
Mesocarnivore Distribution along Gradients of Anthropogenic Disturbance in Mediterranean Landscapes




Abstract
:1. Introduction
2. Materials and Methods
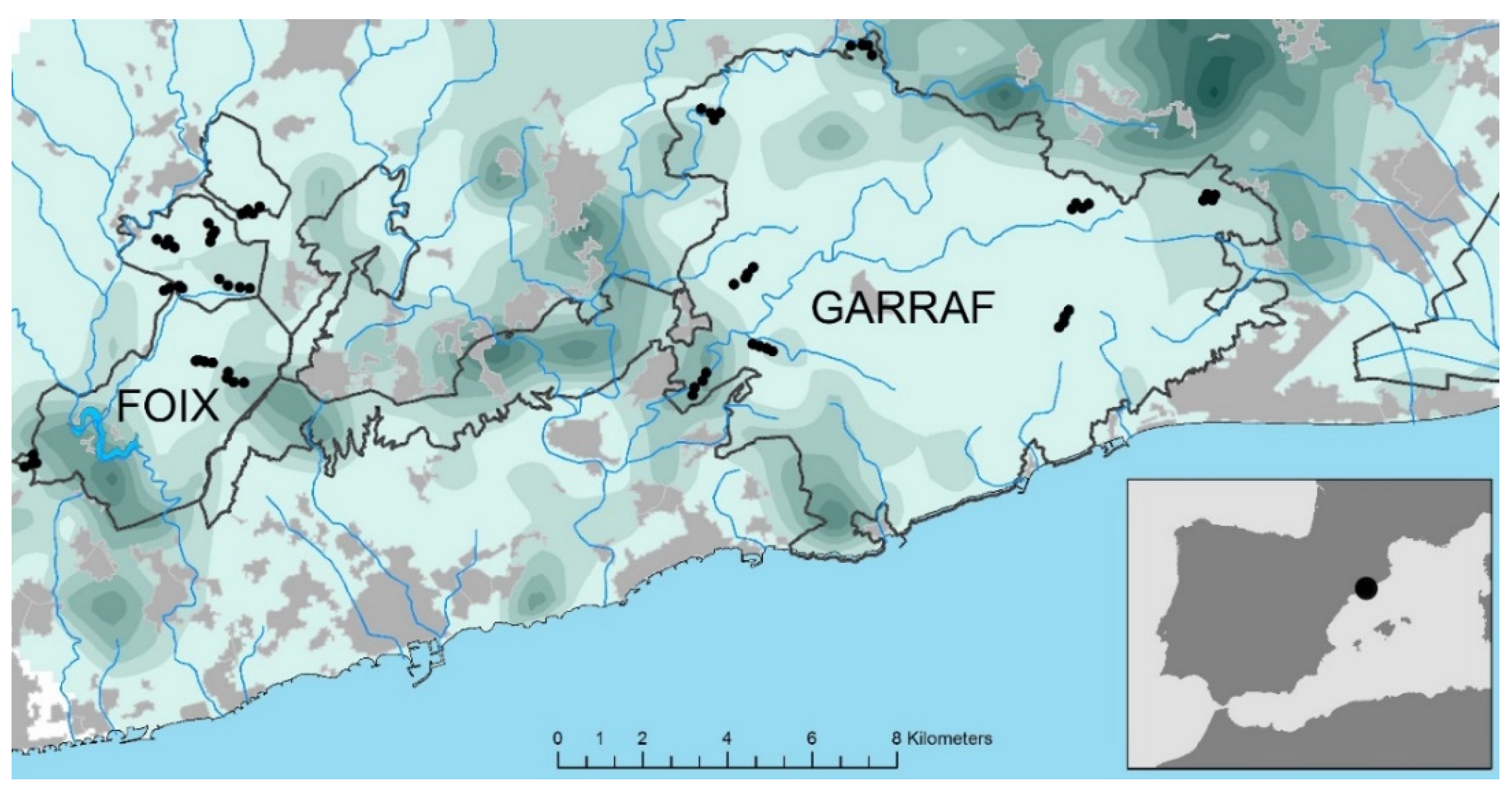
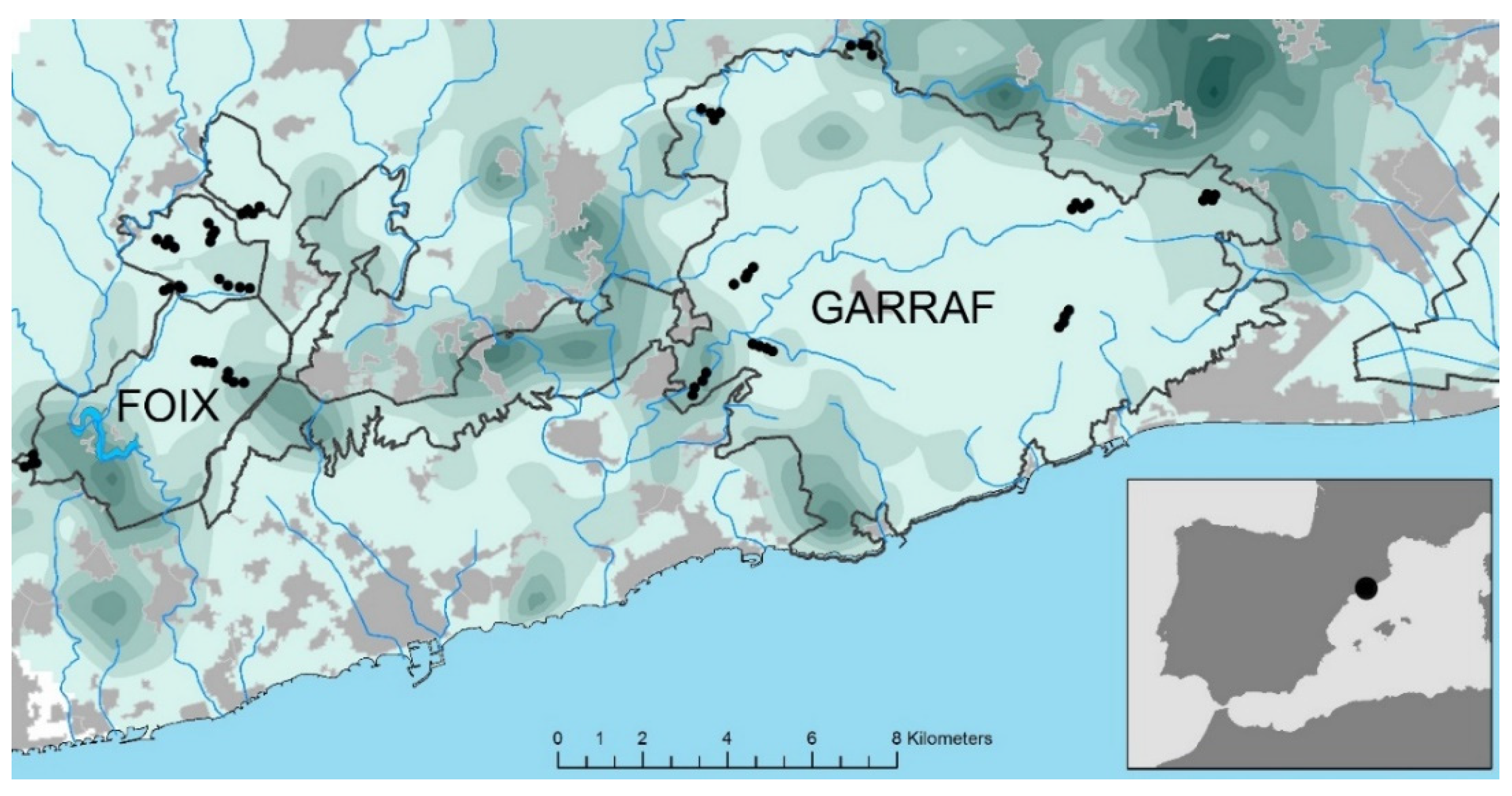
2.1. Study Area and Sampling Design
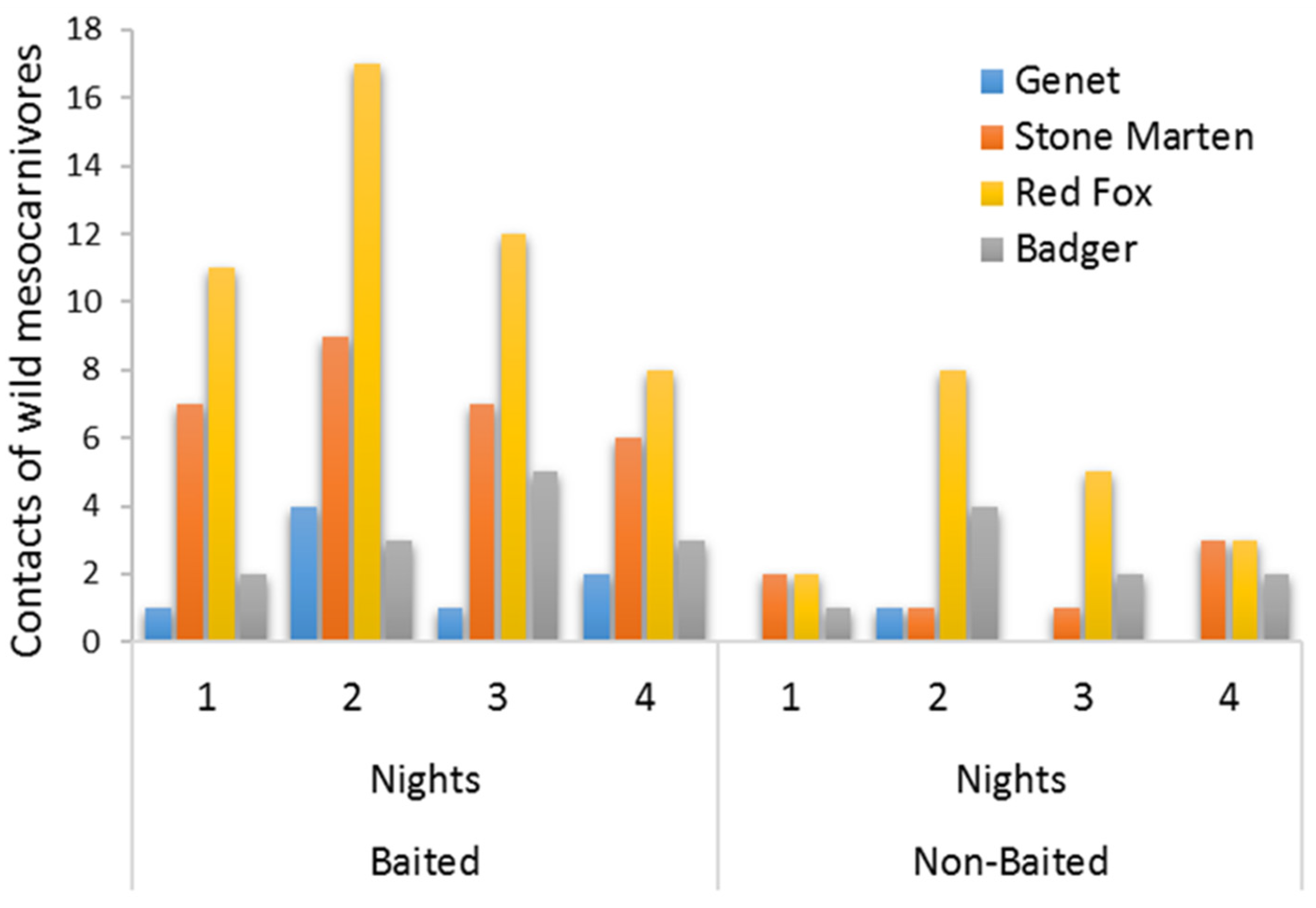
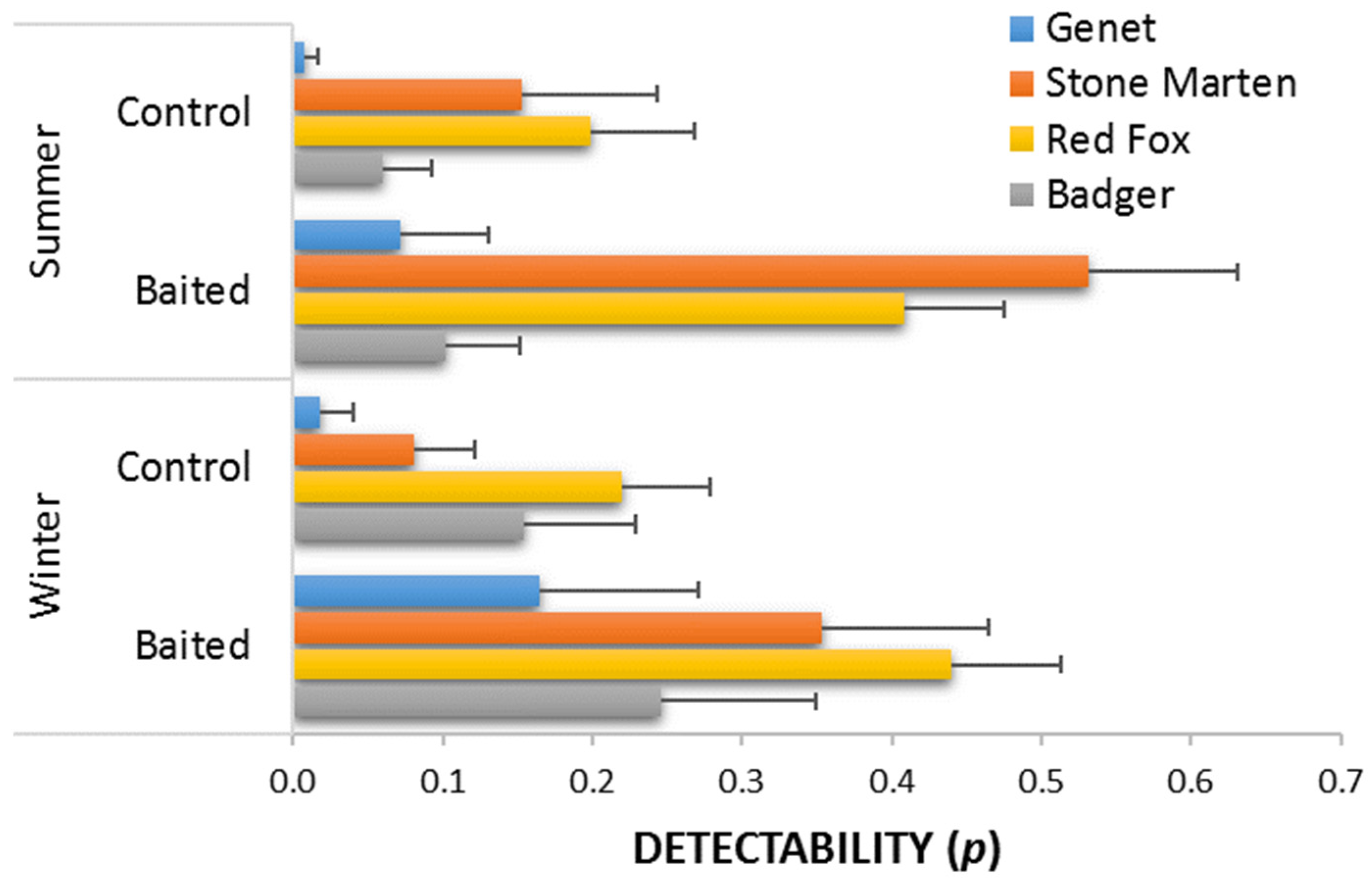
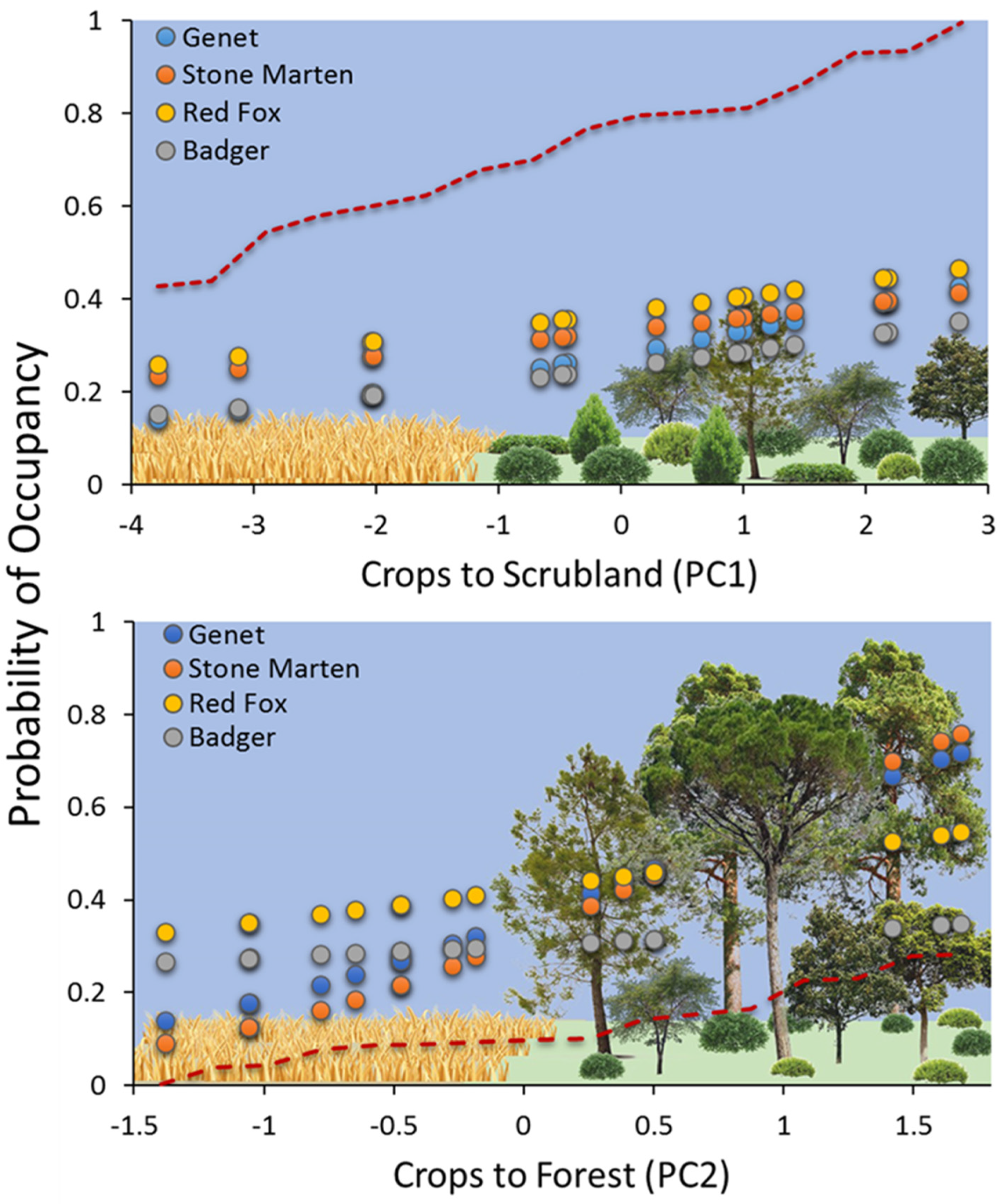
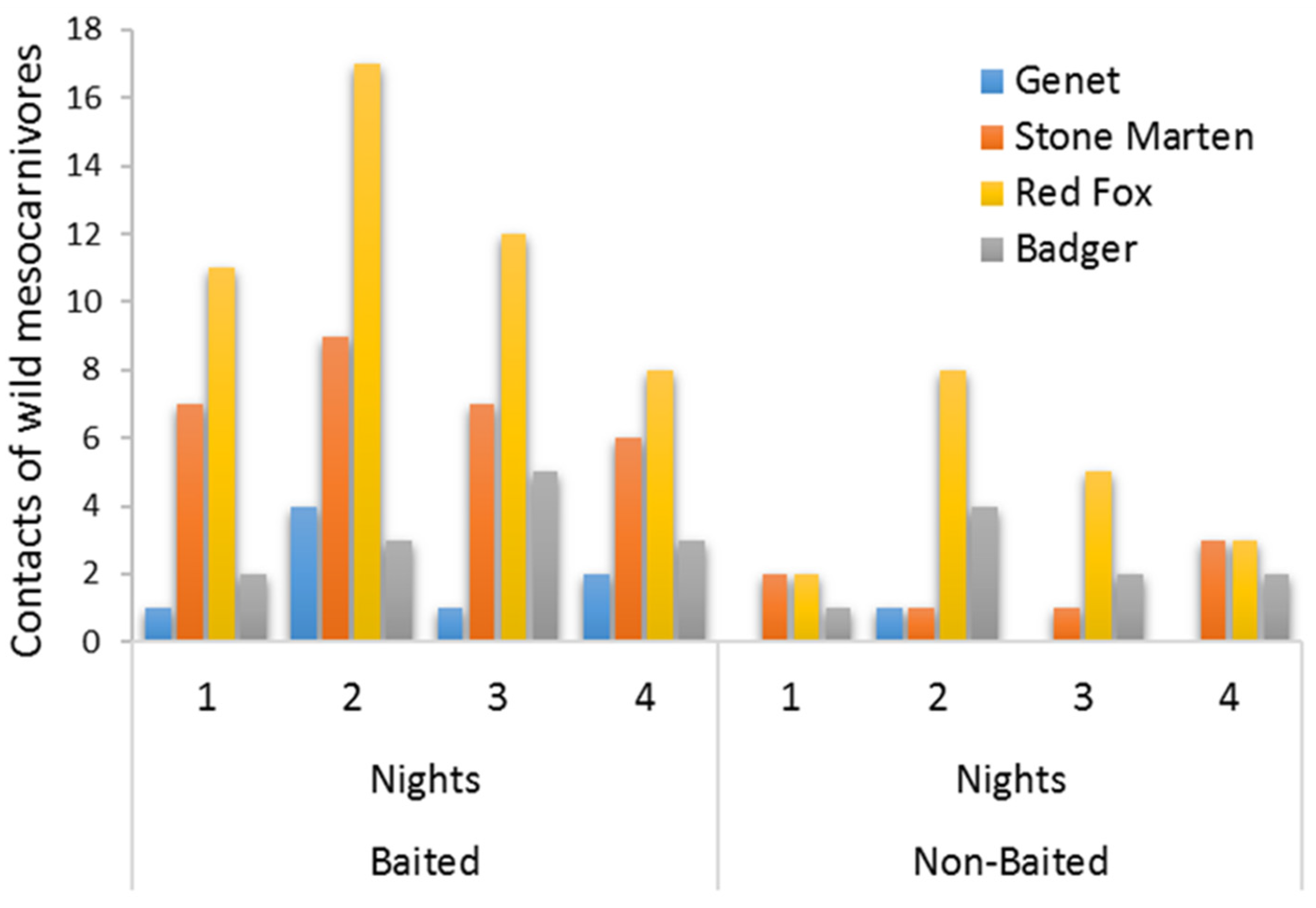
2.2. Statistical Analyses
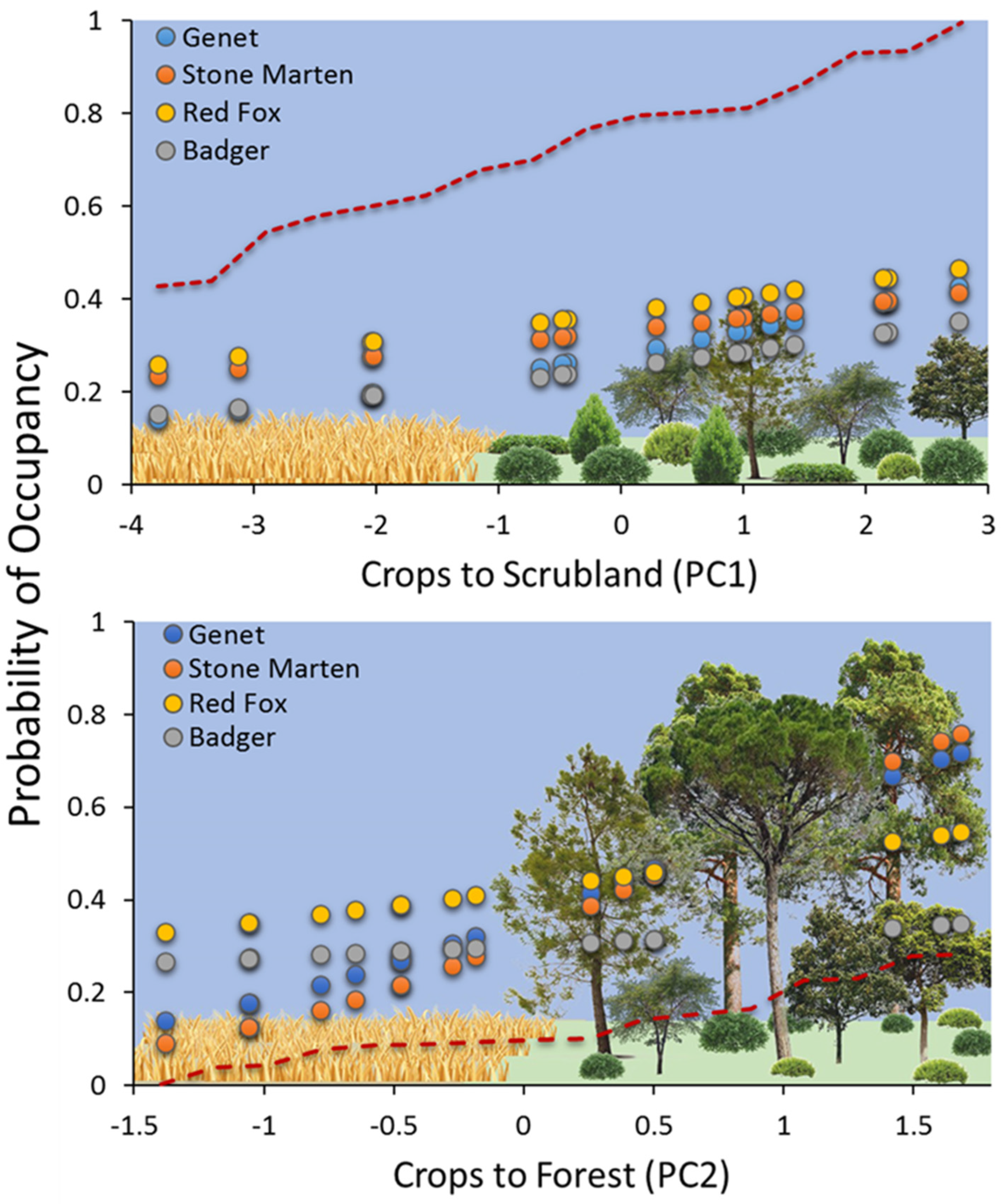
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Díaz, M.; Grim, T.; Markó, G.; Morelli, F.; Ibáñez-Alamo, J.D.; Jokimäki, J.; Kaisanlahti-Jokimäki, M.L.; Tätte, K.; Tryjanowski, P.; Møller, A.P. Effects of climate variation on bird escape distances modulate community responses to global change. Sci. Rep. 2021, 11, 12826. [Google Scholar] [CrossRef] [PubMed]
- Crooks, K.R.; Soulé, M.E. Mesopredator release and avifaunal extinctions in a fragmented system. Nature 1999, 400, 563–566. [Google Scholar] [CrossRef]
- Palomares, F.; Delibes, M.; Revilla, E.; Calzada, J.; Fedriani, J.M. Spatial ecology of Iberian lynx and abundance of European rabbits in southwestern Spain. Wildl. Monogr. 2001, 148, 1–36. [Google Scholar]
- Diaz, M.; Torre, I.; Peris, A.; Tena, L. Foraging behavior of wood mice as related to presence and activity of genets. J. Mammal. 2005, 86, 1178–1185. [Google Scholar] [CrossRef]
- Navarro-Castilla, Á.; Barja, I.; Díaz, M. Foraging, feeding, and physiological stress responses of wild wood mice to increased illumination and common genet cues. Curr. Zool. 2018, 64, 409–417. [Google Scholar] [CrossRef] [Green Version]
- Gallego, D.; Morán-López, T.; Torre, I.; Navarro-Castilla, Á.; Barja, I.; Díaz, M. Context dependence of acorn handling by the Algerian mouse (Mus spretus). Acta Oecol. 2017, 84, 1–7. [Google Scholar] [CrossRef]
- Sergio, F.; Newton, I.; Marchesi, L. Conservation: Top predators and biodiversity. Nature 2005, 436, 192. [Google Scholar] [CrossRef]
- Mendoza, M.; Araújo, M.B. Climate shapes mammal community trophic structures and humans simplify them. Nat. Commun. 2019, 10, 1–9. [Google Scholar] [CrossRef]
- Newbold, T.; Hudson, L.N.; Hill, S.L.L.; Contu, S.; Lysenko, I.; Senior, R.A.; Börger, L.; Bennett, D.J.; Choimes, A.; Collen, B.; et al. Global effects of land use on local terrestrial biodiversity. Nature 2015, 520, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Amici, V.; Battisti, C. Selecting focal species in ecological network planning following an expert-based approach: A case study and a conceptual framework. Landsc. Res. 2009, 34, 545–561. [Google Scholar] [CrossRef]
- Roemer, G.W.; Gompper, M.E.; Van Valkenburgh, B. The Ecological Role of the Mammalian Mesocarnivore. Bioscience 2009, 59, 165–173. [Google Scholar] [CrossRef]
- Alexandre, M.; Hipólito, D.; Ferreira, E.; Fonseca, C.; Rosalino, L.M. Humans do matter: Determinants of red fox (Vulpes vulpes) presence in a western Mediterranean landscape. Mammal. Res. 2020, 65, 203–214. [Google Scholar] [CrossRef]
- Torre, I.; Jaime-González, C.; Díaz, M. Habitat Suitability for Small Mammals in Mediterranean Landscapes: How and Why Shrubs Matter. Sustainability 2022, 14, 1562. [Google Scholar] [CrossRef]
- Curveira-Santos, G.; Marques, T.A.; Björklund, M.; Santos-Reis, M. Mediterranean mesocarnivores in spatially structured managed landscapes: Community organisation in time and space. Agric. Ecosyst. Environ. 2017, 237, 280–289. [Google Scholar] [CrossRef] [Green Version]
- Virgós, E.; Tellería, J.L.; Santos, T. A comparison on the response to forest fragmentation by medium-sized Iberian carnivores in central Spain. Biodivers. Conserv. 2002, 11, 1063–1079. [Google Scholar] [CrossRef]
- Regos, A.; D’Amen, M.; Titeux, N.; Herrando, S.; Guisan, A.; Brotons, L. Predicting the future effectiveness of protected areas for bird conservation in Mediterranean ecosystems under climate change and novel fire regime scenarios. Divers. Distrib. 2016, 22, 83–96. [Google Scholar] [CrossRef]
- Puerta-Piñero, C.; Espelta, J.M.; Sanchez-Humanes, B.; Rodrigo, A.; Coll, L.; Brotons, L. History matters: Previous land use changes determine post-fire vegetation recovery in forested Mediterranean landscapes. For. Ecol. Manag. 2012, 279, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Pausas, J.G. Changes in Fire and Climate in the Eastern Iberian Peninsula (Mediterranean Basin). Clim. Chang. 2004, 63, 337–350. [Google Scholar] [CrossRef]
- Doblas-Miranda, E.; Martínez-Vilalta, J.; Lloret, F.; Álvarez, A.; Ávila, A.; Bonet, F.J.; Brotons, L.; Castro, J.; Curiel Yuste, J.; Díaz, M.; et al. Reassessing global change research priorities in mediterranean terrestrial ecosystems: How far have we come and where do we go from here? Glob. Ecol. Biogeogr. 2015, 24, 25–43. [Google Scholar] [CrossRef] [Green Version]
- Virgós, E. Relative value of riparian woodlands in landscapes with different forest cover for medium-sized Iberian. Biodivers. Conserv. 2001, 10, 1039–1049. [Google Scholar] [CrossRef]
- Swetnam, T.W.; Allen, C.D.; Betancourt, J. Applied historical ecology: Using the past to manage for the future. Ecol. Appl. 1999, 9, 1189–1206. [Google Scholar] [CrossRef]
- Riera, J.; Castell, C. Efectes dels incendis forestals recurrents sobre la distribució de dues espècies del Parc Natural del Garraf: El pi blanc (Pinus halepensis) i la savina (Juniperus phoenicea). Butlletí Inst. Catalana D’història Nat. 1997, 65, 105–116. [Google Scholar]
- Baśnou, C.; Álvarez, E.; Bagaria, G.; Guardiola, M.; Isern, R.; Vicente, P.; Pino, J. Spatial patterns of land use changes across a mediterranean metropolitan landscape: Implications for biodiversity management. Environ. Manag. 2013, 52, 971–980. [Google Scholar] [CrossRef] [PubMed]
- Dytham, C. Choosing and Using Statistics. A biologist’s Guide, 3rd ed.; B lackwell Science, Ed.; Wiley-Blackwell: Oxford, UK, 2011. [Google Scholar]
- Ferreras, P.; DÍaz-Ruiz, F.; Monterroso, P. Improving mesocarnivore detectability with lures in camera-trapping studies. Wildl. Res. 2018, 45, 505–517. [Google Scholar] [CrossRef]
- Molina-Vacas, G.; Bonet-Arbolí, V.; Rodríguez-Teijeiro, J.D. Habitat selection of two medium-sized carnivores in an isolated and highly anthropogenic Mediterranean park: The importance of riverbank vegetation. Ital. J. Zool. 2012, 79, 128–135. [Google Scholar] [CrossRef]
- Schuette, P.; Wagner, A.P.; Wagner, M.E.; Creel, S. Occupancy patterns and niche partitioning within a diverse carnivore community exposed to anthropogenic pressures. Biol. Conserv. 2013, 158, 301–312. [Google Scholar] [CrossRef] [Green Version]
- Barrull, J.; Mate, I.; Ruiz-Olmo, J.; Casanovas, J.G.; Gosàlbez, J.; Salicrú, M. Factors and mechanisms that explain coexistence in a Mediterranean carnivore assemblage: An integrated study based on camera trapping and diet. Mamm. Biol. 2014, 79, 123–131. [Google Scholar] [CrossRef]
- Recio, M.R.; Arija, C.M.; Cabezas-Díaz, S.; Virgós, E. Changes in Mediterranean mesocarnivore communities along urban and ex-urban gradients. Curr. Zool. 2015, 61, 793–801. [Google Scholar] [CrossRef] [Green Version]
- Ferreras, P.; Díaz-Ruiz, F.; Alves, P.C.; Monterroso, P. Optimizing camera-trapping protocols for characterizing mesocarnivore communities in south-western Europe. J. Zool. 2017, 301, 23–31. [Google Scholar] [CrossRef]
- Requena-Mullor, J.M.; López, E.; Castro, A.J.; Cabello, J.; Virgós, E.; González-Miras, E.; Castro, H. Modeling spatial distribution of European badger in arid landscapes: An ecosystem functioning approach. Landsc. Ecol. 2014, 29, 843–855. [Google Scholar] [CrossRef]
- Grajera, J.; Vilella, M.; Torre, I. A pilot study of the use of dry dog food as an alternative attractant in mesocarnivore studies. Mammalia 2021, 85, 422–427. [Google Scholar] [CrossRef]
- Torre, I.; Arrizabalaga, A.; Freixas, L.; Ribas, A.; Flaquer, C.; Diaz, M. Using scats of a generalist carnivore as a tool to monitor small mammal communities in Mediterranean habitats. Basic Appl. Ecol. 2013, 14, 155–164. [Google Scholar] [CrossRef]
- Magurran, A.E.; Baillie, S.R.; Buckland, S.T.; Dick, J.M.; Elston, D.A.; Scott, E.M.; Smith, R.I.; Somerfield, P.J.; Watt, A.D. Long-term datasets in biodiversity research and monitoring: Assessing change in ecological communities through time. Trends Ecol. Evol. 2010, 25, 574–582. [Google Scholar] [CrossRef] [PubMed]
- O’Hara, R.B.; Kotze, D.J. Do not log-transform count data. Methods Ecol. Evol. 2010, 1, 118–122. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Barton, K. Package “MuMIn”, Multi-Model Inference 2020. version 1.43.17. Available online: www.cran.r-project.org/web/packages (accessed on 1 January 2019).
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Mackenzie, D.L.; Nichols, J.D.; Royle, J.A.; Pollock, K.H.; Bailey, L.L.; Hines, J.E. Occupancy Estimation and Modeling Inferring Patterns and Dynamics of Species Occurrence; Elsevier: London, UK, 2018. [Google Scholar]
- Mackenzie, D.I.; Nichols, J.D.; Lachman, G.B.; Droege, S.; Royle, J.A.; Langtimm, C.A. Estimating site occupancy rates when detection probabilities are less than one. Ecology 2002, 83, 2248–2255. [Google Scholar] [CrossRef]
- MacKenzie, D.I. PRESENCE User Manual. Duned. N. Z. Proteus Wildl. Res. Consult. 2012, 36, 1–78. [Google Scholar]
- Pita, R.; Morgado, R.; Moreira, F.; Mira, A.; Beja, P. Roads, forestry plantations and hedgerows affect badger occupancy in intensive Mediterranean farmland. Agric. Ecosyst. Environ. 2020, 289, 106721. [Google Scholar] [CrossRef]
- Virgós, E.; Romero, T.; Mangas, J.G. Factors determining “gaps” in the distribution of a small carnivore, the common genet (Genetta genetta), in central Spain. Can. J. Zool. 2001, 79, 1544–1551. [Google Scholar] [CrossRef]
- Ferreira-Rodriguez, N.; Pombal, M.A. Bait effectiveness in camera trap studies in the Iberian Peninsula. Mammal. Res. 2019, 64, 155–164. [Google Scholar] [CrossRef]
- Hegglin, D.; Bontadina, F.; Gloor, S.; Romer, J.; Müller, U.; Breitenmoser, U.; Deplazes, P. BAITING RED FOXES IN AN URBAN AREA: A CAMERA TRAP STUDY. J. Wildl. Manag. 2002, 68, 1010–1017. [Google Scholar] [CrossRef]
- Tews, J.; Brose, U.; Grimm, V.; Tielborger, K.; Wichmann, M.C.; Schwager, M.; Jeltsch, F. Animal species diversity driven by habitat heterogeneity/diversity: The importance of keystone structures. J. Biogeogr. 2004, 31, 79–92. [Google Scholar] [CrossRef] [Green Version]
- Mangas, J.G.; Lozano, J.; Cabezas-Díaz, S.; Virgós, E. The priority value of scrubland habitats for carnivore conservation in Mediterranean ecosystems. Biodivers. Conserv. 2008, 17, 43–51. [Google Scholar] [CrossRef]
- Šálek, M.; Červinka, J.; Pavluvčík, P.; Poláková, S.; Tkadlec, E. Forest-edge utilization by carnivores in relation to local and landscape habitat characteristics in central European farmland. Mamm. Biol. 2014, 79, 176–182. [Google Scholar] [CrossRef]
- Chapron, G.; Kaczensky, P.; Linnell, J.D.C.; Von Arx, M.; Huber, D.; Andrén, H.; López-Bao, J.V.; Adamec, M.; Álvares, F.; Anders, O.; et al. Recovery of large carnivores in Europe’s modern human-dominated landscapes. Science 2014, 346, 1517–1519. [Google Scholar] [CrossRef] [Green Version]
- Clinchy, M.; Zanette, L.Y.; Roberts, D.; Suraci, J.P.; Buesching, C.D.; Newman, C.; Macdonald, D.W. Fear of the human “super predator” far exceeds the fear of large carnivores in a model mesocarnivore. Behav. Ecol. 2016, 27, 1826–1832. [Google Scholar] [CrossRef] [Green Version]
- Suraci, J.P.; Smith, J.A.; Clinchy, M.; Zanette, L.Y.; Wilmers, C.C. Humans, but not their dogs, displace pumas from their kills: An experimental approach. Sci. Rep. 2019, 9, 12214. [Google Scholar] [CrossRef] [Green Version]
- Vanak, A.T.; Gompper, M.E. Dogs canis familiaris as carnivores: Their role and function in intraguild competition. Mamm. Rev. 2009, 39, 265–283. [Google Scholar] [CrossRef]
- Vilella, M.; Ferrandiz-Rovira, M.; Sayol, F. Coexistence of predators in time: Effects of season and prey availability on species activity within a Mediterranean carnivore guild. Ecol. Evol. 2020, 10, 11408–11422. [Google Scholar] [CrossRef]
- Carricondo-Sanchez, D.; Odden, M.; Kulkarni, A.; Vanak, A.T. Scale-dependent strategies for coexistence of mesocarnivores in human-dominated landscapes. Biotropica 2019, 51, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Torre, I.; Díaz, M. Small mammal abundance in Mediterranean post-fire habitats: A role for predators? Acta Oecologica 2004, 25, 137–142. [Google Scholar] [CrossRef]
- Oro, D.; Sanz-Aguilar, A.; Carbonell, F.; Grajera, J.; Torre, I. Multi-species prey dynamics influences local survival in resident and wintering generalist predators. Oecologia 2021, 197, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Torre, I.; Raspall, A.; Arrizabalaga, A.; Díaz, M. SEMICE: An unbiased and powerful monitoring protocol for small mammals in the Mediterranean Region. Mamm. Biol. 2018, 88, 161–167. [Google Scholar] [CrossRef]
- Goncalves, P.; Alcobia, S.; Simoes, L.; Santos-Reis, M. Effects of management options on mammal richness in a Mediterranean agro-silvo-pastoral system. Agrofor. Syst. 2012, 85, 383–395. [Google Scholar] [CrossRef]
- Palomares, F.; Delibes, M. Comparative ecology of the common genet Genetta genetta and the Egyptian mongoose Herpestes ichneumon Mammalia viverridae at Doñana SW Iberian peninsula. Bol. Real Soc. Esp. Hist. Nat. Secc. Biol. 1991, 87, 257–266. [Google Scholar]
- Palomares, F.; Delibes, M. Predation upon European rabbits and their use of open and closed patches in Mediterranean Habitats. Oikos 1997, 80, 407. [Google Scholar] [CrossRef]
- López-Bao, J.V.; González-Varo, J.P. Frugivory and spatial patterns of seed deposition by carnivorous mammals in anthropogenic landscapes: A multi-scale approach. PLoS ONE 2011, 6, e14569. [Google Scholar] [CrossRef] [PubMed]
- Escribano-Avila, G.; Calviño-Cancela, M.; Pías, B.; Virgós, E.; Valladares, F.; Escudero, A. Diverse guilds provide complementary dispersal services in a woodland expansion process after land abandonment. J. Appl. Ecol. 2014, 51, 1701–1711. [Google Scholar] [CrossRef] [Green Version]
- Valladares, F.; Benavides, R.; Rabasa, S.G.; Díaz, M.; Pausas, J.G.; Paula, S.; Simonson, W.D. Global change and Mediterranean forests: Current impacts and potential responses. In Forests and Global Change; Cambridge University Press: Cambridge, UK, 2014; pp. 47–76. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wild Carnivores (Richness) | Wild Carnivores | Vulpes vulpes | Martes foina | Meles meles | Genetta genetta | Canis lupus familiaris | Felis silvestris catus | |
|---|---|---|---|---|---|---|---|---|
| (Intercept) | −2.04 *** | −1.74 *** | −2.61 *** | −4.51 *** | −2.52 *** | −6.76 ** | −2.70 *** | −4.63 ** |
| (0.46) | (0.39) | (0.54) | (1.10) | (0.53) | (2.13) | (0.52) | (1.56) | |
| Attractant | 1.91 *** | 2.29 *** | 2.35 *** | 3.16 ** | _ | 2.42 * | 0.88 ** | _ |
| (0.49) | (0.41) | (0.54) | (1.12) | (1.13) | (0.31) | |||
| Season | 1.48 ** | 1.28 ** | 1.34 * | 1.90 | _ | 1.16 | 0.13 | 0.99 * |
| (0.49) | (0.42) | (0.56) | (1.17) | (0.91) | (0.33) | (0.46) | ||
| PC1 | 0.29 * | 0.30 * | -0.02 | _ | _ | _ | −0.30 | _ |
| (0.14) | (0.13) | (0.19) | (0.19) | |||||
| PC2 | 0.29 * | 0.37 * | 0.09 | 0.62 * | _ | _ | 0.92 * | 1.49 * |
| (0.14) | (0.16) | (0.25) | (0.28) | (0.42) | (0.65) | |||
| Attractant: Season | −1.47 ** | −1.50 ** | −1.89 ** | −2.11 | _ | _ | _ | _ |
| (0.56) | (0.56) | (0.72) | (1.43) | |||||
| Attractant:PC1 | −0.22 | −0.32** | −0.27 | _ | _ | _ | _ | _ |
| (0.16) | (0.12) | (0.13) | ||||||
| Season: PC2 | _ | _ | _ | _ | _ | _ | −0.75 * | −0.86 |
| (0.34) | (0.44) | |||||||
| PC1: PC2 | _ | _ | −0.49 ** | _ | _ | _ | _ | _ |
| (0.17) | ||||||||
| AICc | 262.40 | 392.18 | 284.40 | 185.16 | 135.82 | 71.87 | 194.54 | 145.04 |
| ∆AICc | 0.18 | 1.36 | 1.10 | 0.90 | 0.00 | 0.27 | 1.94 | 0.00 |
| AICc Weight | 0.17 | 0.24 | 0.10 | 0.21 | 0.18 | 0.24 | 0.05 | 0.73 |
| Pseudo-R² m. | 0.36 | 0.46 | 0.40 | 0.30 | 0.00 | 0.15 | 0.17 | 0.17 |
| Pseudo-R² c. | 0.39 | 0.70 | 0.63 | 0.60 | 0.28 | 0.56 | 0.53 | 0.99 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torre, I.; Pulido, T.; Vilella, M.; Díaz, M. Mesocarnivore Distribution along Gradients of Anthropogenic Disturbance in Mediterranean Landscapes. Diversity 2022, 14, 133. https://doi.org/10.3390/d14020133
Torre I, Pulido T, Vilella M, Díaz M. Mesocarnivore Distribution along Gradients of Anthropogenic Disturbance in Mediterranean Landscapes. Diversity. 2022; 14(2):133. https://doi.org/10.3390/d14020133
Chicago/Turabian StyleTorre, Ignasi, Tomàs Pulido, Marc Vilella, and Mario Díaz. 2022. "Mesocarnivore Distribution along Gradients of Anthropogenic Disturbance in Mediterranean Landscapes" Diversity 14, no. 2: 133. https://doi.org/10.3390/d14020133
APA StyleTorre, I., Pulido, T., Vilella, M., & Díaz, M. (2022). Mesocarnivore Distribution along Gradients of Anthropogenic Disturbance in Mediterranean Landscapes. Diversity, 14(2), 133. https://doi.org/10.3390/d14020133