Abstract
The click beetles (Elateridae) originated in the Mesozoic and recently form a relatively large family with approximately 10,000 described species worldwide. However, the Mesozoic, and particularly Cretaceous, click beetle fauna remains very poorly known. Here we describe Burmogonus cretaceus gen. et sp. nov. based on a single, relatively well-preserved, specimen from the mid-Cretaceous Burmese amber. This species can be assigned with confidence to the subfamily Elaterinae, and based on the supra-antennal carinae being incomplete across the head and directed to the labrum, the shape of metacoxal plates, and simple tarsi, we tentatively place it in the tribe Elaterini. We discuss the morphology of a new genus and other Elaterinae described from Burmese amber.
1. Introduction
The fossil record of Elateridae (click beetles) includes 264 species classified in 102 genera and 9 subfamilies [1,2,3,4,5]. More than half of the fossil species diversity has been described from the Jurassic Karatau [6,7,8], although at least some taxa might belong to a different family [2,9,10]. Considering high diversity of the described click beetles in Jurassic (113 species), it is surprising that only 27 species are known from the Cretaceous period [2,3,4,5]. Eight Cretaceous deposits with described Elateridae are in the Lulworth Formation in England, Mirsanovo and Zaza Formations in Russia, Yixian Formation in China, Jinju Formation in South Korea, Ajka Coal Formation in Hungary, Kzyl-Zhar locality in Kazakhstan, and the amber-rich Hukawng Valley in northern Myanmar (e.g., [2,4,11,12,13,14,15]).
Only five click beetle species have been recorded from the mid-Cretaceous amber of Myanmar to date [3,5,16,17], despite the fact that this deposit has turned out to be an extraordinarily rich source of extinct organisms since it was rediscovered (e.g., [18,19,20,21]). In this study, we describe a new genus and species of click beetles based on a relatively well-preserved specimen from Burmese amber. It belongs to the subfamily Elaterinae and we tentatively place it in the tribe Elaterini based on its supra-antennal carinae being incomplete across the head, metacoxal plates that are only weakly narrowed outwards, and simple tarsi.
2. Materials and Methods
The amber piece was polished by hand, allowing for improved views of the included specimen, and was not subjected to any additional treatment. Photographs were taken by a digital camera mounted on a stereomicroscope Olympus SZX12. Stacks of photographs were combined with the software Helicon Focus Pro (version 7.6.4, Helicon Soft Ltd., Kharkiv, Ukraine), applying the rendering method ‘depth map’ or ‘weighted average’. Adobe Photoshop CC (version 2019-20.0.5) was used for the noise reduction, sharpening, and contrast enhancing, as well as for the assembly of the final image plates.
Morphological terminology follows Costa et al. [22] and Kundrata et al. [3], and the Elateridae classification follows Douglas et al. [23]. Body length of the examined specimen was measured from the frons to apex of elytra, body width at the widest part of the body, pronotal length at midline, and pronotal width at the widest part. The holotype is deposited in the collection of the Department of Palaeontology of the National Museum, Prague, Czech Republic (NMPC). The ZooBank LSID number for this publication is: urn:lsid:zoobank.org:pub:75DD83CF-A69F-4431-930D-F6344AF9BC7E.
3. Results
Systematic Palaeontology
- Order Coleoptera Linnaeus, 1758
- Suborder Polyphaga Emery, 1886
- Series Elateriformia Crowson, 1960
- Superfamily Elateroidea Leach, 1815
- Family Elateridae Leach, 1815
- Subfamily Elaterinae Leach, 1815
- Tribe Elaterini Leach, 1815
- Genus Burmogonus gen. nov.
 Figure 1. Morphology of Burmogonus cretaceus gen. et sp. nov. (A) Habitus, dorsal view; (B) habitus, ventral view. Scale bars: 2.0 mm.
Figure 1. Morphology of Burmogonus cretaceus gen. et sp. nov. (A) Habitus, dorsal view; (B) habitus, ventral view. Scale bars: 2.0 mm. Figure 2. Morphology of Burmogonus cretaceus gen. et sp. nov. (A) Head and anterior part of prothorax, dorsal view; (B) head and anterior part of prothorax, ventral view; (C) head, pronotum, scutellar shield, and base of elytra, dorsal view; (D) head, prothorax, and mesothorax, ventral view; (E) left antenna, ventral view; (F) right mesoleg, dorsal view; (G) right pro- and mesoleg, ventral view; (H) metacoxal plates, ventral view; (I) last four abdominal ventrites, ventral view. Scale bars: 0.5 mm.
Figure 2. Morphology of Burmogonus cretaceus gen. et sp. nov. (A) Head and anterior part of prothorax, dorsal view; (B) head and anterior part of prothorax, ventral view; (C) head, pronotum, scutellar shield, and base of elytra, dorsal view; (D) head, prothorax, and mesothorax, ventral view; (E) left antenna, ventral view; (F) right mesoleg, dorsal view; (G) right pro- and mesoleg, ventral view; (H) metacoxal plates, ventral view; (I) last four abdominal ventrites, ventral view. Scale bars: 0.5 mm.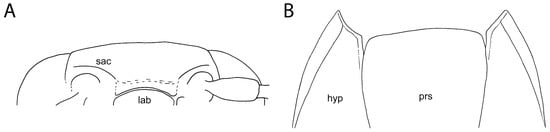 Figure 3. Morphology of Burmogonus cretaceus gen. et sp. nov., drawings. (A) Anterior part of head, frontal view; (B) anterior portion of prothorax with shortly open pronotosternal sutures, ventral view. lab, labrum; hyp, hypomeron; prs, prosternum; sac, supra-antennal carina. Not to scale.
Figure 3. Morphology of Burmogonus cretaceus gen. et sp. nov., drawings. (A) Anterior part of head, frontal view; (B) anterior portion of prothorax with shortly open pronotosternal sutures, ventral view. lab, labrum; hyp, hypomeron; prs, prosternum; sac, supra-antennal carina. Not to scale.
Type species. Burmogonus cretaceus sp. nov., here designated.
Etymology. Derived from the word “burmite” (mid-Cretaceous amber of Myanmar) and part of the word “Brachygonus”, which is a genus in Elateridae: Elaterinae superficially resembling Burmogonus gen. nov. in some aspects. Gender: masculine.
Diagnosis. Burmogonus gen. nov. (Figure 1, Figure 2 and Figure 3) can be readily recognized by the following combination of characters: body short, weakly oblong; head deflexed, mouthparts hypognathous; supra-antennal carinae incomplete across the head (i.e., not forming sharp frontal carina) and directed to labrum; pronotum rather flat, only weakly convex medially; lateral part of pronotum anteriorly bent ventrad; prosternal process rather robust, short, with blunt subapical tooth; mesoventrite rather short, its posterior part between mesocoxae broad and rectangular, with posterior margin straight; metacoxal plates more or less strongly broadened at proximal part; tarsi simple; claws simple, without basal setae. For the full description, see the respective part under B. cretaceus sp. nov. below.
Composition and distribution. Burmogonus gen. nov. is a monotypic genus and is known exclusively from Cretaceous Burmese amber.
- Burmogonus cretaceus sp. nov.
Type material. Holotype, adult specimen, sex unknown, NM-T3507, BUR0020 (NMPC). A complete beetle is included in a transparent, yellow amber piece with dimensions of 16 × 12 × 3.5 mm, with numerous minute syninclusions (debris).
Type stratum and age. Mid-Cretaceous (Upper Albian to Lower Cenomanian) [24,25,26].
Type locality. Hukawng Valley, Tanai Township, Myitkyina District, Kachin State, northern Myanmar.
Etymology. The specific epithet “cretaceus” refers to the Cretaceous period.
Diagnosis. As for the genus (vide supra).
Description. Adult. Body (Figure 1A,B) oval-elongate, rather flattened, about 6.70 mm long and 2.30 mm wide; surface rather smooth (possibly due to bad preservation without visible punctures and pubescence).
Head (Figure 2A,B and Figure 3A) partly retracted to prothorax, deflexed, including eyes 0.55 times as wide as pronotum width. Eyes strongly protuberant. Labrum more than twice as wide as long, dorsally slightly convex. Antenna (Figure 2E) 2.10 mm long, serrate from antennomere 4, relatively short, not reaching posterior angles of pronotum; scape robust, apparently wider then antennomeres 2 and 3, and slightly longer than combined lengths of antennomeres 2 and 3; pedicel and antennomere 3 short, slightly longer than wide, subequal in length; antennomere 4 about 1.65 times as long as antennomere 3; antennomeres 4–10 slightly elongate, 1.6–1.7 times as long as wide, with short serrations; terminal antennomere simple, irregularly elongate, about 2.7 times as long as wide, about 1.3 times as long as antennomere 10, widest medially, apically gradually narrowed and rounded. Mandible robust, relatively short. Maxillary palpus short; palpomere 3 slightly longer than wide; terminal palpomere longer than palpomere 3, securiform.
Pronotum (Figure 2C) weakly convex, 2.05 mm long, 2.30 mm wide, widest at posterior third, 1.10 times as wide as long, and 1.10 times as wide as elytral width at humeri. Anterior margin bisinuate, medially gradually rounded; anterior angles not developed; lateral sides very slightly rounded, gradually slightly widened toward base; posterior margin with distinct sharp notch near base of each posterior angle, medially slightly produced backwards and shallowly emarginate to fit anterior portion of scutellar shield; posterior angles relatively short but distinct, about as long as one quarter of pronotal length when measured along midline, gradually narrowed toward apex, sharp, oriented posteriorly. Lateral carina distinct, complete, not visible from above, from about midlength running ventrad of lateral edge of pronotum (Figure 3B). Disk rather smooth; sublateral carina distinct, coming from subapical inner part of posterior angle, running medially across whole length of posterior angle and slightly surpassing it. Hypomeron rather smooth, with posterior margin on each side subapically abruptly produced backward to meet apices of pronotal posterior angles. Pronotosternal sutures (Figure 3B) moderately strongly sinuate, simple, anteriorly shortly open. Prosternum (Figure 1B, Figure 2D, and Figure 3B) elongate, about twice as long as wide including prosternal process, about 1.2 times as long as wide without prosternal process; prosternal lobe very short, anteriorly widely rounded; prosternal process robust, rather short, about 0.6 times as long as prosternum in front of procoxal cavities, abruptly narrowed right after coxae, then subparallel-sided and subapically narrowed towards apex, with short blunt subapical tooth, apically narrowly rounded; ventral surface with shallow longitudinal median furrow.
Scutellar shield (Figure 2C) about 1.35 times as long as wide, widely rounded anteriorly, with sides subparallel, narrowly rounded posteriorly. Elytra (Figure 1A) 4.50 mm long, together about 2.15 as long as wide and 2.20 times as long as pronotum; weakly convex, widest at humeri, then gradually slightly tapering towards apex, conjointly narrowly rounded; each elytron laterobasally weakly emarginate to fit to cavity formed by posterior margins of dorsal part of pronotum and hypomeron; elytral epipleuron very slightly narrowed toward metacoxae, then abruptly narrowed. Mesocoxa open to both mesanepisternum and mesepimeron. Mesoventrite with anterior margin slightly emarginate on each side, medially not visible, sides almost straight, procoxal rests very shallow, mesoventral process reduced, posterior margin truncate, almost straight, mesoventral cavity deep, with walls more defined posteriorly. Mesometaventral junction straight. Metaventrite about 1.5 times as wide as long, moderately convex. Metacoxal plate (Figure 2H) well developed, rather broadened at proximal part, posteriorly short but distinctly emarginate, from its half gradually obliquely narrowed towards body edge. Hind wing not visible. Leg (Figure 2F,G) slender, femur robust, elongate; tibia about as long as femur, sparsely covered with thick short sharp setae, and with pair of distinct apical spurs; tarsus slender, elongate, slightly shorter than tibia. All tarsomeres simple, gradually slightly widened toward apex, without ventral lobes or lamellae, only with rows of long distinct setae apically; tarsomere I longest, about as long as tarsomeres II and III combined; length ratio of mesotarsomeres I–V 3.5:2.0:1.3:1.0:2.2; claws simple, slender, long, moderately curved.
4. Discussion
The accurate identification and systematic placement of fossil specimens is crucial for further conclusions about the origin and evolution of particular (not only) insect groups. Burmogonus cretaceus gen. et sp. nov. described here clearly belongs to Elateridae based on its external morphology, including the exposed labrum and the pedicel apically attached to scape [9,22,27]. Its placement in the subfamily Elaterinae is supported by the typical habitus, hypognathous head, and pretarsal claws without setae [22,23]. However, the recent most comprehensive phylogenomic study of Elateridae [23] confirmed findings of earlier molecular-based studies (e.g., [28]) that the classification of Elaterinae is far from natural, and found the tribes Agriotini, Ampedini, Dicrepidiini, Elaterini, Megapenthini, and Physorhinini all nonmonophyletic. This situation makes the correct tribal placement of elaterine fossils even more problematic.
Here, Burmogonus gen. nov. is tentatively placed in the tribe Elaterini based on the supra-antennal carinae being incomplete across head (i.e., no sharp frontal carina present; Figure 3A) and directed to the labrum, metacoxal plates only weakly narrowed outwards, and simple tarsomeres without any ventral lobes or lamellae [29,30,31]. The general habitus of Burmogonus gen. nov. might resemble that of Ampedini; however, representatives of that tribe have a complete frontal carina, lateral carina of pronotum anteriorly not bent ventrad, and metacoxal plates usually more abruptly narrowed outwards [29,30,32]. Megapenthini have a complete frontal carina, pronotal disk convex, sloping posteriorly, and the prosternal process is usually more or less thin, without a subapical tooth [29,30,33]. Agriotini, which often share with Burmogonus gen. nov. the pronotal lateral carina anteriorly bent ventrad, differ in the supra-antennal carinae usually being directed laterally from the labrum; the pronotal disk usually being rather convex, sloping posteriorly; and prosternal sutures usually being more deeply furrowed anteriorly [29,30,32]. In Physorhinini, the frontal carina is complete, the metacoxal plates are much more strongly broadened at the proximal part, and tarsomere III bears a ventral lobe [29,31,32]. Dicrepidiini have a complete frontal carina, and at least some tarsomeres bear ventral lobes [30,32,34]. Pomachillini have a complete frontal carina, and pronotum that is elongate and often widened anteriorly [29,31,32]. Synaptini have pectinate pretarsal claws and usually also have distinct, elongate basal furrows on pronotum [29,30,32]. Melanotini have a complete frontal carina and pectinate pretarsal claws [29,30,32]. Other tribes such as Aplastini, Cebrionini, Odontonychini, and Eudicronychini are much more distinctive. It should be noted that the short and posteriorly broadly rectangular mesoventrite of Burmogonus gen. nov. is mostly unusual for Elateridae, and occurs in several unrelated groups, e.g., in some Agrypninae, in most Hypnoidinae, Negastriinae, and Cardiophorinae, and rarely in Dendrometrinae.
Regarding the mid-Cretaceous Burmese amber, only five click beetle species were described before this study. Cretopityobius pankowskiorum Otto, 2019 and Cretopachyderes burmitinus Kundrata, Prosvirov, Long and Packova, 2021 are both very distinctive and belong to Pityobiinae and Agrypninae, respectively [3,17]. The recently described Dariuszelater tarnawskii Zhao, Shih and Ren, 2022 [5] was classified in Elaterinae but was not assigned to any tribe. This species differs from B. cretaceus gen. et sp. nov. in many aspects, including, e.g., the distinctly hairy body surface, relatively longer antennomeres, complete frontal carina (medially obsolete in B. cretaceus sp. nov.), pronotum widest at posterior angles (widest at the posterior third in B. cretaceus sp. nov.) and with much longer and more pronounced hind angles, double pronotosternal sutures (simple sutures in B. cretaceus sp. nov.; Figure 3B), and metacoxal plates strongly narrowed outwards (metacoxal plates only weakly narrowed outwards in B. cretaceus sp. nov.). The remaining two species were described by Cockerell more than a century ago [16], and their systematic placement is uncertain. Elater burmitinus Cockerell, 1917 was placed tentatively in the genus Elater Linnaeus, 1758 because Cockerell himself was not able to assign it to any other genus [16]. It differs clearly from B. cretaceus sp. nov. in its darker and larger body (black, about 11 mm versus brown, 6.7 mm in B. cretaceus sp. nov.) and the shape of the pronotum, especially its posterior angles, which are obliquely truncate and lack sublateral carina (versus narrow sharp posterior angles, each with a distinct sublateral carina in B. cretaceus sp. nov.). The second Cockerell’s species, “Acmaeodera“ burmitina Cockerell, 1917, was originally placed in Buprestidae but is currently tentatively placed in Elateridae incertae sedis based on the work by Bellamy [35] (for more information, see Kundrata et al. [2]). It is much larger than B. cretaceus sp. nov. (19 mm), has a prothorax wider than it is long (versus being clearly longer than wide in B. cretaceus sp. nov.), a differently shaped pronotum, relatively longer elytra compared to the length of the pronotum, and laterally serrated elytra at the apical third [16].
The discovery of a new Cretaceous elaterine species is not surprising, as this subfamily represents one of the earliest branches in Elateridae [23] and it is currently the most diverse click beetle subfamily in regard to genera and species [22]. Therefore, we expect many more species of Elaterinae to be discovered in the mid-Cretaceous amber of Myanmar in near future.
Author Contributions
Conceptualization, R.K.; methodology, K.T., G.P. and R.K.; validation, R.K.; formal analysis, K.T., A.S.P. and R.K.; investigation, K.T., G.P., A.S.P. and R.K.; resources, R.K.; data curation, K.T., G.P. and R.K.; writing—original draft preparation, R.K. and K.T.; writing—review and editing, A.S.P. and G.P.; visualization, K.T., G.P. and R.K.; supervision, R.K.; project administration, R.K. Results of this study are partly used in the unpublished Master’s thesis of K.T. supervised by R.K. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the internal grant of the Faculty of Science, UP Olomouc (IGA_PrF_2022_024; to R.K., G.P. and K.T.) and the Russian Science Foundation research project No. 21-74-10024 (to A.S.P.). R.K. received support from the SYNTHESYS+ project, which is financed by European Community Research Infrastructure Action under the H2020 Integrating Activities Programme (project GB-TAF-8656 for stay in NHMUK, London).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank J. Kvaček and J. Dašková (both NMPC) for their help during our research. R.K. would like to thank M. Geiser, M. Barclay, K. Matsumoto, D. Telnov, J. Wilkinson, and L. Stevens for their hospitality, kindness, and help during his stay in NHMUK, London.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Kundrata, R.; Packova, G.; Hoffmannova, J. Fossil genera in Elateridae (Insecta, Coleoptera): A Triassic origin and Jurassic diversification. Insects 2020, 11, 394. [Google Scholar] [CrossRef] [PubMed]
- Kundrata, R.; Packova, G.; Prosvirov, A.S.; Hoffmannova, J. The Fossil Record of Elateridae (Coleoptera: Elateroidea): Described Species, Current Problems and Future Prospects. Insects 2021, 12, 286. [Google Scholar] [CrossRef] [PubMed]
- Kundrata, R.; Prosvirov, A.S.; Long, R.; Packova, G. Cretopachyderes gen. nov., a new remarkable click beetle (Coleoptera: Elateridae: Agrypninae) from the mid-Cretaceous Burmese amber. Foss. Rec. 2021, 24, 347–355. [Google Scholar] [CrossRef]
- Szabó, M.; Kundrata, R.; Hoffmannova, J.; Németh, T.; Bodor, E.; Szenti, I.; Prosvirov, A.S.; Kukovecz, Á.; Ősi, A. The first mainland European Mesozoic click-beetle (Coleoptera: Elateridae) revealed by x-ray micro-computed tomography scanning of an Upper Cretaceous amber from Hungary. Sci. Rep. 2022, 12, 24. [Google Scholar] [CrossRef]
- Zhao, Y.; Shih, C.; Ren, D. A new genus and species of click beetle (Coleoptera, Elateridae) from mid-Cretaceous amber of northern Myanmar. Cret. Res. 2023, 142, 105387. [Google Scholar] [CrossRef]
- Dolin, V.G. K sistematike mezozoiskikh zhukov-schchelkunov (Coleoptera, Elateridae). Paleontol. Zhurnal 1975, 9, 51–62. [Google Scholar]
- Dolin, V.G. Fossil click beetles (Coleoptera, Elateridae) of the subfamilies Negastriinae and Cardophorinae from Upper Jurassic of Karatau. Vestn. Zool. 1976, 1976, 68–75. [Google Scholar]
- Dolin, V.G. Click beetles (Coleoptera, Elateridae) from the Upper Jurassic of Karatau. In Fossil Insects of the Mesozoic; Dolin, V.G., Panfilov, D.V., Ponomarenko, A.G., Pritykina, D.N., Eds.; Naukova Dumka: Kiev, Ukraine, 1980; pp. 17–81. [Google Scholar]
- Muona, J.; Chang, H.; Ren, D. The clicking Elateroidea from Chinese Mesozoic deposits (Insecta, Coleoptera). Insects 2020, 11, 875. [Google Scholar] [CrossRef]
- Li, H.; Chang, H.L.; Muona, J.; Zhao, Y.; Ren, D. Subfamily Anischiinae (Coleoptera: Eucnemidae) in early Cretaceous of Northeast China. Insects 2021, 12, 105. [Google Scholar] [CrossRef]
- Dolin, V.G.; Nel, A. Trois nouveaux Elateridae fossiles du Mésozoïque supérieur de Chine (Coleoptera). Bull. De La Société Entomol. De Fr. 2002, 107, 341–346. [Google Scholar] [CrossRef]
- Alekseev, A.V. New jewel beetles (Coleoptera: Buprestidae) and similar beetles from the Cretaceous and Early Paleogene of Asia. Paleontol. J. 2008, 42, 53–59. [Google Scholar]
- Chang, H.L.; Zhang, F.; Ren, D. A new genus and two new species of fossil Elaterids from the Yixian Formation of western Liaoning, China (Coleoptera: Elateridae). Zootaxa 2008, 1785, 54–62. [Google Scholar] [CrossRef]
- Alekseev, A.V. New click beetles (Coleoptera: Elateridae) from the Cretaceous of Russia and Kazakhstan. Paleontol. J. 2011, 45, 423–431. [Google Scholar]
- Sohn, J.C.; Nam, G.S.; Choi, S.W.; Ren, D. New fossils of Elateridae (Insecta, Coleoptera) from Early Cretaceous Jinju Formation (South Korea) with their implications to evolutionary diversity of extinct Protagrypninae. PLoS ONE 2019, 14, e0225502. [Google Scholar] [CrossRef]
- Cockerell, T.D.A. Insects in Burmese amber. Ann. Entomol. Soc. Am. 1917, 10, 323–329. [Google Scholar] [CrossRef]
- Otto, R.L. Descriptions of two new elateroid beetles (Coleoptera: Eucnemidae, Elateridae) from Burmese amber. Insecta Mundi 2019, 702, 1–6. [Google Scholar]
- Grimaldi, D.A.; Engel, M.S.; Nascimbene, P.C. Fossiliferous Cretaceous amber from Myanmar (Burma): Its rediscovery, biotic diversity, and paleontological significance. Amer. Mus. Novitates 2002, 3361, 1–71. [Google Scholar] [CrossRef]
- Ross, A.J. Burmese (Myanmar) amber checklist and bibliography 2018. Palaeoentomology 2019, 2, 22–84. [Google Scholar] [CrossRef]
- Ross, A.J. Supplement to the Burmese (Myanmar) amber chcecklist and bibliography, 2019. Palaeoentomology 2020, 3, 103–118. [Google Scholar] [CrossRef]
- Ross, A.J. Supplement to the Burmese (Myanmar) amber checklist and bibliography, 2020. Palaeoentomology 2021, 4, 57–76. [Google Scholar] [CrossRef]
- Costa, C.; Lawrence, J.F.; Rosa, S.P. Elateridae Leach, 1815. In Handbook of Zoology, Arthropoda: Insecta; Coleoptera, Beetles; Volume 2: Morphology and Systematics (Elateroidea, Bostrichiformia, Cucujiformia partim); Leschen, R.A.B., Beutel, R.G., Lawrence, J.F., Eds.; Walter de Gruyter GmbH & Co. KG: Berlin, Germany, 2010; pp. 75–103. [Google Scholar]
- Douglas, H.B.; Kundrata, R.; Brunke, A.J.; Escalona, H.E.; Chapados, J.T.; Eyres, J.; Richter, R.; Savard, K.; Ślipiński, A.; McKenna, D.; et al. Anchored Phylogenomics, Evolution and Systematics of Elateridae: Are All Bioluminescent Elateroidea Derived Click Beetles? Biology 2021, 10, 451. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.; Grimaldi, D.A.; Harlow, G.E.; Wang, J.; Wang, J.; Yang, M.; Lei, W.; Li, Q.; Li, X. Age constraint on Burmese amber based on U–Pb dating of zircons. Cret. Res. 2012, 37, 155–163. [Google Scholar] [CrossRef]
- Mao, Y.; Liang, K.; Su, Y.; Li, J.; Rao, X.; Zhang, H.; Xia, F.; Fu, Y.; Cai, C.; Huang, D. Various amberground marine animals on Burmese amber with discussions on its age. Palaeoentomology 2018, 1, 91–103. [Google Scholar] [CrossRef]
- Yu, T.; Kelly, R.; Mu, L.; Ross, A.; Kennedy, J.; Broly, P.; Xia, F.; Zhang, H.; Wang, B.; Dilcher, D. An ammonite trapped in Burmese amber. Proc. Natl. Acad. Sci. USA 2019, 116, 11345–11350. [Google Scholar] [CrossRef]
- Calder, A.A. Click Beetles: Genera of the Australian Elateridae (Coleoptera). Monographs on Invertebrate Taxonomy; CSIRO Publishing: Victoria, Australia, 1996; Volume 2, pp. 1–401. [Google Scholar]
- Kundrata, R.; Gunter, N.L.; Douglas, H.; Bocak, L. Next step toward a molecular phylogeny of click-beetles (Coleoptera: Elateridae): Redefinition of Pityobiinae, with a description of a new subfamily, Parablacinae, from the Australasian Region. Austral. Entomol. 2016, 55, 291–302. [Google Scholar] [CrossRef]
- Gurjeva, E.L. Zhuki-shchelkuny (Elateridae). Podsemeistvo Elaterinae. Triby Megapenthini, Physorhinini, Ampedini, Elaterini, Pomachiliini [Click-beetles (Elateridae). Subfamily Elaterinae. Tribes Megapenthini, Physorhinini, Ampedini, Elaterini, Pomachiliini]. In Fauna of the USSR. New series. N 118. Coleoptera. Vol. 12. Issue 4; Nauka, Leningrad branch: Leningrad, Russia, 1979; pp. 1–453. [Google Scholar]
- Kishii, T. A Taxonomic Study of the Japanese Elateridae (Coleoptera), with the Keys to the Subfamilies, Tribes and Genera; T. Kishii: Kyoto, Japan, 1987; pp. 1–262. [Google Scholar]
- Mathison, B.A. Click Beetles (Coleoptera: Elateridae) of the Southeastern United States. Occas. Pap. Fla. State Collect. Arthropods 2021, 13, 1–414. [Google Scholar]
- Stibick, J.N.L. Classification of the Elateridae (Coleoptera). Relationships and classification of the subfamilies and tribes. Pac. Insects 1979, 20, 145–186. [Google Scholar]
- Schimmel, R. Die Megapenthini-Arten Süd- und Südostasiens. Vierter Teil: Phylogenese, Zoogeographie und Katalog (Coleoptera: Elateridae); Pollichia-Buch Nr. 48, Pollichia: Bad Dürkheim, Germany, 2005; pp. 1–411. [Google Scholar]
- Casari, S.A. A phylogenetic study of the subtribe Dicrepidiina (Elateridae, Elaterinae, Ampedini). Rev. Brasil. Entomol. 2008, 52, 182–260. [Google Scholar] [CrossRef]
- Bellamy, C.L. Buprestidae (Coleoptera) from amber deposits: A brief review and family switch. Coleopt. Bull. 1995, 49, 175–177. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).