Abstract
Plant diversity in relatively harsh environments, such as metal-polluted areas tends to be relatively low. Invasive plants may invade harsh environments more easily than native plants. However, studies often find fewer invasive species in stressful edaphic habitats (such as serpentine soils). Those examples may represent relatively extreme conditions. Moderately stressful habitats may be more invaded given the advantages of invasive plants. We surveyed the plant diversity in four site pairs across three seasons. Sites consist of abandoned mines and reference sites. The mine sites have calcareous soils with relatively high iron, basic pH, and lower nutrients than reference sites. Results: There were 153 plant species among the four site pairs. Around 80 and 66% of species in calcareous and reference sites were introduced species respectively. Diversity varied across seasons but tended to be lower in the mine sites. One of the mines was significantly more invaded. Across sites, the number of invasive species and their abundances was not different from that of native species. Invasive plants are as capable of invading moderately stressful calcareous sites as native species, with some sites tending to be even more invaded.
1. Introduction
For over a century, biological invasion is an important part of global environmental change, with increasing research interest worldwide [1]. Many alien species are widely distributed in non-native areas, affecting ecosystems, livelihoods, and having significant economic consequences [2]. Invasive plants are usually widely distributed and have a strong ability to adapt to different environments. These invasive plants can displace native plant species and alter the structure and function of native plant communities [3], with the effect of reducing local or regional biodiversity [4]. Invasive plant species tend to have advantages over native species in disturbed or resource-rich conditions. Even under low resource availability, invasive plants may have higher resource utilization efficiency than native plants [5,6,7,8]. Anthropogenic activities such as mining change the structure and function of plant communities, while the species diversity in mining areas could be high, disturbance from mining activities may promote the invasion of alien plants. Studies found that many invasive plants have the ability to settle in sites polluted by heavy metals [9,10,11]. Some alien invasive plants have a high tolerance to pollutants, such as heavy metals [12,13,14]. Therefore, it is very necessary to study these questions.
Past studies have found that invasive plants can invade metal-contaminated areas [15]. Toxicity from metal pollution can seriously interfere with the growth and photosynthesis of plants [16,17]. Moreover, adaptive changes to detoxify metals and other stressors present in metal-polluted sites (such as low soil moisture or low nutrients) can reduce the performance of plants inhabiting such sites. Although the toxic effects of heavy metals can hinder the growth and fecundity of most plants, tolerant plants can proliferate in metal-polluted habitats [18,19]. Hyperaccumulator and metallophyte species are commonly found in metal-contaminated environments such as mine sites. Hyperaccumulators are species that can accumulate high concentrations of metals in their tissues without detriment to their growth [20]. Metallophytes are plant species that are able to grow and form sustained populations in polluted soils, with obligatory metallophytes being found only in metal-contaminated sites, and facultative metallophytes found in both non-contaminated and contaminated sites [21]. In their introduced ranges, invasive plants found in mine sites can be assumed to be facultative metallophytes (as they are highly abundant at the regional level and not restricted to mine sites). Studies have confirmed that many invasive plants have a strong tolerance to metals, with some able to hyperaccumulate metals in tissues [22,23,24,25]. Moreover, many invasive plants have inherent advantages in growth, fecundity, and other physiological characteristics (such as high phenotypic plasticity and allelopathy). Under competition, invasive plants usually perform better than or even inhibit native plant species [26,27]. Although competition may not be as important as tolerance to stress in stressful sites, few habitats are truly non-competitive.
Mine sites represent relatively new, highly disturbed habitats appearing relatively suddenly in time. Several invasion hypotheses suggest that the richness of invasive plants’ metal-polluted sites should be higher compared to normal sites. The ideal weed hypothesis (IWH) posits that successfully established introduced plants have relatively high growth rates, germination rates, and competitive ability [28], and are likely to colonize new areas through long-distance dispersal [29]. Similarly, the empty niche hypothesis (ENH) predicts that ecosystems with low species diversity and more empty niches are more vulnerable to invasion [30,31]. The invasion meltdown hypothesis (IMH) holds that the positive interaction between invasive species will lead to positive group-level feedback, which will intensify the impact of invaders and promote additional invasion by other alien species [32]. Similarly, invasive plants can competitively exclude native plants (e.g., in calcareous grasslands [33]).
Some studies have found that plant communities in abiotically stressful or ‘harsh’ environments are less often invaded by alien plant species than those in milder environments. A review by Zefferman [34] found that invasive plants do not often invade harsh environments such as those with high soil salinity (e.g., salt marshes) or sodic and/or alkaline soils, which have been observed to be less invaded [28]. The proportion of invasive species may be higher in less stressful sites such as mine sites under restoration (e.g., Bauman et al. [35]) and calcareous sites because stressors are less extreme. For example, some studies have shown that there is a unimodal relationship between the fine-scale species richness and the main vegetation gradient in the dry calcareous grassland in the Baltic Sea [36]. Some studies, such as Bauman et al. [35], show that there is no difference in plant diversity due to the increased coverage of invasive and naturalized species in mine areas under reclamation, and plant diversity has decreased in over five years. However, the alkalinity of the unweathered soil in the reclaimed mining areas is significantly higher (>6.5 pH), which may be conducive to the colonization of non-native herbaceous plants that can adapt to this condition [37]. A survey by Aguiar et al. [38] found that alien species are present in 91% of the survey sites, and are more common in sites at two river types (siliceous and calcareous rivers) than in the reference sites, and calcareous sites have higher invasibility, in the non-reference sites of the two river types, alien species have been more common, and the calcareous background is more sensitive to invasion. Globally, calcareous soil accounts for about 30% of the land area, and its characteristics mainly include high calcium carbonate content, low organic matter content, and low plant nutrient elements absorbed and utilized by plants [39]. Calcareous soil mainly exists in areas with higher altitudes, and the pH value is mainly around 7.8. Compared with most soils, calcareous soil is more vulnerable to heavy metal pollution [40]. Restoration work such as the addition of lime, fertilizer, or mulch used to improve the soil in barren sites may ameliorate stressors and promote colonization by adjacent plant species [41]. It is not yet clear if plant communities in certain harsh habitats such as sites with lower metal contamination and calcareous sites would be less invaded. If such sites have lower native plant diversity then invasive plants should be able to invade more easily [42]. Sites vary widely in harshness. Previous studies finding a lower presence of invaders in harsh sites may represent the most extreme conditions, such as heavily polluted sites. Communities with low species diversity and functional diversity in moderately stressful sites should be more seriously invaded than those with high species diversity and functional diversity.
In this study, we tested the plant diversity in an area with multiple mine sites, and test the following hypothesis: (1) the proportion (by the number of individuals and number of species) of invasive plants in the mine sites will be higher than that of native plants compared to adjacent reference sites, (2) the plant diversity in mine sites will be lower than reference sites; and (3) invasive species are abundant in both mine sites and non-mine areas, but native species will be restricted to either site.
2. Materials and Methods
2.1. Study Area
Four mine sites were located on hills around Zhenjiang in Jiangsu Province, China (Figure 1). These are Xianglushan (119°13′41″ E, 32°07′42.9″ N), Goutoushan (119°20′12″ E, 32°03′48″ N), Majishan (119°38′13″ E, 32°06′39″ N, Dantu District), and Longwangshan (119°36′55″ E, 32°08′12″ N). Henceforth, these ‘Locations’ will be referred to as XLS, GTS, MJS, and LWS, respectively. ‘Sites’ refer to the site type (mine site or reference site) within each ‘Location’. The mine site vegetation consisted of low herbs and grasses, with few trees and shrubs. Adjacent woodland and open sites with sparse or no tree cover were chosen for the reference sites paired with the mine sites (1–1.5 km from the respective mine sites). The soil chemical properties of all sites are presented in Table 1. Mine site soils are calcareous and have relatively high levels of iron and calcium, with a basic pH (7.6–8.2) and lower C and N content. The MJS mine site soil is also relatively high in zinc (>500 mg/kg). Mine soils are very rocky with sparser vegetation cover, particularly during spring (Figure 2). Reference sites had neutral to slightly acidic pH and lower levels of iron.
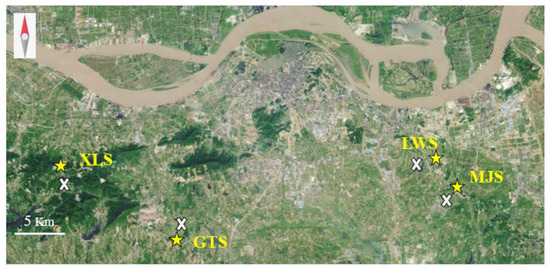
Figure 1.
Map of survey site pairs in the areas around Zhenjiang City on the Yangtze River, Jiangsu Province. Yellow stars indicate mine sites and white crosses indicate reference sites.

Table 1.
Soil properties from the sampling locations for plant diversity located around Zhenjiang (all values are total content in mg/kg, except for total carbon and nitrogen in g/kg). The soil was sampled at 10–30 cm depth. Metal concentration values are means ± SD (n = 3). Different lowercase letters represent significant differences at the location × site level (p < 0.05).

Figure 2.
Photographs detailing the local site conditions: (a) MJS mine site, (b) GTS reference site, (c) MJS mine site vegetation in spring, (d) MJS mine site vegetation in autumn, (e) GTS mine site, and (f) LWS mine site.
In the early 1960s to 70s, large-scale quarrying occurred in the areas surrounding Zhenjiang. Mining activities ceased around 2001, after which efforts were made to restore some of the sites. A portion of the mine sites at MJS and XLS was planted with the shrubs Lespedeza bicolor and Swida walteri in restoration work, so we avoided sampling those areas of the mines. The soil type in the area is highly ferric yellow-brown soil, and occasionally brownish-red soft plastic loam. The rock stratum is mostly covered with 2–3 m of residual soil, and the lithology mainly includes quartz sandstone, basalt, limestone, dolomite, etc. The climate is highly seasonal and is within the subtropical monsoon climate zone, with an annual average temperature of 16 °C. The dominant wind direction is southwest in summer and northwest in winter. The annual average precipitation is 989 mm, and the annual average relative humidity is 76%.
2.2. Plant Diversity Survey and Soil Sample Collection
We surveyed the diversity at three site pairs (mine site, and adjacent reference sites across 3 seasons in April and October (i.e., MJS, GTS, and XLS in spring and autumn 2021, and spring 2022). During the last sampling season (spring 2022), one of the site pairs (GTS) was inaccessible due to covid restrictions therefore a substitute site pair was used instead (LWS). Three 5 × 5 m quadrats were randomly placed at each sub-site, and the total number of plant species and number of individuals for each species in each quadrat were recorded. Individuals were counted non-destructively, but for some sprawling or dense clump-forming species (such as Medicago sp. or some Poaceae), plants were trimmed for better visibility. Soil samples were taken using a soil auger from a depth of 10–30 cm in three replicates, depending on the penetrability of the substrate. For each replicate, soil from three cores taken from within a 2 × 2 m area was pooled. The soil depth sampled depended on the penetrability of the substrate. A total of 2–3 kg of soil was taken at each site. The soil samples were sieved and air-dried for a week, and then sieved through a 40-mesh sieve to facilitate the measurement of the physical and chemical properties of the soil. After the soil was air-dried, samples were sent to Nanjing Convinced-Test Technology Co., Ltd. (Nanjing, China), which conducted the soil analysis. The soil’s total nitrogen content was determined by the Kjeldahl method after digestion with sulfuric acid and an accelerator. Soil pH was measured using the glass electrode method. Soil organic carbon was determined by the potassium dichromate oxidation external heating method. Hydrofluoric acid perchloric acid digestion was used to determine the content of calcium and magnesium by ICP-OES. Cadmium, chromium, lead, zinc, copper, and iron concentrations in soil were determined by four acid digestion inductively coupled plasma spectrometry.
2.3. Statistical Analysis
The proportions of invasive and native species for the number of species and the number of individuals (i.e., abundance) were tested using Chi-square tests. To quantify the species diversity at the sites, we calculated the Shannon-Wiener diversity index (H) and Simpson diversity index (D) of each sampling site. The Simpson evenness index (E) was used to quantify the distribution of invasive and native plants at each sampling site. Species similarity was quantified using the Sørensen similarity index (SI). These are calculated using the following:
where Ni is the number of individuals of the ith species, Pi = Ni/N, is the proportion of the individual number of the ith species, S is the number of plant species in the sample, and N is the total number of individuals. The higher the H and D values, the higher the plant diversity at the site. E represents the species’ evenness in the investigation area. The higher the E-value, the more uniform the plant distribution. The Sorensen Index was used to measure the similarity between two samples of data. EC is the total number of elements in common between the sets, E1 is the number of elements in site 1 and E2 is the number of elements in site 2. Hutcheson’s t-test was used to analyze differences in the Shannon-Weiner index (plant diversity) within mine sites versus reference sites [43]. Linear regression was used to test the relationship between abundance (number of individuals) in the mine sites versus abundance in reference sites for invasive and native species. Shapiro-Wilk and Levene’s tests for checking the normality and homogeneity of variance, the results of the W test show that all p-values were less than 0.05, so the logarithmic transformation was conducted on the data. All analyses were conducted using SPSS statistical software (ver. 26, IBM, Armonk, NY, USA). A p-value < 0.05 is considered statistically significant.
3. Results
There was a total of 153 species belonging to 127 genera and 52 families of plants were recorded (Supplementary materials Table S1). The distribution in the presence of species across sites is summarized in Figure 3.
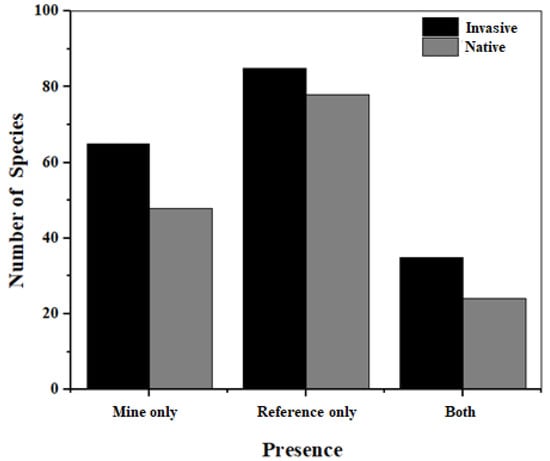
Figure 3.
The total number of invasive and native plant species present across site type combinations.
3.1. Abundance and Species Richness of Invasive Plants across Site Pairs
The average abundance of most species across sites at all locations (i.e., XLS, MJS, LWS, and GTS) was not high (Figure 4). Fewer species were present in both mine and reference sites than those present at either of the site types (Figure 3). By focusing on site pairs, the mine sites can be considered to be a filter of the existing species pool in the local area (Figure 5). Interestingly, there was overall little difference between the proportion of invasive plants and the number of native plants at each site, paired by the number of individuals (Table 2) or by the number of species (Table 3). From the average abundance distribution of plant species in mine sites and reference sites in spring 2022 (Figure S1), it can be seen that more scattered points deviate from the coordinate axis. As in the spring of 2021, although most plants seem to prefer one site type over the other (Table 4), there are still more plant species that are present in both site types than in autumn. The difference among mine sites and reference sites in the proportions of invasive plants and native plants among site pairs at each location was not as obvious in spring 2021 and 2022 as in autumn 2021. Basically, there is little difference in the number of plant individuals among site types, but, on the whole, there are more invasive plants (particularly at the MJS site pair). Spring had a significantly higher proportion of invasive species than in autumn in the number of individuals only, but did not differ significantly in the number of species (Table 5).
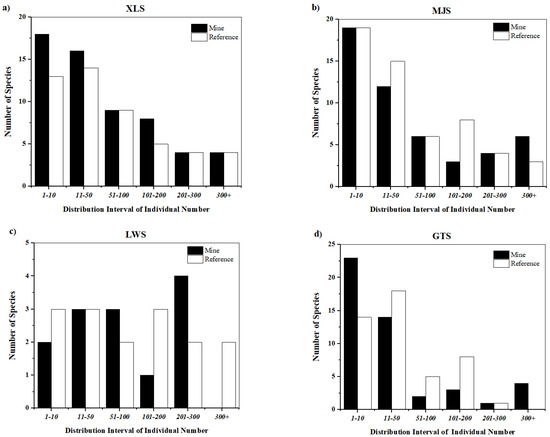
Figure 4.
Histogram showing the distribution of plant species across varying levels of abundance at each location: (a) XLS, (b) MJS, (c) LWS, and (d) GTS. The sampling at LWS was conducted only during spring 2022 when the GTS site pair was inaccessible.
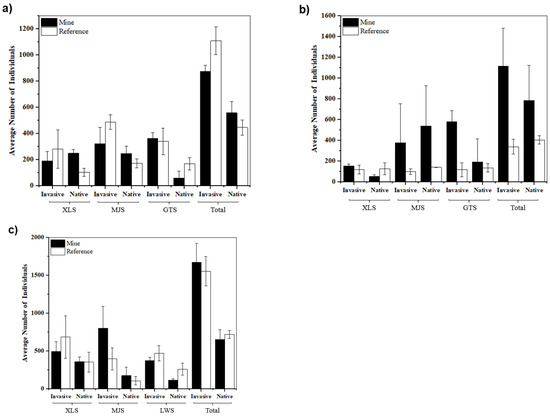
Figure 5.
Comparison of average individual numbers of invasive plants and native plants in each sampling area for: (a) spring 2021 (April), (b) autumn 2021 (October), and (c) spring (April) 2022.
Table 2.
Chi-square test results comparing the number of individuals of invasive versus native species: (a) across all sites, (b) among site pairs (mine sites and adjacent reference sites). Bold p-values indicate statistically significant differences.
Table 3.
Chi-square test results comparing the number of species of invasive versus native species: (a) across all sites, (b) among site pairs (mine sites and adjacent reference sites). Bold p-values indicate statistically significant differences.
Table 4.
Species similarity results among site pairs at each location (mine and adjacent reference sites).
Table 5.
Chi-square test results for the comparison in the proportion of invasive species to native species during spring vs. autumn across all sites. Bold p-values indicate statistically significant differences.
There was a significant positive relationship between abundance in the mine site and abundance in reference sites for both native and invasive plants, where species that are relatively abundant are also abundant across sites (Figure 6). The total plant number and the plant species richness among the mine sites and reference sites were similar (Figure 3; Table 4). Overall, calcareous mine sites did not differ significantly in the proportion of individuals that are invasive compared to reference sites (Table 2a, p > 0.05). However, when the site pairs are considered together, we can find that the difference between the individual numbers of invasive plants and native plants in each sampling area is quite significant, and invasive plants have a higher abundance (Table 2b). The results from the chi-square tests showed that: in general, the abundance of invasive plants is slightly higher than that of native plants, but compared with calcareous areas and reference areas there was no obvious difference between the number of invasive plants and native plant species (Figure 3 and Figure 4; Table 4). Looking at each sampling site separately, we found that, generally, the number of invasive plant species in all mine sites was higher than that of native plants, but in reference sites of GTS and XLS, the number of native plant species is slightly higher than that of invasive plants. Among the four sampling sites, only the number distribution of invasive plants and native plant species in GTS is different, while the number of invasive plants and native plant species in other sampling sites is not significantly different (Table 3).
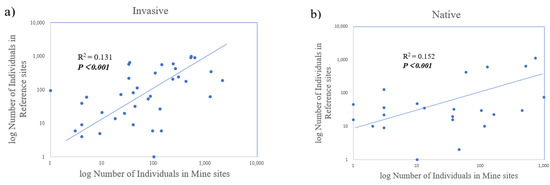
Figure 6.
Linear regressions of plant abundance in mine sites versus reference sites for the site pairs at all locations: (a) log number of individuals for invasive plants (n = 37) and (b) log number of individuals for native plants (n = 23). Each dot point represents an individual species. Values are of the total within each site type, and species absent from either site type were excluded.
3.2. Diversity in Mine Sites Versus Reference Sites
The plant species highly abundant in the reference sites include herbs of Asteraceae, Leguminosae, Rubiaceae, and Geraniaceae, such as Solidago canadensis, Galium spurium, Vicia sativa, Geranium wilfordii, etc. Among them, S. canadensis accounts for nearly 10% of the total number of individuals. Only a few shrub and tree species were present at both sites (less than 1%), such as Rubus hirsutus, Paederia cruddasiana, Ginkgo biloba, and Celtis sinensis. Herbs are abundant in reference sites, accounting for 79% of the plant species (mainly Poaceae and Asteraceae). Interestingly, mine sites had relatively low evenness compared to reference sites at GTS and MJS, but only during autumn. There are 62 species of plants present in both mine sites and reference sites, including 38 invasive plants and 24 native species. Plants species were mainly herbs accounting for > 80% (especially Asteraceae and Poaceae which account for 19% and 31%, respectively). In the mine sites, this is similar where Poaceae accounted for 17% and Asteraceae accounted for 27%, including S. canadensis, Setaria viridis, Erigeron annuus, and Elymus dahuricus. There were only a few shrub species, such as Rubus hirsutus, Duchesnea indica, and Rosa multiflora. Tree species were common, including 15 species in 12 families, such as Ginkgo biloba, Populus tomentosa, Pterocarya stenoptera, Celtis sinensis, and Broussonetia papyrifera, all of which are drought-resistant and barren tree species [44]. Patterns in plant diversity varied across seasons. Mine sites tended to have lower diversity than reference sites (i.e., MJS during spring 2021 and 2022, LS during spring 2021, and GTS during autumn 2021), except for the MJS mine site during autumn, which had higher diversity than the reference site (Table 6). The other indices of diversity are presented in Table S2.
Table 6.
Results from Hutcheson’s t-tests on the differences in plant diversity among mine versus reference sites (Shannon-Weiner index, H).
3.3. Distribution and Abundance of Native and Invasive Species among Sites
Most species are found only at one of the site types, especially at the MJS site pair (mine and reference sites) in autumn (Figure S1). S. canadensis and Setaria viridis were the most abundant species across the site pairs at XLS and GTS (Figure S1). In autumn (Figure 5), there were large differences in abundance among site pairs in each sampling location (e.g., GTS mine site S. canadensis individuals and GTS reference site Eleusine indica individuals; Figure S1). In mine sites, abundance was significantly higher than that of reference sites in spring than in autumn, and most of them are introduced or invasive plants (particularly at the MJS site pair). However, the number of species was similar between autumn and spring (χ2 = 0.04, p = 0.83), but invasive species had higher abundance (χ2 = 473, p < 0.001). The plant species diversity in spring 2022 was similar to that of spring in the preceding year (Table 3a). Mine sites had similar diversity to reference sites (Table S2).
4. Discussion
Our results suggest that invasions are commonplace in moderately stressful habitats such as calcareous mine sites, and certain stressful sites may be significantly more invaded than others. In the study region, alien and invasive plants account for the majority (>50%) of all species. Invasive plant abundance in the mine sites was generally higher than that of native plants. Invasive plants still dominate in most cases in reference sites.
Our hypotheses were: (1) the proportion of invasive plants in the mine sites will be higher than that of native plants. On the whole, invasive plants are very prevalent across sites but did not differ significantly between mine sites and reference areas with the exception of the GTS mine site, which had a significantly higher proportion of invasive plant species relative to natives compared to the proportions in the adjacent reference site. The results for the proportion of invasive species by the total number of individuals were not consistent, with some mine sites having a significantly greater proportion of invasive plants compared to that of the adjacent sites (GTS and LWS) and others had an increased proportion of native species in the mine site than in the adjacent sites (XLS and MJS). Overall, the first prediction has only limited support; (2) the plant diversity in mine sites will be lower than in reference sites. Conversely, we found that species diversity, evenness, and richness were similar across sites and seasons. Evenness was relatively low at the GTS and MJS mine sites only during the autumn sampling. The patterns in plant diversity within site pairs varied across seasons. There were no significant differences among sites in some seasons, but mine sites tend to have lower diversity than adjacent sites (except for MJS during autumn 2021). The species similarity tended to be lower in the mine sites. Thus, there was overall slight support for the second prediction; (3) Invasive species are abundant in both mine sites and reference areas, but native species will be restricted to either site. We found that native and invasive species both vary widely in which sites they inhabit and in similar proportions, and that the common species present at both sites tend to be also common in both sites. Therefore, we found no support for the third prediction.
The high presence of invasive species at the calcareous mine sites was not too unexpected. The overall result was similar to findings from other types of calcareous environments. For instance, in a calcareous grassland ecological restoration project [45], the reconstruction of local calcareous grassland plant communities was affected by the high coverage of invasive plants in the calcareous areas. They similarly observed relatively low evenness in the old restoration site compared to reference sites. This may be due to the dominance of invasive grass in these areas. Bart et al. (2011) studied the plant communities in calcareous marshes and found that those environments were mainly composed of woody invasive and weedy native species. Compared with flat land, the area in the marshland that has been plowed or disturbed there were more pioneer native and invasive herbaceous species. Weeds, herbs, and rhizomes are particularly abundant and frequent in these areas. Despite the existence of these invasive weeds and invasive species, the local pioneer species of marsh continue to live in the wing marsh, and the central community is the remains of the local herbaceous calcareous marsh vegetation [46]. There were few invasive species in the center of the calcareous marsh, but more invasive species on the disturbed edges. Thus, the relatively high number of invasive species at some mine sites (e.g., GTS) might be associated with disturbance. The presence of relatively high soil iron or heavy metals (such as high zinc in the MJS mine) did not seem to impact the overall level of plant diversity.
Our investigation into the plant diversity at ferrous calcareous sites found lower similarity among sites within site pairs, particularly between mine sites and adjacent reference sites. This suggests that the stressors present at the mines apply selective pressures on plants. However, these stressors do not limit the number of plant species able to grow in such sites and may promote a different suite of species that are less common outside of the mines. In autumn, the evenness of species in mine sites was much lower than in reference sites, meaning that the plants in reference sites are more evenly distributed in space, but not in the mine sites, where the plants are more clustered. This distribution may be related to the rocky soils found at mine sites. We found that the iron content in the soil of the region is high (i.e., from 45 to 50 g/kg in reference soils, and from 57 to 85 g/kg in mine soils), which may have had an impact on the species diversity of the region. Presently, there is no literature to investigate the impact of iron on plant diversity, Species diversity was much lower in autumn compared to spring (such as at the MJS), but invasive plants are still predominant across the site. This suggests that invasive plants have stronger environmental tolerance than native plants, and have a competitive advantage over plants with poor tolerance in polluted soil [47]. Although invasive plants have a strong tolerance to the environment, there is a sharp contrast between plant species diversity in spring and autumn. It may also be that as resources are reduced during autumn (e.g., lower solar irradiation and reduced rainfall), the growth of many plants becomes slow or stagnant, and invasive plants can make better use of the resources than local plants, which increases the survival probability [6]. However, in general, the richness or number of invasive plants in different seasons and reference sites was not different from that of local plants. In different stages of vegetation restoration, environmental factors have a phased impact on plant diversity. At different stages and locations, they will be more dominant than alien invasive plants, or in most cases, they are equal to invasive plants [48]. On the one hand, local resident plants (both invasive and native plants) may have adapted to this stressful environment for a long time through phenotypic plasticity or ecological differentiation [49,50,51,52,53,54] has made them completely adapt to environmental pressure like invasive plants. On the other hand, the stressors at the mine sites likely produced a filtering effect, where a portion of the surrounding resident species was excluded from the mines, leaving other species able to tolerate the stressors to become (and not necessarily involving adaptation). Whether plants at the mine sites are adapted to stressors may be addressed in future studies.
The overall diversity is similar across sites, which is not unexpected for moderately stressful habitats such as calcareous sites. Interestingly, the number of species that were only found at mine sites was the greatest (Figure 4). Even in GTS reference sites, there is no species advantage compared with local plants (Table 4). At the same time, due to ore mining, the original vegetation community of the mine has been seriously altered. The survey area belongs to the early stage of natural succession with many Poaceae (especially Imperata cylindrica, Digitaria sanguinalis, Setaria viridis) and Asteraceae (especially Erigeron annuus, E. canadensis and S. canadensis), which are commonly found in wastelands and most of them being invasive species. Poaceae and Asteraceae species are dominant plants in the early stages of natural succession in the restoration of vegetation community [55]. These species have a wide range of distribution, strong adaptability, fast growth, large numbers, and form local small communities. Many Asteraceae and Poaceae plants can adapt to environments with high soil Mn, Cr, Cd, and other heavy metals and other stress factors such as exposure to winds and sun, low nutrients, and drought, so they can be considered pioneer plants for ecological restoration [56,57].
This study aims to explore whether invasive plant species can readily colonize moderately stressful habitats, such as calcareous mine sites, and provide information on species suitable for ecological restoration. The mechanisms of plant invasion are complex and diverse, and an understanding of whether those mechanisms are the same when invasive plants colonize stressful habitats is still developing. The results suggest invasive species are not inhibited from colonizing moderately stressful habitats and may have advantages in some stressful sites such as the GTS mine site which was drastically more invaded than the other mine sites (especially by Setaria viridis, Figure S1). The reasons for this are still unknown and warrant further investigation. Environmental protection and ecological restoration of vegetation at abandoned mine sites are important for ecological health. However, the impacts of invasive plant species in polluted sites should also be considered, particularly as they can easily invade many of such sites which can impede ecological restoration. Future work can address the major problems in global ecological protection.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d14110986/s1, Figure S1: Average abundance distribution of plant species in the mining area and reference sites: (a) April, spring 2021; (b) October, autumn 2021; (c) April, spring 2022. Each dot point represents an individual plant species; Table S1: The presence and abundance of each individual species at each site. Table S2: Plant diversity, abundance and evenness between mine sites and reference sites across all seasons (H is Shannon’s Diversity Index, D is Simpson’s Diversity Index, R is Margalef Index, and E is Evenness index). The sampling at LWS was conducted only during spring 2022 when the GTS site pair was inaccessible. Indices were calculated from three quadrats.
Author Contributions
Conceptualization, J.S.H.W. and S.R.; methodology, J.-H.L., S.R. and J.S.H.W.; software, D.-L.D.; validation, Z.-C.D., J.-F.S. and X.-Q.S.; formal analysis, J.-H.L. and J.S.H.W.; investigation, J.-H.L. and H.L.; resources, J.S.H.W.; data curation, J.-H.L.; writing-original draft, J.-H.L.; writing-review and editing, J.-H.L., J.S.H.W., S.R., A.A.-N. and X.-Q.S.; visualization, J.-H.L.; supervision, J.S.H.W. and D.-L.D.; project administration, J.S.H.W.; funding acquisition, J.S.H.W. Additional details: J.-H.L. conducted the surveys, analyzed the data, collected soil, and wrote the first draft of the manuscript. J.S.H.W. designed the study, provided materials and logistics, collected soil, and edited the manuscript. S.R. designed the study and contributed to data collection. H.L., Z.-C.D. and J.-F.S. contributed to data collation. J.-H.L. and X.-Q.S. identified the plant species. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Jiangsu Natural Science Foundation (BK20200905). JSHW was supported by the Jiangsu University Research Foundation Fund (20JDG056) and the Jiangsu province’s ‘Double Innovation’ talent program. SR was supported by the Jiangsu University Research Foundation Fund (20JDG055), and DZC by the National Natural Science Foundation of China (32271587), carbon peak and carbon neutrality technology innovation foundation of Jiangsu Province (BK20220030).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data is available in the Supplementary Materials.
Acknowledgments
We thank Rahil Afzal for his help with soil collection.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dawson, W.; Schrama, M.; Austin, A. Identifying the role of soil microbes in plant invasions. J. Ecol. 2016, 104, 1211–1218. [Google Scholar] [CrossRef]
- Pyšek, P.; Hulme, P.E.; Simberloff, D.; Bacher, S.; Blackburn, T.M.; Carlton, J.T.; Dawson, W.; Essl, F.; Foxcroft, L.C.; Genovesi, P.; et al. Scientists’ warning on invasive alien species. Biol. Rev. 2020, 95, 1511–1534. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Zhong, C.; Li, X.; Guo, Z.; Shi, S. Hybridization with natives augments the threats of introduced species in sonneratia mangroves. Aquat. Bot. 2019, 160, 103166. [Google Scholar] [CrossRef]
- Dai, Z.C.; Zhu, B.; Wan, J.S.H.; Rutherford, S. Editorial: Global Changes and Plant Invasions. Front. Ecol. 2022, 10, 845816. [Google Scholar] [CrossRef]
- Morris, T.L.; Esler, K.J.; Barger, N.N.; Jacobs, S.M.; Cramer, M.D. Ecophysiological traits associated with the competitive ability of invasive Australian acacias. Divers. Distrib. 2011, 17, 898–910. [Google Scholar] [CrossRef]
- Matzek, V. Superior performance and nutrient-use efficiency of invasive plants over non-invasive congeners in a resource-limited environment. Biol. Invasions 2011, 13, 3005–3014. [Google Scholar] [CrossRef]
- Firn, J.; Prober, S.M.; Buckley, Y.M. Plastic traits of an exotic grass con-tribute to its abundance but are not always favorable. PLoS ONE 2012, 7, e35870. [Google Scholar] [CrossRef] [PubMed]
- Funk, J.L. Differences in plasticity between invasive and native plants from a low resource environment. J. Ecol. 2008, 96, 1162–1174. [Google Scholar] [CrossRef]
- Lee, W.G.; Mark, A.F.; Wilson, J.B. Ecotypic differentiation in the ultramafic flora of the South Island, New Zealand. N. Z. J. Bot. 1983, 21, 141–156. [Google Scholar] [CrossRef]
- Gulezian, P.Z.; Jennifer, L.I.; Kelly, J.G. Establishment of an Invasive Plant Species (Conium maculatum) in Contaminated Roadside Soil in Cook County, Illinois. Am. Midl. Nat. 2012, 168, 375–395. [Google Scholar] [CrossRef]
- Prabakaran, K.; Li, J.; Anandkumar, A.; Leng, Z.; Zou, C.B.; Du, D. Managing environmental contamination through phytoremediation by invasive plants: A review. Ecol. Eng. 2019, 138, 28–37. [Google Scholar] [CrossRef]
- Gong, W.; Wang, Y.; Chen, C.; Xiong, Y.; Zhou, Y.; Xiao, F.; Li, B.; Wang, Y. The rapid evolution of an invasive plant due to increased selection pressures throughout its invasive history. Ecotoxicol Environ. Saf. 2022, 233, 113322. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Chao, C.; Yxa, B.; Yi, W.; Qla, B. Combination effects of heavy metal and inter-specific competition on the invasiveness of Alternanthera philoxeroides. Environ. Exp. Bot. 2021, 189, 104532. [Google Scholar]
- Cai, H.H. Effects of Heavy Metal cd on Alien Invasive Plants and Their Resistance to Disease. Master’s Thesis, Jiangsu University, Zhenjiang, China, 2016. [Google Scholar]
- Dai, Z.C.; Cai, H.H.; Qi, S.S.; Li, J.; Du, D.L. Cadmium hyperaccumulation as an inexpensive metal armor against disease in Crofton weed. Environ. Pollut. 2020, 267, 115649. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.F. Effects of Cu, Zn, Cd, Pb on the Growth of Three Leguminous Plants and Their Adsorption Properties. Master’s Thesis, Northeastern University, Shenyang, China, 2008. [Google Scholar]
- Wang, H.; Jiao, Z.H.; Wu, H.; Wang, Y.G.; Wang, X.X.; Wang, L.S. Review on evaluation methods of heavy metal pollution in soil around lead zinc mine. J. Shenyang Univ. (Nat. Sci.) 2021, 33, 300–306. [Google Scholar]
- Chowdhury, A.; Maiti, S.K. Identification of metal tolerant plant species in mangrove ecosystem by using community study and multivariate analysis: A case study from Indian Sunderban. Environ. Earth Sci. 2016, 75, 744. [Google Scholar] [CrossRef]
- Tovar-Sánchez, E.; Hernández-Plata, I.; Martínez, M.S.; Valencia-Cuevas, L.; Galante, P.M. ‘Heavy Metal Pollution as a Biodiversity Threat’ in Heavy Metals; IntechOpen: London, UK, 2018. [Google Scholar]
- Jaffré, T.; Brooks, R.R.; Lee, J.; Reeves, R.D. Sebertia acuminata: A Hyperaccumulator of Nickel from New Caledonia. Science 1976, 193, 579–580. [Google Scholar] [CrossRef]
- Li, M.J.; Yu, F.X.; Ding, J.N.; Xiong, Z.T. Water Metabolism Changes of Metallophyte-Elsholtzia haichowensis Treated with Copper and Water Stress. Jiangxi Sci. 2014, 32, 46–50. [Google Scholar]
- Chen, C.; Zhang, H.X.; Wang, A.G.; Lu, M.; Shen, Z.G.; Lian, C.L. Phenotypic plasticity accounts for most of the variation in leaf manganese concentrations in Phytolacca americana growing in manganese-contaminated environments. Plant Soil 2015, 396, 215–227. [Google Scholar] [CrossRef]
- Chen, L.; Gao, J.; Zhu, Q.; Wang, Y.; Yang, Y. Accumulation and output of heavy metals by the invasive plant Spartina alterniflora in a coastal salt marsh. Pedosphere 2018, 28, 884–894. [Google Scholar] [CrossRef]
- Liu, X.W.; Qi, C.M.; Wang, Z.C.; Ouyang, C.B.; Li, Y.; Yan, D.D.; Wang, Q.X.; Guo, M.X.; Yuan, Z.H.; He, F.L. Biochemical and ultrastructural changes induced by lead and cadmium to Crofton weed (Eupatorium adenophorum Spreng.). Int. J. Environ. Res. 2018, 12, 597–607. [Google Scholar] [CrossRef]
- Dai, Z.C.; Wan, L.Y.; Qi, S.S.; Rutherford, S.; Du, D.L. Synergy among hypotheses in the invasion process of alien plants: A road map within a timeline. Perspect. Plant Ecol. 2020, 47, 125575. [Google Scholar] [CrossRef]
- Davidson, A.M.; Jennions, M.; Nicotra, A.B. Do invasive species show higher phenotypic plasticity than native species and, if so, is it adaptive? A meta-analysis. Ecol. Lett. 2015, 14, 419–431. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.L.; Feng, Y.L.; Zhang, L.K.; Callaway, R.M.; Alfonso, V.B.; Luo, D.Q.; Liao, Z.Y.; Lei, Y.B.; Barclay, G.F.; Carlos, S.P. Integrating novel chemical weapons and evolutionarily increased competitive ability in success of a tropical invader. New Phytol. 2015, 205, 1350–1359. [Google Scholar] [CrossRef] [PubMed]
- Baker, H.G. The Evolution of weeds. Annu. Rev. Ecol. Evol. 1974, 5, 1–4. [Google Scholar] [CrossRef]
- Wang, M.N.; Dai, Z.C.; Qi, S.S.; Wang, X.Y.; Du, D.L. Main hypotheses and research progress of invasion mechanism of exotic plants. Jiangsu Agric. Sci. 2014, 42, 378–382. [Google Scholar]
- Hierro, J.L.; Callaway, M. A biogeographical approach to plant invasions: The importance of studying exotics in their introduced and native range. J. Ecol. 2005, 93, 5–15. [Google Scholar] [CrossRef]
- Harvey, K.J.; Nipperess, D.A.; Britton, D.R.; Hughes, L. Australian family ties: Does a lack of relatives help invasive plants escape natural enemies? Biol. Invasions 2012, 14, 2423–2434. [Google Scholar] [CrossRef]
- Green, P.T.; O’Dowd, D.J.; Abbott, K.L.; Jeffery, M.; Retallick, K.; Mac, N.R. Invasional meltdown: Invader-invader mutualism facilitates a secondary invasion. Ecology 2011, 92, 1758–1768. [Google Scholar] [CrossRef]
- Pitz, C.; Mahy, G.; Harzé, M.; Uyttenbroeck, R.; Monty, A. Comparison of mining spoils to determine the best substrate for rehabilitating limestone quarries by favoring native grassland species over invasive plants. Ecol. Eng. 2018, 127, 510–518. [Google Scholar] [CrossRef]
- Zefferman, E.; Stevens, J.; Charles, G.; Dunbar-Irwin, M.; Emam, T.; Fick, S.; Morales, L.V.; Wolf, K.M.; Young, D.J.; Young, T.P. Plant communities in harsh sites are less invaded: A summary of observations and proposed explanations. AoB Plants 2015, 7, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Bauman, J.M.; Cochran, C.; Chapman, J.; Gilland, K. Plant community development following restoration treatments on a legacy reclaimed mine site. Ecol. Eng. 2015, 83, 521–528. [Google Scholar] [CrossRef]
- Triin, R.; Aveliina, H.; Meelis, P.; Karin, B.; Pille, G.; Ejvind, R.; Krista, T.; Sergey, Z.; Honor, C.P. Determinants of fine-scale plant diversity in dry calcareous grasslands within the Baltic Sea region. Agric. Ecosyst. Environ. 2014, 182, 9–68. [Google Scholar]
- Franklin, J.A.; Zipper, C.E.; Burger, J.A.; Skousen, J.G.; Jacobs, D.F. Influence of herbaceous ground cover on forest restoration of eastern US coal surface mines. New For. 2012, 43, 905–924. [Google Scholar] [CrossRef]
- Aguiar, F.C.; Ferreira, M.T.; Albuquerque, A.; Moreira, I. Alien and endemic flora at reference and non-reference sites in Mediterranean-type streams in Portugal. Aquat. Conserv. 2010, 17, 335–347. [Google Scholar] [CrossRef]
- Hu, Y.; Huang, Y.; Su, J.; Gao, Z.; Nan, Z. Temporal changes of metal bioavailability and extracellular enzyme activities in relation to afforestation of highly contaminated calcareous soil. Sci. Total Environ. 2018, 622, 1056–1066. [Google Scholar] [CrossRef]
- Moore, T.J.; Loeppert, R.H. Significance of potassium chloride pH of calcareous soils. Soil Sci. Soc. Am. J. 1987, 51, 908–912. [Google Scholar] [CrossRef]
- Johnson, C.D.; Skousen, J.G. Minesoil Properties of 15 Abandoned Mine Land Sites in West Virginia. J. Environ. Qual. 1995, 24, 635–643. [Google Scholar] [CrossRef]
- Maron, J.; Marler, M. Native plant diversity resists invasion at both low and high resource levels. Ecology 2007, 88, 2651–2661. [Google Scholar] [CrossRef]
- Hutcheson, K. A test for comparing diversities based on Shannon formula. J. Theor. Biol. 1970, 29, 151–154. [Google Scholar] [CrossRef]
- Hu, S.; Xiao, X.; Jia, H.S.; Zhou, J. Comparison and analysis on dust-retention ability of major deciduous greening species in Xuzhou. Chin. Agric. Sci. Bull. 2012, 28, 95–98. [Google Scholar]
- Piqueray, J.; Bottin, G.; Delescaille, L.M.; Bisteau, E.; Colinet, G.; Mahy, G. Rapid restoration of a species-rich ecosystem assessed from soil and vegetation indicators: The case of calcareous grasslands restored from forest stands. Ecol. Indic. 2011, 11, 724–733. [Google Scholar] [CrossRef]
- Bart, D.; Simon, M.; Carpenter, Q.; Graham, S. Historical Land Use and Plant-Community Variability in a Wisconsin Calcareous Fen. Rhodora 2011, 113, 160–186. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, R.; Tang, J.; Chen, X. Competitive interaction between the invasive Solidago canadensis and native Kummerowia striata in lead contaminated soil. Bot. Stud. 2008, 49, 385–391. [Google Scholar]
- Liu, X.P.; Zhang, W.J.; Yang, F.; Zhou, X.; Liu, Z.J.; Qu, F.; Lian, S.Q.; Wang, C.L.; Tang, X.G. Changes in vegetation–environment relationships over long-term natural restoration process in Middle Taihang Mountain of North China. Ecol. Eng. 2012, 49, 193–200. [Google Scholar] [CrossRef]
- Schweitzer, J.A.; Larson, K. Greater morphological plasticity of exotic honeysuckle species may make them better invaders than native species. Torrey Bot. Soc. 1999, 126, 15–23. [Google Scholar] [CrossRef]
- Geng, Y.P.; Pan, X.Y.; Xu, C.Y.; Zhang, W.J.; Li, B.; Chen, J.K.; Lu, B.R.; Song, Z.P. Phenotypic plasticity rather than locally adapted ecotypes allows the invasive alligator weed to colonize a wide range of habitats. Biol. Invasions 2007, 9, 245–256. [Google Scholar] [CrossRef]
- Molina-Montenegro, M.A.; Palma-Rojas, C.; Alcayaga-Olivares, Y.; Oses, R.; Corcuera, L.J.; Cavieres, L.A.; Gianoli, E. Ecophysiological traits suggest local adaptation rather than plasticity in the invasive Taraxacum officinale (dandelion) from native and introduced habitat range. Plant Ecol. Divers. 2011, 4, 36–42. [Google Scholar]
- Molina-Montenegro, M.A.; Palma-Rojas, C.; Alcayaga-Olivares, Y.; Oses, R.; Corcuera, L.J.; Cavieres, L.A.; Gianoli, E. Ecophysiological plasticity and local differentiation help explain the invasion success of Taraxacum officinale (dandelion) in South America. Ecography 2012, 36, 718–730. [Google Scholar] [CrossRef]
- Molina-Montenegro, M.A.; Naya, D.E. Latitudinal patterns in phenotypic plasticity and fitness-related traits: Assessing the climatic variability hypothesis (CVH) with an invasive plant species. PLoS ONE 2012, 7, e47620. [Google Scholar] [CrossRef]
- Molina-Montenegro, M.A.; Peñuelas, J.; Munné-Bosch, S.; Sardans, J. Higher plasticity in ecophysiological traits enhances the performance and invasion success of Taraxacum officinale (dandelion) in alpine environments. Biol. Invasions 2012, 14, 21–33. [Google Scholar] [CrossRef]
- Han, F.; Li, C.R.; Sun, M.G.; Fan, Y.X.; Zhao, F.; Lu, S.S.; Fu, Y. Plant community structure at an early ecological restoration stage on an abandoned quarry in Sibao Mount. J. Cent. South Univ. 2008, 28, 35–39. [Google Scholar]
- Yang, S.X.; Li, M.S.; Li, Y.; Huang, H.R. Research status of ecological restoration and pollution of heavy metals by plants in Pingle manganese mine. Minging Saf. Environ. Prot. 2006, 33, 21–23. [Google Scholar]
- Luo, Y.P.; Li, M.S.; Zhang, X.H.; Liu, J.; Huang, H.T.; Cai, X.W. Characteristics of bioaccumulation of heavy metals by dominant plants in Lipu manganese mine, Guangxi. J. GuangXi Norm. Univ. 2005, 23, 89–93. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).