1. Introduction
Hemichordates are a phylum of marine animals comprised of the solitary enteropneusts and the colonial, tubicolous pterobranchs; the latter include
Cephalodiscus and the graptolite clade, including thousands of species. Graptolites dominated the Palaeozoic seas. Their fossil record begins in the Middle Cambrian, followed by considerable radiation, and then almost disappeared in the Middle Carboniferous. Fossils consist of the decay-resistant tubaria, used as stratigraphy index fossils.
Rhabdopleura is a graptolite with fossil members dating from the Middle Cambrian to five living species, making it one of the longest-lived genera in Earth’s history. Its most distinguishing feature is the tubes that form a tubarium, the housing construction of Pterobranchia [
1]. Most of what is known about tube and tubarium development are from a few observations of living
Rhabdopleura. A colony starts from a larva that secretes a dome (or prosiculum), where it metamorphoses into the foundational sicular zooid. The zooid emerges from the prosiculum and then secretes, using glandular cells on the ventral cephalic shield, the first tube of the colony [
2,
3]. Stebbing [
2] provided information on the early steps of tubarium development in
R. compacta. The sicular zooid is attached to the dome with the contractile stalk. From this stalk, a single bud develops into a second zooid. The two zooids are then separated by the secretion of a septum that is perforated with a hole for the growth of the stolon system. Further asexual development is not clear, but during colony growth, a “naked” stolon (or gymnocaulus) is produced by a terminal zooid. The gymnocaulus forms new zooid buds before it hardens into a sclerotized black stolon [
4]. Each zooid may contribute a new tube, or tubes, to the colony, resulting in a tubarium that is flexible and transparent, tending to brown with age [
5]. The tube composition is still debated, although recent evidence corroborate the hypothesis of a chitin-like major component (Beli unpublished observations). Tubes may be creeping or erect and are made of rings and half rings called fuselli [
6,
7]. An individual tube may be used by several generations of zooids. The tubarium, with some exceptions [
8], is usually a few millimeters to a few centimeters in size. Fossil graptolites, except for a few tentative zooid impressions [
9,
10,
11,
12,
13], consist entirely of tubes and tubaria. The tubaria are among the most extraordinary examples of diverse extracellular secreted structures in nature [
4,
14] that take a dizzying array of forms, particularly among the planktonic Graptoloidea, which radiated from the Early Ordovician through the Silurian period, and then went extinct in the early Devonian. The earliest forms, including rhabdopleurids [
7,
15], were benthic encrusting Dendroids that had creeping and erect tubes.
Rhabdopleura, though a modern species, belongs to this primitive fossil clade. Its tubaria are less complex than those of the planktonic graptolites.
An enduring challenge for palaeontologists is the species definition of fossils based on morphology. Fossils provide no evidence of reproductive isolation that is frequently used by biologists to define a living species [
16], and in the absence of genetic data, there is no way to determine heritable characteristics, including variation, from non-heritable ones. Among the thousands of graptolite species, the family to genus level taxonomy is well established [
14]. Graptolites are diverse and variable in form; taxonomic definitions of early benthic graptolite species, including rhabdopleurids, however, can be narrow [
17]. These minor morphological differences are a conundrum to graptolite taxonomists who must decide on the validity of tube traits to define a species, not knowing if differences are due to taphonomy, genetic variation of the species, or environmentally induced phenotypic plasticity [
5,
18,
19,
20,
21,
22,
23]. In marine environments, phenotypic plasticity in response to water flow velocity is a potent, selective force [
24,
25,
26] due to biomechanical drag. Dramatic phenotypic plastic responses to flow have been documented in the ostia of sponges [
27], barnacle legs [
26], and the tube feet of sea urchins [
28]. Water flow constrains the function of the acorn worm pharynx [
29]. Benthic graptolites like
Rhabdopleura may also be expected to show plasticity in the tube length and number in response to flow. The recent discovery that
Rhabdopleura is a graptolite [
30] allows us to experimentally test the plastic response to tube length and number in response to water flow velocity to shed light on our confidence of minor taxonomic characters used on Dendroid graptolite species designations and phylogenetic character decisions.
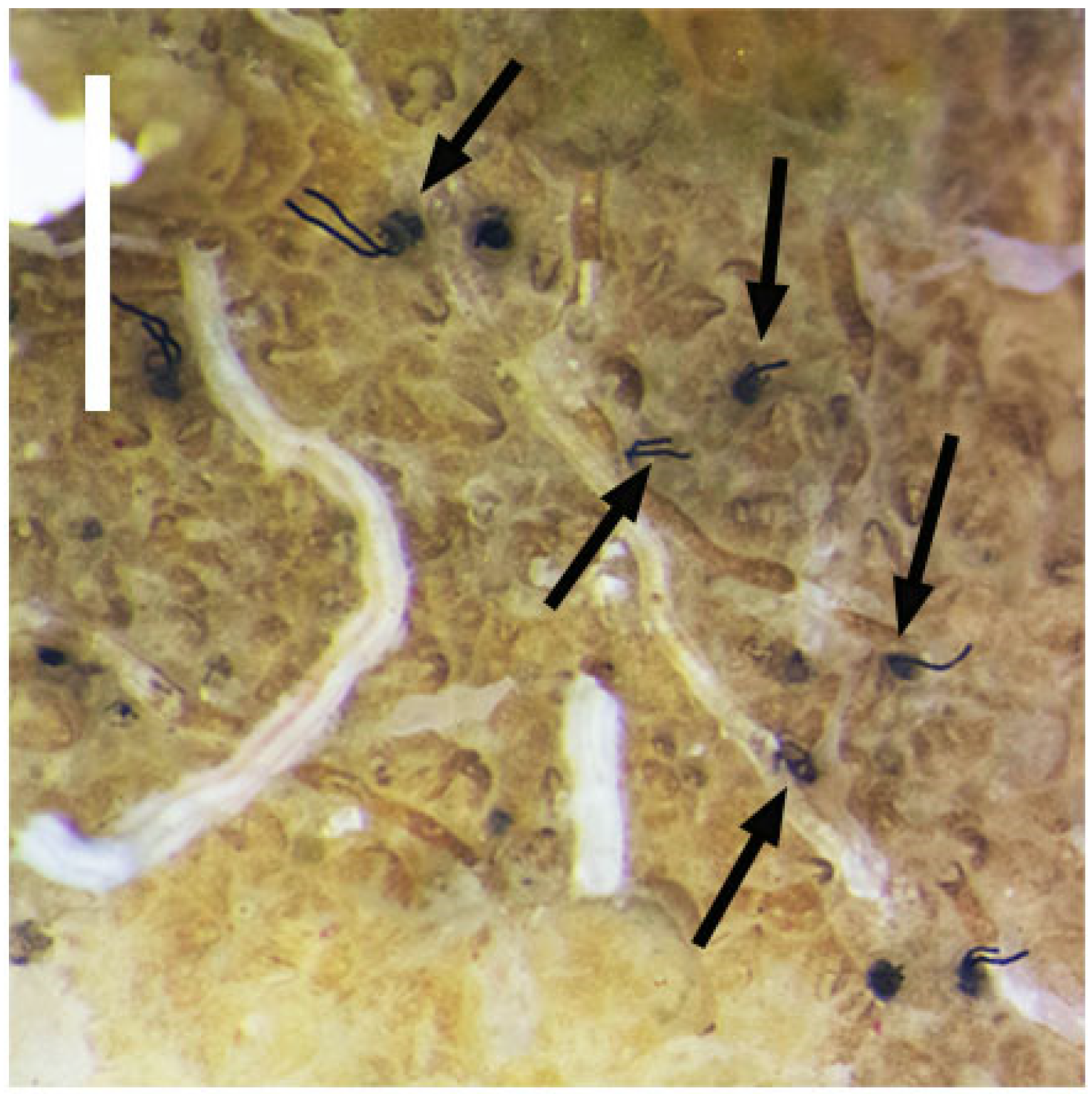
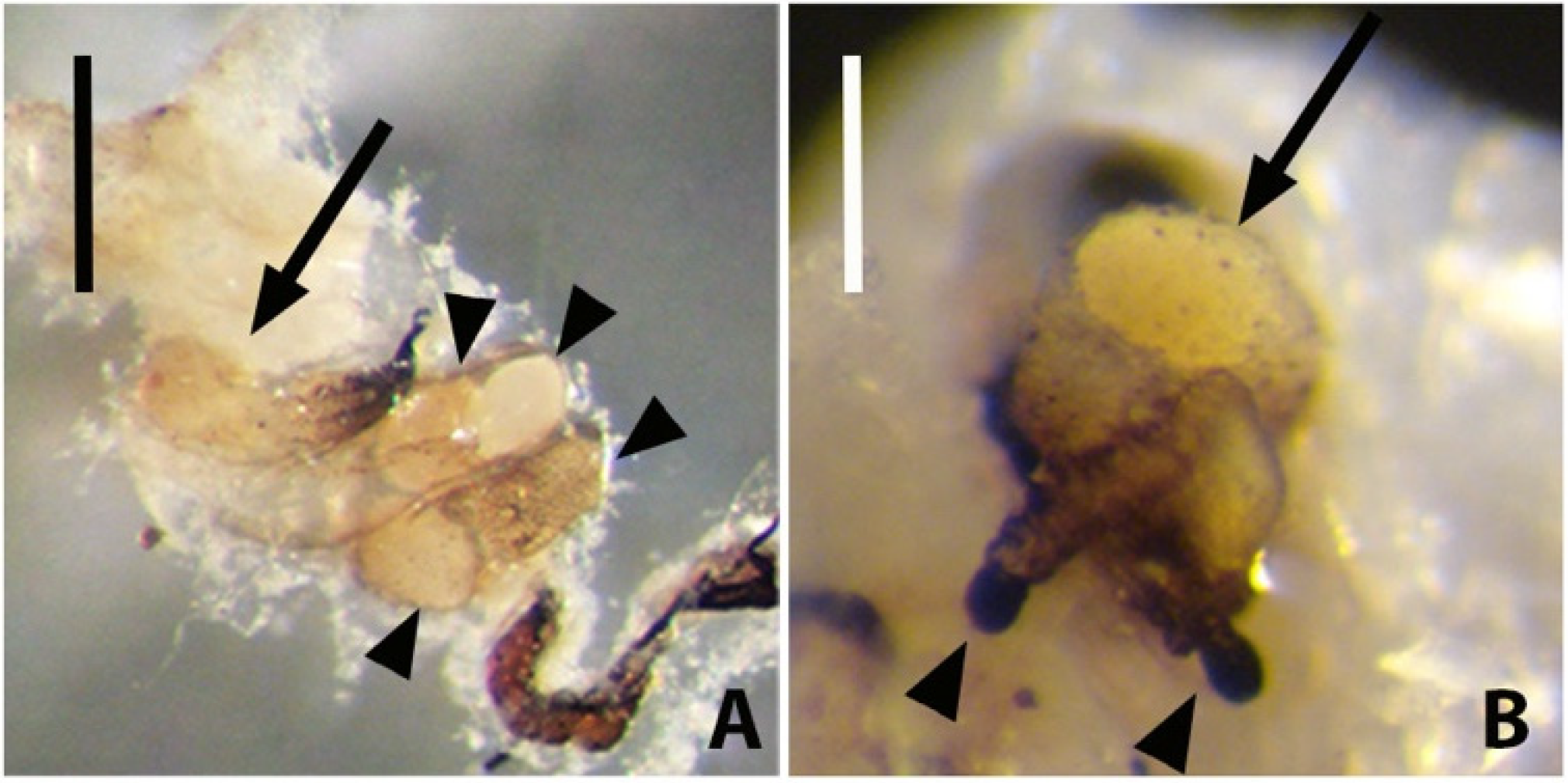
Rhabdopleura recondita Beli, Cameron, Piraino, 2018 is a benthic graptolite discovered on the southern Italian coasts, but it is possibly distributed throughout the Western Mediterranean basin [
31]. It is an atypical rhabdopleurid in that the colonies reside inside of the calcareous zoarium of dead bryozoan colonies, such as
Myriapora truncata (Pallas, 1766) and
Schizoretepora serratimargo Hincks, 1886. The burrows and the zooecia (i.e., the skeletal body walls of chambers previously inhabited by the bryozoan zooids) in the bryozoan skeleton (or zoarium) provide habitat for the
R. recondita zooids, whose interconnecting creeping tubes are smooth and lack fuselli. The erect tubes are a few millimeters in length and extend perpendicularly from the bryozoan zoarium into the water column; they are typically graptolite-like, with fuselli (
Figure 1). Here we make observations and experiments on the development and tubes of
R. recondita kept in captivity. We observe (i) dome secretion, larval metamorphosis, tube secretion, and tube repair by adult zooids, and (ii) we experimentally evaluate the plastic phenotypic response of the number of tubes and tube form to different water flow velocities.
2. Materials and Methods
Rhabdopleura recondita colonies are not visible to the naked eye because they are concealed inside clumps of dead bryozoan zoaria of S. serratimargo and M. truncata. These clumps were collected by hand by SCUBA diving between 10–20 m depth along the Salento coast, south of Otranto, Italy. They were transported in thermally insulated coolers to the University of Salento, where they were kept in an aquarium at 19 °C, the temperature of the collection site. Colonies were kept at a 12-h photoperiod alternating between light and dark.
2.1. Larval and Adult Tube Building
Larvae, zooids, and tube-building behavior were observed daily with a Leica MZ6 ocular stereo microscope, and pictures were taken with a Sony-RX100-I camera. The search for larvae occurred by breaking the bryozoan skeletons using steel probes and chisels. We found six larvae which were recovered with a pipette and transferred to six water bowls with 3 mL of filtered seawater at 19 °C. The bowls were covered with a nylon mesh and immersed together in a one-liter vessel of seawater with an aerator. The vessel also held fragments of colonized bryozoans. (i) Five larvae, one per bowl, were observed without a substrate to evaluate their ability to settle, metamorphose and build a colony, and (ii) one larva was provided with a deeply fragmented M. truncata to let it feel inside a host allowing external observation instead.
Rhabdopleura recondita zooids were collected in the same manner as the larvae and placed into bowls to evaluate their ability to build tubes and colonies. We prepared a total of five bowls, each covered with a nylon mesh. Zooids were fed coral food twice a week (H&O by Sicce) and kept together in a one-liter vessel with an aerator. (i) Into three separate bowls, to observe progress in tube building, we isolated one developing zooid and two adult zooids exposed naked, which means without an erect tube, but with attached stolon, creeping tube, and bryozoan calcium carbonate zoarium where they could not shelter. (ii) In the fourth bowl, we observed the behavior of two zooids exposed partially naked, attached to a common stolon, and with a reduced capacity to shelter inside the bryozoan zoarium. (iii) In a fifth bowl, to observe the ability of zooids to secrete new tubes and the timing of tube development, we cut all 23 standing tubes from a 1.5 cm bryozoan fragment containing an R. recondita colony and observed it over time.
Finally, one colony was fixed in alcohol and placed in vinegar overnight to remove the calcium carbonate matrix of the bryozoan skeleton. This treatment made it possible to observe the colony in the absence of the host skeleton.
2.2. Flow-Induced Response
To test the plastic phenotypic response of R. recondita tube secretion and development to different water velocities, a preliminary trial was performed for four colonies deprived of tubes and exposed to 4.8 cm/s.
Following the preliminary experiment, 16 colony fragments were put in lower velocity channels to form tubes. The channel velocities were established based on velocities measured at the collection sites as reported in the Istituto Superiore per la Protezione e Ricerca Ambientale (ISPRA, 2012
http://www.strategiamarina.isprambiente.it/consultazioni/consultazione-2012/files/3.1%20mar_adr%20caratteristiche%20fisiche.pdf accessed on 5 September 2016). Colony fragments with abundant
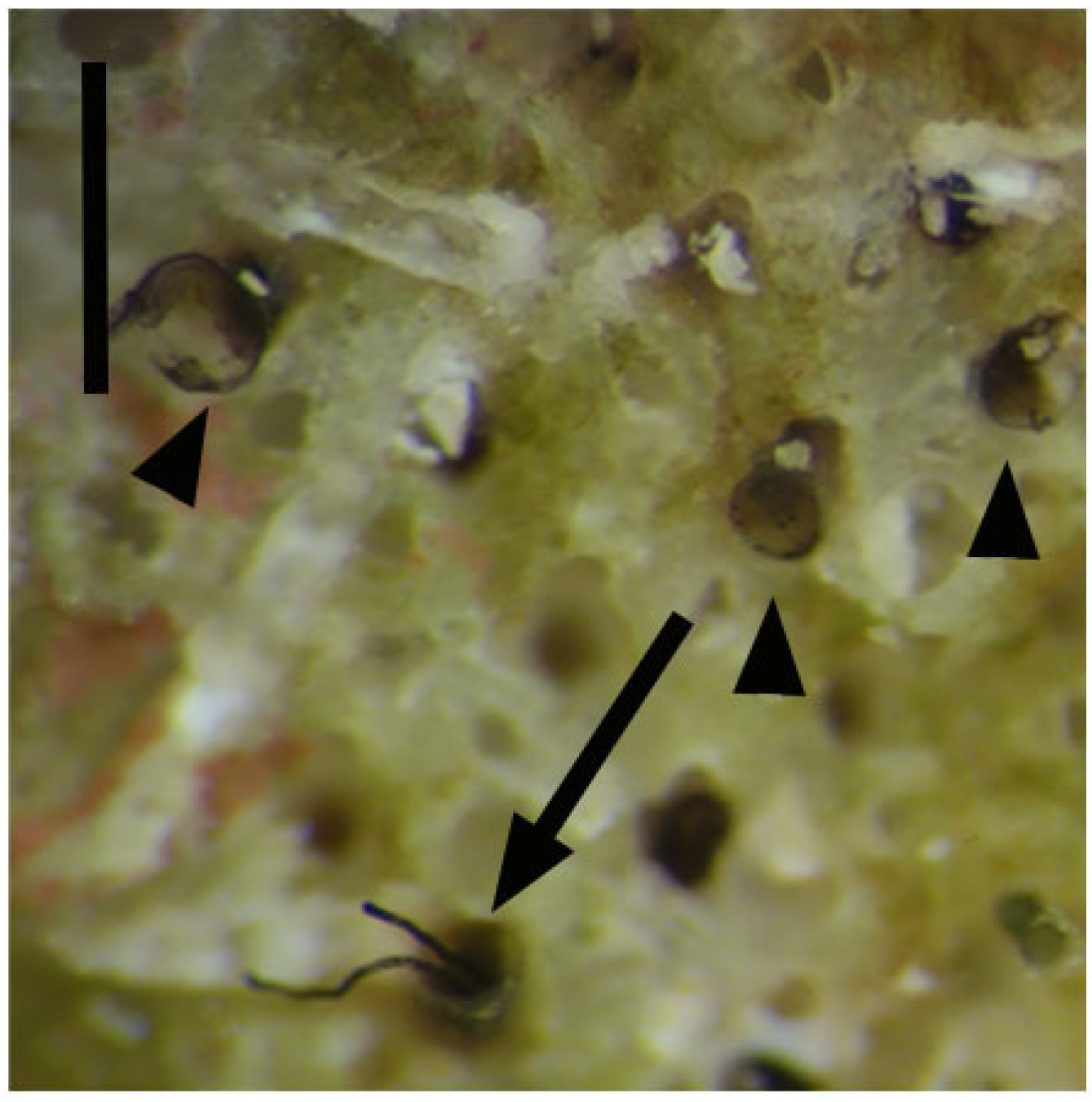
R. recondita tubes and zooids were chosen, though the number of zooids inside of any fragments was unknown because of the hidden nature of zooids. Images were captured with an SMZ 25 stereo microscope equipped with a DS-Ri2 video camera and a video-interactive image analysis system NIS–Elements BR 4.30.02 Nikon Instruments software, and with a Sony RX100 digital camera. Colonies were drawn with particular attention paid to the number and position of erect tubes. The tubes were then removed using tweezers, measured, washed in filtered seawater, and preserved in Picric Acid-Formaldehyde-Glutaraldehyde (PAFG).
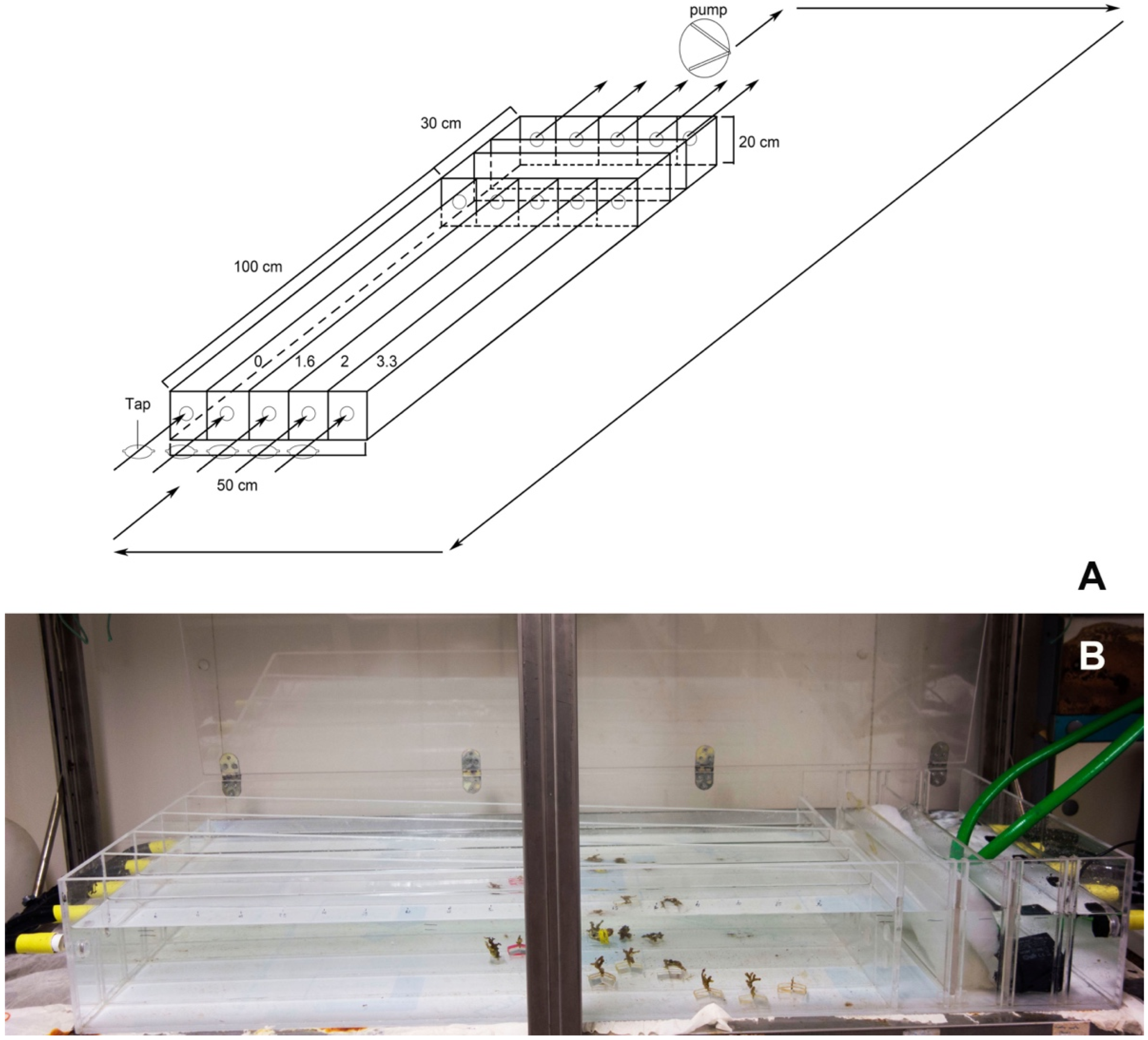
These 16 colonies, denuded of erect tubes (
Figure 2), were then placed into flow channels with one of four experimental flow velocities. Colonies were numbered from 1 to 16 based on the flow velocity treatment. Numbers 1–4 were maintained at 1.6 cm/s; 5–8 were maintained at 2 cm/s; 9–12 were maintained at 3.3 cm/s, and the control colonies 13–16 were maintained at 0 cm/s.
The flow channel aquarium was equipped with a 12,300 L/h pump, filtering and refrigerating system, and taps to regulate the flow velocities through four parallel channels (
Figure 3A,B). Filtered seawater from the
Rhabdopleura collection site was maintained at the collection site temperature of 19 °C. Salinity, dissolved oxygen, and pH were checked weekly and kept at 38‰, 7 mg/L, and 8.2, respectively. Distilled water was sometimes added to counter evaporation. Full spectrum fluorescent bulbs were set to a 12-h daylight photoperiod. The precise flow velocities, controlled with taps, were established using a fluorescein dye. The channel velocities were 1.6 cm/s, 2 cm/s, and 3.3 cm/s.
Four colonies were placed in each channel where the flow was laminar. The control channel at the velocity of 0 cm/s had a drip that prevented stagnation, though no flow could be detected. To minimize a boundary layer effect, colonies were attached to a weighted Plexiglas base that was 1cm deep and projected the fragments up into the center of the flow channel (
Figure 4). Animals were fed twice a week with coral food (H&O by Sicce) and allowed to secrete new tubes for 40 days. The progress of tube construction was observed and photographed every eight days (i.e., days 0, 8, 16, 24, 32, and 40).
2.3. Statistical Analysis
Initially, the data related to the number of new tubes formed and the length of tubes (original and new tubes) were tested for homogeneity and normality using graphical analysis and the Shapiro-Wilk [
32] test. No data tested fit the assumptions of normality, and non-parametric tests were performed. We used the Kruskal-Wallis [
33] analysis to test if (i) the number of new tubes formed differed significantly between water velocity treatments and (ii) the new tube lengths differed significantly between velocity treatments. If the result of the Kruskal-Wallis were significant, the Dunn [
34] post-hoc test was performed to highlight the significance of the differences between treatments.
Additionally, we tested if there were differences between the new tubes’ lengths and the original pre-tweezered tube lengths with respect to velocity. To test if there was a difference in tube length before and after the experiment (new length versus original length) and, if different, was it conditioned to the velocity treatment, we performed a General Linear Models (GLMs) test. First, a complete model was created using the new tube length as a response variable and the original length and speed treatment as explanatory variables. With the model created, a family selection was made to determine the best distributional family for this model. To select the family, we compared them with the Akaike criteria. With the best family selected, the model was tested to highlight the significance, if there was any. In addition, the adjustment for the model was observed using graphs (model vs. residuals). All analyses were performed using the R software [
35]. The level of significance adopted in this study was 5%.
4. Discussion
Rhabdopleura recondita is unusual in that it inhabits the interior empty burrows and zooecia (skeletal holes) of dead bryozoan colonies [
31].
Zooids and larvae were obtained by fracturing this calcium carbonate matrix, and our best observations on the tube structure inside of the skeletal followed acid digestion of the matrix. We previously reported how the burrows and zooecia walls of the bryozoan zoaria are coated with cortical layers of tube material without fuselli [
31]. Here we add to that an observation of an
R. recondita brood chamber, which is known for
R. normani [
36], and one with a female zooid, embryos, and larvae for
R. compacta [
37]. The brood chamber of
R. recondita had four larvae at different stages of maturity, demonstrating asynchronous brood development. The most mature larva was positioned the most distant from the tube aperture. Like
R. compacta, it would need to squeeze past earlier-stage larvae and the parent zooid to exit the colony tubarium [
38].
Fertilization and early cleavage of pterobranch embryos [
36] and observations of larvae are exceedingly rare [
2,
37,
38] so any new information, including rearing conditions, is valuable. Our observations and experiments with the larvae and zooids highlight the adaptations and constraints to the hidden life of
R. recondita. Congeneric species typically develop on exposed surfaces, frequently of calcium carbonate [
31], though occasionally they occupy internal cavities [
39,
40,
41,
42,
43]. During this study, we did not observe
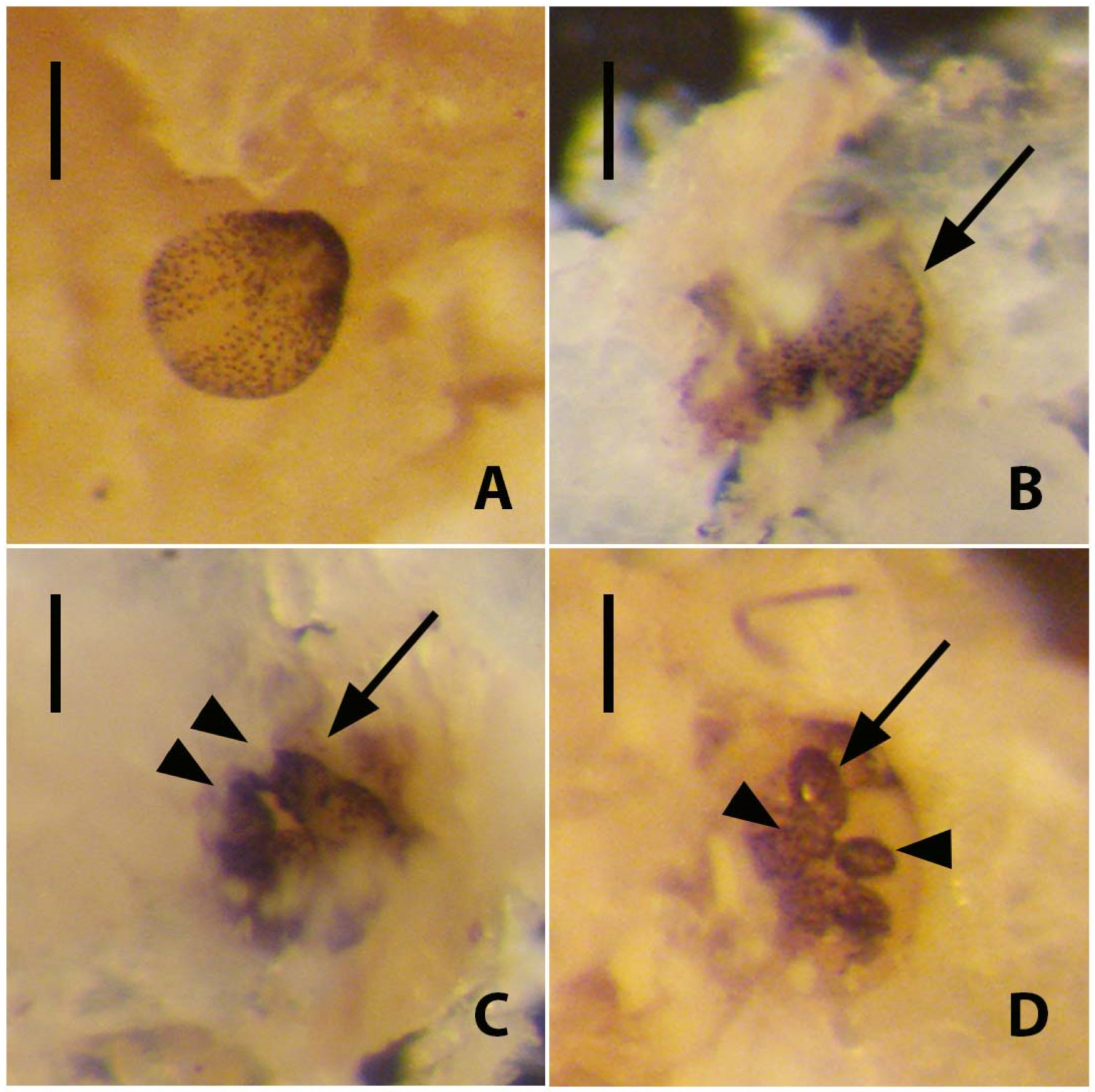
R. recondita building a new tubarium or enlarging a pre-existing one outside of the bryozoan host zoarium. Of the larvae that settled in the absence of bryozoan substrate, some began to secrete a dome (
Figure 5A,B) and swam to the start of metamorphosis, like
R. normani, but only one achieved metamorphosis, suggesting that the host matrix increases normal settlement and metamorphosis (
Figure 5A). This truncation to development and dome secretion may have been due to disturbing the larvae or the lack of ideal developmental conditions [
36], and the absence of a bryozoan zoarium seems most likely. Only one larva without the zoarium substratum settled and completed metamorphosis within 10 days (
Figure 5A). By contrast, the larva in a bowl with bryozoan fragments secreted a dome and metamorphosed after 25 days (
Figure 6). In this case, following its metamorphosis, additional larvae emerged from the fragments, aggregated, settled, and metamorphosed, all in the presence of CaCO
3 substrate. This result supports the finding that host fragments may increase settlement and metamorphosis and that larvae hidden inside the bryozoan zoarium are best positioned for normal development.
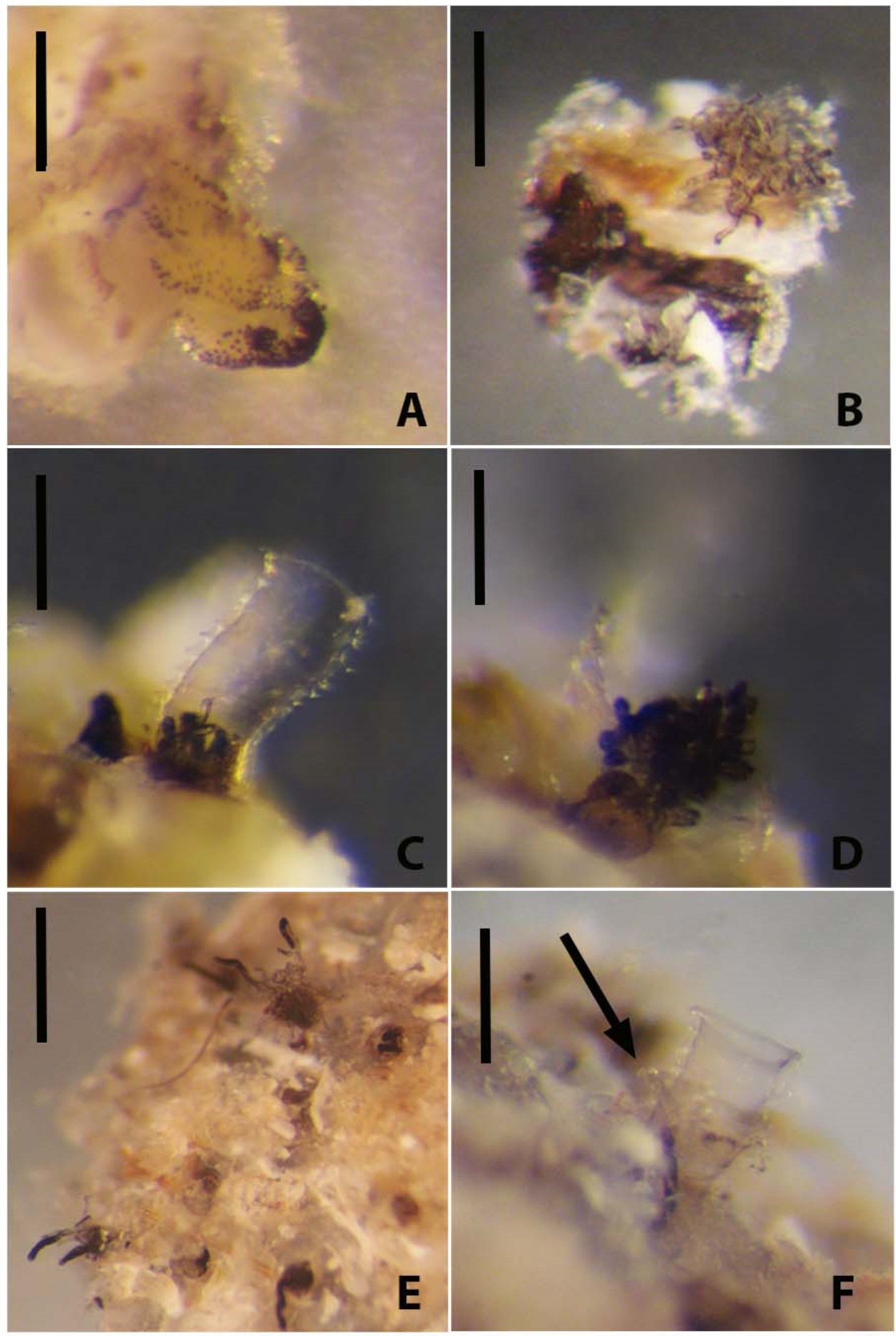
Three experiments were done to observe tube building by adult zooids. Two bowls each held an isolated adult zooid, and a third bowl held an immature zooid. These zooids did not reform a new tube, the immature zooid did not develop further, and none formed a stolon or budded new zooids (
Figure 7A,B). They survived for few hours or days, supporting the idea that the absence of other connected zooids and a tubarium are incompatible with life [
14]. The two zooids connected by a common stolon and cultured in the presence of host zoarium fragments survived and secreted erect tubes with fuselli (
Figure 7C,D). One zooid moved marginally deeper inside the tubarium and built a normally shaped erect tube with fuselli (
Figure 7C), whereas a more exposed zooid (
Figure 7D) secreted a distally flared and abnormally shaped tube. This difference in tube form may suggest that normal tube secretion depends not only on the host material but access to interior burrows of the bryozoan zoarium to build a tube. These observations suggest that
R. recondita tube secretion and colony formation are adapted and constrained to life inside an empty bryozoan host zoarium.
In a fifth bowl, a fragment of bryozoan zoarium contained
R. recondita zooids that extended their arms for filter feeding (
Figure 7E). These zooids’ erect tubes were removed, which did not appear to disturb normal emergence and feeding activities. Of these zooids, only three of the 23 re-secreted a new tube, like what we found in colonies that were subjected to experimental flow velocities. This dependence on the host zoarium for normal tube secretion and the low ratio of zooids that re-secrete tubes posed challenges to our experiments on the plastic response to tube secretion in response to water flow velocity. Also, because most of the tubarium astogeny, or overall colony form, is inside of the host zoarium, and the standing tube spacing is constrained by the availability of channels and exit pores from the zoarium, our experiments on induced plasticity were limited to measurements of standing tube number and lengths. Despite these limitations, we conducted the standing tube plasticity experiments. With five extant species of rhabdopleurid known,
R. recondita is the only one with extensive colonies that can be readily collected and maintained in the lab.
This first experimental attempt of induced phenotypic plasticity in
R. recondita did not find the expected outcome, that longer tubes would be re-secreted at low flow velocities and shorter tubes at higher velocities, similar to what has been reported for sponge ostia, barnacle legs, and regenerated tube feet of sea urchins [
26,
27,
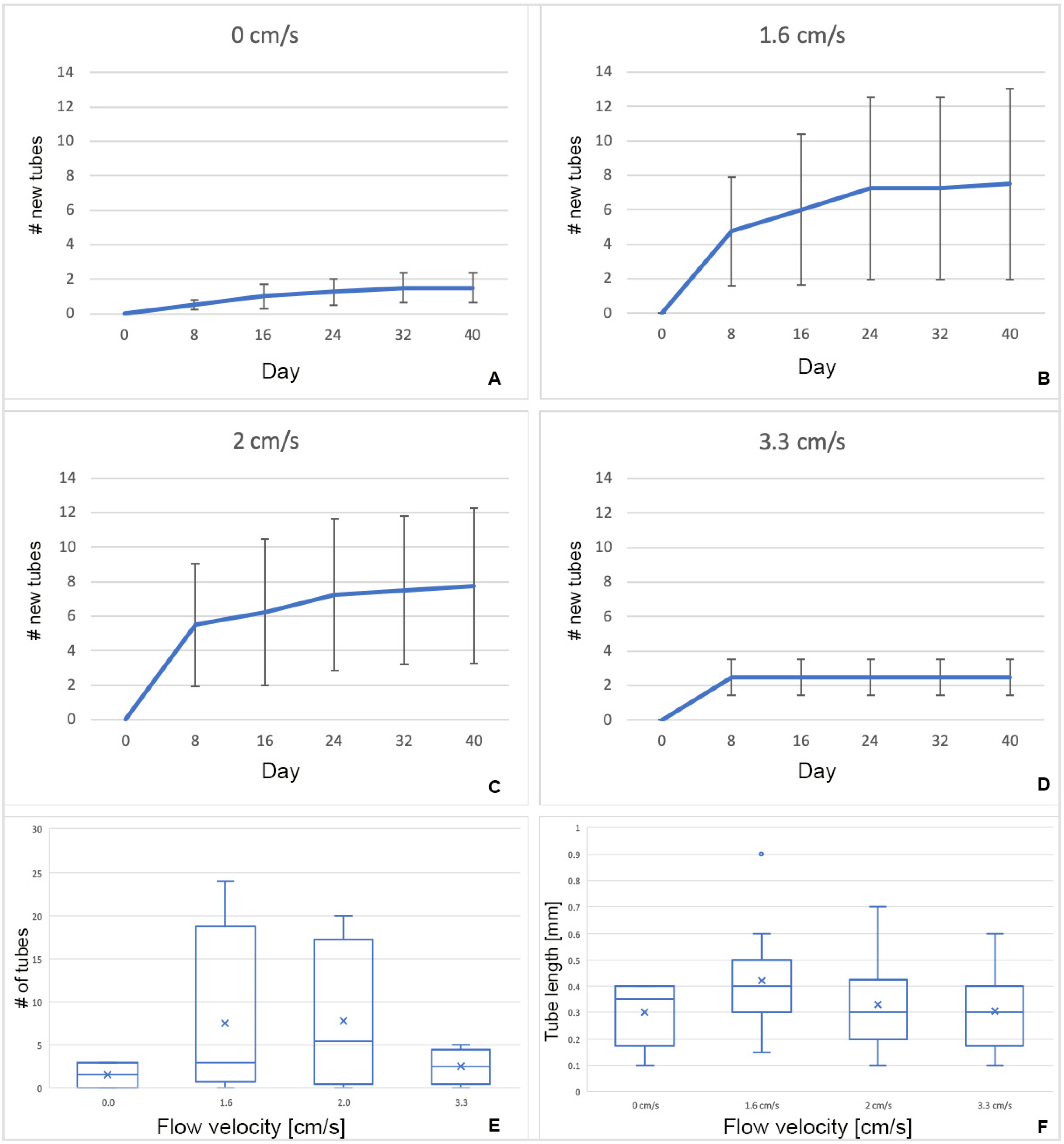
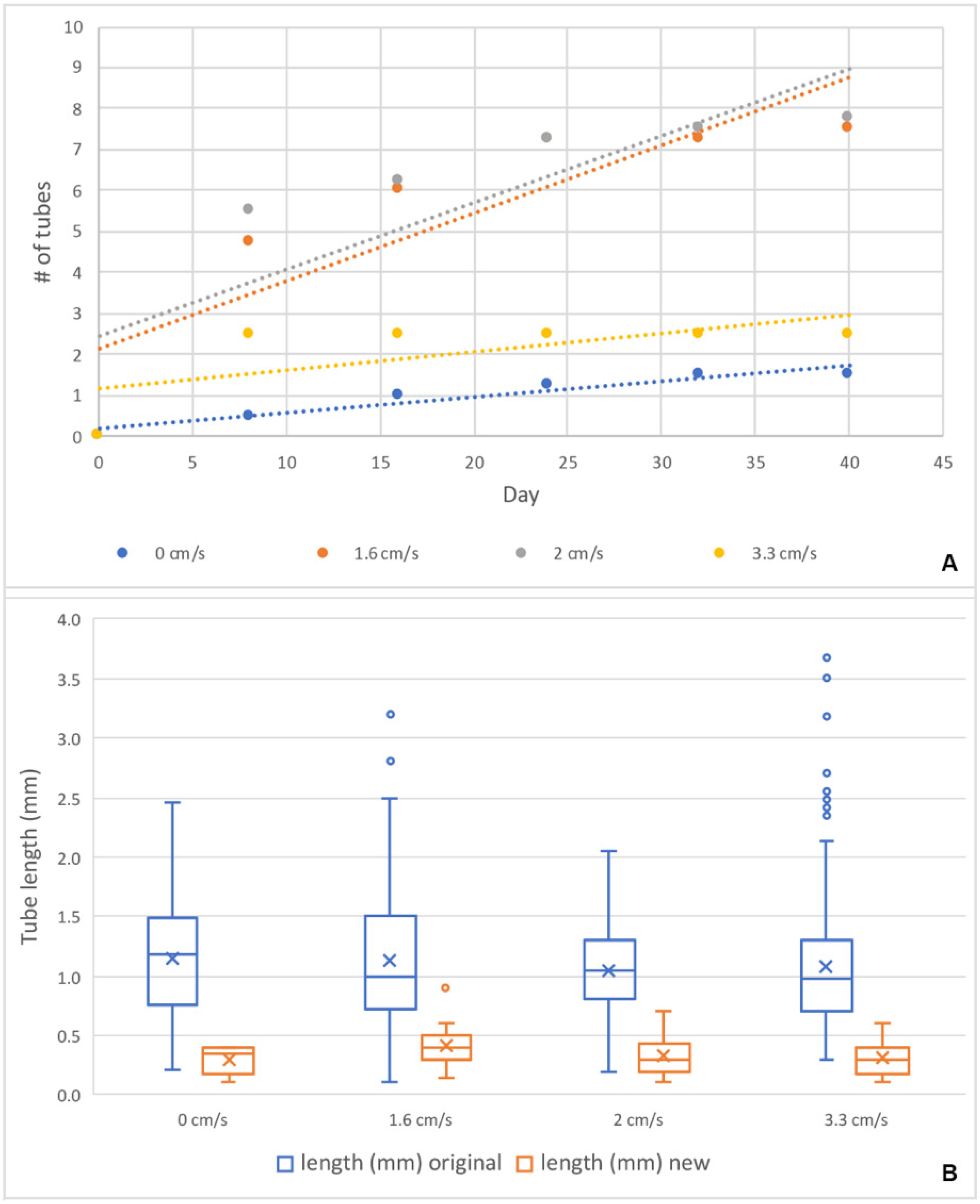
28]. Our preliminary experiment at 4.8 cm/s completely arrested the activity of zooids, and no tube building was done. Further experiments at lower velocities found a tendency for a higher number of tubes to build at intermediate flow velocities and no difference in newly secreted tube length. The number of new erect tubes formed by zooids of
R. recondita exposed to different flow velocities (0, 1.6, 2, and 3.3 cm/s) showed a higher regrowth and potential for growing (
Figure 10E and
Figure 11) at 1.6 and 2 cm/s, though these differences were not statistically significant. More significant results may be obtained with a larger sample size, but we think it is unlikely. Our results do show that
R. recondita lives in a low and narrow range of flow velocity, and within this range, it does not significantly vary the number of tubes re-secreted or tube length. Despite finding no difference in the length of re-secreted tubes among the four velocity treatments, there was a significant length difference between the originally collected colony tubes and the tubes re-secreted after dissection (
Figure 11B). Standing tubes of colonies collected from the Adriatic Sea were longer and more variable than those secreted in the lab (
Figure 11B). Assuming that the tubarium where tubes were removed housed a zooid, the low re-secretion ratio and short lengths may be explained by one of six possibilities. (i) The food that we used, developed for corals, was too limited or nutritionally inadequate for ideal colony growth. Nothing is known about culturing
Rhabdopleura in captivity [
44]. Alternatively, the coral food may have been so abundant that secreting a new tube was unnecessary. (ii) Tube secretion and colony enlargement may be seasonal or sporadic [
44]. (iii) Tube formation is rare because tubes are used from generation to generation [
2,
45,
46]. (iv) The boundary layer effect may have exposed zooids to lower velocities than those set in each flow velocity channel, which in turn may have reduced the gas exchange. (v) Colonies may require much more time, even years, to re-secrete in terms of number and length [
47]. (vi) Stress caused by the loss of all the tubes in each colony and the energy supply required for their reconstruction. We observed zooids that lacked erect tubes emerged from the zoarium to feed, but perhaps they did not have the energy to secrete new tubes. At the terminal end of some excised tubes, we observed a lid on the aperture. These lids are a novel finding, but as to the function, we can only speculate. Rhabdopleurids can produce dormant buds to overcome adverse periods [
2,
7], and these lids may be the sign of zooids’ encystment to face stressful conditions caused by tube removal and aquarium environment.
These results show that the
R. recondita tube number and length exhibit no significant variation in response to water flow velocity. Together, the results of the preliminary experiment, where we found no tube secretion in flow velocities at 4.8 cm/s, suggest that
R. recondita inhabits a low and narrow water flow velocity range. Tube development appears constrained. This suggests that small differences that distinguish early encrusting fossil graptolite species are real. Of course, other living rhabdopleurids, especially those with tubaria that are free of a host, may have a plastic response to flow, food, predation, or temperature. This logic extends to Palaeozoic fossil rhabdopleurids. The developmental toolkit, including plasticity, of early graptolites may have been differed from that of
R. recondita. Peterson et al. [
48] hypothesized that an accumulation of microRNAs over geological time may have increased the silencing and post-translational regulation of gene expression, limiting developmental experimentation. We see no reason that this hypothesis cannot be extended from secreted skeletal matrices (i.e., exoskeletons and endoskeletons) to tube matrices. Others have quantified the number and variation of graptolite forms over time and concluded that these are largely determined by extinctions rather than developmental constraints [
49]. Our results suggest that the tube form of
R. recondita may, in fact, be constrained, but we are cautious about extending this finding to other fossil or extant species. It is conceivable that some graptolite species and clades exhibited tremendous tubarium evolvability. With respect to
Rhabdopleura, its evolvability has nothing to do with survivability. It is among the oldest graptolite lineage that has survived five major extinction events, and it is the only graptolite genus that survives to date.



 ,
, 













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}