
Flower-Visiting Insect Assemblages on Fall-Blooming Native California Sage Scrub Shrubs

 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Sampling
2.2.1. Hand Netting
2.2.2. Photo Sampling
2.2.3. Malaise Trap Sampling
2.3. Insect Sorting and Identification
2.4. Analysis of Flower-Visiting Insect Assemblages
2.5. Pollen Analyses
2.5.1. Species Selection
2.5.2. Pollen Sample Removal
2.5.3. DNA Extraction
2.5.4. Pollen DNA Analyses
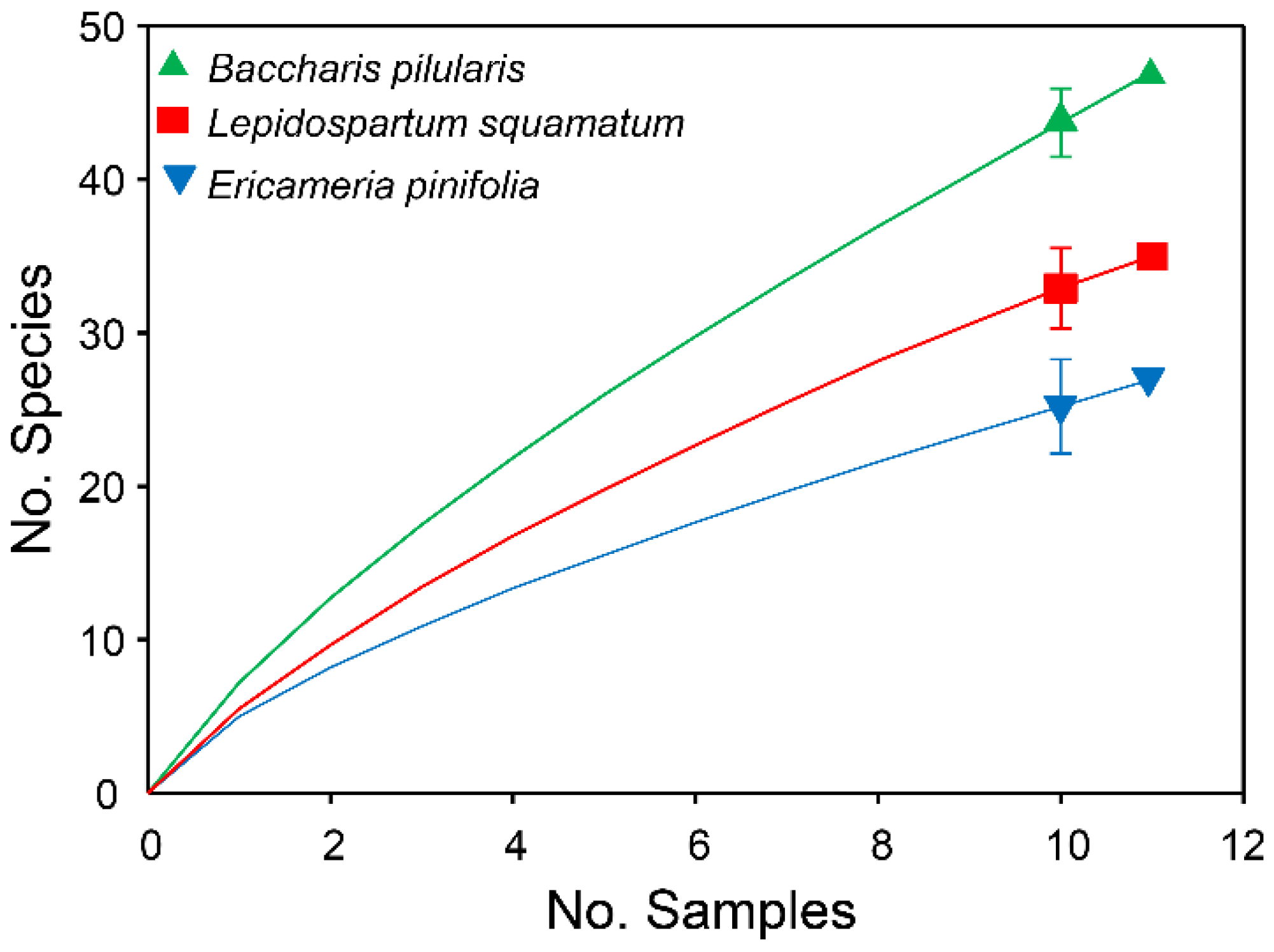
3. Results
3.1. Documentation of Flower-Visiting Insects
3.2. Comparison between Active and Passive Sampling Techniques
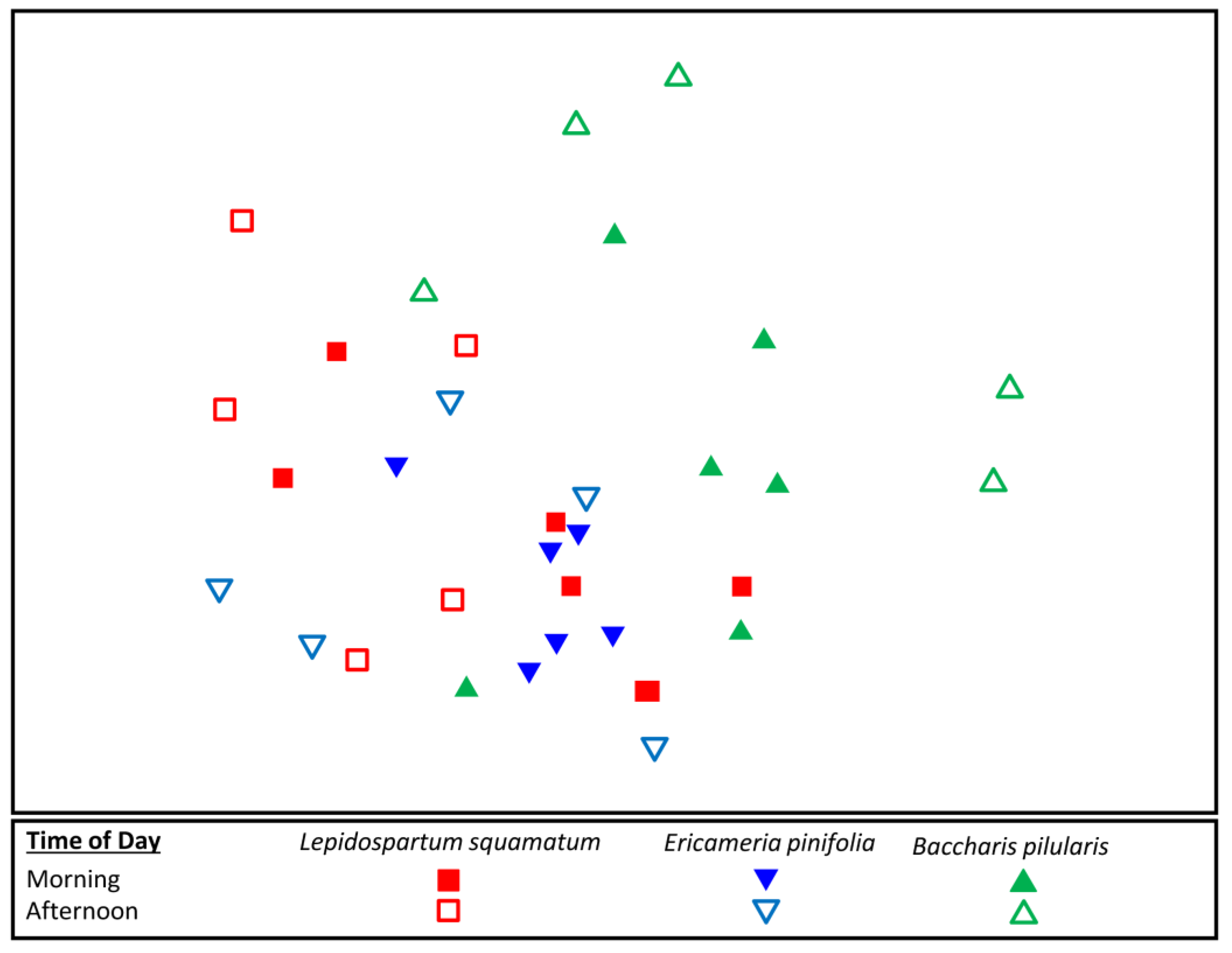
3.3. Flower-Visiting Insect Assemblage Differences on Fall Flowering Shrubs
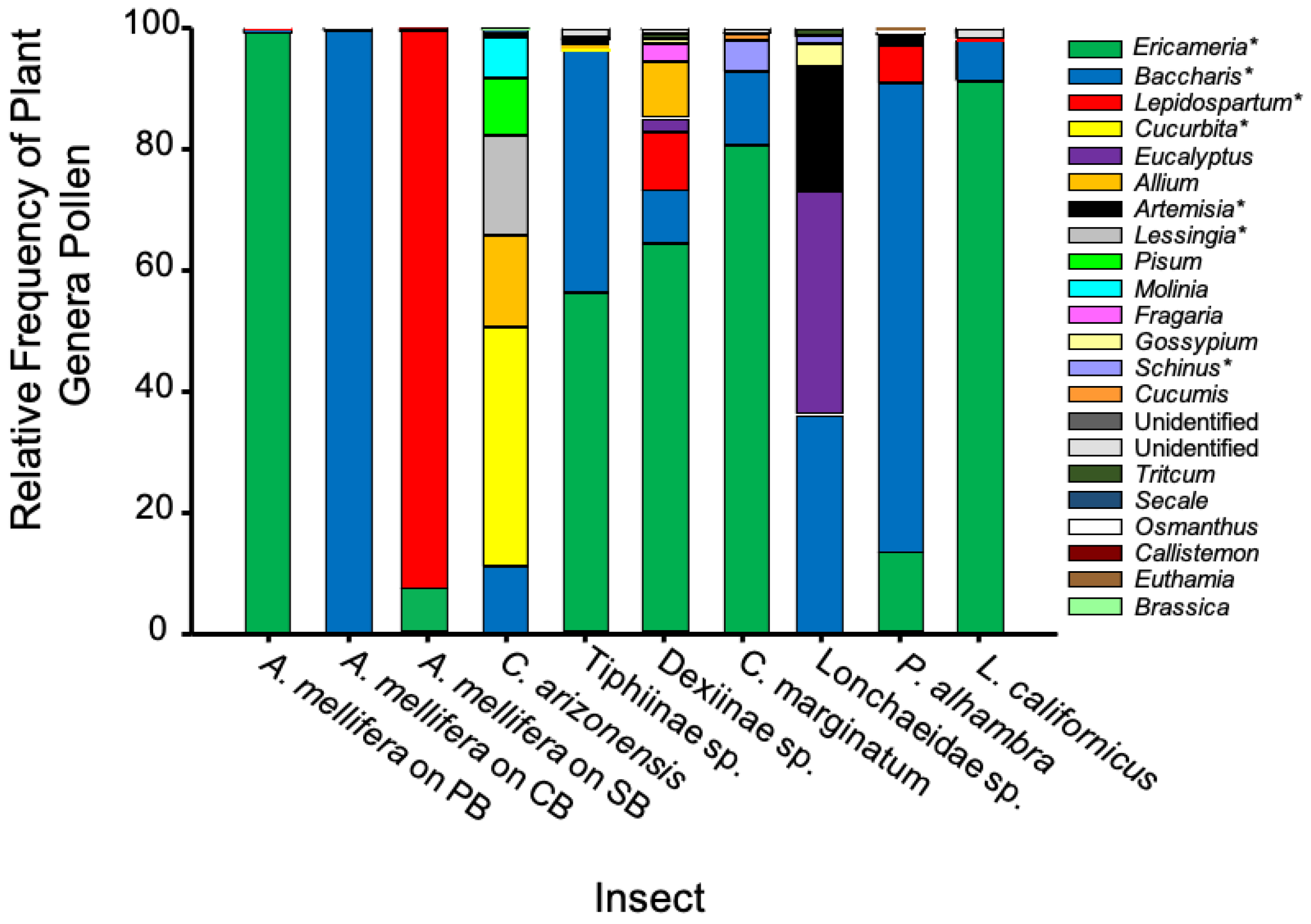
3.4. Pollen Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Michener, C.D. Biogeography of the bees. Ann. Mo. Bot. Garden 1979, 66, 277–347. [Google Scholar] [CrossRef]
- Cincotta, R.P.; Wisnewski, J.; Engelman, R. Human population in the biodiversity hotspots. Nature 2000, 404, 990–992. [Google Scholar] [CrossRef] [PubMed]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Hung, K.-L.J.; Ascher, J.S.; Gibbs, J.; Irwin, R.E.; Bolger, D.T. Effects of fragmentation on a distinctive coastal sage scrub bee fauna revealed through incidental captures by pitfall traps. J. Insect Conserv. 2015, 19, 175–179. [Google Scholar] [CrossRef]
- Moldenke, A.R. California pollinationecology and vegetation types. Phytologia 1976, 34, 305–361. [Google Scholar]
- Hung, K.-L.J.; Ascher, J.S.; Holway, D.A. Urbanization-induced habitat fragmentation erodes multiple components of temporal diversity in a Southern California native bee assemblage. PLoS ONE 2017, 12, e0184136. [Google Scholar] [CrossRef]
- Munz, P.A.; Renshaw, D.L.; Faber, P.M. Introduction to California Desert Wildflowers; University of California Press: Berkeley, CA, USA, 2020. [Google Scholar]
- Westman, W.E. Diversity relations and succession in Californian coastal sage scrub. Ecology 1981, 62, 170–184. [Google Scholar] [CrossRef]
- Bolger, D.T.; Suarez, A.V.; Crooks, K.R.; Morrison, S.A.; Case, T.J. Arthropods in urban habitat fragments in southern California: Area age, and edge effects. Ecol. Appl. 2000, 10, 1230–1248. [Google Scholar] [CrossRef]
- Rubinoff, D. Evaluating the California gnatcatcher as an umbrella species for conservation of southern California coastal sage scrub. Conserv. Biol. 2001, 15, 1374–1383. [Google Scholar] [CrossRef]
- Riordan, E.C.; Rundel, P.W. Modelling the distribution of a threatened habitat: The California sage scrub. J. Biogeogr. 2009, 36, 2176–2188. [Google Scholar] [CrossRef]
- Shreeve, T.G.; Mason, C.F. The number of butterfly species in woodlands. Oecologia 1980, 45, 414–418. [Google Scholar] [CrossRef] [PubMed]
- Bowers, M.A. Bumble bee colonization, extinction, and reproduction in subalpine meadows in northeastern Utah. Ecology 1985, 66, 914–927. [Google Scholar] [CrossRef]
- Jennersten, O. Pollination in Dianthus deltoides (Caryophyllaceae): Effects of habitat fragmentation on visitation and seed set. Conserv. Biol. 1988, 2, 359–366. [Google Scholar] [CrossRef]
- Cane, J.H. Habitat fragmentation and native bees: A premature verdict? Conserv. Ecol. 2001, 5, 1. [Google Scholar] [CrossRef]
- Gezon, Z.J.; Wyman, E.S.; Ascher, J.S.; Inouye, D.W.; Irwin, R.E. The effect of repeated, lethal sampling on wild bee abundance and diversity. Methods Ecol. Evol. 2015, 6, 1044–1054. [Google Scholar] [CrossRef]
- Sawyer, J.O.; Keeler-Wolf, T.; Evens, J.M. A Manual of California Vegetation; California Native Plant Society Press: Sacramento, CA, USA, 1995. [Google Scholar]
- Tabush, C.P. Temporal Bee Diversity and Abundance within the California Sage Scrub of the San Jose Hills and Chino Hills. Master’s Dissertation, California State Polytechnic University, Pomona, CA, USA, 2019. [Google Scholar]
- Montalvo, A.M.; Riordan, E.C.; Beyers, J. Plant Profile for Lepidospartum squamatum. In Riverside-Corona Resource Conservation District and U.S. Department of Agriculture, Forest Service; Pacific Southwest Research Station: Riverside, CA, USA, 2017. [Google Scholar]
- Francis, J.K. Wildland Shrubs of the United States and Its Territories: Thamnic Descriptions, Volume 1; U.S. Department of Agriculture, Forest Service, International Institute of Tropical Forestry: San Juan, PR, USA; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2004.
- Hoffman, L. Spatiotemporal Variation in Pollinator Taxa on the Santa Ana River Woolly Star Eriastrum densifolium ssp. sanctorum (Milliken) Mason (Polemoniaceae). Master’s Dissertation, California State University, Fullerton, CA, USA, 2010. [Google Scholar]
- Barrios-Garcia, M.N.; Rodriguez-Cabal, M.A.; Rudgers, J.A.; Crutsinger, G.M. Soil fertilization does not alter plant architectural effects on arthropod communities. J. Plant Ecol. 2016, 10, 800–807. [Google Scholar] [CrossRef][Green Version]
- Rudgers, J.A.; Whitney, K.D. Interactions between insect herbivores and a plant architectural dimorphism. J. Ecol. 2006, 94, 1249–1260. [Google Scholar] [CrossRef]
- Huey, S.; Nieh, J.C. Foraging at a safe distance: Crab spider effects on pollinators: Crab spider effects on pollinators. Ecol. Entomol. 2017, 42, 469–476. [Google Scholar] [CrossRef]
- Wania, A.; Kühn, I.; Klotz, S. Plant richness patterns in agricultural and urban landscapes in central Germany—Spatial gradients of species richness. Landsc. Urb. Plan. 2006, 75, 97–110. [Google Scholar] [CrossRef]
- Stout, J.C.; Morales, C.L. Ecological impacts of invasive alien species on bees. Apidologie 2009, 40, 388–409. [Google Scholar] [CrossRef]
- Goddard, M.A.; Dougill, A.J.; Benton, T.J. Scaling up from gardens: Biodiversity conservation in urban environments. Trends Ecol. Evol. 2010, 25, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Schlaepfer, M.A.; Sax, D.F.; Olden, J.D. The potential conservation value of non-native species: Conservation value of non-native species. Conserv. Biol. 2011, 25, 428–437. [Google Scholar] [CrossRef] [PubMed]
- Koyama, A.; Egawa, C.; Taki, H.; Yasuda, M.; Kanzaki, N.; Ide, T.; Okabe, K. Non-native plants are a seasonal pollen source for native honeybees in suburban ecosystems. Urban Ecosyst. 2018, 21, 1113–1122. [Google Scholar] [CrossRef]
- Bjerknes, A.-L.; Totland, Ø.; Hegland, S.J.; Nielsen, A. Do alien plant invasions really affect pollination success in native plant species? Biol. Conserv. 2007, 138, 1–12. [Google Scholar] [CrossRef]
- Frankie, G.; Pawelek, J.; Chase, M.H.; Jadallah, C.C.; Feng, I.; Rizzardi, M.; Thorp, R. Native and non-native plants attract diverse bees to urban gardens in California. J. Pollination Ecol. 2019, 25, 16–23. [Google Scholar] [CrossRef]
- Alarcón, R. Congruence between visitation and pollen-transport networks in a California plant-pollinator community. Oikos 2010, 119, 35–44. [Google Scholar] [CrossRef]
- Thomson, D.M.; Meyer, W.M., III; Whitcomb, I.F. Non-native plant removal and high rainfall years promote post-fire recovery of Artemisia californica in southern California sage scrub. PLoS ONE 2021, 16, e0254398. [Google Scholar] [CrossRef]
- Dartnell, S.D.; Hamlett, N.; Meyer, W.M., III. Monitoring butterfly assemblages in southern California to assess the impacts of habitat and climate modifications. J. Insect Conserv. 2022, 26, 149–162. [Google Scholar] [CrossRef]
- Caspi, T.; Hartz, L.A.; Soto Villa, A.E.; Loesberg, J.A.; Robins, C.R.; Meyer, W.M., III. Impacts of invasive annuals on soil carbon and nitrogen storage in southern California depend on identity of the invader. Ecol. Evol. 2019, 9, 4980–4993. [Google Scholar] [CrossRef]
- Xu, X.; Ren, Z.-X.; Trunschke, J.; Kuppler, J.; Zhao, Y.-H.; Knop, E.; Wang, H. Bimodal activity of diurnal flower visitation at high elevation. Ecol. Evol. 2021, 11, 13487–13500. [Google Scholar] [CrossRef]
- Irwin, R.E.; Brody, A.K.; Waser, N.M. The impact of floral larceny on individuals, populations, and communities. Oecologia 2001, 129, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, A.L.; Harder, L.D.; Johnson, S.D. Consumptive emasculation: The ecological and evolutionary consequences of pollen theft. Biol. Rev. 2009, 84, 259–276. [Google Scholar] [CrossRef] [PubMed]
- Inouye, D.W. The terminology of floral larceny. Ecology 1980, 61, 1251–1253. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. PRIMER V6: User Manual/Tutorial. Plymouth, PRIMER-E. Mar. Biol. 2006, 118, 167–176. [Google Scholar] [CrossRef]
- Bell, K.L.; Fowler, J.; Burgess, K.S.; Dobbs, E.K.; Gruenewald, D.; Lawley, B.; Morozumi, C.; Brosi, B.J. Applying pollen DNA metabarcoding to the study of plant–pollinator interactions. Appl. Plant Sci. 2017, 5, 1600124. [Google Scholar] [CrossRef]
- Sickel, W.; Ankenbrand, M.J.; Grimmer, G.; Holzschuh, A.; Härtel, S.; Lanzen, J.; Steffan-Dewenter, I.; Keller, A. Increased efficiency in identifying mixed pollen samples by meta-barcoding with a dual-indexing approach. BMC Ecol. 2015, 15, 20. [Google Scholar] [CrossRef]
- Bolyen, D.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotech. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Rivers, A.R.; Weber, K.C.; Gardner, T.G.; Liu, S.; Armstrong, S.D. ITSxpress: Software to rapidly trim internally transcribed spacer sequences with quality scores for marker gene analysis. F1000Research 2018, 7, 1418. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2′s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- Banchi, E.; Ametrano, C.G.; Greco, S.; Stanković, D.; Muggia, L.; Pallavicini, A. PLANiTS: A curated sequence reference dataset for plant ITS DNA metabarcoding. Database 2020, 2020, baz155. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, A.H.; Thomas, M.; Bhandari, R.; Meshram, H. Malaise trap and insect sampling: Mini review. Biol. Bull. 2016, 2, 35–40. [Google Scholar]
- Cane, J.H.; Minckley, R.L.; Kervin, L.J. Sampling bees (Hymenoptera: Apiformes) for pollinator community studies: Pitfalls of pan-trapping. J. Kans. Entomol. Soc. 2000, 73, 225–231. [Google Scholar]
- Trent, T.R.; Evans, E.W.; Tepedino, V.J. Pan-trapping for bees (Hymenoptera: Apiformes) in Utah’s West Desert: The importance of color diversity. Pan-Pac. Entomol. 2005, 81, 103–113. [Google Scholar]
- Roulston, T.H.; Smith, S.A.; Brewster, A.L. A comparison of pan trap and intensive net sampling techniques for documenting a bee (Hymenoptera: Apiformes) fauna. J. Kans. Entomol. Soc. 2007, 80, 179–181. [Google Scholar] [CrossRef]
- Westphal, C.; Bommarco, R.; Carré, G.; Lamborn, E.; Morison, N.; Petanidou, T.; Potts, S.G.; Roberts, S.P.M.; Szentgyôrgyi, H.; Tscheulin, T.; et al. Measuring bee diversity in different European habitats and biogeographical regions. Ecol. Monogr. 2008, 78, 653–671. [Google Scholar] [CrossRef]
- Matthews, R.W.; Matthews, R.J. Malaise trap: The townes trap catches more insects. Contrib. Am. Entomol. Inst. 1983, 20, 428–432. [Google Scholar]
- Torchio, P.F.; Trostle, G.E. Biological notes on Anthophora urbana urbana and its parasite, Xeromelecta californica (Hymenoptera: Anthophoridae), including descriptions on late embryogenesis and hatching. Annal. Entomol. Soc. Am. 1986, 79, 434–447. [Google Scholar] [CrossRef]
- Bossert, S.; Wood, T.J.; Patiny, S.; Michez, D.; Almeida, E.A.B.; Minckley, R.L.; Packer, L.; Neff, J.L.; Copeland, R.S.; Straka, J.; et al. Phylogeny, biogeography and diversification of the mining bee family Andrenidae. Syst. Entomol. 2021, 47, 283–302. [Google Scholar] [CrossRef]
- Timberlake, P.H. A Revisional Study of the Bees of the Genus Perdita F. Smith, with Special Reference to the Fauna of the Pacific Coast (Hymenoptera, Apoidea) Part III; University of California Press: Berkeley, CA, USA; Los Angeles, CA, USA, 1958. [Google Scholar]
- Riley, J.R.; Greggers, U.; Smith, A.D.; Reynolds, D.R.; Menzel, R. The flight paths of honeybees recruited by dance. Nature 2005, 425, 205–207. [Google Scholar] [CrossRef]
- Grüter, C.; Farina, W. The honeybee waggle dance: Can we follow the steps? Trends Ecol. Evol. 2009, 24, 242–247. [Google Scholar] [CrossRef]
- Nürnberger, F.; Keller, A.; Härtel, S.; Steffan-Dewenter, I. Honey bee waggle dance communication increases diversity of pollen diets in intensively managed agricultural landscapes. Mol. Ecol. 2019, 28, 3602–3611. [Google Scholar] [CrossRef] [PubMed]
- Abrol, D.P. Foraging behavior of bees as influenced by quality and quantity of rewards from flowers. J. Asia-Pac. Entomol. 2006, 9, 145–148. [Google Scholar] [CrossRef]
- Al Toufailia, H.; Couvillon, M.J.; Ratnieks, F.L.W.; Grüter, C. Honey bee waggle dance communication: Signal meaning and signal noise affect dance follower behaviour. Behav. Ecol. Sociobiol. 2013, 67, 549–556. [Google Scholar] [CrossRef]
- Saritha, R.; Vinod Chandra, S.S. Multi dimensional honey bee foraging algorithm based on optimal energy consumption. J. Inst. Eng. India Ser. B 2017, 98, 527–531. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Plant Species | Sample Start Date | Sample End Date |
|---|---|---|
| L. squamatum | 16 September | 1 October |
| E. pinifolia | 12 October | 5 November |
| B. pilularis | 20 October | 5 November |
| Species | No. Indiv. | Netting Location | Extracted DNA (ng/µL) | No. Raw Reads | No. Plant Genera | % Pollen from BFS 2 |
|---|---|---|---|---|---|---|
| Apis mellifera | 44 | L. squamatum | 2.45 | 34,630 | 7 | 99.79 |
| Apis mellifera | 46 | B. pilularis | 9.0 | 32,878 | 2 | 100 |
| Apis mellifera | 62 | E. pinifolia | 4.1 | 21,249 | 6 | 99.93 |
| Ceratina arizonensis | 19 | All 3 hosts | 1.55 | 13,933 | 9 | 67.17 |
| Copestylum marginatum | 8 | All 3 hosts | 0 1 | 12,684 | 5 | 97.90 |
| Dexiinae sp. | 18 | L. squamatum & E. pinifolia | 0.05 | 23,660 | 9 | 82.84 |
| Largus californicus | 6 | All 3 hosts | 0 | 17,021 | 5 | 98.23 |
| Lonchaeidae sp.1 | 13 | B. pilularis | 0 | 11,869 | 6 | 58.16 |
| Palpada alhambra | 27 | All 3 hosts | 0.3 | 12,684 | 11 | 99.05 |
| Tiphiinae sp.1 | 22 | B. pilularis | 1.8 | 15,634 | 7 | 97.63 |
| Species | Pine Bush | Coyote Bush | Scale Broom | Malaise | |||
|---|---|---|---|---|---|---|---|
| Net | Photo | Net | Photo | Net | Photo | ||
| Agapostemon texanus | X | X | |||||
| Agapostemon sp. | X | ||||||
| Anthophora urbana | X | X | X | ||||
| Anthophora sp. | X | ||||||
| Apis mellifera | X | X | X | X | X | ||
| Ashmeadiella sp. | X | ||||||
| Ceratina arizonensis | X | X | X | X | |||
| Ceratina (Zadontomerus) sp. | X | ||||||
| Ceratina sp. | X | ||||||
| Colletes sp.1 | X | ||||||
| Colletes sp.2 | X | ||||||
| Dialictus sp.1 | X | ||||||
| Dialictus sp.2 | X | ||||||
| Dialictus sp.3 | X | ||||||
| Hylaeus sp. | X | ||||||
| Halictus farinosus | X | ||||||
| Halictus ligatus | X | ||||||
| Perdita ericameriae | X | X | |||||
| Rophitinae sp. | X | ||||||
| Xeromelecta californica | X | ||||||
| Xylocopa sonarina | X | ||||||
| Total: 21 bee species | 8 | 4 | 8 | 0 | 6 | 6 | 0 |
| Species | Pine Bush | Coyote Bush | Scale-Broom | Malaise | |||
|---|---|---|---|---|---|---|---|
| Net | Photo | Net | Photo | Net | Photo | ||
| Atlides halesus | X | ||||||
| Brephidium exilis | X | ||||||
| Danaus gilippus | X | ||||||
| Danaus plexippus | X | ||||||
| Erynnis tristis | X | X | |||||
| Euptoieta claudia | X | ||||||
| Heliopetes ericetorum | X | ||||||
| Hemiargus ceraunus | X | ||||||
| Hylephila phyleus | X | X | X | ||||
| Junonia coenia | X | X | |||||
| Plebejus acmon | X | ||||||
| Pyrgus albescens | X | ||||||
| Strymon melinus | X | X | X | ||||
| Vanessa cardui | X | X | X | ||||
| Total: 14 butterfly species | 1 | 4 | 0 | 1 | 2 | 14 | 0 |
| Species | Pine Bush | Coyote Bush | Scale Broom | Malaise | |||
|---|---|---|---|---|---|---|---|
| Net | Photo | Net | Photo | Net | Photo | ||
| Ancistrocerus sp. | X | ||||||
| Anomalon sp. | X | ||||||
| Bembix sp. | X | ||||||
| Braconidae sp.1 | X | ||||||
| Braconidae sp.2 | X | ||||||
| Brasema sp. | X | X | |||||
| Cerceris convergens | X | ||||||
| Cerceris sp. | X | ||||||
| Chalcidoidea sp. | X | ||||||
| Eulophidae sp. | X | ||||||
| Eurytomidae sp.1 | X | X | |||||
| Eurytomidae sp.2 | X | ||||||
| Gasteruptiidae sp.1 | X | ||||||
| Gasteruptiidae sp.2 | X | ||||||
| Leptochilus sp. | X | ||||||
| Oxybelus sp. | X | ||||||
| Paratiphia sp.1 | X | ||||||
| Paratiphia sp. 2 | X | ||||||
| Philanthus gibbosus | X | ||||||
| Philanthus sp.1 | X | ||||||
| Philanthus sp. 2 | X | ||||||
| Polistes aurifer | X | ||||||
| Tiphiinae sp. 1 | X | X | |||||
| Tiphiinae sp. 2 | X | ||||||
| Tiphiinae sp. 3 | X | ||||||
| Vespula pensylvanica | X | ||||||
| Total: 26 wasp species | 4 | 0 | 8 | 0 | 8 | 9 | 0 |
| Species | Pine Bush | Coyote Bush | Scale-Broom | Malaise | |||
|---|---|---|---|---|---|---|---|
| Net | Photo | Net | Photo | Net | Photo | ||
| Bagrada hilaris | X | ||||||
| Largus californicus | X | X | X | X | X | ||
| Largus sp. | X | ||||||
| Murgantia histrionica | X | ||||||
| Neacoryphus bicrucis | X | ||||||
| Total: 5 hemipterans | 1 | 1 | 1 | 0 | 2 | 4 | 0 |
| Species | Pine Bush | Coyote Bush | Scale-Broom | Malaise | |||
|---|---|---|---|---|---|---|---|
| Net | Photo | Net | Photo | Net | Photo | ||
| Acreophthiria sp. | X | X | |||||
| Allograpta obliqua | X | ||||||
| Aphoebantus sp.1 | X | X | X | ||||
| Aphoebantus sp.2 | X | ||||||
| Bombyliidae sp.1 | X | X | |||||
| Bombyliidae sp.2 | X | ||||||
| Bombyliidae sp.3 | X | ||||||
| Calliphoridae sp.1 | X | ||||||
| Calliphoridae sp.2 | X | ||||||
| Ceratitis capitata | X | ||||||
| Coenosia sp. | X | ||||||
| Conopidae sp. | X | ||||||
| Copestylum marginatum | X | X | X | ||||
| Copestylum mexicanum | X | X | X | X | X | ||
| Copestylum satur | X | X | X | X | |||
| Copestylum violaceum | X | ||||||
| Desmometopa sp. | X | ||||||
| Dexiinae sp. | X | X | X | X | |||
| Dilophus sp. | X | ||||||
| Dioprosopa clavata | X | X | X | X | |||
| Diptera sp.1 | X | ||||||
| Diptera sp.2 | X | X | |||||
| Diptera sp.3 | X | X | |||||
| Diptera sp.4 | X | ||||||
| Diptera sp.5 | X | ||||||
| Diptera sp.6 | X | ||||||
| Diptera sp.7 | X | ||||||
| Diptera sp.8 | X | ||||||
| Diptera sp.9 | X | N/A * | |||||
| Diptera sp.10 | X | N/A * | |||||
| Eristalinus taeniops | X | X | |||||
| Eristalis hirta | X | X | |||||
| Eupeodes fumipennis | X | ||||||
| Eupeodes volucris | X | X | |||||
| Geron sp. | X | X | |||||
| Lonchaeidae sp.1 | X | ||||||
| Lonchaeidae sp.2 | X | ||||||
| Lucilia sp. | X | ||||||
| Musciodea sp. | X | X | |||||
| Oscinellinae sp. | X | ||||||
| Palpada alhambra | X | X | X | X | X | ||
| Palpada mexicana | X | X | X | ||||
| Paragus haemorrhous | X | ||||||
| Paragus sp. | X | ||||||
| Phthiriinae sp.1 | X | X | |||||
| Spilomyia interrupta | X | ||||||
| Tachinidae sp.1 | X | ||||||
| Tachinidae sp.2 | X | ||||||
| Tachinidae sp.3 | X | ||||||
| Tephritini sp. | X | ||||||
| Thaumatomyia sp. | X | ||||||
| Toxomerus marginatus | X | X | X | X | |||
| Trichopoda pennipes | X | X | |||||
| Villa lateralis | X | ||||||
| Villa sp. | X | X | |||||
| Zodion sp. | X | ||||||
| Total: 56 fly species | 13 | 8 | 30 | 5 | 16 | 15 | 7 |
| Plant Species | No. Species Collected | No.Unique Species | No. Singleton Specimens | Predicted Species Richness (SD) | % Inventory Completeness |
|---|---|---|---|---|---|
| L. squamatum | 35 | 22 | 14 | 76.1 (23.8) | 45.99 |
| E. pinifolia | 27 | 12 | 8 | 117.3 (76.7) | 23.02 |
| B. pilularis | 47 | 36 | 26 | 209 (98.2) | 22.49 |
| Pairwise Comparison | R Value | p Value |
|---|---|---|
| L. squamatum, E. pinifolia | −0.025 | 0.614 |
| L. squamatum, B. pilularis | 0.319 | 0.002 * |
| E. pinifolia, B. pilularis | 0.283 | 0.005 * |
| Pairwise Comparison Insect Species | Proportion of Samples Present | Avg. Dissimilarity ± 1 SD | Contributed % Dissimilarity | |
|---|---|---|---|---|
| B. pilularis vs. E. pinifolia | B. pilularis | E. pinifolia | ||
| Tiphiinae sp.1 | 0.55 | 0.00 | 4.66 ± 1.01 | 5.94 |
| Palpada alhambra | 0.55 | 0.64 | 4.40 ± 0.91 | 5.62 |
| Largus californicus | 0.09 | 0.36 | 3.12 ± 0.76 | 3.97 |
| Copestylum marginatum | 0.09 | 0.36 | 3.10 ± 0.73 | 3.96 |
| Lonchaeidae sp.2 | 0.36 | 0.00 | 2.78 ± 0.70 | 3.54 |
| Braconidae sp.1 | 0.36 | 0.00 | 2.68 ± 0.71 | 3.42 |
| Dexiinae sp. | 0.00 | 0.27 | 2.33 ± 0.58 | 2.97 |
| Desmometopa sp. | 0.27 | 0.00 | 2.13 ± 0.58 | 2.71 |
| Ceratina arizonensis | 0.18 | 0.09 | 2.05 ± 0.51 | 2.61 |
| Braconidae sp.2 | 0.00 | 0.27 | 2.05 ± 0.60 | 2.61 |
| B. pilularis vs. L. squamatum | B. pilularis | L. squamatum | ||
| Tiphiinae sp.1 | 0.55 | 0.00 | 4.39 ± 1.03 | 5.34 |
| Palpada alhambra | 0.55 | 0.36 | 4.24 ± 0.96 | 5.17 |
| Dexiinae sp. | 0.00 | 0.45 | 4.02 ± 0.87 | 4.90 |
| Copestylum marginatum | 0.09 | 0.27 | 2.74 ± 0.65 | 3.34 |
| Lonchaeidae sp.2 | 0.36 | 0.00 | 2.63 ± 0.71 | 3.20 |
| Braconidae sp.1 | 0.36 | 0.00 | 2.55 ± 0.72 | 3.10 |
| Ceratina arizonensis | 0.18 | 0.18 | 2.22 ± 0.62 | 2.71 |
| Desmometopa sp. | 0.27 | 0.00 | 2.01 ± 0.58 | 2.45 |
| Lonchaeidae sp.1 | 0.27 | 0.00 | 1.89 ± 0.59 | 2.30 |
| Apis mellifera | 0.82 | 1.00 | 1.69 ± 0.46 | 2.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dartnell, S.; Cavalcanti, A.R.O.; Misaki Bluebell, A.; Hamlett, N.V.; Crane, E.J., III; Meyer, W.M., III. Flower-Visiting Insect Assemblages on Fall-Blooming Native California Sage Scrub Shrubs. Diversity 2022, 14, 958. https://doi.org/10.3390/d14110958
Dartnell S, Cavalcanti ARO, Misaki Bluebell A, Hamlett NV, Crane EJ III, Meyer WM III. Flower-Visiting Insect Assemblages on Fall-Blooming Native California Sage Scrub Shrubs. Diversity. 2022; 14(11):958. https://doi.org/10.3390/d14110958
Chicago/Turabian StyleDartnell, Sofia, Andre R. O. Cavalcanti, Ayame Misaki Bluebell, Nancy V. Hamlett, Edward J. Crane, III, and Wallace M. Meyer, III. 2022. "Flower-Visiting Insect Assemblages on Fall-Blooming Native California Sage Scrub Shrubs" Diversity 14, no. 11: 958. https://doi.org/10.3390/d14110958
APA StyleDartnell, S., Cavalcanti, A. R. O., Misaki Bluebell, A., Hamlett, N. V., Crane, E. J., III, & Meyer, W. M., III. (2022). Flower-Visiting Insect Assemblages on Fall-Blooming Native California Sage Scrub Shrubs. Diversity, 14(11), 958. https://doi.org/10.3390/d14110958