
New Records of Powdery Mildews from Taiwan: Erysiphe ipomoeae comb. nov., E. aff. betae on Buckwheat, and E. neolycopersici comb. nov. on Cardiospermum halicacabum




Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Morphology
2.2. Molecular Phylogeny
2.3. Pathogenicity Tests
3. Results
3.1. Morphology
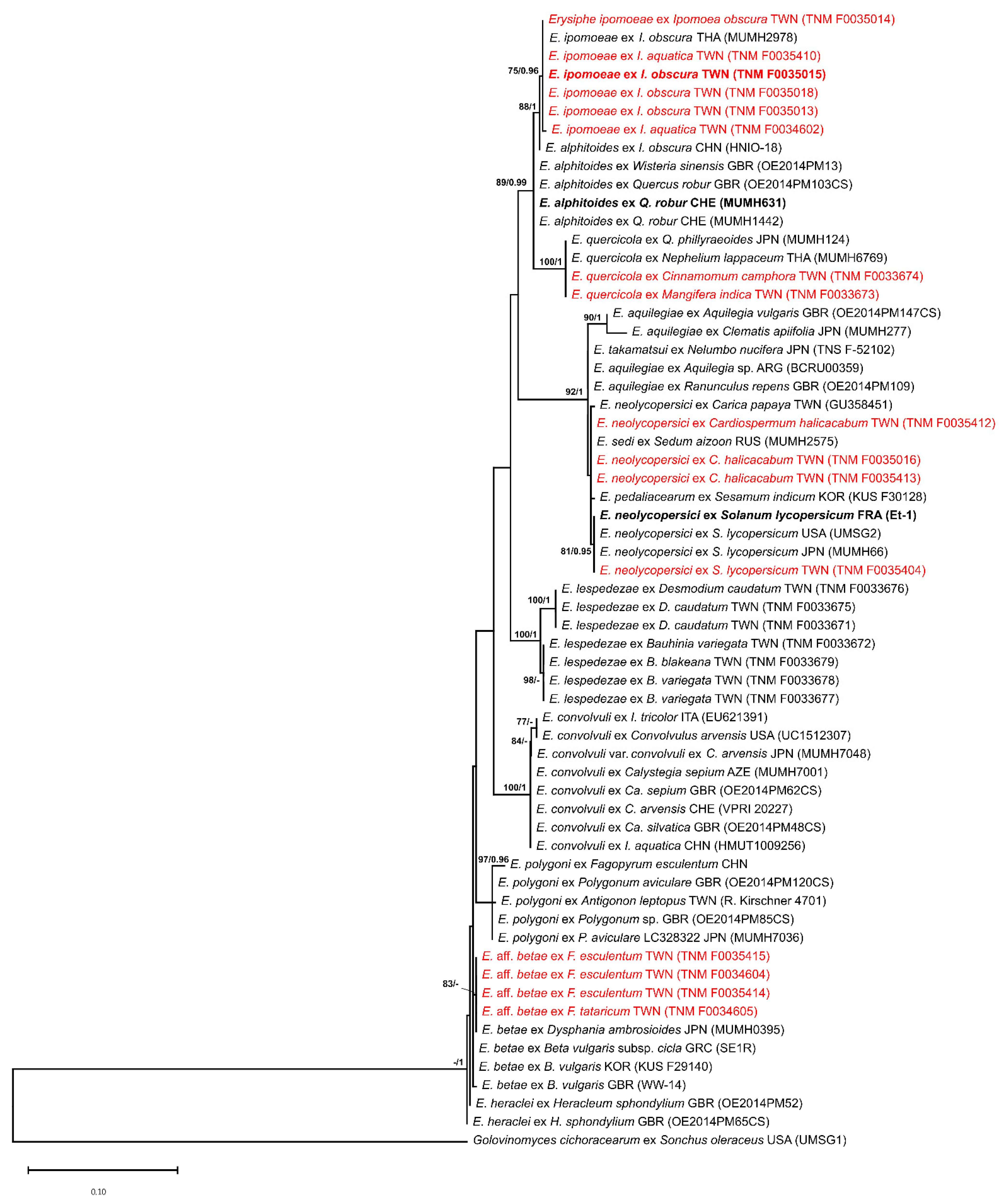
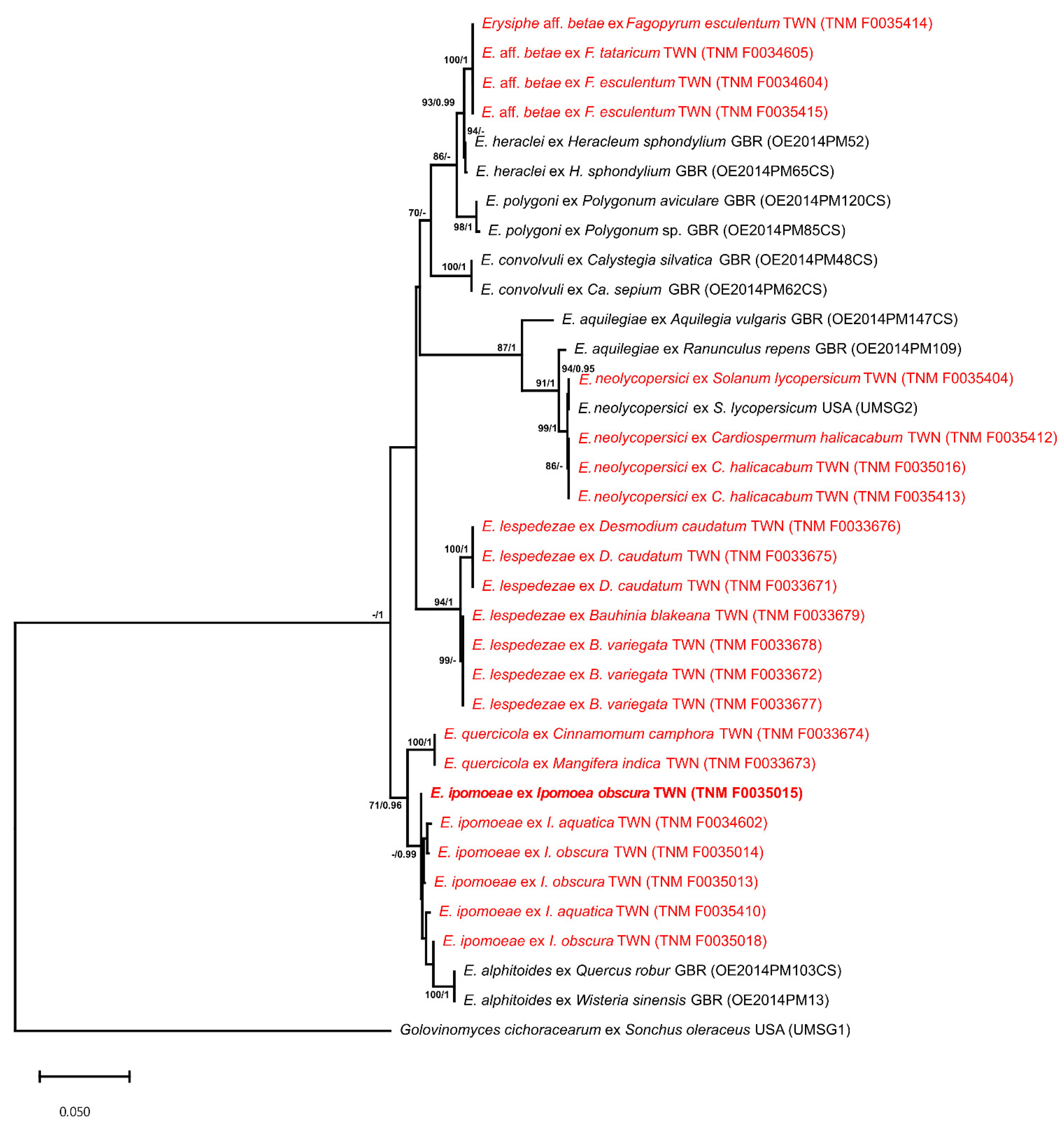
3.2. Phylogentic Analyses
3.3. Pathogenicity Tests
3.4. Taxonomy
4. Discussion
4.1. Erysiphe ipomoeae and E. convolvuli, the Two Distinct Species Causing Powdery Mildews on Convolvulaceae
4.2. Powdery Mildews on Buckwheat and the Host Range of Erysiphe betae
4.3. Erysiphe aquilegiae Complex Needs Further Exploration
4.4. Perennation of Powdery Mildews in the Subtropics and the Impact on Crop Production
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johnston, P.R.; Quijada, L.; Smith, C.A.; Baral, H.-O.; Hosoya, T.; Baschien, C.; Pärtel, K.; Zhuang, W.Y.; Haelewaters, D.; Park, D.; et al. A multigene phylogeny toward a new phylogenetic classification of Leotiomycetes. IMA Fungus 2019, 10, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Braun, U.; Cook, R.T.A. Taxonomic Manual of the Erysiphales (Powdery Mildews); CBS-KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2012; pp. 1–707. [Google Scholar]
- Takamatsu, S. Studies on the evolution and systematics of powdery mildew fungi. J. Gen. Plant Pathol. 2018, 84, 422–426. [Google Scholar] [CrossRef]
- Kiss, L.; Vaghefi, N.; Bransgrove, K.; Dearnaley, J.D.W.; Takamatsu, S.; Tan, Y.P.; Marston, C.; Liu, S.Y.; Jin, D.N.; Adorada, D.L.; et al. Australia: A continent without native powdery mildews? The first comprehensive catalog indicates recent introductions and multiple host range expansion events, and leads to the re-discovery of Salmonomyces as a new lineage of the Erysiphales. Front. Microbiol. 2020, 11, 1571. [Google Scholar] [CrossRef] [PubMed]
- Hawksworth, D.L. The magnitude of fungal diversity: The 1.5 million species estimate revisited. Mycol. Res. 2001, 105, 1422–1432. [Google Scholar] [CrossRef] [Green Version]
- Amano, K. Host Range and Geographical Distribution of the Powdery Mildew Fungi; Japan Science Society Press: Tokyo, Japan, 1986; pp. 1–741. [Google Scholar]
- Nasir, M.; Mughal, S.M.; Mukhtar, T.; Awan, M.Z. Powdery mildew of mango: A review of ecology, biology, epidemiology and management. Crop Protect. 2014, 64, 19–26. [Google Scholar] [CrossRef]
- Mieslerová, B.; Kitner, M.; Křístková, E.; Majeský, L.; Lebeda, A. Powdery mildews on Lactuca species—A complex view of host-pathogen interactions. Crit. Rev. Plant Sci. 2020, 39, 44–71. [Google Scholar] [CrossRef]
- Takamatsu, S.; Ito, H.; Shiroya, Y.; Kiss, L.; Heluta, V. First comprehensive phylogenetic analysis of the genus Erysiphe (Erysiphales, Erysiphaceae) I. The Microsphaera lineage. Mycologia 2015, 107, 475–489. [Google Scholar] [CrossRef]
- Takamatsu, S.; Ito, H.; Shiroya, Y.; Kiss, L.; Heluta, V. First comprehensive phylogenetic analysis of the genus Erysiphe (Erysiphales, Erysiphaceae) II: The Uncinula lineage. Mycologia 2015, 107, 903–914. [Google Scholar] [CrossRef] [Green Version]
- Abasova, L.V.; Aghayeva, D.N.; Takamatsu, S. Notes on powdery mildews of the genus Erysiphe from Azerbaijan. Curr. Res. Environm. Appl. Mycol. 2018, 8, 30–53. [Google Scholar] [CrossRef]
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.-H.; Li, D.-Z.; Marhold, K.; et al. International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code) Adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017; Koeltz Botanical Books: Glashütten, Germany, 2018. [Google Scholar]
- Sawada, K. Formosa Agricultural Experiment Station Special Report No. 9; Taiwan Sōtokufu Nōji Shikenjō: Taihoku, Formosa, 1914. [Google Scholar]
- Sawada, K. Descriptive Catalogue of Taiwan (Formosan) Fungi. Part I; College of Agriculture National Taiwan University: Taipei, Taiwan, 1919. [Google Scholar]
- Kou, K.C. Checklist of Erysiphaceae in Taiwan. Fung. Sci. 1998, 13, 39–59. [Google Scholar]
- Tzean, S.S.; Tzeng, K.C.; Chang, C.A.; Tsay, T.T.; Yen, S.F. List of Plant Diseases in Taiwan, 5th ed.; Taiwan Phytopathological Society: Taipei, Taiwan, 2019; pp. 1–329. [Google Scholar]
- Kirschner, R.; Liu, W. Two new hosts of anamorphic Erysiphe quercicola: Cinnamomum camphora and Murraya paniculata. Mycoscience 2014, 55, 190–195. [Google Scholar] [CrossRef]
- Kirschner, R. First record of the sexual stage of the powdery mildew Erysiphe pseudolonicerae in Taiwan. Fung. Sci. 2015, 30, 61–66. [Google Scholar]
- Wang, C.T.; Yeh, Y.W.; Lin, L.D.; Kirschner, R. First record of the powdery mildew fungus Erysiphe polygoni on the introduced coral vine Antigonon leptopus (Polygonaceae) in Taiwan. Plant Pathol. Quar. 2019, 9, 160–165. [Google Scholar] [CrossRef]
- Wang, C.T.; Yeh, Y.W.; Lin, L.D.; Kirschner, R. First record of Erysiphe magnifica on the new host Magnolia× Alba in Taiwan indicates high morphological plasticity of the anamorph under tropical conditions. Plant Pathol. Quar. 2020, 10, 59–65. [Google Scholar] [CrossRef]
- Wang, C.T.; Yeh, Y.W.; Lin, L.D.; Kirschner, R. First record of the powdery mildew fungi of mulberry and plane trees, Erysiphe mori and E. Platani, in Taiwan. J. Exp. For. Nat. Taiwan Univ. 2021, 35, 39–48. [Google Scholar]
- Kirschner, R.; Lotz-Winter, H.; Piepenbring, M. A new species of the elm powdery mildew species complex (Erysiphaceae) on Chinese elm (Ulmus parvifolia) in East Asia segregated from Erysiphe ulmi. Phytotaxa 2020, 447, 276–282. [Google Scholar] [CrossRef]
- Yeh, Y.W.; Chou, P.Y.; Hou, H.Y.; Kirschner, R. First records of powdery mildew fungi (Erysiphales) on medicinal plants in Taiwan. Bot. Stud. 2021, 62, 1. [Google Scholar] [CrossRef]
- Xiao, Y.T.; Wang, C.J.; Huang, T.C.; Shen, Y.M. Erysiphe lespedezae causing powdery mildew on Bauhinia variegata, B. blakeana and Desmodium caudatum in Taiwan. For. Pathol. 2020, 50, e12631. [Google Scholar] [CrossRef]
- Cunnington, J.H.; Takamatsu, S.; Lawrie, A.C.; Pascoe, I.G. Molecular identification of anamorphic powdery mildews (Erysiphales). Australas. Plant Pathol. 2003, 32, 421–428. [Google Scholar] [CrossRef]
- Ellingham, O.; David, J.; Culham, A. Enhancing identification accuracy for powdery mildews using previously underexploited DNA loci. Mycologia 2019, 111, 798–812. [Google Scholar] [CrossRef]
- Bolay, A.; Clerc, P.; Braun, U.; Götz, M.; Takamatsu, S. New species, new records and first sequence data of powdery mildews (Erysiphaceae) from Europe with special emphasis on Switzerland. Austrian J. Mycol. 2021, 28, 131–160. [Google Scholar]
- Yen, J.M.; Wang, C.C. Étude sur les champignons parasites du Sud-Est asiatique XXII: Les Oidium de Formose (II). Rev. Mycol. 1973, 37, 125–153. [Google Scholar]
- Meeboon, J.; Takamatsu, S. Notes on powdery mildews (Erysiphales) in Thailand II. Erysiphe species on Adoxaceae Anacardiaceae, Apocynaceae, Araliaceae, Aristolochiaceae, Bixaceae, Brassicaceae, Cleomaceae, Convolvulaceae, Cucurbitaceae and Euphorbiacea. Trop. Plant Pathol. 2016, 41, 357–369. [Google Scholar]
- Lee, L.C.; Hou, Y.C.; Hsieh, Y.Y.; Chen, Y.H.; Shen, Y.C.; Lee, I.J.; Shih, M.-C.M.; Hou, W.C.; Liu, H.K. Dietary supplementation of rutin and rutin-rich buckwheat elevates endogenous glucagon-like peptide 1 levels to facilitate glycemic control in type 2 diabetic mice. J. Funct. Foods 2021, 85, 104653. [Google Scholar] [CrossRef]
- Takamatsu, S.; Kano, Y. PCR primers useful for nucleotide sequencing of rDNA of the powdery mildew fungi. Mycoscience 2001, 42, 135–139. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics; Academy Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Mori, Y.; Sato, Y.; Takamatsu, S. Evolutionary analysis of the powdery mildew fungi using nucleotide sequences of the nuclear ribosomal DNA. Mycologia 2000, 92, 74–93. [Google Scholar] [CrossRef]
- Pan, Y.W.; Wu, H.; Di, R. First report of powdery mildew caused by Erysiphe alphitoides on Ipomoea obscura in China. Plant Dis. 2020, 104, 1870. [Google Scholar] [CrossRef]
- Takamatsu, S.; Limkaisang, S.; Kom-un, S.; Bolay, A.; To-anun, C. Identity of a powdery mildew fungus occurring on Paeonia and its relationship with Erysiphe hypophylla on oak. Mycoscience 2006, 47, 367–373. [Google Scholar] [CrossRef]
- Takamatsu, S.; Braun, U.; Limkaisang, S.; Kom-Un, S.; Sato, Y.; Cunnington, J.H. Phylogeny and taxonomy of the oak powdery mildew Erysiphe alphitoides sensu lato. Mycol. Res. 2007, 111, 809–826. [Google Scholar] [CrossRef]
- Joa, J.H.; Seong, K.C.; Choi, I.Y.; Cho, S.E.; Shin, H.D. First report of powdery mildew caused by Erysiphe betae on sugar beet in Korea. Plant Dis. 2017, 101, 254. [Google Scholar] [CrossRef]
- Francis, S.A.; Roden, B.C.; Adams, M.J.; Weiland, J.; Asher, M.J. Comparison of ITS sequences from UK and North American sugar-beet powdery mildews and the designation of Erysiphe betae. Mycol. Res. 2007, 111, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Vakalounakis, D.J.; Kavroulakis, N. Occurrence of powdery mildew caused by Erysiphe betae on chard [Beta vulgaris L. subsp. cicla (L.) Koch] in Greece. Crop Protect. 2017, 99, 128–131. [Google Scholar] [CrossRef]
- Saenz, G.S.; Taylor, J.W. Phylogeny of the Erysiphales (powdery mildews) inferred from internal transcribed spacer ribosomal DNA sequences. Can. J. Bot. 1999, 77, 150–168. [Google Scholar]
- Tsay, J.G.; Chen, R.S.; Wang, H.L.; Wang, W.L.; Weng, B.C. First report of powdery mildew caused by Erysiphe diffusa, Oidium neolycopersici, and Podosphaera xanthii on papaya in Taiwan. Plant Dis. 2011, 95, 1188. [Google Scholar] [CrossRef] [PubMed]
- Kiss, L.; Cook, R.T.A.; Saenz, G.S.; Cunnington, J.H.; Takamatsu, S.; Pascoe, I.; Bardin, M.; Nicot, P.C.; Sato, Y.; Rossman, A.Y. Identification of two powdery mildew fungi, Oidium neolycopersici sp. nov. and O. lycopersici, infecting tomato in different parts of the world. Mycol. Res. 2001, 105, 684–697. [Google Scholar] [CrossRef]
- Wu, Y.; Ma, X.F.; Pan, Z.Y.; Kale, S.D.; Song, Y.; King, H.; Zhang, Q.; Presley, C.; Deng, X.X.; Wei, C.I. Comparative genome analyses reveal sequence features reflecting distinct modes of host-adaptation between dicot and monocot powdery mildew. BMC Genom. 2018, 19, 705. [Google Scholar] [CrossRef]
- Shin, H.D.; Meeboon, J.; Takamatsu, S.; Adhikari, M.K.; Braun, U. Phylogeny and taxonomy of Pseudoidium pedaliacearum. Mycol. Prog. 2019, 18, 237–246. [Google Scholar] [CrossRef]
- Lu, W.J.; Wang, L.H.; Wang, Y.Q.; Li, C.H. First report of powdery mildew caused by Erysiphe polygoni on buckwheat in Yunnan, China. Plant Dis. 2015, 99, 1281. [Google Scholar] [CrossRef]
- Limkaisang, S.; Kom-un, S.; Furtado, E.L.; Liew, K.W.; Salleh, B.; Sato, Y.; Takamatsu, S. Molecular phylogenetic and morphological analyses of Oidium heveae, a powdery mildew of rubber tree. Mycoscience 2005, 46, 220–226. [Google Scholar] [CrossRef]
- Meeboon, J.; Takamatsu, S. Hosts of asexual morph of Erysiphe quercicola from Thailand. Trop. Plant Pathol. 2020, 45, 122–135. [Google Scholar] [CrossRef]
- Meeboon, J.; Takamatsu, S. Erysiphe takamatsui, a powdery mildew of lotus: Rediscovery of teleomorph after 40 years, morphology and phylogeny. Mycoscience 2015, 56, 159–167. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edler, D.; Klein, J.; Antonelli, A.; Silvestro, D. raxmlGUI 2.0: A graphical interface and toolkit for phylogenetic analyses using RAxML. Methods Ecol. Evol. 2021, 12, 373–377. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolay, A. Les Oïdiums de Suisse (Erysiphacées); Cryptogamica Helvtica: Birmensdorf, Switzerland, 2005; pp. 1–176. [Google Scholar]
- Thite, S.V.; Kore, B.A. Diversity of powdery mildew fungi from Satara district (M.S.). Int. J. Multidiscip. Res. 2013, 2, 119–122. [Google Scholar]
- Braun, U. Taxonomic notes on some powdery mildews of various genera. Mycotaxon 1986, 25, 259–269. [Google Scholar]
- Sawada, K. Descriptive Catalogue of Taiwan (Formosan) Fungi. Part XI; College of Agriculture National Taiwan University: Taipei, Taiwan, 1959; pp. 1–268. [Google Scholar]
- Delmail, D.; Autret, J.-L. First description of Oidium neolycopersici (Erysiphaceae) in France, on a new host plant extinct in the wild. Mycotaxon 2010, 113, 269–271. [Google Scholar] [CrossRef]
- Hidayat, I.; Meeboon, J.; Takamatsu, S. First report of Pseudoidium aff. neolycopersici in Indonesia. Australas. Plant Dis. Notes 2014, 9, 139. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.Y.; Liu, S.Y.; Jiang, W.T.; Li, Y. First report of powdery mildew caused by Pseudoidium cf. neolycopersici on Physalis pubescens in China. Plant Dis. 2015, 99, 1862. [Google Scholar]
- Brielmaier-Liebetanz, U.; Field, A.E.; Warfield, C.Y.; Braun, U. Powdery mildew (Erysiphaceae) on Calibrachoa hybrids in Germany, Nicaragua and the USA. Plant Pathol. Quar. 2015, 5, 1–5. [Google Scholar] [CrossRef]
- Mukhtar, I.; van Peer, A.F. Occurrence of powdery mildew caused by Pseudoidium neolycopersici on papaya (Carica papaya) in China. Plant Dis. 2018, 102, 2645. [Google Scholar] [CrossRef]
- Liu, X.M.; Wei, Y.X.; Zhang, H.; Zhou, F.X.; Pu, J.J. First report of powdery mildew (Pseudoidium neolycopersici) on Croton (Codiaeum variegatum var. pictum) in China. Plant Dis. 2015, 99, 288. [Google Scholar] [CrossRef]
- Xu, D.D.; Huang, Z.H.; Zhang, X.M.; Li, M.H.; Huang, H.Q.; Zeng, Q.Q. First report of powdery mildew of Excoecaria cochinchinensis caused by Pseudoidium neolycopersici in China. Plant Dis. 2020, 104, 594. [Google Scholar] [CrossRef]
- Liu, Y.X.; Cai, Z.Y.; Yang, Y.; Long, J.M.; Li, H.Q.; Zhang, Z.B. First report of powdery mildew caused by Pseudoidium neolycopersici on Moringa oleifera in China. Plant Dis. 2019, 103, 1768. [Google Scholar] [CrossRef]
- Mukhtar, I.; Ashraf, H.J.; Khokhar, I.; van Peer, A.F.; Huang, Q.; Chen, B.; Yan, J.; Ashraf, A.; Chou, T.; Jiang, S. First report of powdery mildew caused by Pseudoidium neolycopersici (Erysiphe aquilegiae complex) on Senna bicapsularis in China. Plant Dis. 2019, 103, 2479. [Google Scholar] [CrossRef]
- Farr, D.F.; Rossman, A.Y. Fungal Databases, U.S. National Fungus Collections, ARS, USDA. Available online: https://nt.ars-grin.gov/fungaldatabases/ (accessed on 8 July 2021).
- Takamatsu, S.; Shiroya, Y.; Seko, Y. Geographical and spatial distributions of two Erysiphe species occurring on lilacs (Syringa spp.). Mycoscience 2016, 57, 349–355. [Google Scholar] [CrossRef]
- Bradshaw, M.; Braun, U.; Götz, M.; Takamatsu, S.; Brand, T.; Cabrera, M.G.; Dirchwolf, P.; Kummer, V.; Medina, R.; Moparthi, S.; et al. Contributions to the knowledge of the phylogeny and taxonomy of the Erysiphaceae (powdery mildews)—Part 1. Sydowia 2021, 73, 89–112. [Google Scholar]
- Thaung, M.M. Powdery mildews in Burma with reference to their global host-fungus distributions and taxonomic comparisons. Australas. Plant Pathol. 2007, 36, 543–551. [Google Scholar] [CrossRef]
- Hong, S.H.; Lee, Y.H.; Choi, Y.J.; Shin, H.D. First report of powdery mildew caused by Erysiphe convolvuli on Calystegia hederacea in Korea. J. Plant Pathol. 2021, 103, 351. [Google Scholar] [CrossRef]
- Sert, H.B.; Sümbül, H.; Sterflinger, K. Occurrence of powdery mildews on new hosts in Turkey. Phytoparasitica 2006, 34, 474–476. [Google Scholar] [CrossRef]
- Braun, U.; Kiehr, M.; Delhey, R. Some new records of powdery mildew fungi from Argentina. Sydowia 2000, 53, 34–43. [Google Scholar]
- Siahaan, S.A.S.; Hidayat, I.; Kramadibrata, K.; Meeboon, J.; Takamatsu, S. Bauhinia purpurea, Durio zibethinus, and Nephelium lappaceum: Additional hosts of the asexual morph of Erysiphe quercicola. Mycoscience 2016, 57, 375–383. [Google Scholar] [CrossRef]
- Xu, B.; Braun, U.; Zhang, S.; Yang, H.; Cao, Z.; Guo, Y.; Song, J. Powdery mildew caused by Erysiphe lespedezae (including Pseudoidium caesalpiniacearum, syn. nov.) on Bauhinia blakeana and B. purpurea in China. Phytotaxa 2018, 345, 35–42. [Google Scholar] [CrossRef]
- Desprez-Loustau, M.-L.; Massot, M.; Feau, N.; Fort, T.; de Vicente, A.; Torés, J.A.; Ortuño, D.F. Further support of conspecificity of oak and mango powdery mildew and first report of Erysiphe quercicola and Erysiphe alphitoides on mango in mainland Europe. Plant Dis. 2017, 101, 1086–1093. [Google Scholar] [CrossRef] [Green Version]
- Ajitomi, A.; Takushi, T.; Sato, Y.; Arasaki, C.; Ooshiro, A. First report of powdery mildew of mango caused by Erysiphe quercicola in Japan. J. Gen. Plant Pathol. 2020, 86, 316–321. [Google Scholar] [CrossRef]
- Desprez-Loustau, M.-L.; Massot, M.; Toïgo, M.; Fort, T.; Kaya, A.G.A.; Boberg, J.; Braun, U.; Capdevielle, X.; Cech, T.; Chandelier, A.; et al. From leaf to continent: The multi-scale distribution of an invasive cryptic pathogen complex on oak. Fungal Ecol. 2018, 36, 39–50. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, F.L.; Cao, P.; Liu, Y.; Liu, K.; Xu, K.D.; Liu, K.; Zhang, J.; Tan, G.X.; Li, C.W. First report of powdery mildew caused by Erysiphe betae on Swiss chard in China. Plant Dis. 2014, 98, 1429. [Google Scholar] [CrossRef]
- Anwar, A.; Afshan, N.U.S.; Ishaq, A.; Riaz, M.; Khalid, A.N.; Uddin, S. First Pakistani report of Erysiphe betae on the invasive weed Chenopodium ambrosioides. Mycotaxon 2020, 135, 649–655. [Google Scholar] [CrossRef]
- Gautam, A.K.; Avasthi, S. Diversity of powdery mildew fungi from North Western Himalayan region of Himachal Pradesh-a checklist. Plant Pathol. Quar. 2018, 8, 78–99. [Google Scholar] [CrossRef]
- Pande, A. Ascomycetes of Peninsular India; Scientific Publishers: Jodhpur, India, 2008. [Google Scholar]
- Piepenbring, M. Checklist of fungi in Panama. Puente Biol. 2006, 1, 1–96. [Google Scholar]
- Morris, C.E.; Moury, B. Revisiting the concept of host range of plant pathogens. Annu. Rev. Phytopathol. 2019, 57, 63–90. [Google Scholar] [CrossRef]
- Whipps, J.M.; Budge, S.P.; Fenlon, J.S. Characteristics and host range of tomato powdery mildew. Plant Pathol. 1998, 47, 36–48. [Google Scholar] [CrossRef]
- Jankovics, T.; Bai, Y.; Kovács, G.M.; Bardin, M.; Nicot, P.C.; Toyoda, H.; Matsuda, Y.; Niks, R.E.; Kiss, L. Oidium neolycopersici: Intraspecific variability inferred from amplified fragment length polymorphism analysis and relationship with closely related powdery mildew fungi infecting various plant species. Phytopathology 2008, 98, 529–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wadl, P.A.; Mack, B.M.; Beltz, S.B.; Moore, G.G.; Baird, R.E.; Rinehart, T.A.; Molnar, T.J.; Staton, M.E.; Hadziabdic, D.; Trigiano, R.N. Development of genomic resources for the powdery mildew, Erysiphe pulchra. Plant Dis. 2019, 103, 804–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polonio, Á.; Díaz-Martínez, L.; Fernández-Ortuño, D.; de Vicente, A.; Romero, D.; López-Ruiz, F.J.; Pérez-García, A. A hybrid genome assembly resource for Podosphaera xanthii, the main causal agent of powdery mildew disease in cucurbits. Mol. Plant Microbe Interact. 2021, 34, 319–324. [Google Scholar] [CrossRef]
- Glawe, D.A. The powdery mildews: A review of the world’s most familiar (yet poorly known) plant pathogens. Annu. Rev. Phytopathol. 2008, 46, 27–51. [Google Scholar] [CrossRef]
- Gent, D.H.; Claassen, B.J.; Twomey, M.C.; Wolfenbarger, S.N.; Woods, J.L. Susceptibility of hop crown buds to powdery mildew and its relation to perennation of Podosphaera macularis. Plant Dis. 2018, 102, 1316–1325. [Google Scholar] [CrossRef] [Green Version]
- Gent, D.H.; Mahaffee, W.F.; Turechek, W.W.; Ocamb, C.M.; Twomey, M.C.; Woods, J.L.; Probst, C. Risk factors for bud perennation of Podosphaera macularis on hop. Phytopathology 2019, 109, 74–83. [Google Scholar] [CrossRef] [Green Version]
- Pirondi, A.; Pérez-García, A.; Battistini, G.; Muzzi, E.; Brunelli, A.; Collina, M. Seasonal variations in the occurrence of Golovinomyces orontii and Podosphaera xanthii, causal agents of cucurbit powdery mildew in Northern Italy. Ann. Appl. Biol. 2015, 167, 298–313. [Google Scholar] [CrossRef]
- Liu, N.; Gong, G.; Zhang, M.; Zhou, Y.; Chen, Z.; Yang, J.; Chen, H.; Wang, X.; Lei, Y.; Liu, K. Over-summering of wheat powdery mildew in Sichuan Province, China. Crop Protect. 2012, 34, 112–118. [Google Scholar] [CrossRef]
- Yeh, Y.W.; Wu, T.Y.; Wen, H.L.; Jair, H.W.; Lee, M.Z.; Kirschner, R. Host plants of the powdery mildew fungus Podosphaera xanthii in Taiwan. Trop. Plant Pathol. 2020, 46, 44–61. [Google Scholar] [CrossRef]
- Austin, D.F. Water spinach (Ipomoea aquatica, Convolvulaceae): A food gone wild. Ethnobot. Res. Appl. 2007, 5, 123–146. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.W. Studies on the reasonable application of fertilizers for the cultivation of Ipomoea Aquatica. Tainan Dares 2016, 67, 38–48. [Google Scholar]
- Cook, R.T.; Henricot, B.; Henrici, A.; Beales, P. Morphological and phylogenetic comparisons amongst powdery mildews on Catalpa in the UK. Mycol. Res. 2006, 110, 672–685. [Google Scholar] [CrossRef]
- Mulpuri, S.; Soni, P.K.; Gonela, S.K. Morphological and molecular characterization of powdery mildew on sunflower (Helianthus annuus L.), alternate hosts and weeds commonly found in and around sunflower fields in India. Phytoparasitica 2016, 44, 353–367. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Host Family | Host Species | Voucher 1 | Accession Number 2 | Reference | |
|---|---|---|---|---|---|---|
| ITS | TUB2 | |||||
| Erysiphe alphitoides | Convolvulaceae | Ipomoea obscura | HNIO-18 | MN186769 | n.a. | Pan et al. [34] |
| Fabaceae | Wisteria sinensis | OE2014PM13 | KY660754 | KY786697 | Ellingham et al. [26] | |
| Fagaceae | Quercus robur | OE2014PM103CS | KY660753 | KY786753 | Ellingham et al. [26] | |
| Fagaceae | Q. robur | MUMH1442 | AB257430 * | n.a. | Takamatsu et al. [35] | |
| Fagaceae | Q. robur | MUMH631(t) | AB292708 | n.a. | Takamatsu et al. [36] | |
| E. aquilegiae | Ranunculaceae | Aquilegia sp. | BCRU00359 | LC009883 * | n.a | Takamatsu et al. [9] |
| Ranunculaceae | A. vulgaris | OE2014PM147CS | KY653191 | KY786773 | Ellingham et al. [26] | |
| Ranunculaceae | Clematis apiifolia | MUMH277 | LC009938 * | n.a | Takamatsu et al. [9] | |
| Ranunculaceae | Ranunculus repens | OE2014PM109 | KY653197 | KY786756 | Ellingham et al. [26] | |
| E. betae | Amaranthaceae | Beta vulgaris | KUS F29140 | KX574674 | n.a. | Joa et al. [37] |
| Amaranthaceae | B. vulgaris | WW-14 | DQ164433 | n.a. | Francis et al. [38] | |
| Amaranthaceae | B. vulgaris subsp. cicla | SE1R | KY399969 | n.a. | Vakalounakis and Kavroulakis [39] | |
| Amaranthaceae | Dysphania ambrosioides | MUMH0395 | LC009946 * | n.a. | Takamatsu et al. [9] | |
| E. aff. betae | Polygonaceae | Fagopyrum esculentum | TNM F0034604 | OM033347 * | OM056697 | This study |
| Polygonaceae | F. esculentum | TNM F0035414 | OM368494 * | OM423597 | This study | |
| Polygonaceae | F. esculentum | TNM F0035415 | OM368495 * | OM423598 | This study | |
| Polygonaceae | F. tataricum | TNM F0034605 | OM033348 * | OM056698 | This study | |
| E. convolvuli | Convolvulaceae | Calystegia sepium | MUMH7001 | LC270835 * | n.a. | Abasova et al. [11] |
| Convolvulaceae | Ca. sepium | OE2014PM62CS | KY660794 | KY786726 | Ellingham et al. [26] | |
| Convolvulaceae | Ca. silvatica | OE2014PM48CS | KY660793 | KY786719 | Ellingham et al. [26] | |
| Convolvulaceae | Convolvulus arvensis | UC1512307 | AF011298 | n.a. | Saenz and Taylor [40] | |
| Convolvulaceae | C. arvensis | VPRI 20227 | AF154327 | n.a. | Cunnington et al. [25] | |
| Convolvulaceae | I. aquatica | HMUT1009256 | KJ885178 | n.a. | Abasova et al. [11] | |
| Convolvulaceae | I. tricolor | n.a. | EU621391 | n.a. | Takamatsu et al. [9] | |
| E. convolvuli var. convolvuli | Convolvulaceae | C. arvensis | MUMH7048 | LC328325 | n.a. | Abasova et al. [11] |
| E. heraclei | Apiaceae | Heracleum sphondylium | OE2014PM52 | KY660792 | KY786720 | Ellingham et al. [26] |
| Apiaceae | H. sphondylium | OE2014PM65CS | KY660830 | KY786728 | Ellingham et al. [26] | |
| E. ipomoeae | Convolvulaceae | I. obscura | TNM F0035015(t) | OM033351 * | OM056701 | This study |
| Convolvulaceae | I. obscura | TNM F0035013 | OM033349 * | OM056699 | This study | |
| Convolvulaceae | I. obscura | TNM F0035014 | OM033350 * | OM056700 | This study | |
| Convolvulaceae | I. obscura | TNM F0035018 | OM033354 * | OM056704 | This study | |
| Convolvulaceae | I. obscura | MUMH2978 | LC163910 | n.a. | Meeboon and Takamatsu [29] | |
| Convolvulaceae | I. aquatica | TNM F0034602 | OM033346 * | OM056696 | This study | |
| Convolvulaceae | I. aquatica | TNM F0035410 | OM368491 * | OM423594 | This study | |
| E. lespedezae | Fabaceae | Bauhinia variegata | TNM F0033672 | MT471985 | OM056688 | Xiao et al. [24] and this study |
| Fabaceae | B. variegata | TNM F0033677 | MT471988 | OM056693 | Xiao et al. [24] and this study | |
| Fabaceae | B. variegata | TNM F0033678 | MT471989 | OM056694 | Xiao et al. [24] and this study | |
| Fabaceae | B. blakeana | TNM F0033679 | MT471990 | OM056695 | Xiao et al. [24] and this study | |
| Fabaceae | Desmodium caudatum | TNM F0033671 | MT471984 | OM056687 | Xiao et al. [24] and this study | |
| Fabaceae | D. caudatum | TNM F0033675 | MT471986 | OM056691 | Xiao et al. [24] and this study | |
| Fabaceae | D. caudatum | TNM F0033676 | MT471987 | OM056692 | Xiao et al. [24] and this study | |
| E. neolycopersici | Caricaceae | Carica papaya | n.a. | GU358451 | n.a. | Tsay et al. [41] |
| Sapindaceae | Cardiospermum halicacabum | TNM F0035016 | OM033352 * | OM056702 | This study | |
| Sapindaceae | C. halicacabum | TNM F0035412 | OM368492 * | OM423595 | This study | |
| Sapindaceae | C. halicacabum | TNM F0035413 | OM368493 * | OM423596 | This study | |
| Solanaceae | Solanum lycopersicum | Et-1(t) | AF229019 | n.a. | Kiss et al. [42] | |
| Solanaceae | S. lycopersicum | UMSG2 | KX776199 * | MCFK01008520 | Wu et al. [43] | |
| Solanaceae | S. lycopersicum | MUMH66 | LC009912 * | n.a. | Takamatsu et al. [9] | |
| Solanaceae | S. lycopersicum | TNM F0035404 | OM368490 * | OM423593 | This study | |
| E. pedaliacearum | Pedaliacea | Sesamum indicum | KUS F30128 | LC342963 * | n.a. | Shin et al. [44] |
| E. polygoni | Polygonaceae | Antigonon leptopus | R. Kirschner 4701 | MK685172 | n.a. | Wang et al. [19] |
| Polygonaceae | F. esculentum | n.a. | KP076437 | n.a. | Lu et al. [45] | |
| Polygonaceae | Polygonum sp. | OE2014PM85CS | KY660828 | KY786741 | Ellingham et al. [26] | |
| Polygonaceae | P. aviculare | OE2014PM120CS | KY660829 | KY786760 | Ellingham et al. [26] | |
| Polygonaceae | P. aviculare | MUMH7036 | LC328322 | n.a. | Abasova et al. [11] | |
| E. quercicola | Anacardiaceae | Mangifera indica | TNM F0033673 | OM033344 * | OM056689 | This study |
| Fagaceae | Q. phillyraeoides | MUMH124 | AB193591 | n.a. | Limkaisang et al. [46] | |
| Lauraceae | Cinnamomum camphora | TNM F0033674 | OM033345 * | OM056690 | This study | |
| Sapindaceae | Nephelium lappaceum | MUMH6769 | MN081591 * | n.a. | Meeboon and Takamatsu [47] | |
| E. sedi | Crassulaceae | Sedum aizoon | MUMH2575 | LC010045 * | n.a. | Takamatsu et al. [9] |
| E. takamatsui | Nelumbonaceae | Nelumbo nucifera | TNS F-52102 | AB916688 * | n.a. | Meeboon and Takamatsu [48] |
| G. cichoracearum | Asteraceae | Sonchus oleraceus | UMSG1 | HM449077 | MCBS01024998 | Wu et al. [43] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsiao, H.-Y.; Ariyawansa, H.A.; Hsu, C.-C.; Wang, C.-J.; Shen, Y.-M. New Records of Powdery Mildews from Taiwan: Erysiphe ipomoeae comb. nov., E. aff. betae on Buckwheat, and E. neolycopersici comb. nov. on Cardiospermum halicacabum. Diversity 2022, 14, 204. https://doi.org/10.3390/d14030204
Hsiao H-Y, Ariyawansa HA, Hsu C-C, Wang C-J, Shen Y-M. New Records of Powdery Mildews from Taiwan: Erysiphe ipomoeae comb. nov., E. aff. betae on Buckwheat, and E. neolycopersici comb. nov. on Cardiospermum halicacabum. Diversity. 2022; 14(3):204. https://doi.org/10.3390/d14030204
Chicago/Turabian StyleHsiao, Hui-Yu, Hiran A. Ariyawansa, Ching-Ching Hsu, Chao-Jen Wang, and Yuan-Min Shen. 2022. "New Records of Powdery Mildews from Taiwan: Erysiphe ipomoeae comb. nov., E. aff. betae on Buckwheat, and E. neolycopersici comb. nov. on Cardiospermum halicacabum" Diversity 14, no. 3: 204. https://doi.org/10.3390/d14030204
APA StyleHsiao, H.-Y., Ariyawansa, H. A., Hsu, C.-C., Wang, C.-J., & Shen, Y.-M. (2022). New Records of Powdery Mildews from Taiwan: Erysiphe ipomoeae comb. nov., E. aff. betae on Buckwheat, and E. neolycopersici comb. nov. on Cardiospermum halicacabum. Diversity, 14(3), 204. https://doi.org/10.3390/d14030204