

Tracing the Food Web of Changing Arctic Ocean: Trophic Status of Highly Abundant Fish, Gasterosteus aculeatus (L.), in the White Sea Recovered Using Stomach Content and Stable Isotope Analyses

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
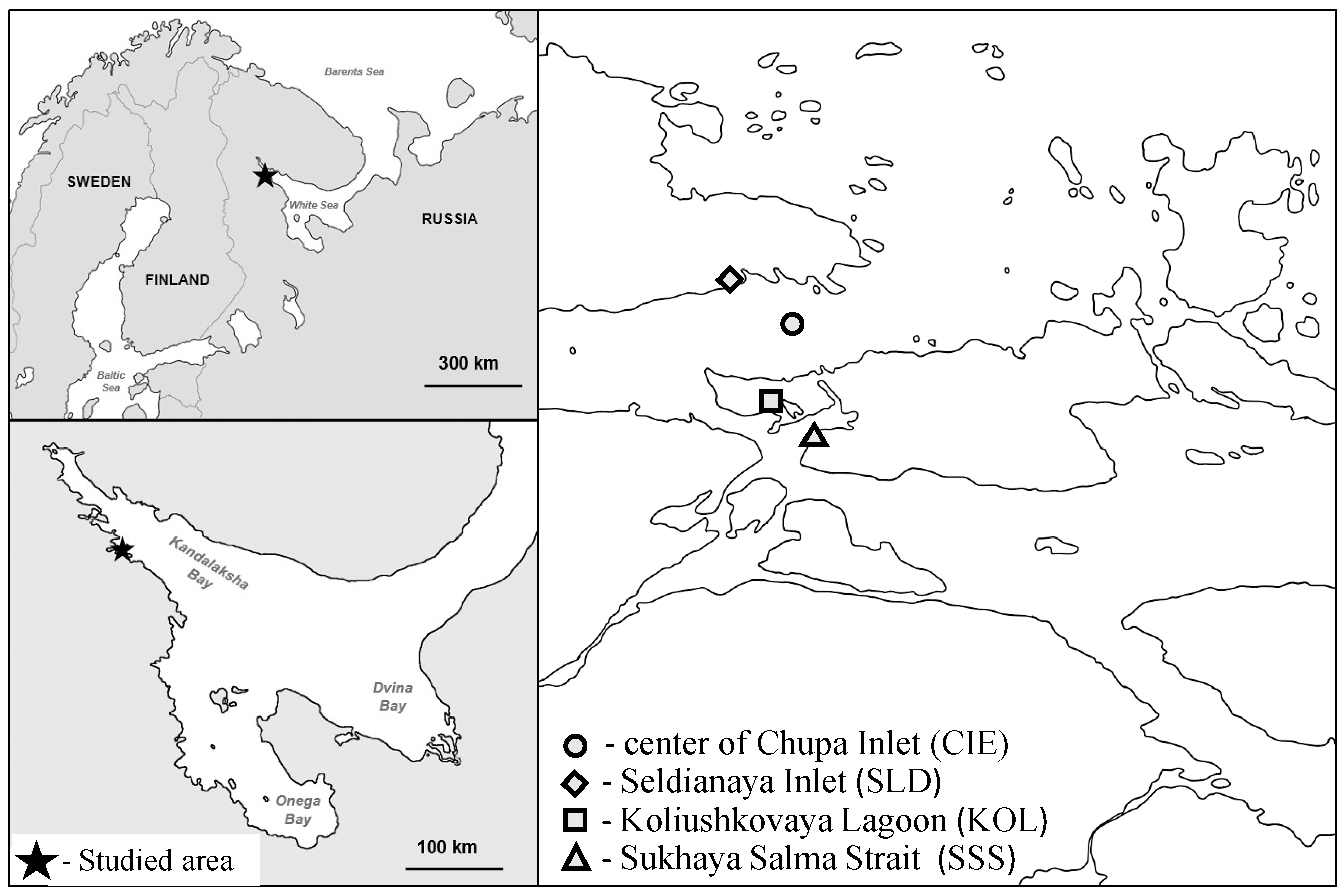
2.1. Study Area and Field Sampling
2.2. Laboratory Analyses
2.3. Data Analysis
3. Results
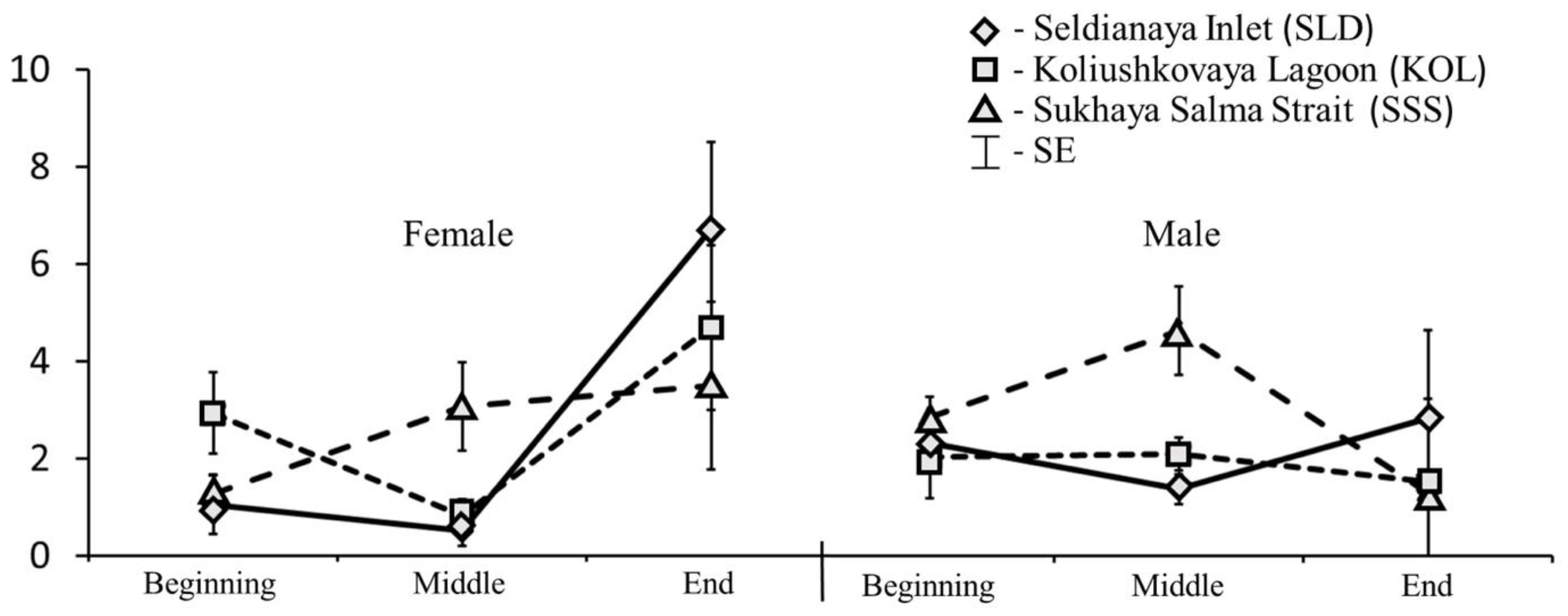
3.1. Feeding Intensity
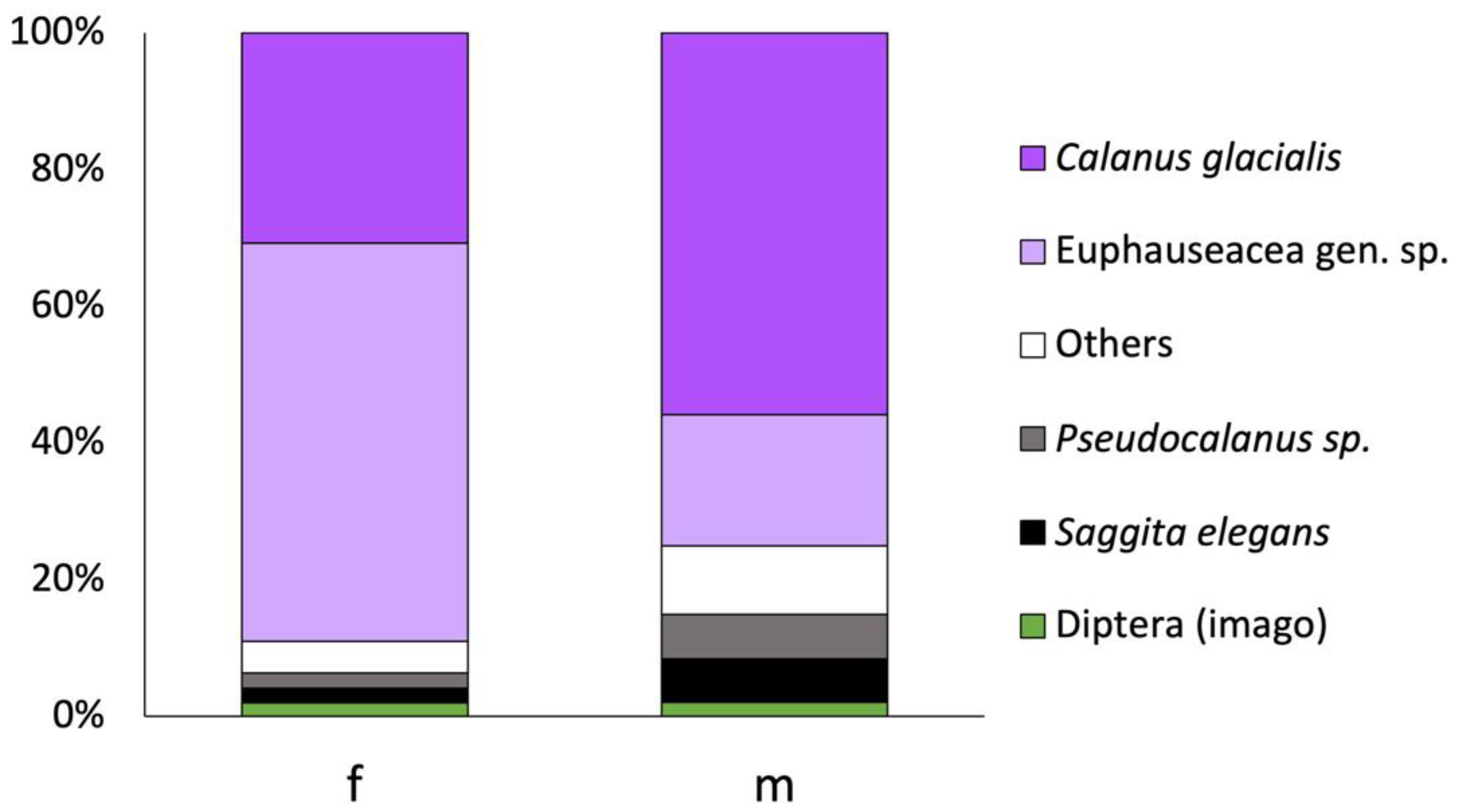
3.2. Stomach Content
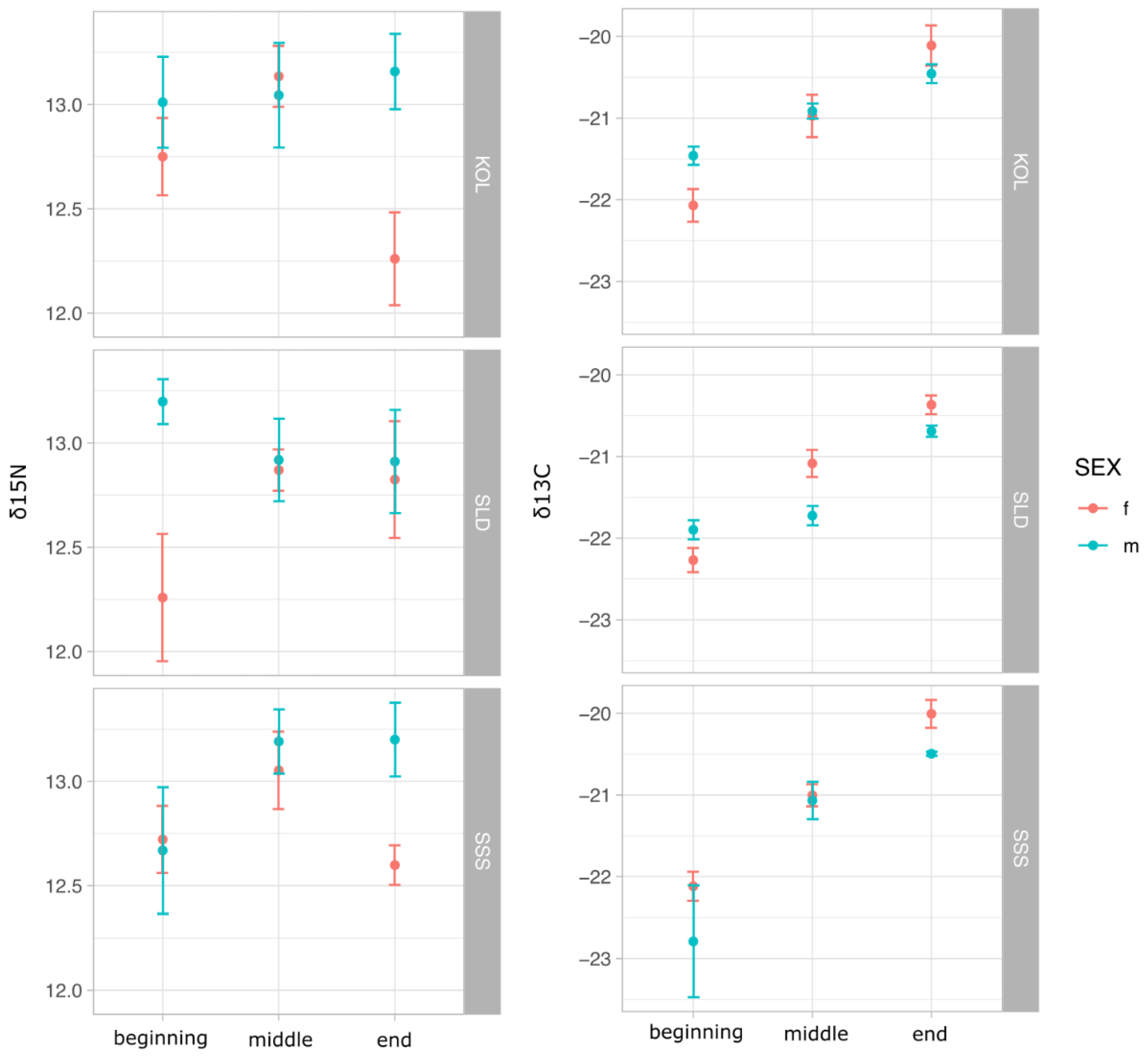
3.3. Stable Isotopes Values in Sticklebacks, Benthic and Planktonic Invertebrates
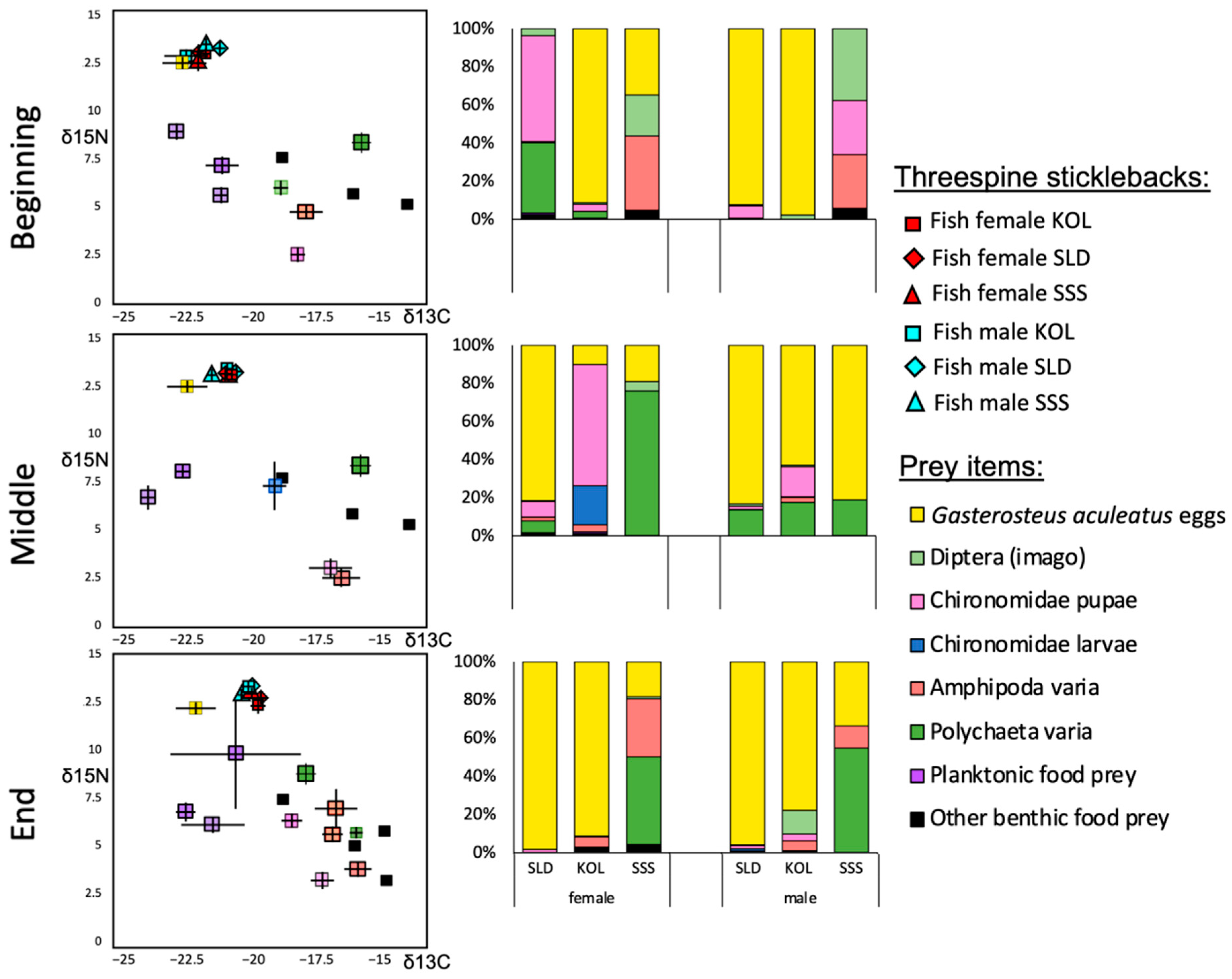
3.4. Comparing Stable Isotopes with Stomach Contents
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Special Report on the Ocean and Cryosphere in a Changing Climate; Pörtner, H.-O., Roberts, D.C., Masson-Delmotte, V., Zhai, P., Tignor, M., Poloczanska, E., Mintenbeck, K., Nicolai, M., Okem, A., Petzold, J., et al., Eds.; Cambridge University Press: New York, NY, USA, 2019; pp. 1–755.
- Steffen, W.; Rockström, J.; Richardson, K.; Lenton, T.M.; Folke, C.; Liverman, D.; Summerhayes, C.P.; Barnosky, A.D.; Cornell, S.E.; Crucifix, M.; et al. Trajectories of the Earth System in the Anthropocene. Proc. Natl. Acad. Sci. USA 2018, 115, 8252–8259. [Google Scholar] [CrossRef] [PubMed]
- Cusa, M.; Berge, J.; Varpe, Ø. Seasonal Shifts in Feeding Patterns: Individual and Population Realized Specialization in a High Arctic Fish. Ecol. Evol. 2019, 9, 11112–11121. [Google Scholar] [CrossRef] [PubMed]
- Fossheim, M.; Primicerio, R.; Johannesen, E.; Ingvaldsen, R.B.; Aschan, M.M.; Dolgov, A.V. Recent Warming Leads to a Rapid Borealization of Fish Communities in the Arctic. Nat. Clim. Chang. 2015, 5, 673–677. [Google Scholar] [CrossRef]
- Filatov, N.; Pozdnyakov, D.; Johannessen, O.M.; Pettersson, L.H.; Bobylev, L.P. White Sea: Its Marine Environment and Ecosystem Dynamics Influenced by Global Change; Springer Science & Business Media: New York, NY, USA, 2007. [Google Scholar]
- Lajus, D.L.; Dmitrieva, Z.V.; Kraikovski, A.V.; Lajus, J.A.; Alexandrov, D.A. Atlantic Salmon Fisheries in the White and Barents Sea Basins: Dynamic of Catches in the 17–18th Century and Comparison with 19–20th Century Data. Fish. Res. 2007, 87, 240–254. [Google Scholar] [CrossRef]
- Studenov, I.; Chupov, D.; Ustyuzhinskiy, G.; Tortsev, A. Results of atlantic salmon investigations in northern dvina river during fishing for research purposes. Fisheries 2020, 2020, 64–70. [Google Scholar] [CrossRef]
- Lajus, D.L.; Alekseeva, Y.I.; Lajus, J.A. Herring Fisheries in the White Sea in the 18th-Beginning of the 20th Centuries: Spatial and Temporal Patterns and Factors Affecting the Catch Fluctuations. Fish. Res. 2007, 87, 255–259. [Google Scholar] [CrossRef]
- Yershov, P.; Sukhotin, A. Age and Growth of Marine Three-Spined Stickleback in the White Sea 50 Years after a Population Collapse. Polar Biol. 2015, 38, 1813–1823. [Google Scholar] [CrossRef]
- Yurtseva, A.; Noreikiene, K.; Lajus, D.; Li, Z.; Alapassi, T.; Ivanova, T.; Ivanov, M.; Golovin, P.; Vesala, S.; Merilä, J. Aging Three-Spined Sticklebacks Gasterosteus Aculeatus: Comparison of Estimates from Three Structures. J. Fish Biol. 2019, 95, 802–811. [Google Scholar] [CrossRef]
- Ivanova, T.S.; Ivanov, M.V.; Golovin, P.V.; Polyakova, N.V.; Lajus, D.L. The White Sea Threespine Stickleback Population: Spawning Habitats, Mortality, and Abundance. Evol. Ecol. Res. 2016, 17, 301–315. [Google Scholar]
- Murzina, S.A.; Nefedova, Z.A.; Pekkoeva, S.N.; Lajus, D.L.; Nemova, N.N. Fatty Acids of the Three-Spined Stickleback (Gasterosteus Aculeatus L.) from the White Sea. Appl. Biochem. Microbiol. 2019, 55, 73–77. [Google Scholar] [CrossRef]
- Lajus, D.L.; Golovin, P.V.; Zelenskaia, A.E.; Demchuk, A.S.; Dorgham, A.S.; Ivanov, M.V.; Ivanova, T.S.; Murzina, S.A.; Polyakova, N.V.; Rybkina, E.V.; et al. Threespine Stickleback of the White Sea: Population Characteristics and Role in the Ecosystem. Contemp. Probl. Ecol. 2020, 13, 132–145. [Google Scholar] [CrossRef]
- Lajus, D.; Ivanova, T.; Rybkina, E.; Lajus, J.; Ivanov, M. Multidecadal Fluctuations of Threespine Stickleback in the White Sea and Their Correlation with Temperature. ICES J. Mar. Sci. 2021, 78, 653–665. [Google Scholar] [CrossRef]
- Cury, P.; Bakun, A.; Crawford, R.J.M.; Jarre, A.; Quiñones, R.A.; Shannon, L.J.; Verheye, H.M. Small Pelagics in Upwelling Systems: Patterns of Interaction and Structural Changes in “wasp-Waist” Ecosystems. ICES J. Mar. Sci. 2000, 57, 603–618. [Google Scholar] [CrossRef]
- Bakhvalova, A.E.; Ivanova, T.S.; Ivanov, M.V.; Demchuk, A.S.; Movchan, E.A.; Lajus, D.L. Long-Term Changes in the Role of Threespine Stickleback (Gasterosteus Aculeatus) in the White Sea: Predatory Fish Consumption Reflects Fluctuating Stickleback Abundance during the Last Century. Evol. Ecol. Res. 2016, 17, 317–334. [Google Scholar]
- Demchuk, A.; Ivanov, M.; Ivanova, T.; Polyakova, N.; Mas-Martí, E.; Lajus, D. Feeding Patterns in Seagrass Beds of Three-Spined Stickleback Gasterosteus Aculeatus Juveniles at Different Growth Stages. J. Mar. Biol. Assoc. UK 2015, 95, 1635–1643. [Google Scholar] [CrossRef]
- Fry, B. Food Web Structure on Georges Bank from Stable C, N, and S Isotopic Compositions. Limnol. Oceanogr. 1988, 33, 1182–1190. [Google Scholar] [CrossRef]
- Zanden, M.J.V.; Rasmussen, J.B. Primary Consumer δ13 C and δ15 N and the Trophic Position of Aquatic Consumers. Ecology 1999, 80, 1395–1404. [Google Scholar] [CrossRef]
- Post, D.M. Using Stable Isotopes to Estimate Trophic Position: Models, Methods, and Assumptions. Ecology 2002, 83, 703–718. [Google Scholar] [CrossRef]
- Golubkov, S.M.; Berezina, N.A.; Gubelit, Y.I.; Demchuk, A.S.; Golubkov, M.S.; Tiunov, A.V. A Relative Contribution of Carbon from Green Tide Algae Cladophora Glomerata and Ulva Intestinalis in the Coastal Food Webs in the Neva Estuary (Baltic Sea). Mar. Pollut. Bull. 2018, 126, 43–50. [Google Scholar] [CrossRef]
- Croizier, G.L.; Schaal, G.; Point, D.; Loc’h, F.L.; Machu, E.; Fall, M.; Munaron, J.-M.; Boyé, A.; Walter, P.; Laë, R. Stable Isotope Analyses Revealed the Influence of Foraging Habitat on Mercury Accumulation in Tropical Coastal Marine Fish. Sci. Total Environ. 2019, 650, 2129–2140. [Google Scholar] [CrossRef]
- Costalago, D.; Forster, I.; Nemcek, N.; Neville, C.; Perry, R.I.; Young, K.; Hunt, B.P.V. Seasonal and Spatial Dynamics of the Planktonic Trophic Biomarkers in the Strait of Georgia (Northeast Pacific) and Implications for Fish. Sci. Rep. 2020, 10, 8517. [Google Scholar] [CrossRef]
- Zanden, M.J.V.; Cabana, G.; Rasmussen, J.B. Comparing Trophic Position of Freshwater Fish Calculated Using Stable Nitrogen Isotope Ratios (Δ15N) and Literature Dietary Data. Can. J. Fish. Aquat. Sci. 1997, 54, 1142–1158. [Google Scholar] [CrossRef]
- Zanden, M.J.V.; Rasmussen, J.B. A Trophic Position Model of Pelagic Food Webs: Impact on Contaminant Bioaccumulation in Lake Trout. Ecol. Monogr. 1996, 66, 451–477. [Google Scholar] [CrossRef]
- Ravinet, M.; Ishikawa, A.; Kitano, J. Trophic Niche Differentiation and Phenotypic Divergence among Cryptic Species of Japanese Ninespine Sticklebacks. Evol. Ecol. Res. 2016, 17, 505–523. [Google Scholar]
- Golovin, P.V.; Bakhvalova, A.E.; Ivanov, M.V.; Ivanova, T.S.; Smirnova, K.A.; Lajus, D.L. Sex-Biased Mortality of Marine Threespine Stickleback (Gasterosteus Aculeatus L.) during Their Spawning Period in the White Sea. Evol. Ecol. Res. 2019, 20, 279–295. [Google Scholar]
- Ivanova, T.S.; Ivanov, M.V.; Bakhvalova, A.E.; Polyakova, N.V.; Golovin, P.V.; Kucheryavyy, A.V.; Yurtseva, A.O.; Smirnova, K.A.; Lajus, D.L. Homing Ability and Site Fidelity of Marine Threespine Stickleback on Spawning Grounds. Evol. Ecol. Res. 2019, 20, 297–315. [Google Scholar]
- Chislenko, L.L. Nomograms for Determining the Weight of Aquatic Organisms by Body Size and Form (Sea Mesobenthos and Plankton); Nauka: Moscow, Russia, 1968. [Google Scholar]
- Alimov, A.F.; Bogatov, V.V.; Golubkov, S.M. Production Hydrobiology; Nauka: Moscow, Russia, 2013. [Google Scholar]
- Pertzova, N.M. Average Mass and Sizes of Abundant Zooplankton Species in the White Sea. Oceanology 1967, 7, 309–313. [Google Scholar]
- Keough, J.R.; Sierszen, M.E.; Hagley, C.A. Analysis of a Lake Superior Coastal Food Web with Stable Isotope Techniques. Limnol. Oceanogr. 1996, 41, 136–146. [Google Scholar] [CrossRef]
- Hyslop, E.J. Stomach Contents Analysis—A Review of Methods and Their Application. J. Fish Biol. 1980, 17, 411–429. [Google Scholar] [CrossRef]
- Pinkas, L.; Olipham, M.S.; Iverson, I.L.K. Food Habits of Albacore; Bluef in Tuna and Bonito in Californian Waters. Calif. Fish Game 1971, 152, 1–105. [Google Scholar]
- Jost, L. Entropy and Diversity. Oikos 2006, 113, 363–375. [Google Scholar] [CrossRef]
- Hureau, J.-C. Biologie Cornparee de Quelques Poissons Antarctiques (Nototheniidae). Bull. L’inst. Oc’Enograph. Monaco 1969, 68, 1–244. [Google Scholar]
- Nilsen, M.; Pedersen, T.; Nilssen, E.M.; Fredriksen, S. Trophic studies in a high-latitude fjord ecosystem—A comparison of stable isotope analyses (δ13C and δ15N) and trophic-level estimates from a mass-balance model. Can. J. Fish. Aquat. Sci. 2008, 65, 2791–2806. [Google Scholar] [CrossRef]
- Stock, B.C.; Jackson, A.L.; Ward, E.J.; Parnell, A.C.; Phillips, D.L.; Semmens, B.X. Analyzing Mixing Systems Using a New Generation of Bayesian Tracer Mixing Models. PeerJ 2018, 6, e5096. [Google Scholar] [CrossRef]
- Britton, J.R.; Busst, G.M.A. Stable Isotope Discrimination Factors of Omnivorous Fishes: Influence of Tissue Type, Temperature, Diet Composition and Formulated Feeds. Hydrobiologia 2018, 808, 219–234. [Google Scholar] [CrossRef]
- Jakubavičiute, E.; Bergström, U.; Eklöf, J.S.; Haenel, Q.; Bourlat, S.J. DNA Metabarcoding Reveals Diverse Diet of the Three-Spined Stickleback in a Coastal Ecosystem. PLoS ONE 2017, 12, e0186929. [Google Scholar] [CrossRef]
- Snowberg, L.K.; Bolnick, D.I. Assortative Mating by Diet in a Phenotypically Unimodal but Ecologically Variable Population of Stickleback. Am. Nat. 2008, 172, 733–739. [Google Scholar] [CrossRef]
- Bolnick, D.I.; Snowberg, L.K.; Hirsch, P.E.; Lauber, C.L.; Knight, R.; Caporaso, J.G.; Svanbäck, R. Individuals’ Diet Diversity Influences Gut Microbial Diversity in Two Freshwater Fish (Threespine Stickleback and Eurasian Perch). Ecol. Lett. 2014, 17, 979–987. [Google Scholar] [CrossRef]
- Rybkina, E.V.; Demchuk, A.S.; Lajus, D.L.; Ivanova, T.S.; Ivanov, M.V.; Galaktionov, K.V. Dynamics of Parasite Community during Early Ontogenesis of Marine Threespine Stickleback, Gasterosteus Aculeatus. Evol. Ecol. Res. 2016, 17, 335–354. [Google Scholar]
- Whoriskey, F.G.; FitzGerald, G.J. Sex, Cannibalism and Sticklebacks. Behav. Ecol. Sociobiol. 1985, 18, 15–18. [Google Scholar] [CrossRef]
- Rohwer, S. Parent Cannibalism of Offspring and Egg Raiding as a Courtship Strategy. Am. Nat. 1978, 112, 429–440. [Google Scholar] [CrossRef]
- Mehlis, M.; Bakker, T.C.M.; Frommen, J.G. Nutritional Benefits of Filial Cannibalism in Three-Spined Sticklebacks (Gasterosteus aculeatus). Naturwissenschaften 2009, 96, 399–403. [Google Scholar] [CrossRef]
- Vadeboncoeur, Y.; Zanden, M.J.V.; Lodge, D.M. Putting the Lake Back Together: Reintegrating Benthic Pathways into Lake Food Web Models: Lake Ecologists Tend to Focus Their Research on Pelagic Energy Pathways, but, from Algae to Fish, Benthic Organisms Form an Integral Part of Lake Food Webs. BioScience 2002, 52, 44–54. [Google Scholar] [CrossRef]
- Kainz, M.J.; Hager, H.H.; Rasconi, S.; Kahilainen, K.K.; Amundsen, P.-A.; Hayden, B. Polyunsaturated Fatty Acids in Fishes Increase with Total Lipids Irrespective of Feeding Sources and Trophic Position. Ecosphere 2017, 8, e01753. [Google Scholar] [CrossRef]
- Reimchen, T.E.; Steeves, D.; Bergstrom, C.A. Sex Matters for Defence and Trophic Traits of Threespine Stickleback. Evol. Ecol. Res. 2016, 17, 459–485. [Google Scholar]
- MacNeil, M.; Skomal, G.; Fisk, A. Stable Isotopes from Multiple Tissues Reveal Diet Switching in Sharks. Mar. Ecol. Prog. Ser. 2005, 302, 199–206. [Google Scholar] [CrossRef]
- Richert, J.E.; Galván-Magaña, F.; Klimley, A.P. Interpreting Nitrogen Stable Isotopes in the Study of Migratory Fishes in Marine Ecosystems. Mar. Biol. 2015, 162, 1099–1110. [Google Scholar] [CrossRef]
- Demchuk, A.S.; Ivanov, M.V.; Ivanova, T.S.; Polyakova, N.V.; Golovin, P.V.; Lajus, D.L. Feeding of the threespine stickleback Gasterosteus aculeatus (Linnaeus, 1758) in spawning grounds. Tr. KNC RAN 2018, 4, 42–58. [Google Scholar] [CrossRef][Green Version]
- Bentzen, P.; McPhail, J.D. Ecology and Evolution of Sympatric Sticklebacks (Gasterosteus): Specialization for Alternative Trophic Niches in the Enos Lake Species Pair. Can. J. Zool. 1984, 62, 2280–2286. [Google Scholar] [CrossRef]
- Kristjánsson, B.K.; Skúlason, S.; Noakes, D.L. Rapid Divergence in a Recently Isolated Population of Threespine Stickleback (Gasterosteus Aculeatus L.). Evol. Ecol. Res. 2002, 4, 659–672. [Google Scholar]
- Reimchen, T.E.; Nosil, P. Replicated Ecological Landscapes and the Evolution of Morphological Diversity among Gasterosteus Populations from an Archipelago on the West Coast of Canada. Can. J. Zool. 2006, 84, 643–654. [Google Scholar] [CrossRef]
- Kitano, J.; Mori, S.; Peichel, C.L. Sexual Dimorphism in the External Morphology of the Threespine Stickleback (Gasterosteus Aculeatus). Copeia 2007, 2007, 336–349. [Google Scholar] [CrossRef]
- Spoljaric, M.A.; Reimchen, T.E. Habitat-Dependent Reduction of Sexual Dimorphism in Geometric Body Shape of Haida Gwaii Threespine Stickleback: Reduction in sexual dimorphism in stickleback. Biol. J. Linn. Soc. 2008, 95, 505–516. [Google Scholar] [CrossRef]
- Tamelander, T.; Renaud, P.E.; Hop, H.; Carroll, M.L.; Ambrose, W.G., Jr.; Hobson, K.A. Trophic Relationships and Pelagic–Benthic Coupling during Summer in the Barents Sea Marginal Ice Zone, Revealed by Stable Carbon and Nitrogen Isotope Measurements. Mar. Ecol. Prog. Ser. 2006, 310, 33–46. [Google Scholar] [CrossRef]
- Peltonen, H.; Vinni, M.; Lappalainen, A.; Pönni, J. Spatial Feeding Patterns of Herring (Clupea Harengus L.), Sprat (Sprattus Sprattus L.), and the Three-Spined Stickleback (Gasterosteus aculeatus L.) in the Gulf of Finland, Baltic Sea. ICES J. Mar. Sci. 2004, 61, 966–971. [Google Scholar] [CrossRef][Green Version]
- Grey, J. Trophic Fractionation and the Effects of Diet Switch on the Carbon Stable Isotopic ‘Signatures’ of Pelagic Consumers. Int. Ver. Für. Theor. Angew. Limnol. Verh. 2000, 27, 3187–3191. [Google Scholar] [CrossRef]
- Healy, K.; Kelly, S.B.A.; Guillerme, T.; Inger, R.; Bearhop, S.; Jackson, A.L. Predicting Trophic Discrimination Factor Using Bayesian Inference and Phylogenetic, Ecological and Physiological Data. DEsIR: Discrimination Estimation in R. PeerJ Preprints 2017, 1–21. [Google Scholar] [CrossRef]
- Brush, J.M.; Fisk, A.T.; Hussey, N.E.; Johnson, T.B. Spatial and Seasonal Variability in the Diet of Round Goby (Neogobius melanostomus): Stable Isotopes Indicate That Stomach Contents Overestimate the Importance of Dreissenids. Can. J. Fish. Aquat. Sci. 2012, 69, 573–586. [Google Scholar] [CrossRef]
- Beltran, R.S.; Peterson, S.H.; McHuron, E.A.; Reichmuth, C.; Hückstädt, L.A.; Costa, D.P. Seals and Sea Lions Are What They Eat, plus What? Determination of Trophic Discrimination Factors for Seven Pinniped Species. Rapid Commun. Mass Spectrom. 2016, 30, 1115–1122. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Seldianaya Inlet (SLD) | Koliushkovaya Lagoon (KOL) | Sukhaya Salma Strait (SSS) |
|---|---|---|---|
| Geographical coordinates | 66.33822° N, 33.62149° E | 66.31307° N, 33.64644° E | 66.31208° N, 33.65021° E |
| General description | Triangular inlet 120 × 240 m with wide entrance and shallow top. Average depth is 3.0 m | Isolated lagoon 200 × 540 m with average depth 1.5 m | Open strait with slope 6–8 cm/m in the study area |
| Tide amplitude, m | Up to 2.5 | Up to 0.3 | Up to 2.5 |
| Surface water temperature at sampling in 2016 | 12 °C (May)–20 °C (July) | 14 °C (May)–22 °C (July) | 12 °C (May)–20 °C (July) |
| Surface salinity at sampling in 2016 | 23 ppt (May)–24 ppt (July) | 15 ppt (May)–15 ppt (July) | 21 ppt (May)–19 ppt (July) |
| Bottom type | Stony littoral and muddy sublittoral zones | Muddy littoral and sublittoral zones | Stony littoral and muddy and sandy sublittoral zones |
| Aquatic vegetation | Fucoids in the littoral zone, dense eelgrass Zostera marina beds with dry biomass 1 kg/m2 and projective cover—up to 100% | Eelgrass beds near the sea entrance with dry biomass up to 0.1 kg/m2 and projective cover up to 30%, filamentous algae | Fucoids in the littoral, eelgrass with dry biomass up to 0.003 kg/m2 in sublittoral zone |
| Taxa | Offshore Site | Inshore Sites | ||
|---|---|---|---|---|
| IRI, % (F) | IRI, % (M) | IRI, % (F) | IRI, % (M) | |
| Diatomeae gen. sp. | 3.2 | 0.3 | 1.5 | 0.0 |
| Calanus glacialis | 20.7 | 61.6 | - | - |
| Oithona similis | 2.5 | 0.2 | - | - |
| Pseudocalanus sp. | 9.3 | 19.7 | - | - |
| Copepoditii Copepoda | 16.0 | 3.0 | - | - |
| Euphauseacea gen. sp. | 33.9 | 9.5 | - | - |
| Gastropoda varia | 0.0 | 0.2 | 5.8 | 0.5 |
| Polychaeta varia | 0.0 | 0.0 | 6.5 | 1.7 |
| Amphipoda varia | 0.5 | 0.0 | 1.7 | 0.5 |
| Chironomidae varia | - | - | 8.0 | 4.2 |
| Diptera (imago) | 1.1 | 1.0 | 1.8 | 1.8 |
| Gasterosteus aculeatus eggs | - | - | 67.8 | 82.0 |
| Other planktonic food prey | 12.7 | 3.4 | 5.6 | 9.3 |
| Other benthic food prey | 0.0 | 1.0 | 0.4 | 0.01 |
| Sex and Spawning Period | n | δ13C | δ15N | δC13 Range | δN15 Range | TP Observed | TP Expected |
|---|---|---|---|---|---|---|---|
| Threespine stickleback Gasterosteus aculeatus | |||||||
| Females, beginning | 15 | −22.15 ± 0.101 | 12.58 ± 0.140 | −22.76 to −21.51 | 11.19 to 13.24 | 5.22 ± 0.044 | 4.2 |
| Females, middle | 15 | −21.02 ± 0.108 | 13.02 ± 0.087 | −21.57 to −20.33 | 12.41 to 13.65 | 5.32 ± 0.027 | 4.2 |
| Females, end | 15 | −20.16 ± 0.111 | 12.56 ± 0.135 | −20.75 to -19.34 | 11.53 to 13.59 | 5.04 ± 0.042 | 3.8 |
| Males, beginning | 15 | −22.05 ± 0.272 | 12.96 ± 0.138 | −25.48 to −21.26 | 12.07 to 13.8 | 5.34 ± 0.043 | 4.4 |
| Males, middle | 15 | −21.24 ± 0.131 | 13.05 ± 0.117 | −22.1 to −20.64 | 12.11 to 13.75 | 5.33 ± 0.037 | 4.1 |
| Males, end | 15 | −20.55 ± 0.052 | 13.09 ± 0.118 | −20.85 to −20.07 | 12.11 to 13.82 | 5.21 ± 0.037 | 4.2 |
| Stickleback eggs | 3 | −22.45 ± 0.315 | 12.63 ± 0.104 | −22.81 to −21.95 | 12.47 to 12.75 | 5.2 ± 0.033 | |
| Prey organism | |||||||
| Amphipoda | 15 | −16.98 ± 0.249 | 4.86 ± 0.451 | −18.93 to −15.67 | 2.4 to 8.61 | 2.66 ± 0.124 | |
| Chironomidae | 21 | −18.47 ± 0.217 | 4.96 ± 0.421 | −20.12 to −15.79 | 2.27 to 6.88 | 2.73 ± 0.125 | |
| Gastropoda | 12 | −15.06 ± 0.341 | 4.91 ± 0.31 | −16.97 to −13.21 | 3.2 to 6.13 | 2.65 ± 0.095 | |
| Isopoda | 3 | −16.27 ± 0.166 | 5.76 ± 0.177 | −16.49 to −16.03 | 5.6 to 6.05 | 2.86 ± 0.052 | |
| Oligochaeta | 3 | −19.01 ± 0.36 | 7.56 ± 0.214 | −19.52 to −18.51 | 7.31 to 7.9 | 3.56 ± 0.063 | |
| Polychaeta | 5 | −16.48 ± 0.64 | 8.3 ± 0.14 | −18.11 to −15.71 | 8.12 to 8.65 | 3.2 ± 0.567 | |
| Plankton | 22 | −22.22 ± 0.373 | 7.26 ± 0.394 | −24.64 to −16.65 | 5.36 to 12.14 | 3.39 ± 0.114 | |
| Variables | Factor | ||
|---|---|---|---|
| Sex | Period | Site | |
| δ13C | 0.12 | <0.01 | 0.03 |
| δ15N | <0.01 | 0.07 | 0.79 |
| TP | <0.01 | <0.01 | 0.86 |
| Diet | Female | Male | ||||
|---|---|---|---|---|---|---|
| Beginning | Middle | End | Beginning | Middle | End | |
| Amphipoda | <0.01–0.01 0.001 | <0.01–<0.01 <0.001 | <0.01–0.03 0.005 | – – | <0.01–<0.01 <0.001 | – – |
| Chironomidae | <0.01–0.01 0.001 | <0.01–<0.01 <0.001 | <0.01–0.57 0.164 | <0.01–<0.01 <0.001 | <0.01–<0.01 <0.001 | <0.01–<0.01 <0.001 |
| Stickleback eggs | 0.97–1.00 0.994 | 0.80–1.00 0.948 | <0.01–1.00 0.563 | 0.99–1.00 0.999 | 0.87–1.00 0.985 | 0.99–1.00 0.999 |
| Gastropoda | <0.01–0.01 0.001 | <0.01–<0.01 <0.001 | <0.01–0.01 0.001 | <0.01–<0.01 <0.001 | <0.01–<0.01 <0.001 | <0.01–<0.01 <0.001 |
| Isopoda | <0.01–0.01 0.001 | – – | <0.01–0.07 0.009 | <0.01–<0.01 <0.001 | <0.01–<0.01 <0.001 | <0.01–<0.01 <0.001 |
| Oligochaeta | <0.01–0.02 0.002 | <0.01–0.03 0.002 | <0.01–0.16 0.018 | <0.01–<0.01 <0.001 | <0.01–<0.01 <0.001 | <0.01–<0.01 <0.001 |
| Polychaeta | <0.01–0.01 0.001 | <0.01–0.17 0.048 | <0.01–0.28 0.094 | <0.01–<0.01 <0.001 | <0.01–0.13 0.014 | <0.01–<0.01 <0.001 |
| Mixed zooplankton | <0.01–0.01 0.001 | <0.01–<0.01 <0.001 | <0.01–0.50 0.146 | <0.01–<0.01 <0.001 | <0.01–<0.01 <0.001 | <0.01–<0.01 <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Genelt-Yanovskaya, A.S.; Polyakova, N.V.; Ivanov, M.V.; Nadtochii, E.V.; Ivanova, T.S.; Genelt-Yanovskiy, E.A.; Tiunov, A.V.; Lajus, D.L. Tracing the Food Web of Changing Arctic Ocean: Trophic Status of Highly Abundant Fish, Gasterosteus aculeatus (L.), in the White Sea Recovered Using Stomach Content and Stable Isotope Analyses. Diversity 2022, 14, 955. https://doi.org/10.3390/d14110955
Genelt-Yanovskaya AS, Polyakova NV, Ivanov MV, Nadtochii EV, Ivanova TS, Genelt-Yanovskiy EA, Tiunov AV, Lajus DL. Tracing the Food Web of Changing Arctic Ocean: Trophic Status of Highly Abundant Fish, Gasterosteus aculeatus (L.), in the White Sea Recovered Using Stomach Content and Stable Isotope Analyses. Diversity. 2022; 14(11):955. https://doi.org/10.3390/d14110955
Chicago/Turabian StyleGenelt-Yanovskaya, Anna S., Natalia V. Polyakova, Mikhail V. Ivanov, Ekaterina V. Nadtochii, Tatiana S. Ivanova, Evgeny A. Genelt-Yanovskiy, Alexei V. Tiunov, and Dmitry L. Lajus. 2022. "Tracing the Food Web of Changing Arctic Ocean: Trophic Status of Highly Abundant Fish, Gasterosteus aculeatus (L.), in the White Sea Recovered Using Stomach Content and Stable Isotope Analyses" Diversity 14, no. 11: 955. https://doi.org/10.3390/d14110955
APA StyleGenelt-Yanovskaya, A. S., Polyakova, N. V., Ivanov, M. V., Nadtochii, E. V., Ivanova, T. S., Genelt-Yanovskiy, E. A., Tiunov, A. V., & Lajus, D. L. (2022). Tracing the Food Web of Changing Arctic Ocean: Trophic Status of Highly Abundant Fish, Gasterosteus aculeatus (L.), in the White Sea Recovered Using Stomach Content and Stable Isotope Analyses. Diversity, 14(11), 955. https://doi.org/10.3390/d14110955