
Global Patterns of Earwig Species Richness


Abstract
1. Introduction
2. Materials and Methods
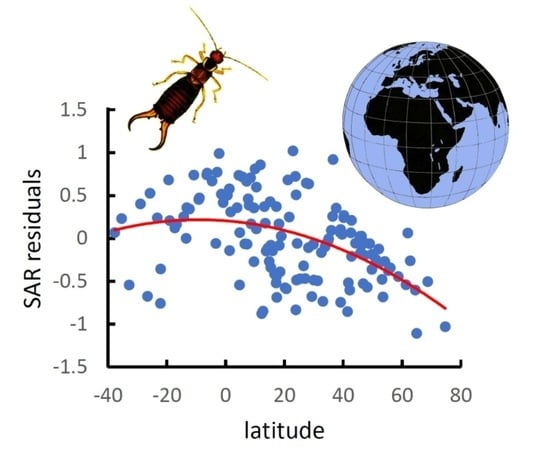
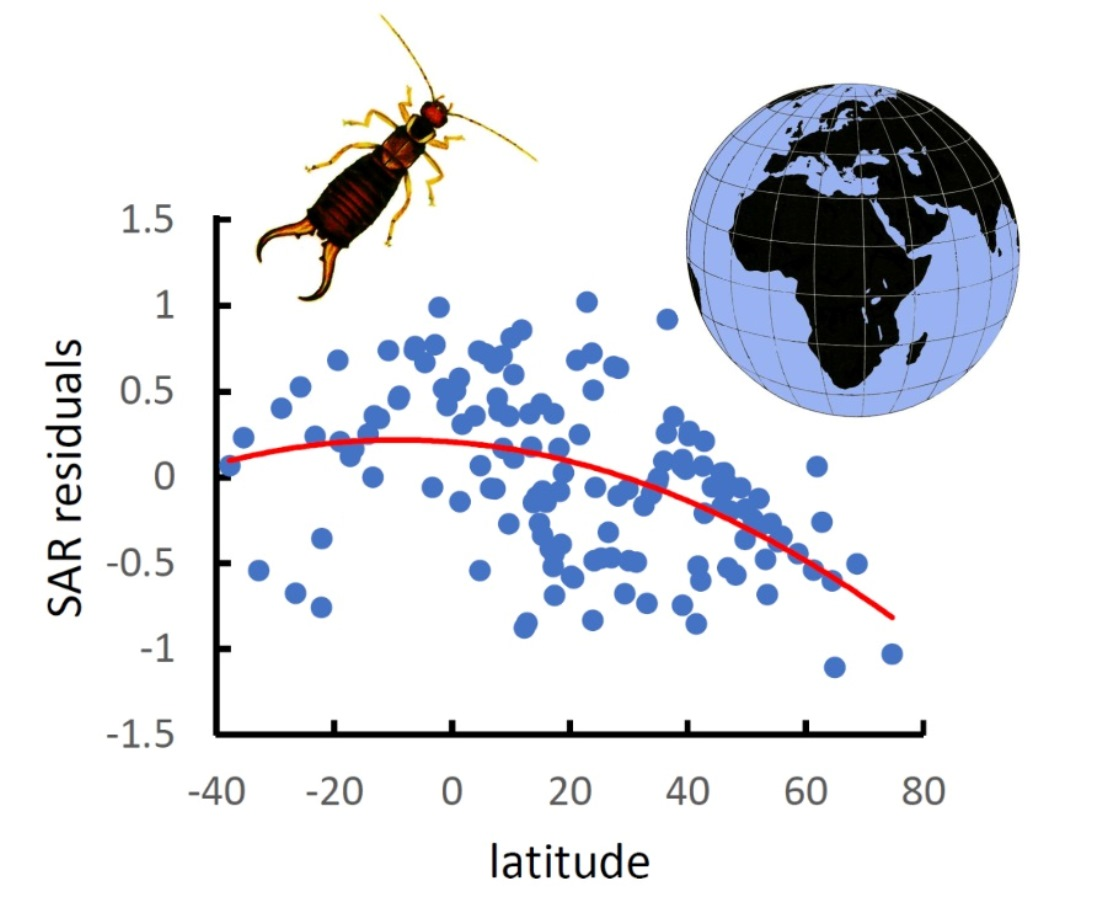
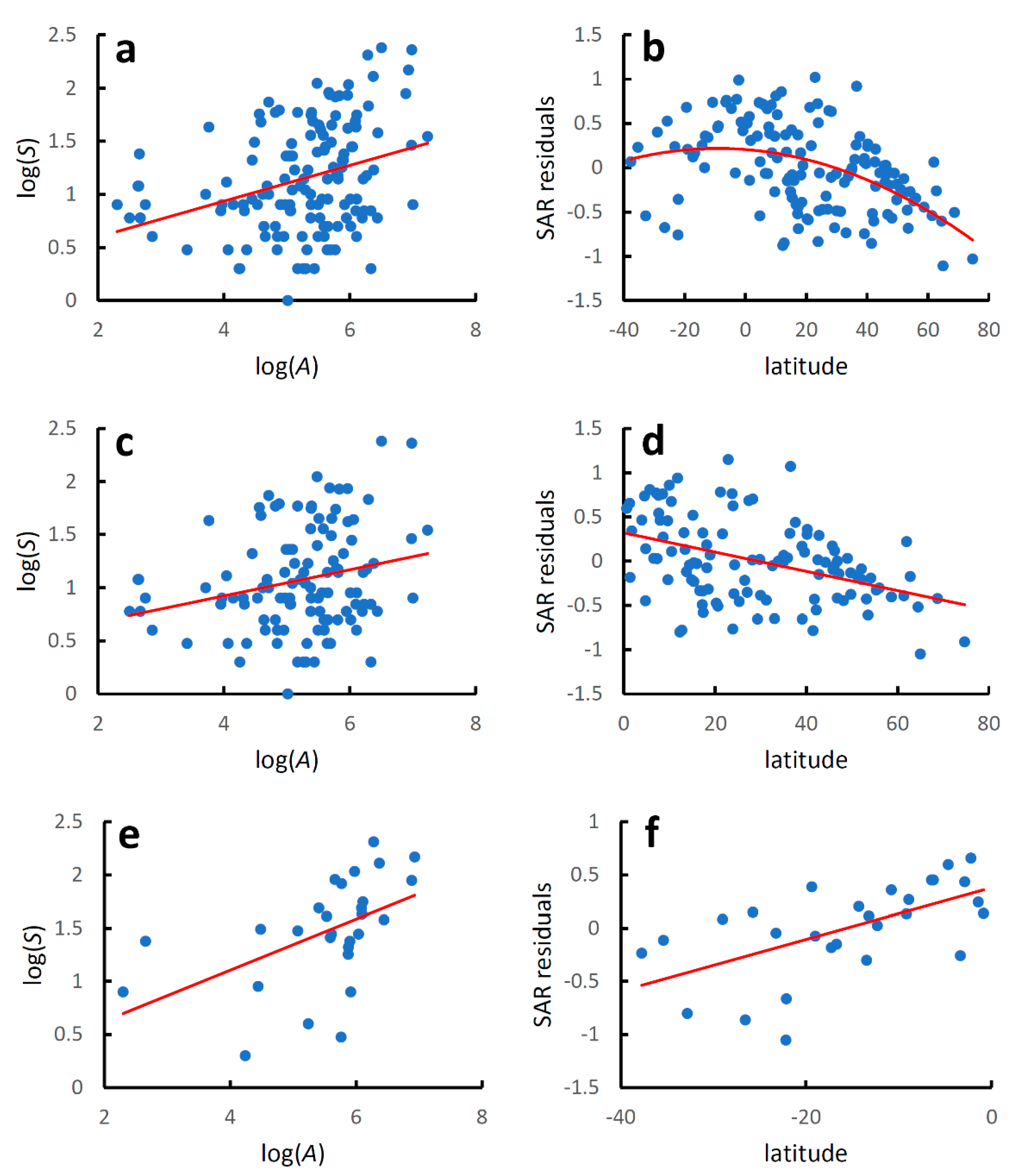
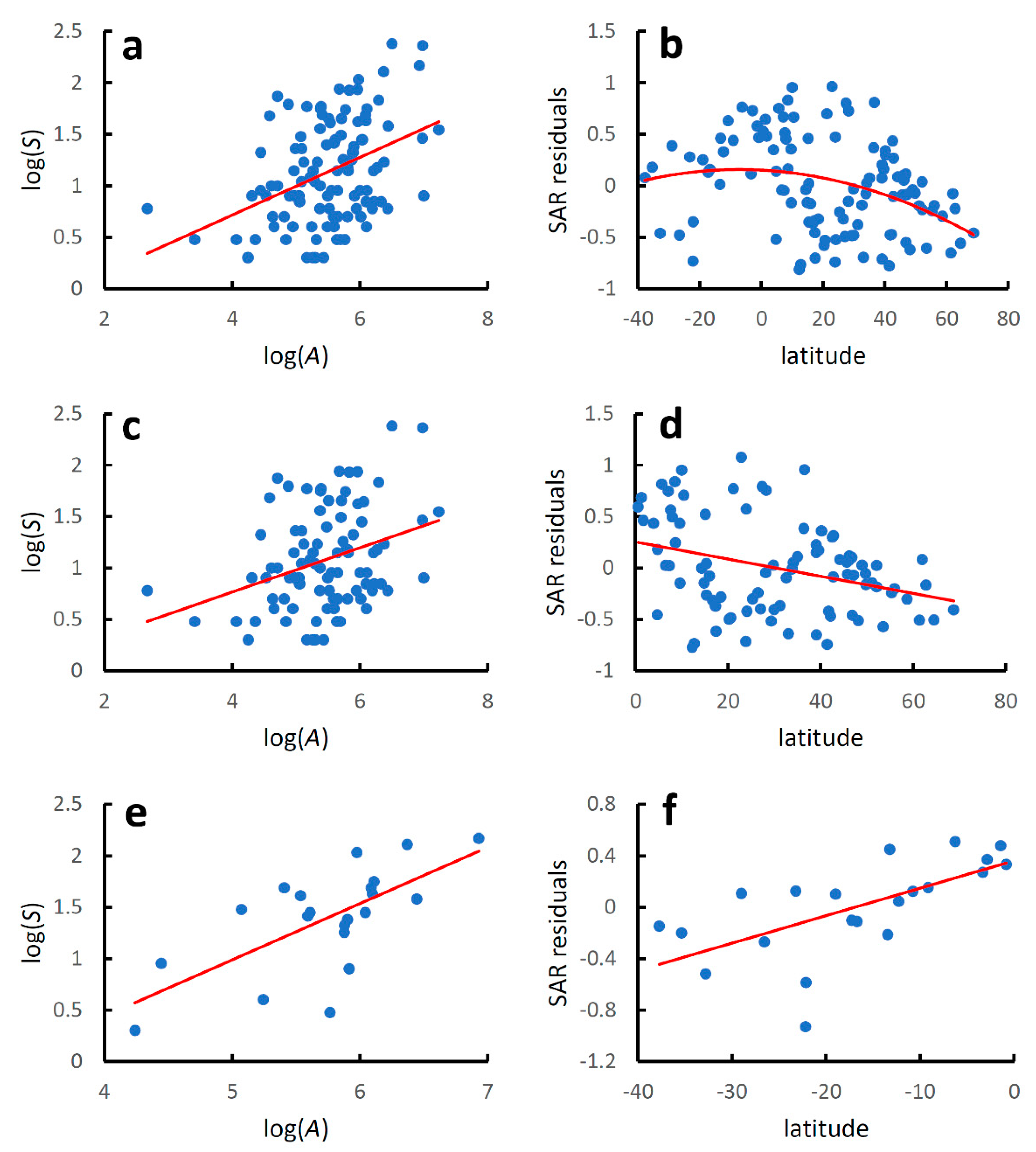
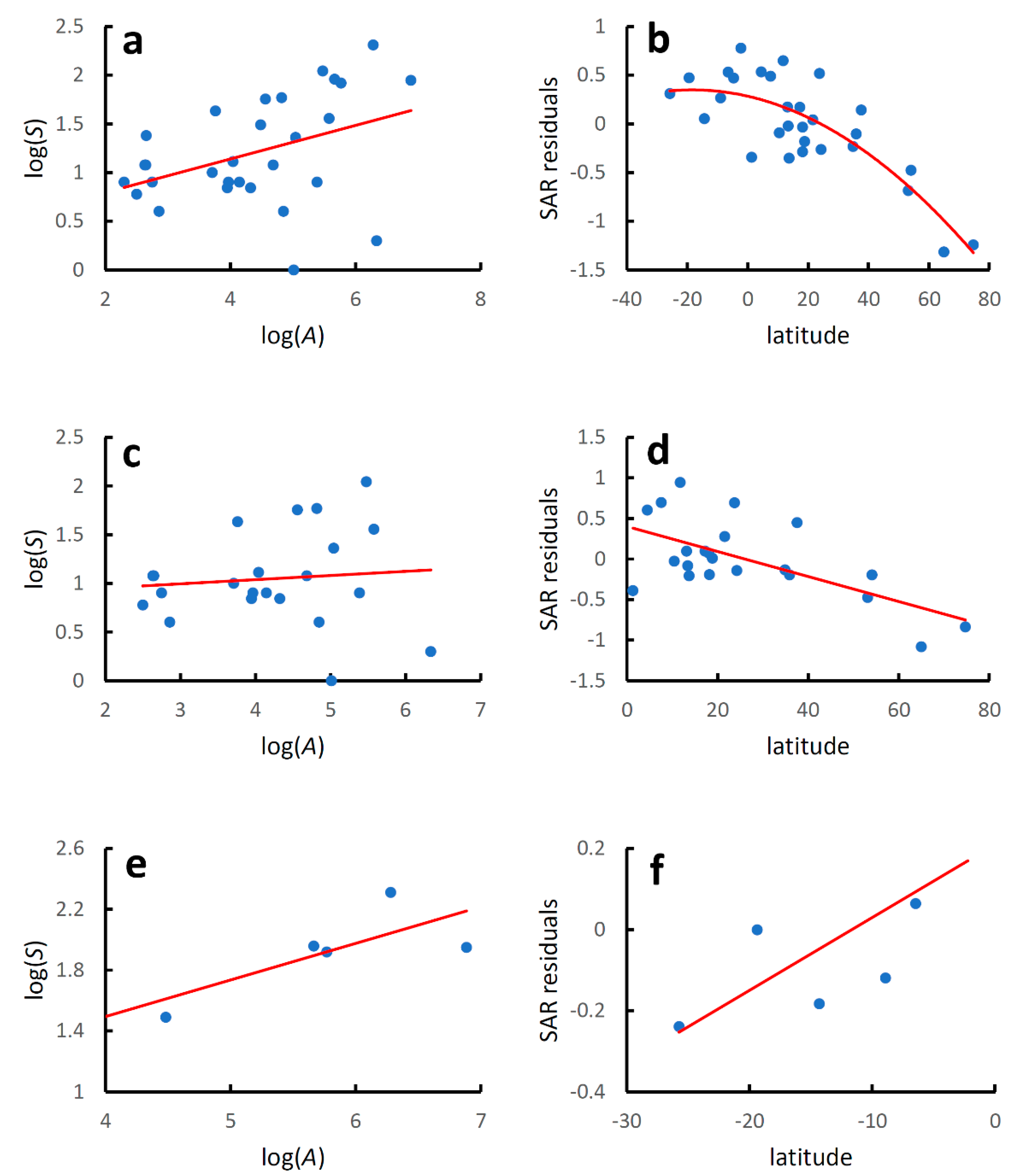
3. Results
4. Discussion
- (1)
- Time: as tropics were not severely affected by glaciations, they were occupied for longer periods, providing more time for cladogenesis;
- (2)
- Environmental heterogeneity: tropical areas have greater environmental heterogeneity that increases the probability of species coexistence through niche differentiation;
- (3)
- Competition: while temperate populations may be more controlled by abiotic factors (seasonality), tropical populations are more regulated by biotic interactions, such as competition, which increases species diversity through specialization;
- (4)
- Predation: tropics have more predators that maintain prey species at low densities, thus decreasing competition among prey species and hence increasing the coexistence of prey species (this hypothesis suggests a mechanism exactly opposite to that of the competition hypothesis);
- (5)
- Productivity: tropics support more species because more resources are available, allowing for more specialization;
- (6)
- Environmental (climatic) stability: severe and/or unpredictable climates tend to have lower habitat diversity, whereas stable climates tend to present higher habitat diversity, which, in turn, promotes species diversity (as in the environmental heterogeneity hypothesis).
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pianka, E.R. Latitudinal Gradients in Species Diversity: A Review of Concepts. Am. Nat. 1966, 100, 33–46. [Google Scholar] [CrossRef]
- Rohde, K. Latitudinal gradients in species diversity: The search for the primary cause. Oikos 1992, 65, 514–527. [Google Scholar] [CrossRef]
- Willig, M.R.; Kaufman, D.M.; Stevens, R.D. Latitudinal gradients of biodiversity: Pattern, process, scale, and synthesis. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 273–309. [Google Scholar] [CrossRef]
- Hillebrand, H. On the generality of the latitudinal diversity gradient. Am. Nat. 2004, 163, 192–211. [Google Scholar] [CrossRef] [PubMed]
- Lomolino, M.V.; Riddle, B.R.; Whittaker, R.J.; Brown, J.H. Biogeography, 4th ed.; Sinauer Associates Inc.: Sunderland, MA, USA, 2010. [Google Scholar]
- Brown, J.H. Why are there so many species in the tropics? J. Biogeogr. 2014, 41, 8–22. [Google Scholar] [CrossRef] [PubMed]
- Gillman, L.N.; Wright, S.D. Species richness and evolutionary speed: The influence of temperature, water and area. J. Biogeogr. 2014, 41, 39–51. [Google Scholar] [CrossRef]
- Fine, P.V.A. Ecological and evolutionary drivers of geographic variation in species diversity. Annu. Rev. Ecol. Evol. Syst. 2015, 46, 369–392. [Google Scholar] [CrossRef]
- Jablonski, D.; Huang, S.; Roy, K.; Valentine, J.W. Shaping the latitudinal diversity gradient: New perspectives from a synthesis of paleobiology and biogeography. Am. Nat. 2017, 189, 1–12. [Google Scholar] [CrossRef]
- Schemske, D.W.; Mittelbach, G.G. “Latitudinal gradients in species diversity”: Reflections on Pianka’s 1966 article and a look forward. Am. Nat. 2017, 189, 599–603. [Google Scholar] [CrossRef] [PubMed]
- Kinlock, N.L.; Prowant, L.; Herstoff, E.M.; Foley, C.M.; Akin-Fajiye, M.; Bender, N.; Umarani, M.; Ryu, H.Y.; Sen, H.Y.; Gurevitch, J.; et al. Explaining global variation in the latitudinal diversity gradient: Meta-analysis confirms known patterns and uncovers new ones. Glob. Ecol. Biogeogr. 2018, 27, 125–141. [Google Scholar] [CrossRef]
- Saupe, E.E.; Myers, C.E.; Townsend Peterson, A.; Soberon, J.; Singarayer, J.; Valdes, P.; Qiao, H. Spatio–temporal climate change contributes to latitudinal diversity gradients. Nat. Ecol. Evol. 2019, 3, 1419–1429. [Google Scholar] [CrossRef] [PubMed]
- Beaugrand, G.; Kirby, R.; Goberville, E. The mathematical influence on global patterns of biodiversity. Ecol. Evol. 2020, 10, 6494–6511. [Google Scholar] [CrossRef] [PubMed]
- Fuhrman, J.A.; Steele, J.A.; Hewson, I.; Schwalbach, M.S.; Brown, M.V.; Green, J.L.; Brown, J.H. A latitudinal diversity gradient in planktonic marine bacteria. Proc. Natl. Acad. Sci. USA 2008, 105, 7774–7778. [Google Scholar] [CrossRef] [PubMed]
- Hillebrand, H.; Azovsky, A.I. Body size determines the strength of the latitudinal diversity gradient. Ecography 2001, 24, 251–256. [Google Scholar] [CrossRef]
- Culver, S.J.; Buzas, M.A. Global latitudinal species diversity gradient in deep–sea benthic foraminifera. Deep-Sea Res. 2000, 47, 259–275. [Google Scholar] [CrossRef]
- Dolan, J.R.; Gallegos, C.L. Estuarine diversity of tintinnids (planktonic ciliates). J. Plankton Res. 2001, 23, 1009–1027. [Google Scholar] [CrossRef][Green Version]
- Nunn, C.L.; Altizer, S.M.; Sechrest, W.; Cunningham, A.A. Latitudinal Gradients of Parasite Species Richness in Primates. Divers. Distrib. 2005, 11, 249–256. [Google Scholar] [CrossRef]
- Flessa, K.W.; Jablonski, D. Biogeography of Recent marine bivalve molluscs and its implications for paleobiogeography and the geography of extinction: A progress report. Histor. Biol. 1995, 10, 25–47. [Google Scholar] [CrossRef]
- Fortes, R.R.; Absalão, R.S. The Applicability of Rapoport’s Rule to the Marine Molluscs of the Americas. J. Biogeogr. 2004, 31, 1909–1916. [Google Scholar] [CrossRef]
- Jablonski, D.; Roy, K.; Valentine, J.W. Out of the tropics: Evolutionary dynamics of the latitudinal diversity gradient. Science 2006, 314, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Jablonski, D.; Belanger, C.L.; Berke, S.K.; Huang, S.; Krug, A.Z.; Roy, K.; Tomašových, A.; Valentine, J.W. Out of the tropics, but how? Fossils, bridge species, and thermal ranges in the dynamics of the marine latitudinal diversity gradient. Proc. Natl. Acad. Sci. USA 2013, 110, 10487–10494. [Google Scholar] [CrossRef]
- Johnson, P.T.J.; Haas, S.E. Why do parasites exhibit reverse latitudinal diversity gradients? Testing the roles of host diversity, habitat and climate. Glob. Ecol. Biogeogr. 2021, 30, 1810–1821. [Google Scholar] [CrossRef]
- Moreno, R.A.; Labra, F.A.; Cotoras, D.D.; Camus, P.A.; Gutiérrez, D.; Aguirre, L.; Rozbaczylo, N.; Poulin, E.; Lagos, N.A.; Zamorano, D.; et al. Evolutionary drivers of the hump–shaped latitudinal gradient of benthic polychaete species richness along the Southeastern Pacific coast. PeerJ 2021, 9, e12010. [Google Scholar] [CrossRef]
- Kotwicki, L.; Szymelfenig, M.; De Troch, M.; Urban–Malinga, B.; Węsławski, J.M. Latitudinal biodiversity patterns of meiofauna from sandy littoral beaches. Biodivers. Conserv. 2005, 14, 461–474. [Google Scholar] [CrossRef]
- Piel, W.H. The global latitudinal diversity gradient pattern in spiders. J. Biogeogr. 2018, 45, 1896–1904. [Google Scholar] [CrossRef]
- Reid, J.W. Latitudinal diversity patterns of continental benthic copepod species assemblages in the Americas. Hydrobiologia 1994, 292/293, 341–349. [Google Scholar] [CrossRef]
- Dworschak, P.C. Global diversity in the Thalassinidea (Decapoda). J. Crustac. Biol. 2000, 20, 238–245. [Google Scholar] [CrossRef]
- Chiu, W.T.R.; Yasuhara, M.; Cronin, T.M.; Hunt, G.; Gemery, L.; Wei, C.L. Marine latitudinal diversity gradients, niche conservatism and out of the tropics and Arctic: Climatic sensitivity of small organisms. J. Biogeogr. 2020, 47, 817–828. [Google Scholar] [CrossRef]
- Enquist, B.J.; Niklas, K.J. Invariant scaling relations across tree–dominated communities. Nature 2001, 410, 655–660. [Google Scholar] [CrossRef]
- Sax, D.F. Latitudinal gradients and geographic ranges of exotic species: Implications for biogeography. J. Biogeogr. 2001, 28, 139–150. [Google Scholar] [CrossRef]
- Qian, H.; Song, J.S.; Krestov, P.; Guo, Q.; Wu, Z.; Shen, X.; Guo, X. Large–scale phytogeographical patterns in East Asia in relation to latitudinal and climatic gradients. J. Biogeogr. 2003, 30, 129–141. [Google Scholar] [CrossRef]
- Davies, T.J.; Barraclough, T.G.; Savolainen, V.; Chase, M.W. Environmental causes for plant biodiversity gradients. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2004, 359, 1645–1656. [Google Scholar] [CrossRef]
- Mutle, J.; Barthlott, W. Patterns of vascular plant diversity at continental to global scales. Biol. Skr. 2005, 55, 521–531. [Google Scholar]
- Kerkhoff, A.J.; Moriarty, P.E.; Weiser, M.D. The latitudinal species richness gradient in New World woody angiosperms is consistent with the tropical conservatism hypothesis. Proc. Natl Acad. Sci. USA 2014, 111, 8125–8130. [Google Scholar] [CrossRef] [PubMed]
- Burnham, R.J.; Santanna, C.V. Distribution, diversity, and traits of native, exotic, and invasive climbing plants in Michigan. Brittonia 2015, 67, 350–370. [Google Scholar] [CrossRef]
- Chen, S.B.; Ferry Slik, J.W.; Gao, J.; Mao, L.F.; Bi, M.J.; Shen, M.W.; Zhou, K.X. Latitudinal diversity gradients in bryophytes and woody plants: Roles of temperature and water availability. J Syst. Evol. 2015, 53, 535–545. [Google Scholar] [CrossRef]
- Liu, H.; Yu, R.; Huang, J.; Liu, Y.; Zang, R.; Guo, Z.; Ding, Y.; Lu, X.; Li, Q.; Chen, H.Y.H. Latitudinal diversity gradients and Rapoport effects in Chinese endemic woody seed plants. Forests 2020, 11, 1029. [Google Scholar] [CrossRef]
- Oberdorff, T.; Guégan, J.-F.; Hugueny, B. Global scale patterns of fish species richness in rivers. Ecography 1995, 18, 345–352. [Google Scholar] [CrossRef]
- Kaufman, D.M.; Willig, M.R. Latitudinal patterns of mammalian species richness in the New World: The effects of sampling method and faunal group. J. Biogeogr. 1998, 25, 795–805. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Porter, E.E.; Diniz–Filho, J.A.F. Productivity and history as predictors of the latitudinal diversity gradient of terrestrial birds. Ecology 2003, 84, 1608–1623. [Google Scholar] [CrossRef]
- Patten, M.A. Correlates of species richness in North American bat families. J. Biogeogr. 2004, 31, 975–985. [Google Scholar] [CrossRef]
- Stevens, R.D. Untangling latitudinal richness gradients at higher taxonomic levels: Familial perspectives on the diversity of New World bat communities. J. Biogeogr. 2004, 31, 665–674. [Google Scholar] [CrossRef]
- Turner, J.R.G.; Hawkins, B.A. The global biodiversity gradient. In Frontiers of Biogeography: New Directions in the Geography of Nature; Lomolino, M.V., Heaney, L.R., Eds.; Sinauer Associates Inc.: Sunderland, MA, USA, 2004; pp. 171–190. [Google Scholar]
- Buckley, L.B.; Jetz, W. Environmental and historical constraints on global patterns of amphibian richness. Proc. R. Soc. Lond. Ser. B 2007, 274, 1167–1173. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, B.A.; Diniz–Filho, J.A.; Jaramillo, C.A.; Soeller, S.A. Climate, niche conservatism, and the global bird diversity gradient. Am. Nat. 2007, 170, S16–S27. [Google Scholar] [CrossRef] [PubMed]
- Fisher, J.A.; Frank, K.T.; Petrie, B.; Leggett, W.C.; Shackell, N.L. Temporal dynamics within a contemporary latitudinal diversity gradient. Ecol. Lett. 2008, 11, 883–897. [Google Scholar] [CrossRef] [PubMed]
- Schipper, J.; Chanson, J.S.; Chiozza, F.; Cox, N.A.; Hoffmann, M.; Katariya, V.; Lamoreux, J.; Rodrigues, A.S.; Stuart, S.N.; Temple, H.J.; et al. The status of the world’s land and marine mammals: Diversity, threat, and knowledge. Science 2008, 322, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Rolland, J.; Condamine, F.L.; Jiguet, F.; Morlon, H. Faster speciation and reduced extinction in the tropics contribute to the mammalian latitudinal diversity gradient. PLoS Biol. 2014, 12, e1001775. [Google Scholar] [CrossRef]
- Rolland, J.; Condamine, F.L.; Beeravolu, C.R.; Jiguet, F.; Morlon, H. Dispersal is a major driver of the latitudinal diversity gradient of Carnivora. Glob. Ecol. Biogeogr. 2015, 24, 1059–1071. [Google Scholar] [CrossRef]
- Pulido–Santacruz, P.; Weir, J.T. Extinction as a driver of avian latitudinal diversity gradients. Evolution 2016, 70, 860–872. [Google Scholar] [CrossRef]
- Hanly, P.J.; Mittelbach, G.G.; Schemske, D.W. Speciation and the latitudinal diversity gradient: Insights from the global distribution of endemic fish. Am. Nat. 2017, 189, 604–615. [Google Scholar] [CrossRef]
- Stork, N.E. How many species of insects and other terrestrial arthropods are there on Earth? Annu. Rev. Entomol. 2018, 63, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Eggleton, P. The state of the world’s insects. Ann. Rev. Environ. Resour. 2020, 45, 61–82. [Google Scholar] [CrossRef]
- Janzen, D.H. The peak in North American ichneumonid species richness lies between 38° and 42° N. Ecology 1981, 62, 532–537. [Google Scholar] [CrossRef]
- Davidowitz, G.; Rosenzweig, M.L. The latitudinal gradient of species diversity among North American grasshoppers (Acrididae) within a single habitat: A test of the spatial heterogeneity hypothesis. J. Biogeogr. 1998, 25, 553–560. [Google Scholar] [CrossRef]
- Dingle, H.; Rochester, W.; Zalucki, M. Relationships among climate, latitude and migration: Australian butterflies are not temperate–zone birds. Oecologia 2000, 124, 196–207. [Google Scholar] [CrossRef]
- Kocher, S.D.; Williams, E.H. The diversity and abundance of north American butterflies vary with habitat disturbance and geography. J. Biogeogr. 2000, 27, 785–794. [Google Scholar] [CrossRef]
- Lobo, J.M. Species diversity and composition of dung beetle (Coleoptera: Scarabaeoidea) assemblages in North America. Canadian Entomologist 2000, 132, 307–321. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Porter, E.E. Water–energy balance and the geographic pattern of species richness of western Palearctic butterflies. Ecol. Entomol. 2003, 28, 678–686. [Google Scholar] [CrossRef]
- Rodriguero, M.S.; Gorla, D.E. Latitudinal Gradient in Species Richness of the New World Triatominae (Reduviidae). Glob. Ecol. Biogeogr. 2004, 13, 75–84. [Google Scholar] [CrossRef]
- Schuldt, A.; Assmann, T. Environmental and historical effects on richness and endemism patterns of carabid beetles in the western Palaearctic. Ecography 2009, 32, 705–714. [Google Scholar] [CrossRef]
- Baselga, A. Determinants of species richness, endemism and turnover in European longhorn beetles. Ecography 2008, 31, 263–271. [Google Scholar] [CrossRef]
- Ulrich, W.; Fiera, C. Environmental correlates of species richness of European springtails (Hexapoda: Collembola). Acta Oecol. 2009, 35, 45–52. [Google Scholar] [CrossRef]
- Bąkowski, M.; Ulrich, W.; Laštůvka, Z. Environmental correlates of species richness of Sesiidae (Lepidoptera) in Europe. Eur. J. Entomol. 2010, 107, 563–570. [Google Scholar] [CrossRef]
- Hawkins, B.A. Multiregional comparison of the ecological and phylogenetic structure of butterfly species richness gradients. J. Biogeogr. 2010, 37, 647–656. [Google Scholar] [CrossRef]
- Fattorini, S.; Ulrich, W. Drivers of species richness in European Tenebrionidae (Coleoptera). Acta Oecol. 2012, 36, 255–258. [Google Scholar] [CrossRef]
- Burington, Z.L.; Inclán–Luna, D.J.; Pollet, M.; Stireman, J.O., III. Latitudinal patterns in tachinid parasitoid diversity (Diptera: Tachinidae): A review of the evidence. Insect Conserv. Divers. 2020, 13, 419–431. [Google Scholar] [CrossRef]
- Fernandes, G.W.; Price, P.W. Biogeographical gradients in galling species richness. Tests of hypotheses. Oecologia 1988, 76, 161–167. [Google Scholar] [CrossRef]
- Cushman, J.; Lawton, J.; Manly, B. Latitudinal patterns in European ant assemblages: Variation in species richness and body size. Oecologia 1993, 95, 30–37. [Google Scholar] [CrossRef]
- Andrew, N.R.; Hughes, L. Diversity and assemblage structure of phytophagous Hemiptera along a latitudinal gradient: Predicting the potential impacts of climate change. Glob. Ecol. Biogeogr. 2005, 14, 249–262. [Google Scholar] [CrossRef]
- Fattorini, S. Testing the latitudinal gradient: A narrow scale analysis of tenebrionid richness (Coleoptera, Tenebrionidae) in the Aegean archipelago (Greece). Ital. J. Zool. 2006, 73, 203–211. [Google Scholar] [CrossRef]
- Cancello, E.M.; Silva, R.R.; Vasconcellos, A.; Reis, Y.T.; Oliveira, L.M. Latitudinal variation in termite species richness and abundance along the Brazilian Atlantic forest hotspot. Biotropica 2014, 46, 441–450. [Google Scholar] [CrossRef]
- Fattorini, S. Tenebrionid beetle distributional patterns in Italy: Multiple colonization trajectories in a biogeographical crossroad. Insect Conserv. Divers. 2014, 7, 144–160. [Google Scholar] [CrossRef]
- Yuan, S.; Huang, M.; Wang, X.; Ji, L.; Zhang, Y. Centers of endemism and diversity patterns for typhlocybine leafhoppers (Hemiptera: Cicadellidae: Typhlocybinae) in China. Insect Sci. 2014, 21, 523–536. [Google Scholar] [CrossRef] [PubMed]
- Heino, J.; Alahuhta, J.; Fattorini, S. Macroecology of ground beetles: Species richness, range size and body size show different geographical patterns across a climatically heterogeneous area. J. Biogeogr. 2019, 46, 2548–2557. [Google Scholar] [CrossRef]
- Fattorini, S. Odonate diversity patterns in Italy disclose intricate colonization pathways. Biology 2022, 11, 886. [Google Scholar] [CrossRef] [PubMed]
- Eggleton, P.E.; Williams, P.H.; Gaston, K.J. Explaining global termite diversity: Productivity or history? Biodivers. Conserv. 1994, 3, 318–330. [Google Scholar] [CrossRef]
- Price, P.W.; Fernandes, G.W.; Lara, A.C.F.; Brawn, J.; Barrios, H.; Wright, M.G.; Ribeiro, S.; Rothcliff, N. Global patterns in local number of insect galling species. J. Biogeogr. 1998, 25, 581–591. [Google Scholar] [CrossRef]
- Condamine, F.L.; Sperling, F.A.H.; Wahlberg, N.; Rasplus, J.Y.; Kergoat, G.J. What causes latitudinal gradients in species diversity? Evolutionary processes and ecological constraints on swallowtail biodiversity. Ecol. Lett. 2012, 15, 267–277. [Google Scholar] [CrossRef]
- Economo, E.P.; Narula, N.; Friedman, N.R.; Weiser, M.D.; Guénard, B. Macroecology and macroevolution of the latitudinal diversity gradient in ants. Nat. Commun. 2018, 9, 1778. [Google Scholar] [CrossRef]
- Pinkert, S.; Barve, V.; Guralnick, R.; Jetz, W. Global geographical and latitudinal variation in butterfly species richness captured through a comprehensive country-level occurrence database. Global Ecol. Biogeogr. 2022, 31, 830–839. [Google Scholar] [CrossRef]
- Cardoso, P.; Erwin, T.L.; Borges, P.A.V.; New, T.R. The seven impediments in invertebrate conservation and how to overcome them. Biol. Conserv. 2011, 144, 2647–2655. [Google Scholar] [CrossRef]
- Kouki, J.; Niemelä, P.; Viitasaari, M. Reversed latitudinal gradients in species richness of sawflies (Hymenoptera, Symphyta). Ann. Zool. Fenn. 1994, 31, 83–88. [Google Scholar]
- Boyero, L.; Pearson, R.G.; Dudgeon, D.; Graca, M.A.S.; Gessner, M.O.; Albarino, R.J.; Ferreira, V.; Yule, C.M.; Boulton, A.J.; Arunachalam, M.; et al. Global distribution of a key trophic guild contrasts with common latitudinal diversity patterns. Ecology 2011, 92, 1839–1848. [Google Scholar] [CrossRef]
- Haas, F. Biodiversity of Dermaptera. In Insect Biodiversity and Society; Foottit, R.G., Adler, P.H., Eds.; Wiley–Blackwell: Hoboken, NJ, USA, 2018; pp. 315–334. [Google Scholar]
- Richards, O.W.; Davies, R.G. Imms’ General Textbook of Entomology; Chapman and Hall: London, UK, 1977. [Google Scholar]
- Albouy, V.; Caussanel, C. Dermaptères ou perce–oreilles. In Faune de France; Fédération française des Sociétés de Sciences Naturelles: Paris, France, 1990. [Google Scholar]
- Haas, F. Earwig Research Centre. Available online: http://www.earwigs-online.de (accessed on 23 March 2021).
- Britannica. List of the Total Areas of the World’s Countries, Dependencies, and Territories. Encyclopedia Britannica. 2020. Available online: https://www.britannica.com/topic/list-of-the-total-areas-of-the-worlds-countries-dependencies-and-territories-2130540 (accessed on 23 March 2021).
- South, A. rworldmap: A New R package for Mapping Global Data. R J. 2011, 3, 35–43. [Google Scholar] [CrossRef]
- Ribera, I.; Foster, G.N.; Vogler, A.P. Does habitat use explain large scale species richness patterns of aquatic beetles in Europe? Ecography 2003, 26, 145–152. [Google Scholar] [CrossRef]
- Ulrich, W.; Sachanowicz, K.; Michalak, M. Environmental correlates of species richness of European bats (Mammalia: Chiroptera). Acta Chiropterol. 2007, 9, 347–360. [Google Scholar] [CrossRef]
- Triantis, K.A.; Guilhaumon, F.; Whittaker, R.J. The island species–area relationship: Biology and statistics. J. Biogeogr. 2012, 39, 215–231. [Google Scholar] [CrossRef]
- Matthews, T.J.; Guilhaumon, F.; Triantis, K.A.; Borregaard, M.K.; Whittaker, R.J. On the form of species–area relationships in habitat islands and true islands. Global Ecol. Biogeogr. 2016, 25, 847–858. [Google Scholar] [CrossRef]
- Matthews, T.J.; Rigal, F.; Triantis, K.A.; Whittaker, R.J. A global model of island species–area relationships. Proc. Natl. Acad. Sci. USA 2019, 116, 12337–12342. [Google Scholar] [CrossRef] [PubMed]
- Matthews, T.; Triantis, K.; Whittaker, R. The species–area relationship: Both general and protean? In The Species–Area Relationship: Theory and Application; Matthews, T., Triantis, K., Whittaker, R., Eds.; Cambridge University Press: Cambridge, UK, 2021; pp. 3–19. [Google Scholar]
- Tjørve, E.; Matthews, T.; Whittaker, R. The history of the species–area relationship. In The Species–Area Relationship: Theory and Application; Matthews, T., Triantis, K., Whittaker, R., Eds.; Cambridge University Press: Cambridge, UK, 2021; pp. 20–48. [Google Scholar]
- Tjørve, E.; Tjørve, K.; Šizlingová, E.; Šizling, A. Determinants of the shape of species–area curves. In The Species–Area Relationship: Theory and Application; Matthews, T., Triantis, K., Whittaker, R., Eds.; Cambridge University Press: Cambridge, UK, 2021; pp. 78–106. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. R Core Team: Vienna, Austria, 2020. Available online: https://www.R–project.org (accessed on 31 August 2021).
- Gaston, K. Global patterns in biodiversity. Nature 2000, 405, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, B.A.; Field, R.; Cornell, H.V.; Currie, D.J.; Guégan, J.; Kaufman, D.M.; Kerr, J.T.; Mittelbach, G.G.; Oberdorff, T.; O’Brien, E.M.; et al. Energy, water, and broad-scale geographic patterns of species richness. Ecology 2003, 84, 3105–3117. [Google Scholar] [CrossRef]
- Evans, K.L.; Greenwood, J.J.; Gaston, K.J. Dissecting the species-energy relationship. Proc. Royal Soc. B 2005, 272, 2155–2163. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Brown, J.H.; Tang, Z.; Fang, J. Temperature dependence, spatial scale, and tree species diversity in eastern Asia and North America. Proc. Natl. Acad. Sci. USA 2009, 106, 13388–13392. [Google Scholar] [CrossRef] [PubMed]
- Massante, J.C.; Götzenberger, L.; Takkis, K.; Hallikma, T.; Kaasik, A.; Laanisto, L.; Hutchings, M.J.; Gerhold, P. Contrasting Latitudinal Patterns in Phylogenetic Diversity between Woody and Herbaceous Communities. Sci. Rep. 2019, 9, 6443. [Google Scholar] [CrossRef] [PubMed]
- Lomolino, M.V. The species–area relationship: New challenges for an old pattern. Prog. Phys. Geogr. 2001, 25, 1–21. [Google Scholar] [CrossRef]
- Lomolino, M.V.; Weiser, M.D. Towards a more general species–area relationship: Diversity on all islands, great and small. J. Biogeogr. 2001, 28, 431–445. [Google Scholar] [CrossRef]
- Scheiner, S.M. Six types of species–area curves. Global Ecol. Biogeogr. 2003, 12, 441–447. [Google Scholar] [CrossRef]
- Drakare, S.; Lennon, J.J.; Hillebrand, H. The imprint of the geographical, evolutionary and ecological context on species–area relationships. Ecol. Lett. 2006, 9, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Dapporto, L.; Dennis, R.L.H. Conservation biogeography of large Mediterranean islands. Butterfly impoverishment, conservation priorities and inferences for an ecological island paradigm. Ecography 2009, 32, 169–179. [Google Scholar] [CrossRef]
- Dapporto, L.; Dennis, R.L.H. Skipper butterfly impoverishment on large Mediterranean islands (Lepidoptera Hesperiidae): Deterministic factors and stochastic events. Biodiv. Conserv. 2010, 19, 2637–2649. [Google Scholar] [CrossRef]
- Whittaker, R.J.; Fernandez–Palacios, J.M. Island Biogeography: Ecology, Evolution, and Conservation, 2nd ed.; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Rosenzweig, M.L. Species Diversity in Space and Time; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Ulrich, W.; Buszko, J. Species-area relationships of butterflies in Europe and species richness forecasting. Ecography 2003, 26, 365–373. [Google Scholar] [CrossRef]
- Owens, H.L.; Lewis, D.S.; Dupuis, J.R.; Clamens, A.L.; Sperling, F.A.H.; Kawahara, A.Y.; Guralnick, R.P.; Condamine, F.L. The latitudinal diversity gradient in New World swallowtail butterflies is caused by contrasting patterns of out–of–and into–the–tropics dispersal. Glob. Ecol. Biogeogr. 2017, 26, 1447–1458. [Google Scholar] [CrossRef]
- Wiens, J.J.; Donoghue, M.J. Historical biogeography, ecology and species richness. Trends Ecol Evol. 2004, 19, 639–644. [Google Scholar] [CrossRef]
- Popham, E.; Manly, B. Geographical distribution of the Dermaptera and the continental drift hypothesis. Nature 1969, 222, 981–982. [Google Scholar] [CrossRef]
- Popham, E.J. The geographical distribution of the Dermaptera (Insecta) with reference to continental drift. J. Nat. Hist. 2000, 34, 2007–2027. [Google Scholar] [CrossRef]
- Bey–Bienko, G.J. The Dermapteran Insects, 5th ed.; The Fauna of USSR, New Series; Academy of Sciences USSR: Moscow & Leningrad, Russia, 1936; (in Russian with English summary). [Google Scholar]
- Semenov Tian-Shanskij, A. Les limites et les subdivisions zoogeographiques de la région paléarctique pour les animaux terrestres, basées sur la distribution géographique des insectes Coléoptères. (Avec une carte géographique). Trav. Del’lnstitut Zool. I’Academie Des Sci. I’URSS 1935, 2–3, 397–410. [Google Scholar]
- Dennis, R.; Williams, W.; Shreeve, T. A multivariate approach to the determination of faunal structures among European butterfly species (Lepidoptera: Rhopalocera). Zool. J. Linn. Soc. 1991, 101, 1–49. [Google Scholar] [CrossRef]
- Hewitt, G.M. Post–glacial re–colonization of European biota. Biol. J. Linn. Soc. 1999, 68, 87–112. [Google Scholar] [CrossRef]
- Taberlet, P.; Fumagalli, L.; Wust–Saucy, A.G.; Cosson, J.F. Comparative phylogeography and postglacial colonization routes in Europe. Mol. Ecol. 1998, 7, 453–464. [Google Scholar] [CrossRef]
- Schmitt, T. Molecular biogeography of Europe: Pleistocene cycles and postglacial trends. Front. Zool. 2007, 4, 11. [Google Scholar] [CrossRef]
- Fattorini, S.; Ulrich, W. Spatial distributions of European Tenebrionidae point to multiple postglacial colonization trajectories. Biol. J. Linn. Soc. 2012, 105, 318–329. [Google Scholar] [CrossRef]
- Dapporto, L.; Cini, A.; Vodӑ, R.; Dincӑ, V.; Wiemers, M.; Menchetti, M.; Magini, G.; Talavera, G.; Shreeve, T.; Bonelli, S.; et al. Integrating three comprehensive data sets shows that mitochondrial DNA variation is linked to species traits and paleogeographic events in European butterflies. Mol. Ecol. Resour. 2019, 19, 1623–1636. [Google Scholar] [CrossRef] [PubMed]
- Stuart, O.P.; Binns, M.; Umina, P.A.; Holloway, J.; Severtson, D.; Nash, M.; Heddle, T.; van Helden, M.; Hoffmann, A.A. Morphological and molecular analysis of Australian earwigs (Dermaptera) points to unique species and regional endemism in the Anisolabididae family. Insects 2019, 10, 72. [Google Scholar] [CrossRef]
- Guillet, S.; Josselin, N.; Vancassel, M. Multiple introductions of the Forficula auricularia species complex (Dermaptera: Forficulidae) in Eastern North America. Can. Entomol. 2000, 132, 49–57. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Equation | r2 | F | p |
|---|---|---|---|---|
| World species-area relationship | y = 0.263 + 0.169x | 0.104 | 16.127 | 9.65 × 10−5 |
| World latitudinal gradient | y = 0.207 − 0.0001x2 − 0.003x | 0.221 | 19.565 | 3.31 × 10−8 |
| North Hemisphere species-area relationship | y = 0.433 + 0.123x | 0.058 | 6.874 | 0.0997 |
| North Hemisphere latitudinal gradient | y = 0.320 − 0.011x | 0.180 | 24.239 | 2.87 × 10−6 |
| South Hemisphere species-area relationship | y = 0.144 + 0.240x | 0.257 | 9.066 | 0.0059 |
| South Hemisphere latitudinal gradient | y = 0.379 + 0.024x | 0.353 | 14.189 | 0.0009 |
| Model | Equation | r2 | F | p |
|---|---|---|---|---|
| World species-area relationship | y = −0.409 + 0.281x | 0.166 | 21.916 | 8.18 × 10−6 |
| World latitudinal gradient | y = 0.152 − 0.0001x2 − 0.002x | 0.114 | 6.988 | 0.0014 |
| North Hemisphere species-area relationship | y = −0.097 + 0.216x | 0.109 | 10.925 | 0.0014 |
| North Hemisphere latitudinal gradient | y = 0.253 − 0.008x | 0.106 | 10.607 | 0.0016 |
| South Hemisphere species-area relationship | y = −1.754 + 0.548x | 0.456 | 15.956 | 0.0008 |
| South Hemisphere latitudinal gradient | y = 0.362 + 0.021x | 0.411 | 13.282 | 0.0017 |
| Model | Equation | r2 | F | p |
|---|---|---|---|---|
| World species-area relationship | y = 0.451 + 0.172x | 0.157 | 5.022 | 0.0335 |
| World latitudinal gradient | y = 0.285 − 0.0002x2 − 0.007x | 0.669 | 26.230 | 5.81 × 10−7 |
| North Hemisphere species-area relationship | y = 0.867 + 0.043x | 0.009 | 0.179 | 0.3230 |
| North Hemisphere latitudinal gradient | y = 0.403 − 0.016x | 0.400 | 13.332 | 0.0016 |
| South Hemisphere species-area relationship | y = 0.533 + 0.240x | 0.831 | 24.630 | 0.0040 |
| South Hemisphere latitudinal gradient | y = 0.210 + 0.018x | 0.629 | 8.470 | 0.0334 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fattorini, S. Global Patterns of Earwig Species Richness. Diversity 2022, 14, 890. https://doi.org/10.3390/d14100890
Fattorini S. Global Patterns of Earwig Species Richness. Diversity. 2022; 14(10):890. https://doi.org/10.3390/d14100890
Chicago/Turabian StyleFattorini, Simone. 2022. "Global Patterns of Earwig Species Richness" Diversity 14, no. 10: 890. https://doi.org/10.3390/d14100890
APA StyleFattorini, S. (2022). Global Patterns of Earwig Species Richness. Diversity, 14(10), 890. https://doi.org/10.3390/d14100890