
Understanding the Effects of Climate Change on the Distributional Range of Plateau Fish: A Case Study of Species Endemic to the Hexi River System in the Qinghai–Tibetan Plateau


Abstract
1. Introduction
2. Materials and Methods
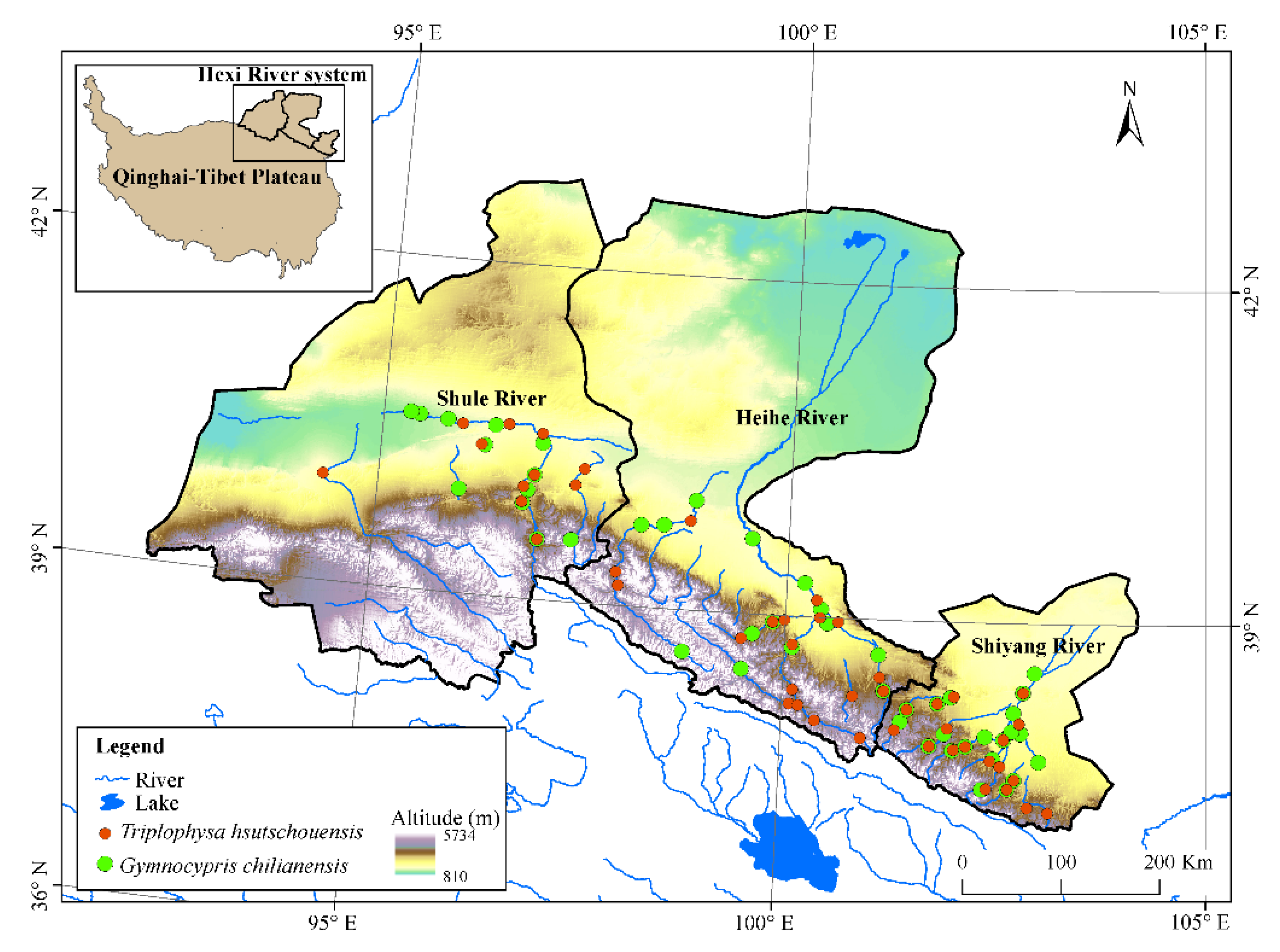
2.1. Occurrence Records
2.2. Current and Future Environmental Variables
2.3. MaxEnt Model Development
3. Results
3.1. Model Evaluation
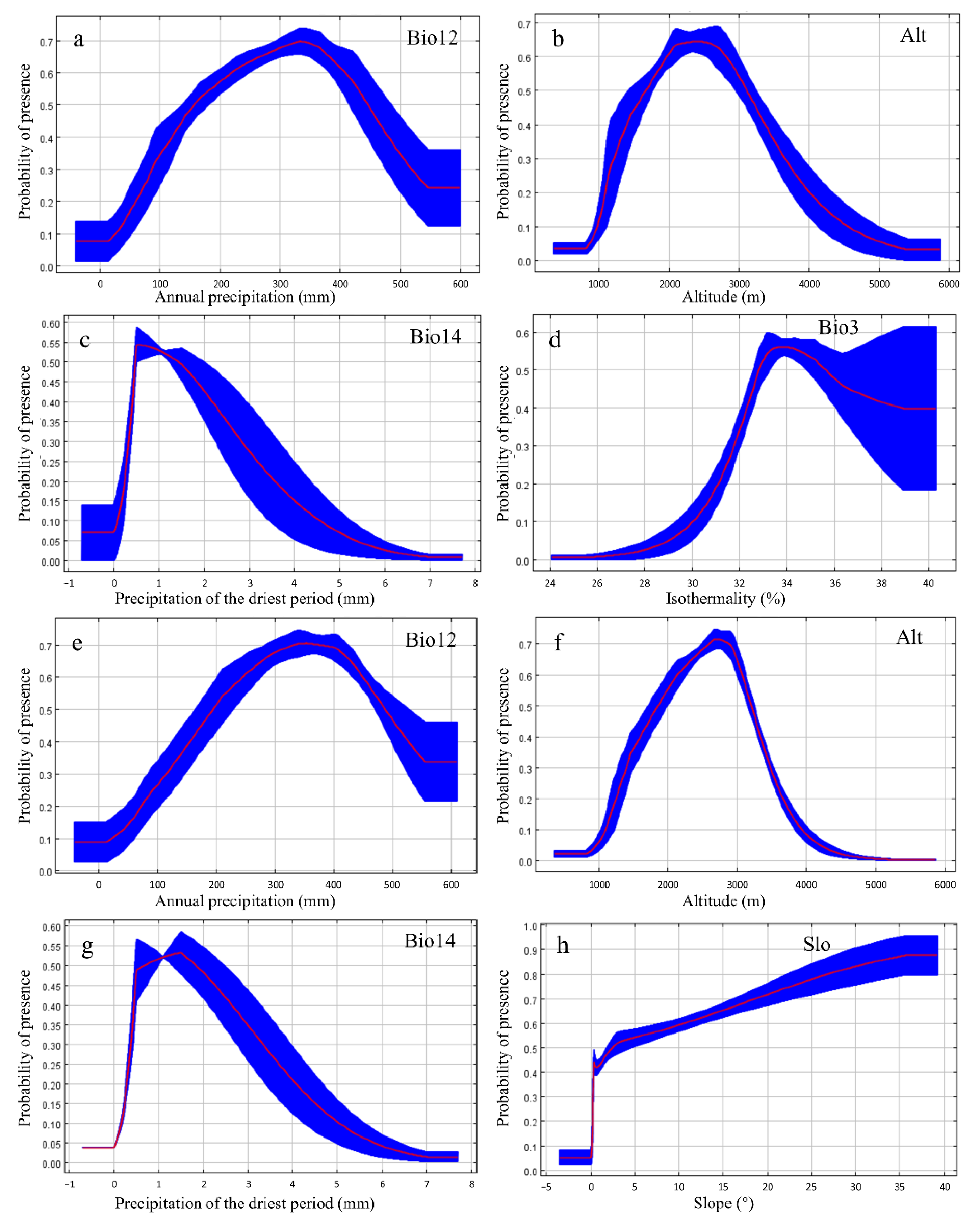
3.2. Important Variables
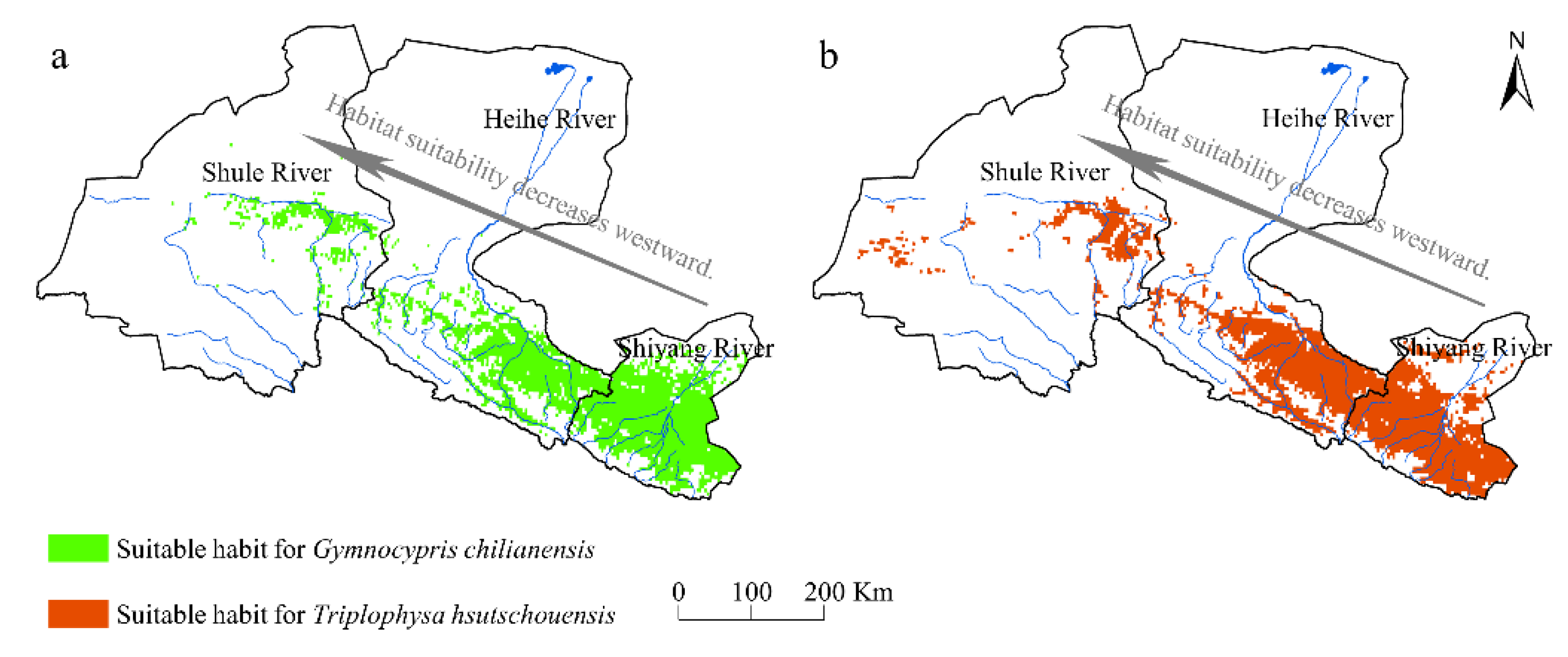
3.3. Current Suitability
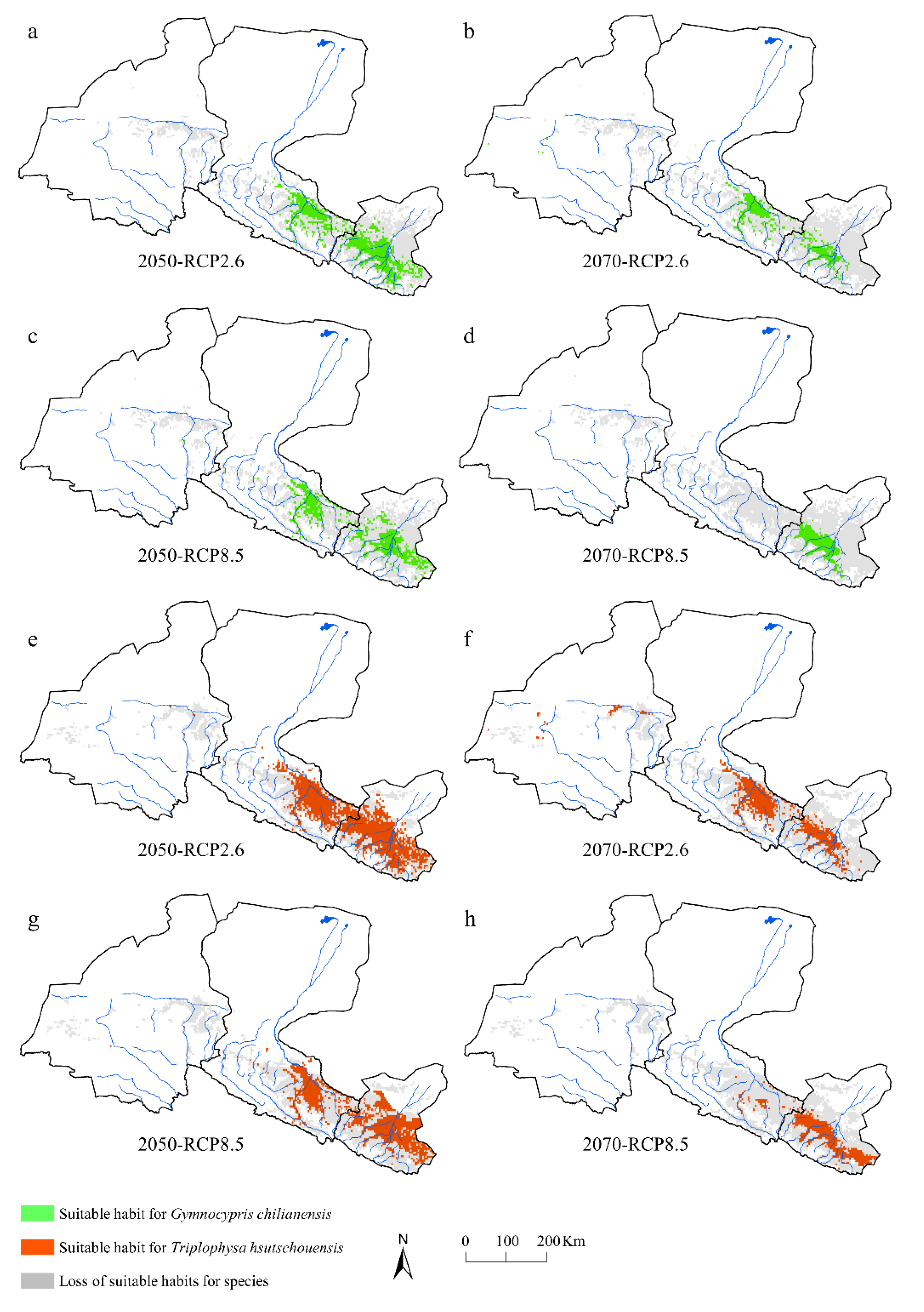
3.4. Predicted Changes in Suitability
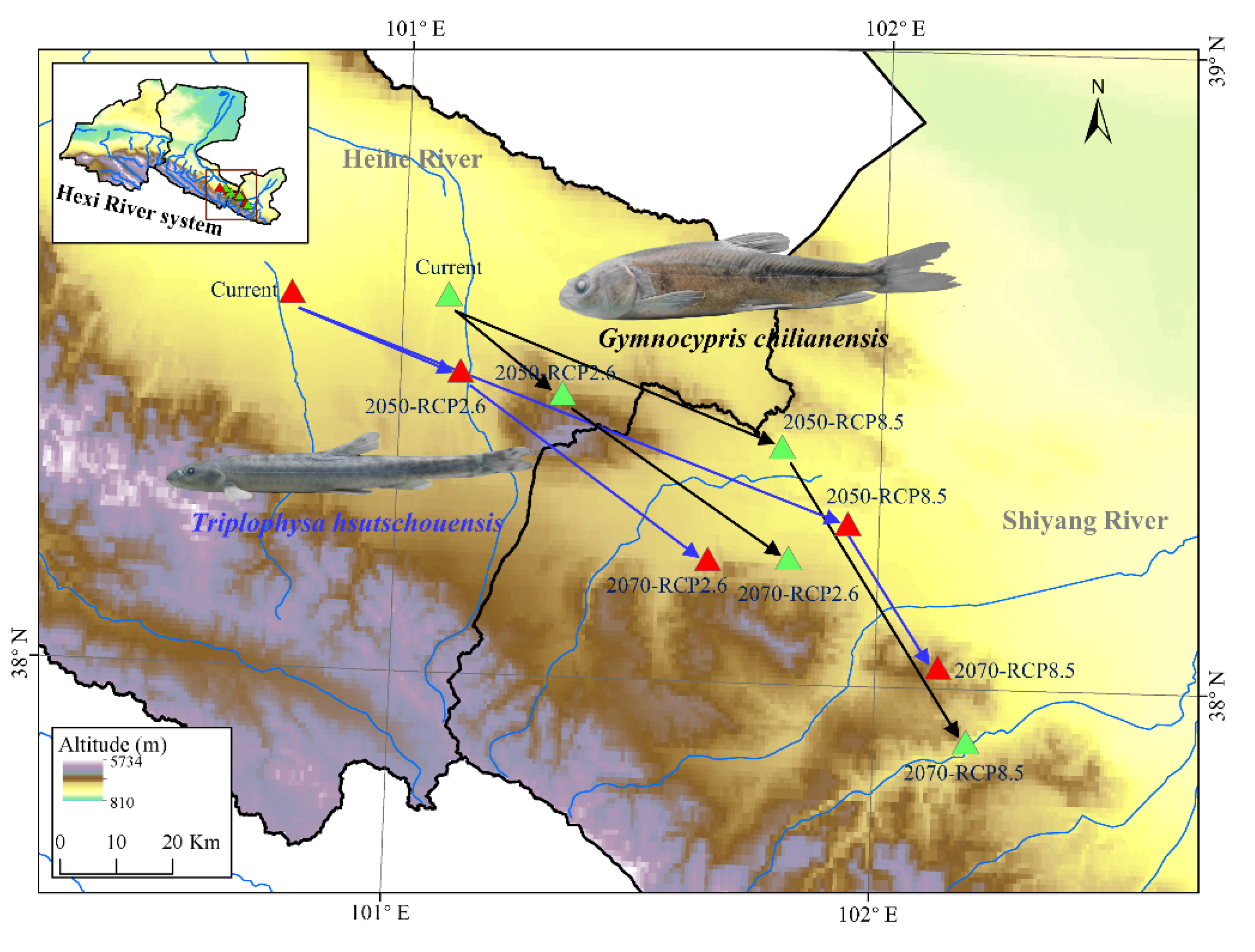
3.5. Distribution Center Change
4. Discussion
4.1. Variable Influences
4.2. Current Distribution Characteristics
4.3. Future Changes
4.4. Conservation
4.5. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mooney, H.; Larigauderie, A.; Cesario, M.; Elmquist, T.; Hoegh-Guldberg, O.; Lavorel, S.; Mace, G.M.; Palmer, M.; Scholes, R.; Yahara, T. Biodiversity, climate change, and ecosystem services. Curr. Opin. Environ. Sustain. 2009, 1, 46–54. [Google Scholar] [CrossRef]
- Barnosky, A.D.; Matzke, N.; Tomiya, S.; Wogan, G.O.U.; Swartz, B.; Quental, T.B.; Marshall, C.; McGuire, J.L.; Lindsey, E.L.; Maguire, K.C.; et al. Has the Earth’s sixth mass extinction already arrived? Nature 2011, 471, 51–57. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Climate Change 2014: Synthesis Report. In Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014; 151p. [Google Scholar]
- Liu, C.; Comte, L.; Xian, W.; Chen, Y.; Olden, J.D. Current and projected future risks of freshwater fish invasions in China. Ecography 2019, 42, 2074–2083. [Google Scholar] [CrossRef]
- Lenoir, J.; Svenning, J.C. Climate-related range shifts–a global multidimensional synthesis and new research directions. Ecography 2015, 38, 15–28. [Google Scholar] [CrossRef]
- Urban, M.C. Accelerating extinction risk from climate change. Science 2015, 348, 571–573. [Google Scholar] [CrossRef]
- Comte, L.; Olden, J.D. Climatic vulnerability of the world’s freshwater and marine fishes. Nat. Clim. Change 2017, 7, 718–722. [Google Scholar] [CrossRef]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2007, 81, 163–182. [Google Scholar] [CrossRef]
- Avaria-Llautureo, J.; Venditti, C.; Rivadeneira, M.M.; Inostroza-Michael, O.; Rivera, R.J.; Hernández, C.E.; Canales-Aguirre, C.B. Historical warming consistently decreased size, dispersal and speciation rate of fish. Nat. Clim. Change 2021, 11, 787–793. [Google Scholar] [CrossRef]
- Knouft, J.H.; Ficklin, D.L. The potential impacts of climate change on biodiversity in flowing freshwater systems. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 111–133. [Google Scholar] [CrossRef]
- Barbarossa, V.; Bosmans, J.; Wanders, N.; King, H.; Bierkens, M.F.P.; Huijbregts, M.A.J.; Schipper, A.M. Threats of global warming to the world’s freshwater fishes. Nat. Commun. 2021, 12, 1701. [Google Scholar] [CrossRef]
- Nogués-Bravo, D.; Araújo, M.B.; Errea, M.P.; Martínez-Rica, J.P. Exposure of global mountain systems to climate warming during the 21st century. Glob. Environ. Change 2007, 17, 420–428. [Google Scholar] [CrossRef]
- Mountain Research Initiative EDW Working Group. Elevation-dependent warming in mountain regions of the world. Nat. Clim. Change 2015, 5, 424–430. [Google Scholar] [CrossRef]
- Klanderud, K.; Totland, Ø. Simulated climate change altered dominance hierarchies and diversity of an alpine biodiversity hotspot. Ecology 2005, 86, 2047–2054. [Google Scholar] [CrossRef]
- Jay, F.; Manel, S.; Alvarez, N.; Durand, E.Y.; Thuiller, W.; Holderegger, R.; Taberlet, P.; François, O. Forecasting changes in population genetic structure of alpine plants in response to global warming. Mol. Ecol. 2012, 21, 2354–2368. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, B. Climatic warming in the Tibetan Plateau during recent decades. Int. J. Climatol. 2000, 20, 1729–1742. [Google Scholar] [CrossRef]
- Liu, J.; Xin, Z.; Huang, Y.; Yu, J. Climate suitability assessment on the Qinghai-Tibet Plateau. Sci. Total Environ. 2022, 816, 151653. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.H.; Ericson, P.G.P.; Lei, F.M.; Li, S.H. Postglacial colonization of the Tibetan plateau inferred from the matrilineal genetic structure of the endemic red-necked snow finch, Pyrgilauda ruficollis. Mol. Ecol. 2005, 14, 1767–1781. [Google Scholar] [CrossRef]
- Zhang, Q.; Chiang, T.Y.; George, M.; Liu, J.Q.; Abbott, R.J. Phylogeography of the Qinghai-Tibetan Plateau endemic Juniperus przewalskii (Cupressaceae) inferred from chloroplast DNA sequence variation. Mol. Ecol. 2005, 14, 3513–3524. [Google Scholar] [CrossRef]
- Qi, D.; Guo, S.; Zhao, X.; Yang, J.; Tang, W. Genetic diversity and historical population structure of Schizopygopsis pylzovi (Teleostei: Cyprinidae) in the Qinghai-Tibetan Plateau. Freshw. Biol. 2007, 52, 1090–1104. [Google Scholar] [CrossRef]
- Zhao, K.; Duan, Z.; Yang, G.; Peng, Z.; He, S.; Chen, Y. Origin of Gymnocypris przewalskii and phylogenetic history of Gymnocypris eckloni (Teleostei: Cyprinidae). Prog. Nat. Sci. 2007, 17, 520–528. [Google Scholar]
- Chen, Z.; Luo, L.; Wang, Z.; He, D.; Zhang, L. Diversity and distribution of fish in the Qilian Mountain Basin. Biodivers. Data J. 2022, 10, e85992. [Google Scholar] [CrossRef]
- Feng, S.W. Evolution of Hexi water system in Gansu. J. Lanzhou Univ. Nat. Sci. 1981, 17, 125–129. [Google Scholar]
- Li, J.J. Discussion of the recent age and the quaternary glaciation of Qilian Mountains. J. Lanzhou Univ. Nat. Sci. 1963, 13, 77–86. [Google Scholar]
- Li, J.J.; Fang, X.M.; Pan, B.T.; Zhao, Z.J.; Song, Y.G. Late Cenozoic intensive uplift of Qinghai-Xizang plateau and its impacts on environments in surrounding area. Quat. Sci. 2001, 21, 381–391. [Google Scholar]
- He, D.; Sui, X.; Sun, H.; Tao, J.; Ding, C.; Chen, Y.; Chen, Y. Diversity, pattern and ecological drivers of freshwater fish in China and adjacent areas. Rev. Fish Biol. Fish. 2020, 30, 387–404. [Google Scholar] [CrossRef]
- Zhao, T.Q. Fish-fauna and zoogeographical division of Hexi-Alashan region, the northwest China. Acta Zool. Sin. 1991, 37, 153–167. [Google Scholar]
- Wang, X.T. Vertebrate Fauna of Gansu; Gansu Science and Technology Press: Lanzhou, China, 1991; pp. 35–36, 102–105. [Google Scholar]
- Wu, Y.F.; Wu, C.Z. The Fishes of the Qinghai-Xizang Plateau; Sichuan Publishing House of Science and Technology: Chengdu, China, 1992; pp. 190–192, 446–448. [Google Scholar]
- Zhao, T.Q. On the taxonomic problem of Gymnocpris eckloni Herzenstein. La Anim. Mondo 1986, 3, 49–55. [Google Scholar]
- Zhao, K.; Duan, Z.; Peng, Z.; Gan, X.; Zhang, R.; He, S.; Zhao, X. Phylogeography of the endemic Gymnocypris chilianensis (Cyprinidae): Sequential westward colonization followed by allopatric evolution in response to cyclical Pleistocene glaciations on the Tibetan Plateau. Mol. Phylogenet. Evol. 2011, 59, 303–310. [Google Scholar] [CrossRef]
- Feng, C.; Zhou, W.; Tang, Y.; Gao, Y.; Chen, J.; Tong, C.; Liu, S.; Wanghe, K.; Zhao, K. Molecular systematics of the Triplophysa robusta (Cobitoidea) complex: Extensive gene flow in a depauperate lineage. Mol. Phylogenet. Evol. 2019, 132, 275–283. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, Y.P.; Yang, Z.Y.; Liu, Z.; Du, Y.Y. DNA barcoding reveals cryptic diversity in the underestimated genus Triplophysa (Cypriniformes: Cobitidae, Nemacheilinae) from the northeastern Qinghai-Tibet Plateau. BMC Evol. Biol. 2020, 20, 151. [Google Scholar] [CrossRef]
- Quan, J.; Zhao, G.; Li, L.; Zhang, J.; Luo, Z.; Kang, Y.; Liu, Z. Phylogeny and genetic diversity reveal the influence of Qinghai-Tibet Plateau uplift on the divergence and distribution of Gymnocypris species. Aquat. Sci. 2021, 83, 6. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Pearson, R.G. Species’ distribution modeling for conservation educators and practitioners. Lessons Conserv. 2010, 3, 54–89. [Google Scholar]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Modell. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Modell. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudik, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Williams, P.H.; Margules, C.R.; Hilbert, D.W. Data requirements and data sources for biodiversity priority area selection. J. Biosci. 2002, 27, 327–338. [Google Scholar] [CrossRef]
- Li, Y.; Qin, T.; Tun, Y.Y.; Chen, X. Fishes of the Irrawaddy River: Diversity and conservation. Aquat. Conserv. 2021, 31, 1945–1955. [Google Scholar] [CrossRef]
- Zhang, C.; Zhu, R.; Sui, X.; Li, X.; Chen, Y. Understanding patterns of taxonomic diversity, functional diversity, and ecological drivers of fish fauna in the Mekong River. Glob. Ecol. Conserv. 2021, 28, e01711. [Google Scholar] [CrossRef]
- WorldClim. Free Climate Data for Ecological Modeling and GIS. Available online: https://www.worldclim.org/ (accessed on 25 May 2022).
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Chen, L.; Frauenfeld, O.W. Surface air temperature changes over the twentieth and twenty-first centuries in China simulated by 20 CMIP5 models. J. Clim. 2014, 27, 3920–3937. [Google Scholar] [CrossRef]
- Hu, W.; Wang, Y.; Dong, P.; Zhang, D.; Yu, W.; Ma, Z.; Chen, G.; Liu, Z.; Du, J.; Chen, B.; et al. Predicting potential mangrove distributions at the global northern distribution margin using an ecological niche model: Determining conservation and reforestation involvement. For. Ecol. Manag. 2020, 478, 118517. [Google Scholar] [CrossRef]
- Meinshausen, M.; Smith, S.J.; Calvin, K.; Daniel, J.S.; Kainuma, M.L.T.; Lamarque, J.F.; Matsumoto, K.; Montzka, S.A.; Raper, S.C.B.; Riahi, K.; et al. The RCP greenhouse gas concentrations and their extensions from 1765 to 2300. Clim. Change 2011, 109, 213–241. [Google Scholar] [CrossRef]
- Sreekumar, E.R.; Nameer, P.O. A MaxEnt modelling approach to understand the climate change effects on the distributional range of White-bellied Sholakili Sholicola albiventris (Blanford, 1868) in the Western Ghats, India. Ecol. Inform. 2022, 70, 101702. [Google Scholar] [CrossRef]
- Fourcade, Y.; Engler, J.O.; Rödder, D.; Secondi, J. Mapping species distributions with MaxEnt using a geographically biased sample of presence data: A performance assessment of methods for correcting sampling bias. PLoS ONE 2014, 9, e97122. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2010, 17, 43–57. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A., Jr. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Liu, C.; White, M.; Newell, G. Selecting thresholds for the prediction of species occurrence with presence-only data. J. Biogeogr. 2013, 40, 778–789. [Google Scholar] [CrossRef]
- Feng, C.; Wu, Y.; Tian, F.; Tong, C.; Tang, Y.; Zhang, R.; Li, G.; Zhao, K. Elevational diversity gradients of Tibetan loaches: The relative roles of ecological and evolutionary processes. Ecol. Evol. 2017, 7, 9970–9977. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Field, R.; Cornell, H.V.; Currie, D.J.; Guégan, J.F.; Kaufman, D.M.; Kerr, J.T.; Mittelbach, G.G.; Oberdorff, T.; O’Brien, E.M.; et al. Energy, water, and broad-scale geographic patterns of species richness. Ecology 2003, 84, 3105–3117. [Google Scholar] [CrossRef]
- Froese, R.; Pauly, D.; FishBase. World Wide Web Electronic Publication. Available online: http://www.fishbase.org,version (accessed on 2 August 2022).
- Hu, Y.; Fan, H.; Chen, Y.; Chang, J.; Zhan, X.; Wu, H.; Zhang, B.; Wang, M.; Zhang, W.; Yang, L.; et al. Spatial patterns and conservation of genetic and phylogenetic diversity of wildlife in China. Sci. Adv. 2021, 7, eabd5725. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.; Huang, X. Mekong fishes: Biogeography, migration, resources, threats, and conservation. Rev. Fish. Sci. Aquac. 2022, 30, 170–194. [Google Scholar] [CrossRef]
- Bruton, M.N. The effects of suspensoids on fish. Hydrobiologia 1985, 125, 221–241. [Google Scholar] [CrossRef]
- Hauer, C.; Leitner, P.; Unfer, G.; Pulg, U.; Habersack, H.; Graf, W. The role of sediment and sediment dynamics in the aquatic environment. In Riverine Ecosystem Management; Schmutz, S., Sendzimir, J., Eds.; Springer: Cham, Switzerland, 2018; Volume 8, pp. 151–169. [Google Scholar]
- O’Brien, E.M. Climatic gradients in woody plant species richness: Towards an explanation based on an analysis of Southern Africa’s woody flora. J. Biogeogr. 1993, 20, 181–198. [Google Scholar] [CrossRef]
- Colwell, R.K.; Hurtt, G.C. Nonbiological gradients in species richness and a spurious Rapoport effect. Am. Nat. 1944, 144, 570–595. [Google Scholar] [CrossRef]
- Colwell, R.K.; Lees, D.C. The mid-domain effect: Geometric constraints on the geography of species richness. Trends Ecol. Evol. 2000, 15, 70–76. [Google Scholar] [CrossRef]
- Li, J.; He, Q.; Hua, X.; Zhou, J.; Xu, H.; Chen, J.; Fu, C. Climate and history explain the species richness peak at mid-elevation for Schizothorax fishes (Cypriniformes: Cyprinidae) distributed in the Tibetan Plateau and its adjacent regions. Glob. Ecol. Biogeogr. 2009, 18, 264–272. [Google Scholar] [CrossRef]
- Stephens, P.R.; Wiens, J.J. Explaining species richness from continents to communities: The time-for-speciation effect in emydid turtles. Am. Nat. 2003, 161, 112–128. [Google Scholar] [CrossRef]
- Parmesan, C. Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Long, D.; Scanlon, B.R.; Mann, M.E.; Li, X.; Tian, F.; Sun, Z.; Wang, G. Climate change threatens terrestrial water storage over the Tibetan Plateau. Nat. Clim. Change 2022, 12, 801–807. [Google Scholar] [CrossRef]
- Guglielmi, G. Climate change is turning more of Central Asia into desert. Nature 2022. [Google Scholar] [CrossRef] [PubMed]
- Barbarossa, V.; Schmitt, R.J.P.; Huijbregts, M.A.J.; Zarfl, C.; King, H.; Schipper, A.M. Impacts of current and future large dams on the geographic range connectivity of freshwater fish worldwide. Proc. Natl. Acad. Sci. USA 2020, 117, 3648–3655. [Google Scholar] [CrossRef]
- Rangel, T.F.; Edwards, N.R.; Holden, P.B.; Diniz-Filho, J.A.F.; Gosling, W.D.; Coelho, M.T.P.; Cassemiro, F.A.; Rahbek, C.; Colwell, R.K. Modeling the ecology and evolution of biodiversity: Biogeographical cradles, museums, and graves. Science 2018, 361, 244–257. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Y. Identification and Assessment of Ecological Networks of Ungulates in Qilian Mountains under Climate Change. Master’s Thesis, Lanzhou University, Lanzhou, China, 2022. [Google Scholar]
- Rahel, F.J. Biogeographic barriers, connectivity and homogenization of freshwater faunas: It’s a small world after all. Freshw. Biol. 2007, 52, 696–710. [Google Scholar] [CrossRef]
- Alabdulhafith, B.; Binothman, A.; Alwahiby, A.; Haig, S.M.; Prommer, M.; Leonardi, G. Predicting the potential distribution of a near-extinct avian predator on the Arabian Peninsula: Implications for its conservation management. Environ. Monit. Assess. 2022, 194, 535. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, L.; Zhou, K.; Zhang, Y.; Song, Z.; He, S. Evidence for adaptation to the Tibetan Plateau inferred from Tibetan loach transcriptomes. Genome Biol. Evol. 2015, 7, 2970–2982. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, Y.; Zhang, Z.; He, S. Comprehensive transcriptome analysis reveals accelerated genic evolution in a Tibet fish, Gymnodiptychus pachycheilus. Genome Biol. Evol. 2015, 7, 251–261. [Google Scholar] [CrossRef]
- Lei, Y.; Yang, L.; Zhou, Y.; Wang, C.; Lv, W.; Li, L.; He, S. Hb adaptation to hypoxia in high-altitude fishes: Fresh evidence from schizothoracinae fishes in the Qinghai-Tibetan Plateau. Int. J. Biol. Macromol. 2021, 185, 471–484. [Google Scholar] [CrossRef] [PubMed]
- Toro, M.A.; Caballero, A. Characterization and conservation of genetic diversity in subdivided populations. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2005, 360, 1367–1378. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Sui, X.; Chen, Y.; He, D. Climate change and spatial distribution shaped the life-history traits of schizothoracine fishes on the Tibetan Plateau and its adjacent areas. Glob. Ecol. Conserv. 2020, 22, e01041. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Brooks, T.M.; Mittermeier, R.A.; da Fonseca, G.A.B.; Gerlach, J.; Hoffmann, M.; Lamoreux, J.F.; Mittermeier, C.G.; Pilgrim, J.D.; Rodrigues, A.S.L. Global biodiversity conservation priorities. Science 2006, 313, 58–61. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, P.; Wu, C.; Li, X.; An, R.; Xie, K. Reservoir ecological operation by quantifying outflow disturbance to aquatic community dynamics. Environ. Res. Lett. 2021, 16, 074005. [Google Scholar] [CrossRef]
- Zhou, B.; Wen, Q.H.; Xu, Y.; Song, L.; Zhang, X. Projected changes in temperature and precipitation extremes in China by the CMIP5 multimodel ensembles. J. Clim. 2017, 27, 6591–6611. [Google Scholar] [CrossRef]
- Giachetta, E.; Willett, S. A global dataset of river network geometry. Sci. Data 2018, 5, 180127. [Google Scholar] [CrossRef]
- Keil, P.; Chase, J.M. Global patterns and drivers of tree diversity integrated across a continuum of spatial grains. Nat. Ecol. Evol. 2019, 3, 390–399. [Google Scholar] [CrossRef]
- Carvajal-Quintero, J.; Villalobos, F.; Oberdorff, T.; Grenouillet, G.; Brosse, S.; Hugueny, B.; Jézéquel, C.; Tedesco, P.A. Drainage network position and historical connectivity explain global patterns in freshwater fishes’ range size. Proc. Natl. Acad. Sci. USA 2019, 116, 13434–13439. [Google Scholar] [CrossRef]
- Yue, T.; Zhao, N.; Fan, Z.; Li, J.; Chen, F.; Lu, Y.; Wang, C.; Xu, B.; Wilson, J. CMIP5 downscaling and its uncertainty in China. Glob. Planet. Change 2016, 146, 30–37. [Google Scholar] [CrossRef]
- Hewitson, B.C.; Crane, R.G. Climate downscaling: Techniques and application. Clim. Res. 1996, 7, 85–95. [Google Scholar] [CrossRef]
- Chen, H.P.; Sun, J.Q.; Li, H.X. Future changes in precipitation extremes over China using the NEX-GDDP high-resolution daily downscaled data-set. Atmos. Oceanic Sci. Lett. 2017, 10, 403–410. [Google Scholar] [CrossRef]
- Räisänen, J. How reliable are climate models? Tellus A 2007, 59, 2–29. [Google Scholar] [CrossRef]
- Raftery, A.E.; Gneiting, T.; Balabdaoui, F.; Polakowski, M. Using Bayesian model averaging to calibrate forecast ensembles. Mon. Weather Rev. 2005, 133, 1155–1174. [Google Scholar] [CrossRef]
- Liu, L.L.; Liu, Z.F.; Ren, X.Y.; Fischer, T.; Xu, Y. Hydrological impacts of climate change in the Yellow River basin for the 21st century using hydrological model and statistical downscaling model. Quat. Int. 2011, 244, 211–220. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor Sets | Code | Variable | Unit | Description |
|---|---|---|---|---|
| Climate † | Bio1 | Annual mean temperature | °C | The average temperature for each month |
| Bio2 | Mean diurnal range | °C | Measure of temperature change over the course of the year using monthly maximum temperatures and monthly minimum temperatures | |
| Bio3 | Isothermality | % | Derived by calculating the ratio of the mean diurnal range (Bio 2) to the annual temperature range (Bio 7, discussed below) and then multiplying by 100 | |
| Bio4 | Temperature seasonality (Standard deviation × 100) | % | The amount of temperature variation over a cause of the year, based on the standard deviation (variation) of monthly temperature averages | |
| Bio5 | Max. temperature of the warmest month | °C | The maximum monthly temperature occurrence over a given year (time series) or averaged span of years (normal) | |
| Bio6 | Min. temperature of the coldest month | °C | The minimum monthly temperature occurrence over a given year (time series) or averaged span of years (normal) | |
| Bio7 | Temperature of annual range | °C | A measure of temperature variation over a given period. (Bio7 = Bio5–Bio6) | |
| Bio8 | Mean temperature of the wettest quarter | °C | Mean temperatures that prevail during the wettest season | |
| Bio9 | Mean temperature of the driest quarter | °C | Quarterly index approximates mean temperatures that prevail during the driest quarter | |
| Bio10 | Mean temperature of the warmest quarter | °C | Quarterly index approximates mean temperatures that prevail during the warmest quarter | |
| Bio11 | Mean temperature of the coldest quarter | °C | Quarterly index approximates mean temperatures that prevail during the coldest quarter | |
| Bio12 | Annual precipitation | mm | Sum of all total monthly precipitation values | |
| Bio13 | Precipitation of the wettest month | mm | The total precipitation that prevails during the wettest month | |
| Bio14 | Precipitation of the driest month | mm | The total precipitation that prevails during the driest month | |
| Bio15 | Precipitation seasonality (Coefficient of variation) | % | Measure of the variation in monthly precipitation totals over the course of the year | |
| Bio16 | Precipitation of the wettest quarter | mm | Total precipitation that prevails during the wettest quarter | |
| Bio17 | Precipitation of the driest quarter | mm | Total precipitation that prevails during the driest quarter | |
| Bio18 | Precipitation of the warmest quarter | mm | Total precipitation that prevails during the warmest quarter | |
| Bio19 | Precipitation of the coldest quarter | mm | Total precipitation that prevails during the coldest quarter | |
| Topography ‡ | Alt | Altitude | m | The potential for habitat diversification |
| Slo | Slope | ° | Computed based on the Alt data using the Surface function of Spatial Analyst Tools in ArcGIS 10.7 | |
| Food resources § | Npp | Net primary productivity | gC/m2 | Net amount of solar energy converted to plant organic matter through photosynthesis |
| Human activities ¶ | Hfp | Human footprint | Created from nine data layers covering human population pressure (population density), human land use and infrastructure (built-up areas, nighttime lights, land use/land cover), and human access (coastlines, roads, railroads, navigable rivers) |
| Model | Gymnocypris chilianensis | Triplophysa hsutschouensis |
|---|---|---|
| Current (1970–2000) | 0.938 | 0.936 |
| Future (2050)-RCP2.6 | 0.926 | 0.942 |
| Future (2070)-RCP2.6 | 0.929 | 0.941 |
| Future (2050)-RCP8.5 | 0.925 | 0.940 |
| Future (2070)-RCP8.5 | 0.930 | 0.942 |
| Code | G. chilianensis | T. hsutschouensis |
|---|---|---|
| Bio12 | 35.8 | 38.7 |
| Alt | 23.7 | 24.4 |
| Bio14 | 20.0 | 19.1 |
| Bio3 | 10.6 | 6.3 |
| Slo | 6.1 | 8.0 |
| Bio15 | 2.2 | 1.7 |
| Bio2 | 0.9 | 1.2 |
| Bio9 | 0.7 | 0.6 |
| Species | Future Climate Scenario | Suitable Area (km2) | Loss of Suitable Area (km2) | Loss of Suitable Habitat (%) |
|---|---|---|---|---|
| G. chilianensis | 2050-RCP2.6 | 14,286.5 | 32,293.8 | 69.3 |
| 2070-RCP2.6 | 9559.4 | 37,020.9 | 79.5 | |
| 2050-RCP8.5 | 11,224.5 | 35,355.8 | 75.9 | |
| 2070-RCP8.5 | 5355.8 | 41,224.6 | 88.5 | |
| T. hsutschouensis | 2050-RCP2.6 | 26,784.3 | 21,967.5 | 45.1 |
| 2070-RCP2.6 | 15,026.5 | 33,725.3 | 69.2 | |
| 2050-RCP8.5 | 18,922.4 | 29,829.4 | 61.2 | |
| 2070-RCP8.5 | 8679.6 | 40,072.2 | 82.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Z.; Chen, L.; Wang, Z.; He, D. Understanding the Effects of Climate Change on the Distributional Range of Plateau Fish: A Case Study of Species Endemic to the Hexi River System in the Qinghai–Tibetan Plateau. Diversity 2022, 14, 877. https://doi.org/10.3390/d14100877
Chen Z, Chen L, Wang Z, He D. Understanding the Effects of Climate Change on the Distributional Range of Plateau Fish: A Case Study of Species Endemic to the Hexi River System in the Qinghai–Tibetan Plateau. Diversity. 2022; 14(10):877. https://doi.org/10.3390/d14100877
Chicago/Turabian StyleChen, Zhaosong, Liuyang Chen, Ziwang Wang, and Dekui He. 2022. "Understanding the Effects of Climate Change on the Distributional Range of Plateau Fish: A Case Study of Species Endemic to the Hexi River System in the Qinghai–Tibetan Plateau" Diversity 14, no. 10: 877. https://doi.org/10.3390/d14100877
APA StyleChen, Z., Chen, L., Wang, Z., & He, D. (2022). Understanding the Effects of Climate Change on the Distributional Range of Plateau Fish: A Case Study of Species Endemic to the Hexi River System in the Qinghai–Tibetan Plateau. Diversity, 14(10), 877. https://doi.org/10.3390/d14100877