
Composition and Distribution on a Host of Avian Lice of White Storks in North-Eastern Algeria

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
Statistical Analysis
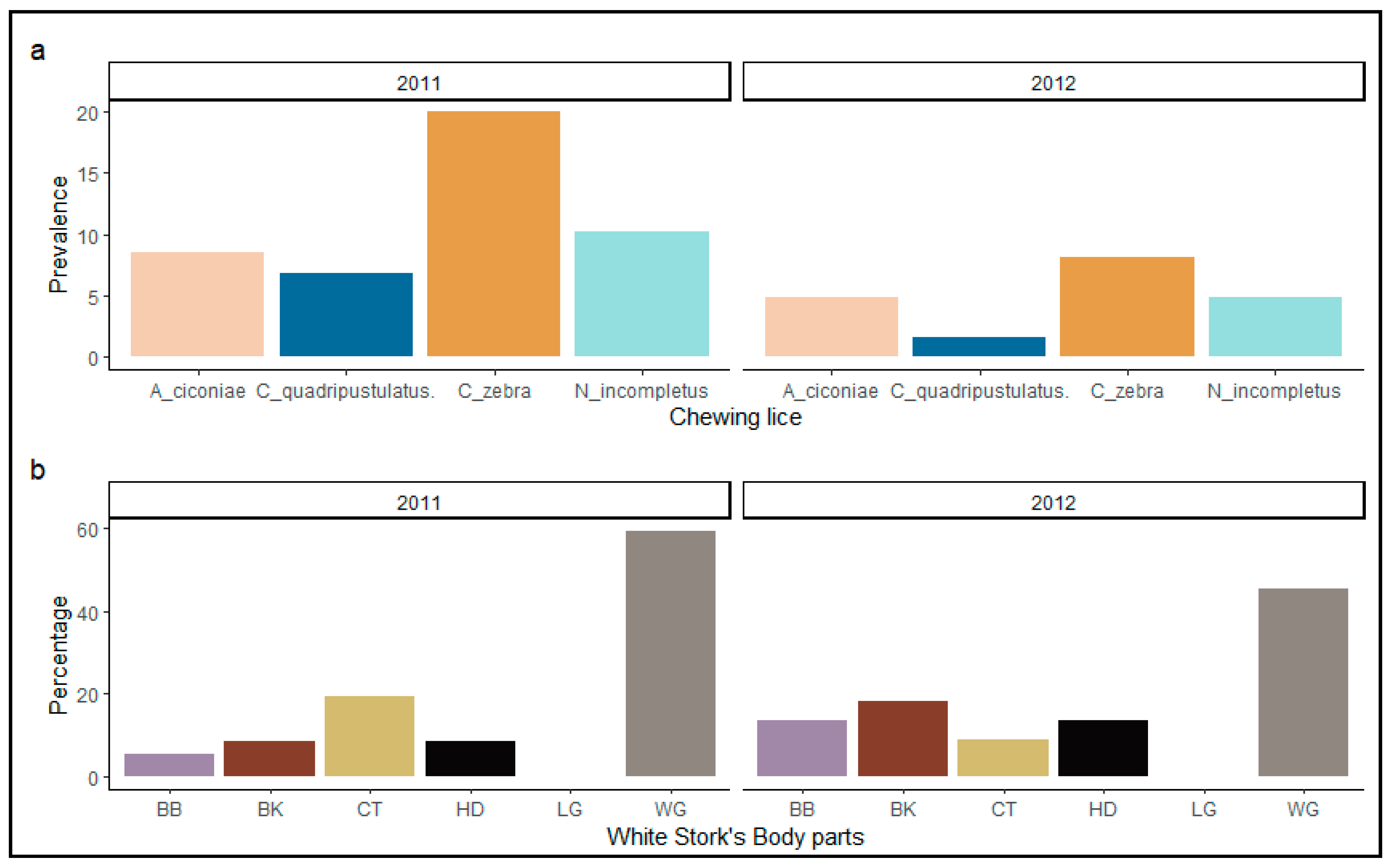
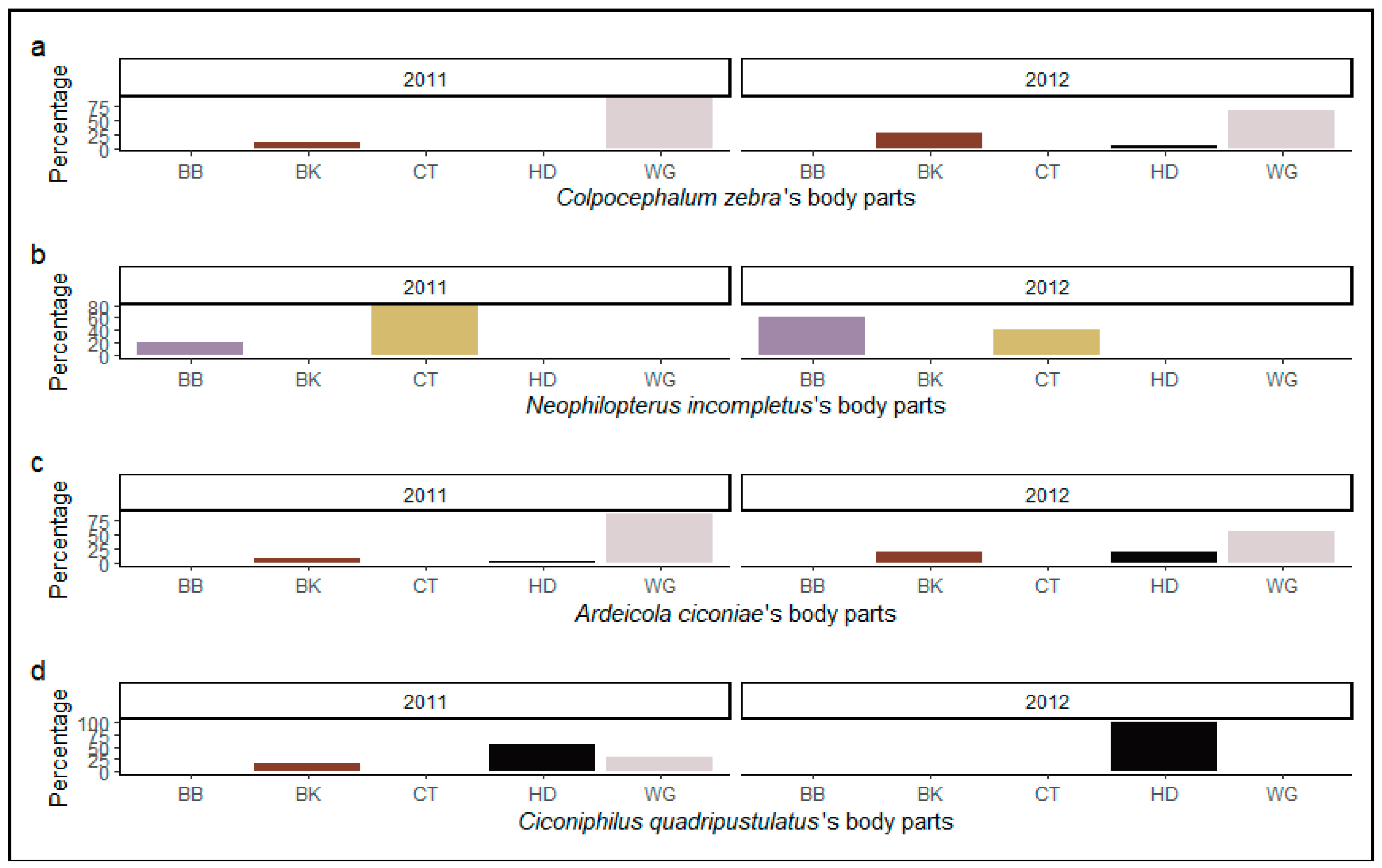
3. Results
3.1. Spatial Distribution of Ectoparasites on Their Host
3.2. Aggregation
4. Discussion
4.1. Aggregation and Sex Ratio
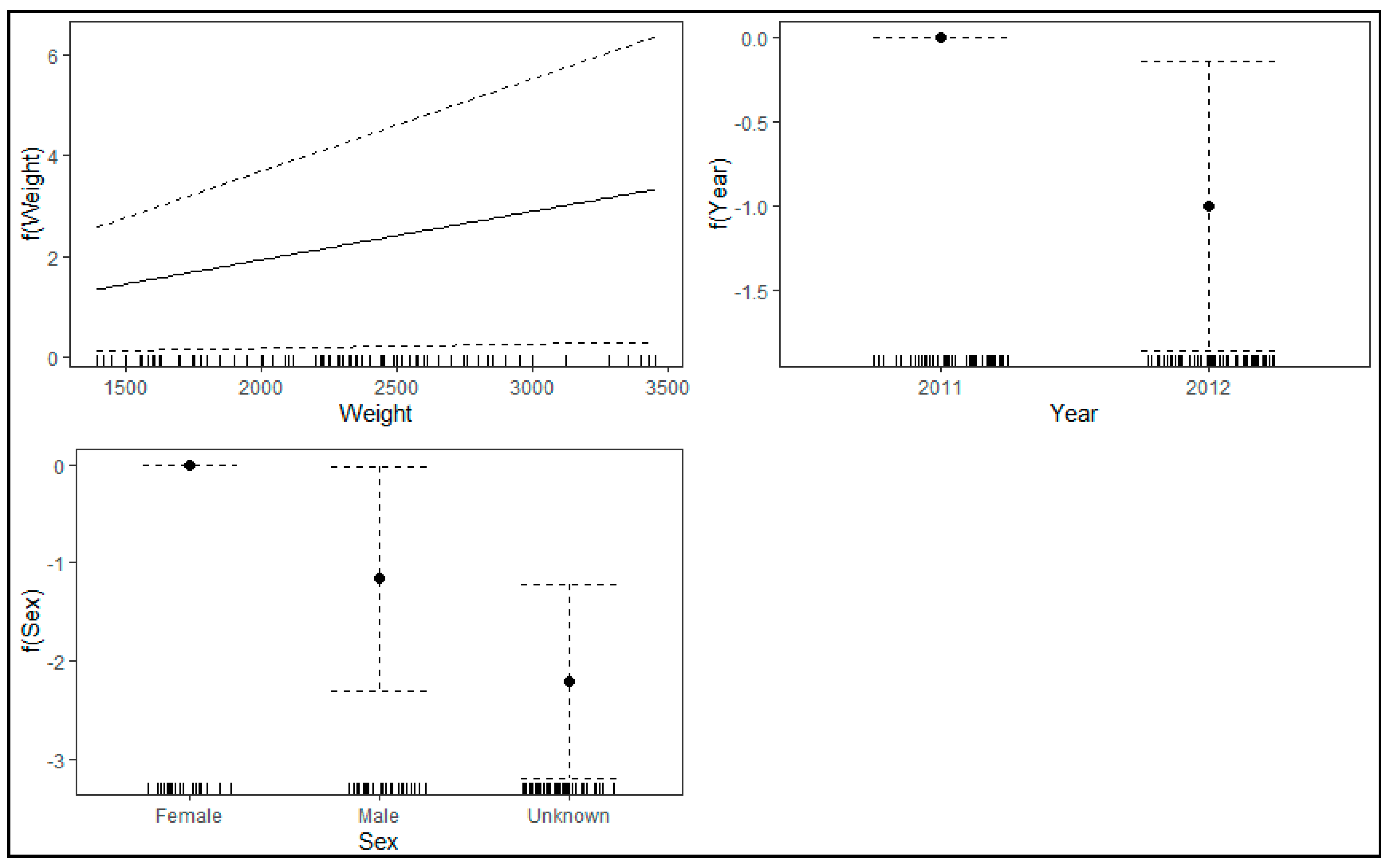
4.2. Factors Affecting Louse Abundance
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Proctor, H.; Owens, I. Mites and birds: Diversity, parasitism and coevolution. Trends Ecol. Evol. 2000, 15, 358–364. [Google Scholar] [CrossRef]
- Møller, A.P. Effects of parasitism by a haematophagous mite on reproduction in the Barn Swallow. Ecology 1990, 71, 2345–2357. [Google Scholar] [CrossRef]
- Lehmann, T. Ectoparasites: Direct impact on host fitness. Parasitol. Today 1993, 9, 8–12. [Google Scholar] [CrossRef]
- Brown, C.R.; Brown, M.B.; Rannala, B. Ectoparasites reduce long-term survival of their avian host. Proc. R. Soc. Lond. Ser. B-Biol. Sci. 1995, 262, 313–319. [Google Scholar]
- Barbosa, A.; Merino, S.; De Lope, F.; Møller, A.P. Effects of feather lice on flight behavior of male Barn Swallows (Hirundo rustica). Auk 2002, 119, 213–216. [Google Scholar] [CrossRef]
- Sheath, D.J.; Dick, J.T.A.; Dickey, J.W.E.; Guo, Z.; Andreou, D.; Britton, J.R. Winning the arms race: Host-parasite shared evolutionary history reduces infection risks in fish final hosts. Biol. Lett. 2018, 14, 20180363. [Google Scholar] [CrossRef] [PubMed]
- Clayton, D.H.; Bush, S.E.; Johnson, K.P. Coevolution of Life on Hosts; University of Chicago Press: Chicago, IL, USA; London, UK, 2016. [Google Scholar]
- Permin, A.; Hansen, J.W. Epidemiology, Diagnosis and Control of Poultry Parasites FAO Animal Health Manuals 4; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 1998. [Google Scholar]
- Fryderyk, S.; Izdebska, J.N. Chewing Lice (Insecta, Phthiraptera,) of the White Stork (Ciconia ciconia L.) in Poland. Ann. Univ. Mariae Curie Sk. 2009, 64, 83–85. [Google Scholar] [CrossRef][Green Version]
- Proctor, H.C. Feather Mites (Acari: Astigmata): Ecology, behavior, and evolution. Annu. Rev. Entomol. 2003, 48, 185–209. [Google Scholar] [CrossRef]
- Horak, I.; Gallivan, G.; Braack, L.; Boomker, J.; Devos, V. Parasites of domestic and Wild animals in South Africa. XLI. Arthropod parasites of impalas, Aepyceros melampus, in the Kruger National Park. Onderstepoort J. Vet. Res. 2005, 70, 131–163. [Google Scholar]
- Hubálek, Z. An Annotated Checklist of Pathogenic Microorganisms Associated with Migratory Birds. J. Wildl. Dis. 2004, 40, 639–659. [Google Scholar] [CrossRef]
- Sychra, O.; Literak, I.; Podzemny, P.; Benidekt, V. Insect ectoparasites from wild passerine birds in the Czech Republic. Parasite 2008, 15, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Palomar, A.; Santibáñez, P.; Mazuelas, D.; Roncero, L.; Santibáñez, S.; Portillo, A.; Otéo, J.A. Role of Birds in Dispersal of Etiologic Agents of Tick-borne Zoonoses, Spain, 2009. Emerg. Infect. Dis. 2012, 18, 1188–1191. [Google Scholar] [CrossRef] [PubMed]
- Arnal, A.; Gómez-Díaz, E.; Cerdà-Cuéllar, M.; Lecollinet, S.; Pearce-Duvet, J.; Busquets, N.; García-Bocanegra, I.; Pages, N.; Vittecoq, M.; Hammouda, A.; et al. Circulation of a Meaban-Like Virus in Yellow-Legged Gulls and Seabird Ticks in the Western Mediterranean Basin. PLoS ONE 2014, 9, e89601. [Google Scholar] [CrossRef] [PubMed]
- De Brooke, M.L.; Nakamura, H. The acquisition of host-specific feather lice by common cuckoos (Cuculus canorus). J. Zool. 1998, 244, 167–173. [Google Scholar] [CrossRef]
- Johnson, K.P.; Bush, S.E.; Clayton, D. Correlated evolution of host and parasite body size: Tests of Harrison’s rule using bird sandlice. Evolution 2005, 59, 1744–1753. [Google Scholar] [CrossRef]
- Clay, T. Geographical distribution of the Mallophaga (Insecta). Bull. Br. Ornithol. Club 1964, 84, 14–16. [Google Scholar]
- Choe, J.C.; Kim, K.C. Ectoparasites of the pelagic cormorant Phalacrocorax pelagicus from the Pribilof Islands, Alaska. J. Med. Entomol. 1987, 24, 584–592. [Google Scholar] [CrossRef] [PubMed]
- Choe, J.C.; Kim, K.C. Microhabitat preference and coexistence of ectoparasites arthropods on Alaskan seabirds. Can. J. Zool. 1988, 66, 987–997. [Google Scholar] [CrossRef]
- Wecksten, J.D. Biogeography explains phylogenetic patterns in toucan chewing lice. Syst. Biol. 2004, 53, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Gotkiewicz, W.; Wittbrodt, K. White stork (Ciconia ciconia L.) as an indicator of natural environment status in agricultural areas, illustrated with an example of Masurian Landscape Park. Environ. Prot. Nat. Resour. 2019, 30, 1–6. [Google Scholar] [CrossRef]
- Cramp, S.; Simmons, K.E.L. HandBook of the Birds of Europe, the Middle East and North Africa: The Birds of the Western Palearctic. Volume 1, Ostrich to Ducks; Oxfords University Press: New York, NY, USA, 1977. [Google Scholar]
- Heim de Balsac, H.; Mayaud, N. Les Oiseaux du Nord-Ouest de l’Afrique; Paul Lechevalier: Paris, France, 1962. [Google Scholar]
- Isenmann, P.; Moali, A. Birds of Algeria; SEOF: Paris, France, 2000. [Google Scholar]
- Samraoui, B.; Samraoui, F. An ornithological survey of the wetlands of Algeria: Important Bird Areas, Ramsar sites and threatened species. Wildfowl 2008, 58, 71–98. [Google Scholar]
- Boukhemza, M.; Boukhemza-Zemmouri, N.; Voisin, J.-F. Biologie et écologie de la reproduction de la Cigogne blanche (Ciconia ciconia) dans la vallée du Sébaou (Kabylie, Algérie). Aves 2007, 44, 213–222. [Google Scholar]
- Djerdali, S.; Tortosa, F.S.; Hillstrom, L.; Doumandji, S. Food supply and external cues limit the clutch size and hatchability in the White Stork Ciconia ciconia. Acta Ornithol. 2008, 43, 145–150. [Google Scholar] [CrossRef]
- Si Bachir, A.; Chenchouni, H.; Djeddou, N.; Barbraud, C.; Céréghino, R.; Santoul, F. Using self-organizing maps to investigate factors regulating colony size and breeding success of the White Stork (Ciconia ciconia). J. Ornithol. 2013, 154, 481–489. [Google Scholar] [CrossRef]
- Benharzallah, N.; Si Bachir, A.; Taleb, F.; Barbraud, C. Factors affecting growth parameters of White Stork nestlings in eastern Algeria. J. Ornithol. 2015, 156, 601–612. [Google Scholar] [CrossRef]
- Bouriach, M.; Samraoui, F.; Souilah, R.; Houma, I.; Razkallah, I.; Alfarhan, A.H.; Samraoui, B. Does core-periphery gradient determine breeding performance in a breeding colony of White Storks Ciconia ciconia? Acta Ornithol. 2015, 50, 149–156. [Google Scholar] [CrossRef]
- Belabed, B.-E.; Athamnia, M.; Touati, L.; Samraoui, F.; Boucheker, A.; Samraoui, B. The early bird catches the worm: Age-specific arrival time influences reproductive performance in the White Stork Ciconia ciconia. Bird Study 2019, 66, 121–129. [Google Scholar] [CrossRef]
- Bouguessa-Cheriak, L.; Doumandji, S.; Ould Messaoud, S.; Marniche, F. Insect ectoparasites on the White Stork, Ciconia ciconia (L.) (Ciconiiformes: Ciconiidae), during breeding period in the extreme east of Algeria. Muzeul Olteniei Craiova. Oltenia-Studii şi comunicări Ştiinţele Naturii. 2017, 33, 72–78. [Google Scholar]
- Rouag-Ziane, N.; Boulahbal, A.; Gauthier-Clerc, M.; Thomas, F.; Chabi, Y. Inventory and quantification of the ectoparasites of the common Coot Fulica atra (Gruiformes: Rallidae) in the northeast of Algeria. Parasite 2007, 14, 253–256. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Touati, L.; Samraoui, B. Diversity and distribution of avian lice on chicks of Greater Flamingo (Phoenicopterus roseus) in Algeria. Avian Biol. Res. 2013, 6, 61–68. [Google Scholar] [CrossRef]
- Touati, L.; Figuerola, J.; Samraoui, B. Patterns in the distribution of avian ectoparasites on Glossy Ibis chicks Plegadis falcinellus in Algeria. Zool. Ecol. 2015, 25, 46–53. [Google Scholar] [CrossRef]
- Temimi, I.; Marniche, F.; Lazli, A.; Milla, A.; Dik, B. The study of the parasites of the bird Egretta garzetta (Linnaeus, 1766) (Aves: Ardeidae) in northeastern of Algeria. Muzeul Olteniei Craiova. Oltenia-Studii şi Comun. Ştiinţele Naturii. 2017, 33, 65–68. [Google Scholar]
- Baziz-Neffah, F.; Bitam, I.; Kernif, T. Contribution to the knowledge of ectoparasites of birds in Algeria. Bull. Soc. Zool. Fr. 2015, 140, 81–98. [Google Scholar]
- Møller, A.P. Advantages and disavantages of coloniality in the swallow (Hirundo rustica). Anim. Behav. 1987, 35, 819–832. [Google Scholar] [CrossRef]
- Møller, A.P.; Dufva, R.; Allander, K. Parasites and the evolution of host sexual behavior. Adv. Study Behav. 1993, 22, 65–102. [Google Scholar]
- Rózsa, L.; Rékázi, J.; Reiczigel, J. Relationship of host coloniality to the population ecology of avian lice (Insecta: Phtiraptera). J. Anim. Ecol. 1996, 65, 242–248. [Google Scholar] [CrossRef]
- Samraoui, B. White Storks wintering in northeast Algeria. Br. Birds 1998, 91, 377. [Google Scholar]
- Palma, R.L. Slide-mounting of lice: A detailed description of the Canada balsam technique. N. Zeal. Entomol. 1978, 6, 432–436. [Google Scholar] [CrossRef]
- Séguy, E. Faune de France, 43. Insectes Ectoparasites (Mallophages, Anoploures, Siphonaptères); Paul Lechevalier: Paris, France, 1944. [Google Scholar]
- Price, R.D.; Hallenthal, R.A.; Palma, R.L.; Johnson, K.P.; Clayton, D.H. The Chewing Lice the World. World Checklist and Biological Overview; Illinois Natural History Survey Special Publication: Champaign, IL, USA, 2003; Volume 24. [Google Scholar]
- Wall, R.; Shearer, D. Veterinary Ectoparasites: Biology, Pathology and Control; Blackwell Science: Hoboken, NJ, USA, 2008. [Google Scholar]
- Griffith, R.; Double, M.C.; Orr, K.; Dawson, R.J.G. A DNA test to sex most birds. Mol. Ecol. 2002, 7, 1071–1075. [Google Scholar] [CrossRef]
- Margolis, L.; Eschi, G.W.; Holmes, J.-C.; Kuris, A.M.; Shad, G.A. The use of ecological terms in parasitology (report of an ad hoc committee of the American Society of Parasitologists). J. Parasitol. 1982, 68, 131–133. [Google Scholar] [CrossRef]
- Fowler, J.; Cohen, L. Statistics for Ornithologists, 2nd ed.; BTO Guide N.2; British Trust for Ornithology: Thetford, UK, 1996. [Google Scholar]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Stak, A.W. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Wilson, K.; Bjornstad, O.N.; Dobson, P.; Merler, S.; Poglayen, G.; Randolph, S.E.; Read, A.F.; Skorping, A. Chapter2: Heterogeneities in macroparasites infections: Patterns and processes. In The Ecology of Wildlife Diseases; Hudson, P.J., Rizzoli, A., Grenfell, B.T., Heesterbeek, H., Dobson, A.P., Eds.; Oxford University Press: Oxford, UK, 2001; pp. 6–44. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Tendeiro, J. Estudos sobre uma Colecção de malóphagos de aves. Bol. Cult. Da Guiné Port. 1955, 35, 497–625. [Google Scholar]
- Carriker, M.A. On the genera “Ciconiphilus” and “Ardeiphilus” with description of six new species. Rev. Bras. de Biol. 1964, 24, 95–108. [Google Scholar]
- Price, R.D.; Beer, J.R. Colpocephalum (Mallophaga: Menoponidae) of the Ciconiiformes. Ann. Entomol. Soc. Am. 1965, 1, 111–131. [Google Scholar] [CrossRef]
- Price, R.D.; Beer, J.R. A review of Ciconiphilus Bedford (Mallophaga: Menoponidae). Can. Entomol. 1965, 97, 657–666. [Google Scholar] [CrossRef]
- Price, R.D.; Emerson, K.C. Two new species of Colpocephalum (Mallophaga: Menoponidae) from Neotropical Ciconiiformes. Ann. Entomol. Soc. Am. 1967, 60, 875–878. [Google Scholar] [CrossRef]
- Tuff, D.W. A review of North American Ardeicola (Mallophaga: Philopteridae). J. Kans. Entomol. Soc. 1967, 40, 241–263. [Google Scholar]
- Price, R.C. A new species of Colpocephalum (Phtiraptera) on Threskiornis (Aves) from Aldabra. Syst. Entomol. 1976, 1, 61–63. [Google Scholar] [CrossRef]
- Hajela, K.P.; Tandan, B.K. Species of Ardeicola (Insecta: Mallophaga) parasitic on birds of the family Threskiornithidae. Zool. J. Linnaean Soc. 1970, 49, 309–334. [Google Scholar] [CrossRef]
- Martin Mateo, M.P. Phtiraptera from Platalea leucorodia (L.) (Aves: Ciconiiformes: Threskiornithidae) in Spain. Res. Rev. Parasitol. 1994, 54, 109–115. [Google Scholar]
- Martin Mateo, M.P. Mallophaga, Amblycera. In Fauna Iberica; Ramos, M.A., et al., Eds.; Nacional de Ciencias Naturales Consejo Superior de Investigaciones Cientificas: Madrid, Spain, 2002; Volume 20, pp. 1–187. [Google Scholar]
- İnci, A.; Yıldırım, A.; Dik, B.; Düzlü, O. Current knowledge of Turkey’s louse fauna. Türkiye Parazitoloji Dern. 2011, 34, 212–220. [Google Scholar] [CrossRef]
- Martin Mateo, M.P. Maláfagos (Insecta) parásitos de la cigüeña comun, Ciconia ciconia (L.) (Aves). Rev. Esp. Entomol. 1988, 64, 147–158. [Google Scholar]
- Dik, B.; Uslu, U. Mallophaga (Insecta) species occurring on Storks (Ciconia ciconia) Linnaeus, 1758). Turk. Soc. Parasitol. 2006, 30, 220–225. [Google Scholar]
- Adam, C. Data on the chewing louse fauna (Phthiraptera: Amblycera, Ischnocera) from some Romanian autochthonous and exotic birds. Trav. du Mus. Natl. d’histoire Nat. “Grigore Antipa” 2007, 50, 145–210. [Google Scholar]
- Clay, T. Some problems in the evolution of a group of ectoparasites. Evolution 1949, 3, 279–299. [Google Scholar] [CrossRef] [PubMed]
- Palma, R.L.; Johnson, A.R.; Cezilly, F.; Thomas, F.; Renaud, F. Diversity and distribution of feather lice on Greater Flamingoes (Phoenicopterus ruber roseus) in the Camargue, Southern France. N. Zeal. Entomol. 2002, 25, 87–89. [Google Scholar] [CrossRef]
- Nelson, B.C.; Murray, M.D. The distribution of Mallophaga on the domestic Pigeon (Columba livia). Int. J. Parasitol. 1971, 1, 21–29. [Google Scholar] [CrossRef]
- Clay, T. The Phtiraptera (Insecta) parasitic on flamingoes (Phoenicopteridae: Aves). J. Zool. Lond. 1974, 172, 483–490. [Google Scholar] [CrossRef]
- Johnson, K.P.; Shreve, S.M.; Smith, V.S. Repeated adaptive divergence of microhabitat specialization in avian feather lice. BMC Biol. 2012, 10, 52. Available online: http://www.biomedcentral.com/1741-7007/10/52 (accessed on 8 January 2022). [CrossRef] [PubMed]
- Crompton, D.W.T. Birds as habitat for parasites. In Host-Parasite Evolution. General Principles and Avian Models; Clayton, D.H., Moore, J., Eds.; Oxford University Press: Oxford, UK, 1997; pp. 253–270. [Google Scholar]
- Bush, S.E.; Sohn, E.; Clayton, D.H. Ecomorphology of parasite attachment: Experiments with feather lice. J. Parasitol. 2006, 92, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Clayton, D.H.; Moyer, B.R.; Bush, S.E.; Jones, T.G.; Gardiner, D.W.; Rhodes, B.B.; Goller, F. Adaptive significance of avian beak morphology for ectoparasites control. Proc. R. Soc. Lond. B 2005, 272, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Fowler, J.A.; Williams, I.R. Population dynamics of Mallophaga and Acari on reed buntings occupying a communal winter roost. Ecol. Entomol. 1985, 10, 377–383. [Google Scholar] [CrossRef]
- Clayton, D.H.; Lee, P.L.M.; Tompkins, D.M.; Brodie, E.D., III. Reciprocal natural selection on host-parasite phenotypes. Am. Nat. 1999, 154, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Rózsa, L. Patterns in the abundance of avian lice (Phthiraptera: Amblycera, Ischnocera). J. Avian Biol. 1997, 28, 249–254. [Google Scholar] [CrossRef][Green Version]
- Clayton, D.H.; Walther, B.A. Influence of host ecology and morphology on the diversity of Neotropical bird lice. Oikos 2001, 94, 455–467. [Google Scholar] [CrossRef]
- Clayton, D.H.; Drown, D.M. Critical evaluation of five methods for quantifying chewing lice (Insecta: Phthiraptera). J. Parasitol. 2001, 87, 1291–1300. [Google Scholar] [CrossRef]
- Miller, M.J.R.; Ewins, P.J.; Galloway, T.D. Records of ectoparasites collected on ospreys from Ontario. J. Wildl. Dis. 1997, 33, 373–376. [Google Scholar] [CrossRef]
- Darolova, A.; Hoi, H.; Kristofik, J.; Hoi, C. Horizontal and vertical ectoparasites transmission of three species of mallophaga, and individual variation in European bee-eaters (Merops apiaster). J. Parasitol. 2001, 87, 256–262. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ectoparasites | Year | Infested Hosts | Prevalence (%) | Mean Abundance (±sd) | Mean Intensity (±sd) |
|---|---|---|---|---|---|
| Colpocephalum zebra (Amblycera, Menoponidae) | 2011 | 10/59 | 19.92 | 0.59 ± 1.52 | 1.4 ± 2.12 |
| 2012 | 5/61 | 8.19 | 0.27 ± 1.02 | 1.41 ± 1.85 | |
| Neophilopterusincompletus (Amblycera, Menoponidae) | 2011 | 6/59 | 10.16 | 0.38 ± 1.27 | 0.92 ± 1.2 |
| 2012 | 3/61 | 4.91 | 0.16 ± 0.72 | 0.83 ± 1.42 | |
| Ardeicola ciconiae (Ischnocera, Philopteridae) | 2011 | 5/59 | 8.47 | 0.37 ± 1.2 | 0.88 ± 1.73 |
| 2012 | 3/61 | 4.91 | 0.24 ± 1.08 | 1.25 ± 2.14 | |
| Ciconiphilus quadripustulatus (Ischnocera, Philopteridae) | 2011 | 4/59 | 6.77 | 0.22 ± 0.99 | 0.52 ± 1.53 |
| 2012 | 1/61 | 1.63 | 0.03 ± 0.25 | 0.16 ± 0.53 |
| Ectoparasites | Year | Variance (S2) | Mean Abundance (±sd) | Index of Aggregation |
|---|---|---|---|---|
| Colpocephalum zebra (Amblycera, Menoponidae) | 2011 | 2.31 | 0.59 ± 1.52 | 3.91 |
| 2012 | 1.02 | 0.27 ± 1.02 | 3.77 | |
| Neophilopterusincompletus (Amblycera, Menoponidae) | 2011 | 1.59 | 0.38 ± 1.27 | 4.18 |
| 2012 | 0.53 | 0.16 ± 0.72 | 3.31 | |
| Ardeicola ciconiae (Ischnocera, Philopteridae) | 2011 | 1.44 | 0.37 ± 1.2 | 3.89 |
| 2012 | 1.20 | 0.24 ± 1.08 | 5.00 | |
| Ciconiphilus quadripustulatus (Ischnocera, Philopteridae) | 2011 | 1.03 | 0.22 ± 0.99 | 4.68 |
| 2012 | 0.06 | 0.03 ± 0.25 | 2.00 |
| Parametric Coefficients | Estimate | Std. Error | z-Value | p-Value |
|---|---|---|---|---|
| Intercept | −0.48 | 1.1 | −0.44 | 0.662 |
| Weight | 9.60 × 10−4 | 4.50 × 10−4 | −2.15 | 0.031 |
| Year_2012 | −1 | 0.44 | −2.29 | 0.022 |
| Sex_Male | −1.16 | 0.58 | −1.99 | 0.046 |
| Sex_Unknown | −2.2 | 0.51 | −4.36 | 1.33 × 10−5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Touati, L.; Athamnia, M.; Nedjah, R.; Boucheker, A.; Samraoui, F.; El-Serehy, H.A.; Samraoui, B. Composition and Distribution on a Host of Avian Lice of White Storks in North-Eastern Algeria. Diversity 2022, 14, 77. https://doi.org/10.3390/d14020077
Touati L, Athamnia M, Nedjah R, Boucheker A, Samraoui F, El-Serehy HA, Samraoui B. Composition and Distribution on a Host of Avian Lice of White Storks in North-Eastern Algeria. Diversity. 2022; 14(2):77. https://doi.org/10.3390/d14020077
Chicago/Turabian StyleTouati, Laïd, Mohammed Athamnia, Riad Nedjah, Abdennour Boucheker, Farrah Samraoui, Hamed A. El-Serehy, and Boudjéma Samraoui. 2022. "Composition and Distribution on a Host of Avian Lice of White Storks in North-Eastern Algeria" Diversity 14, no. 2: 77. https://doi.org/10.3390/d14020077
APA StyleTouati, L., Athamnia, M., Nedjah, R., Boucheker, A., Samraoui, F., El-Serehy, H. A., & Samraoui, B. (2022). Composition and Distribution on a Host of Avian Lice of White Storks in North-Eastern Algeria. Diversity, 14(2), 77. https://doi.org/10.3390/d14020077