuel, O. FastME 2.0: A Comprehensive, Accurate, and Fast Distance-Based Phylogeny Inference Program. Mol. Biol. Evol. 2015, 32, 2798. [Google Scholar] [CrossRef] [Green Version]
- Tang, T.; Sun, X.; Dong, Y.; Liu, Q. Erythrobacter aureus sp. nov., a plant growth-promoting bacterium isolated from sediment in the Yellow Sea, China. 3 Biotech 2019, 9, 430. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acids | 1 | 2 | 3 | 4 |
|---|---|---|---|---|
| Saturated | ||||
| C15:0 | 1.57 | − | − | 5.76 |
| C16:0 | 11.98 | 3.32 | 4.21 | 5.11 |
| C17:0 | 1.91 | 1.93 | − | 4.63 |
| C18:0 | 1.21 | − | − | − |
| C19:0 | − | 1.63 | − | − |
| Branched | ||||
| C17:0a | 0.77 | 0.78 | − | 3.97 |
| Unsaturated | ||||
| C17:1ω6c | 9.54 | 17.76 | 7.66 | 34.69 |
| C18:1ω12t | 40.29 | − | 55.06 | − |
| C16:1ω7c | 9.93 | 6.75 | 10.31 | 3.04 |
| C16:1ω7t | 0.72 | 1.12 | 1.14 | − |
| C17:1ω8c | − | 1.69 | − | − |
| C18:1ω7c | − | − | − | 20.92 |
| Hydroxy | ||||
| C14:02-OH | 5.09 | 3.23 | 5.89 | 4.25 |
| C16:02-OH | 5.89 | 3.77 | 6.36 | − |
| C18:1ω9t and/or C18:1ω7c | − | 40.52 | − | − |
| Reference Type Strains | 16S | ANI | dDDH |
|---|---|---|---|
| Qipengyuania citreus RE35F/1T | 98.80 | 79.38 | 22.10 |
| Erythrobacter aureus YH-07T | 98.38 | 78.93 | 21.40 |
| Qipengyuania pelagi UST081027-248T | 98.31 | 75.43 | 21.40 |
| Qipengyuania seohaensis SW-135T | 98.02 | 75.81 | 19.40 |
| Qipengyuania vulgaris 022 2-10T | 98.15 | 77.31 | 23.50 |
| Sphingomonas phyllosphaerae FA2T | 97.75 | 70.00 | 20.20 |
| Qipengyuania aquimaris SW-110T | 97.74 | 75.68 | 19.60 |
| Qipengyuania gaetbuli SW-161T | 97.66 | 76.43 | 19.70 |
| Erythrobacter litoralis DSM 8509T | 97.22 | 72.61 | 18.80 |
| Properties | 1 | 2 | 3 | 4 |
|---|---|---|---|---|
| Motility | + | − | − | + |
| Esterase (C 4) | w+ | w+ | + | + |
| Esterase lipase (C 8) | w+ | + | + | + |
| Cystine arylamidase | + | w+ | w+ | w+ |
| Trypsin | + | w+ | + | + |
| α-chymotrypsin | w+ | + | − | + |
| Naphthol-AS-BI-phosphohydrolase | + | w+ | + | + |
| α-glucosidase | + | + | − | − |
| lysine decarboxylase | w+ | − | − | − |
| Citrate utilization | w+ | − | − | − |
| Tryptophan deaminase | w+ | + | + | + |
| Gelatinase | − | w+ | − | − |
| Nitrate reduction | − | + | + | + |
| Arginine dihydrolase | + | − | − | − |
| Inositol fermentation | + | − | − | − |
| L-Alanine | − | + | + | w+ |
| L-Galactonic acid lactone | + | w+ | + | w+ |
| L-Lactic acid | + | − | w+ | − |
| D-Salicin | + | − | − | w+ |
| Citric acid | − | + | + | − |
| L-Histidine | − | − | w+ | + |
| L-pyroglutamic acid | + | w+ | + | + |
| Stachyose | − | − | w+ | + |
| N-acetyl Neuraminic acid | + | − | w+ | + |
| D-serine | − | + | − | + |
| Polar lipids | PC, SGL, PG, DPG, 3GL, 1L. | * DPG, PC, PE, PG, 2L, 1P. | ** PE, PG, DPG, PC, SGL. | *** DPG, PC, PE, PG, SGL, 2L. |
| Fatty acids | ||||
| C16:0 | 11.98% | 5.11% | 3.32% | 4.21% |
| C18:1ω12t | 40.29% | ND | ND | 55.06 |
| C16:02-OH | 5.89 | ND | 3.77% | 6.36% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tareen, S.; Risdian, C.; Müsken, M.; Wink, J. Qipengyuania pacifica sp. nov., a Novel Carotenoid-Producing Marine Bacterium of the Family Erythrobacteraceae, Isolated from Sponge (Demospongiae), and Antimicrobial Potential of Its Crude Extract. Diversity 2022, 14, 295. https://doi.org/10.3390/d14040295
Tareen S, Risdian C, Müsken M, Wink J. Qipengyuania pacifica sp. nov., a Novel Carotenoid-Producing Marine Bacterium of the Family Erythrobacteraceae, Isolated from Sponge (Demospongiae), and Antimicrobial Potential of Its Crude Extract. Diversity. 2022; 14(4):295. https://doi.org/10.3390/d14040295
Chicago/Turabian StyleTareen, Sanaullah, Chandra Risdian, Mathias Müsken, and Joachim Wink. 2022. "Qipengyuania pacifica sp. nov., a Novel Carotenoid-Producing Marine Bacterium of the Family Erythrobacteraceae, Isolated from Sponge (Demospongiae), and Antimicrobial Potential of Its Crude Extract" Diversity 14, no. 4: 295. https://doi.org/10.3390/d14040295
APA StyleTareen, S., Risdian, C., Müsken, M., & Wink, J. (2022). Qipengyuania pacifica sp. nov., a Novel Carotenoid-Producing Marine Bacterium of the Family Erythrobacteraceae, Isolated from Sponge (Demospongiae), and Antimicrobial Potential of Its Crude Extract. Diversity, 14(4), 295. https://doi.org/10.3390/d14040295