
The Long and Winding Road of Coral Reef Recovery in the Anthropocene: A Case Study from Puerto Rico



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Sampling Design
2.3. Statistical Analyses
3. Results
3.1. Coral Reef Benthic Community Analysis
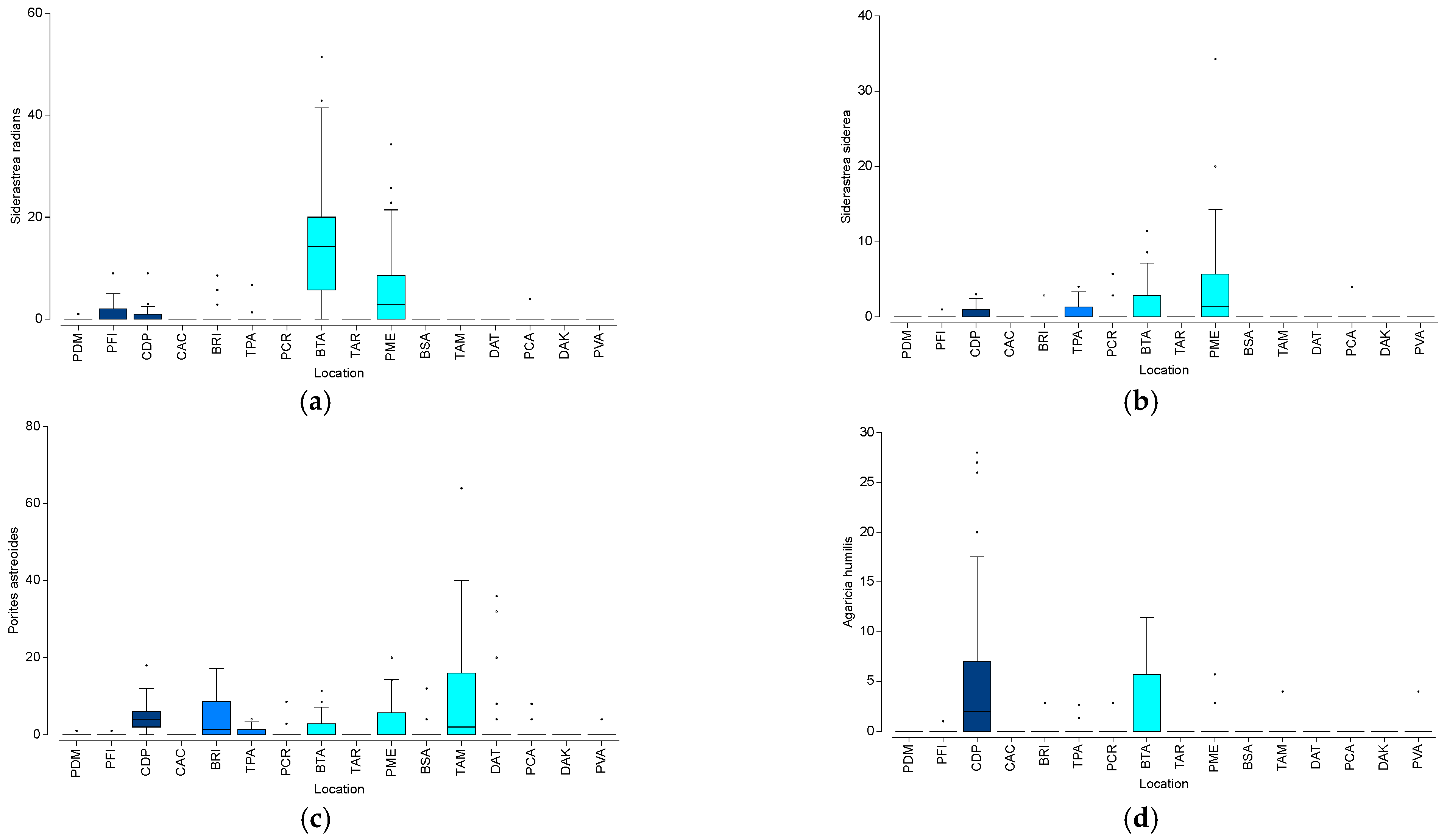
3.2. Coral Recruit Analysis
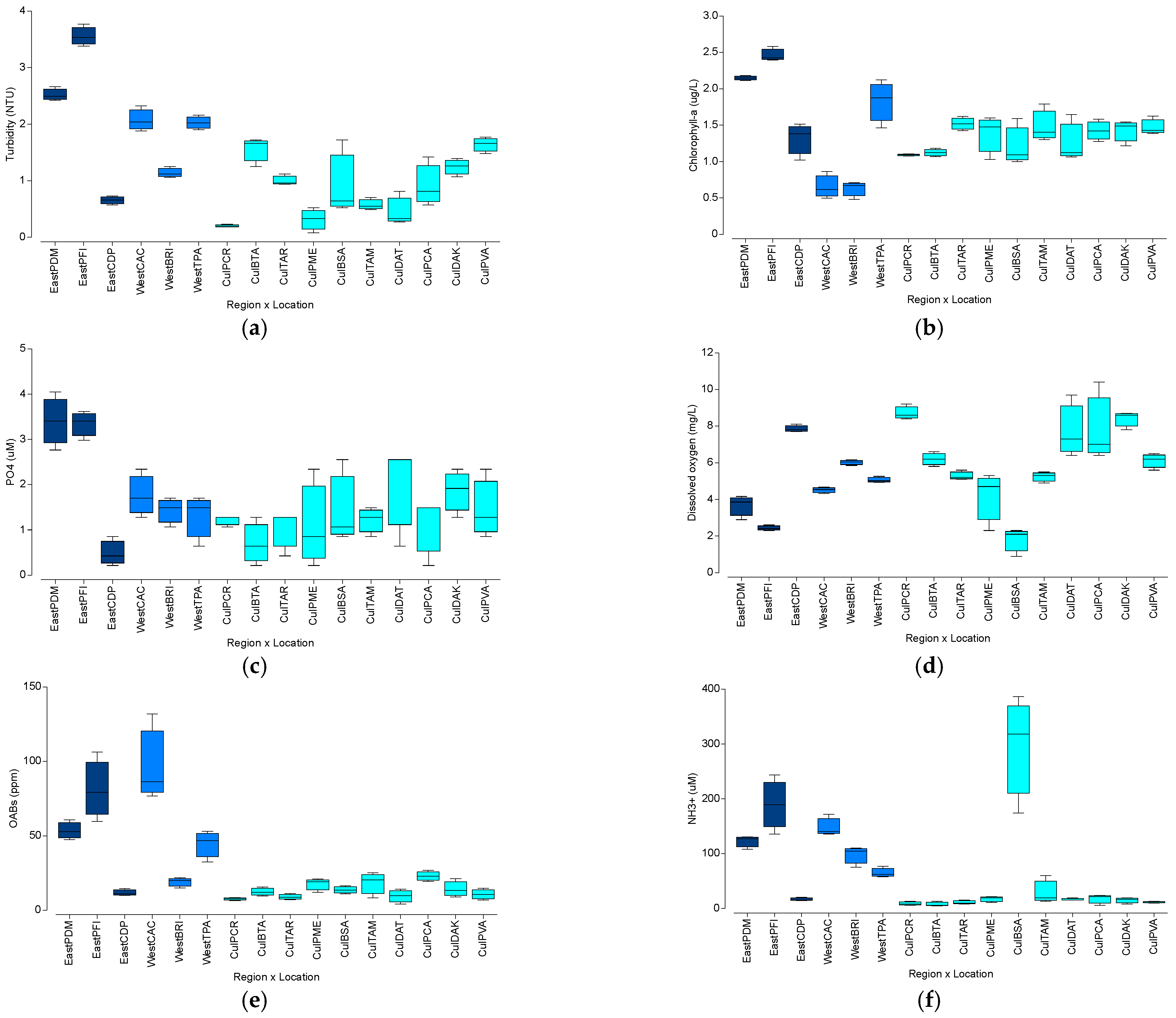
3.3. Water Quality Analysis
3.4. Relationship between Water Quality and Coral Recruit Community Structure
3.5. Relationship between Coral Recruit and Benthic Communities
3.6. Relationship between Water Quality and the Benthic Community Structure
4. Discussion
4.1. The Role of Poor Water Quality
4.2. The Role of Strong Hurricanes
4.3. Potential Impacts on Coral Larval Dynamics
4.4. Long-Term Consequences for Coral Reef Ecological Functions and Services
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cesar, H.S.J. Coral reefs: Their functions, threats and economic value. In Collected Essays on the Economics of Coral Reefs; Cesar, H.S.J., Ed.; CORDIO: Växjö, Sweden, 2007; pp. 14–39. [Google Scholar]
- Leeworthy, V.R.; Schwarzmann, D.; Hughes, S.; Vaughn, J.; Dato, C.; Padilla, G. Economic Contribution of Reef Using Visitor Spending to the Puerto Rican Economy; Office of National Marine Sanctuaries, National Oceanic and Atmospheric Administration: Silver Spring, MD, USA, 2018; pp. 1–23. [Google Scholar]
- Storlazzi, C.D.; Reguero, B.G.; Cole, A.D.; Lowe, E.; Shope, J.B.; Gibbs, A.E.; Nickel, B.A.; McCall, R.T.; van Dongeren, A.R.; Beck, M.W. Rigorously valuing the role of US coral reefs in coastal hazard risk reduction. In Open-File Report-US Geological Survey; U.S. Geological Survey: Reston, VA, USA, 2019. [Google Scholar]
- Miller, J.; Muller, E.; Rogers, C.; Waara, R.; Atkinson, A.; Whelan, K.R.T.; Patterson, M.; Witcher, B. Coral disease following massive bleaching in 2005 causes 60% decline in coral cover on reefs in the US Virgin Islands. Coral Reefs 2009, 28, 925–937. [Google Scholar] [CrossRef]
- Edmunds, P.J. Decadal-scale changes in the community structure of coral reefs of St. John, US Virgin Islands. Mar. Ecol. Prog. Ser. 2013, 489, 107–123. [Google Scholar] [CrossRef]
- Hughes, T.P.; Barnes, M.L.; Bellwood, D.R.; Cinner, J.E.; Cumming, G.S.; Jackson, J.B.; Kleypas, J.; Van De Leemput, I.A.; Lough, J.M.; Morrison, T.H.; et al. Coral reefs in the Anthropocene. Nature 2017, 546, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Kerry, J.T.; Simpson, T. Large-scale bleaching of corals on the Great Barrier Reef. Ecology 2018, 99, 501. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Kerry, J.T.; Baird, A.H.; Connolly, S.R.; Dietzel, A.; Eakin, C.M.; Heron, S.F.; Hoey, A.S.; Hoogenboom, M.O.; Liu, G.; et al. Global warming transforms coral reef assemblages. Nature 2018, 556, 492–496. [Google Scholar] [CrossRef]
- National Oceanic and Atmospheric Administration. NOAA Coral Reef Ecosystems. 2019. Available online: https://www.noaa.gov/education/resource-collections/marine-life-education-resources/coral-reef-ecosystems (accessed on 1 May 2021).
- Ferrario, F.; Beck, M.W.; Storlazzi, C.D.; Micheli, F.; Shepard, C.C.; Airoldi, L. The effectiveness of coral reefs for coastal hazard risk reduction and adaptation. Nat. Commun. 2014, 5, 3794. [Google Scholar] [CrossRef]
- ICRI What Are Corals? International Coral Reef Initiative. 2015. Available online: https://www.icriforum.org/about-coral-reefs/what-are-corals (accessed on 1 May 2021).
- Scheffer, M.; Barrett, S.; Carpenter, S.R.; Folke, C.; Green, A.J.; Holmgren, M.; Hughes, T.P.; Kosten, S.; Van de Leemput, I.A.; Nepstad, D.C.; et al. Creating a safe operating space for iconic ecosystems. Science 2015, 347, 1317–1319. [Google Scholar] [CrossRef]
- Hughes, T.P.; Kerry, J.T.; Baird, A.H.; Connolly, S.R.; Chase, T.J.; Dietzel, A.; Hill, T.; Hoey, A.S.; Hoogenboom, M.O.; Jacobson, M.; et al. Global warming impairs stock–recruitment dynamics of corals. Nature 2019, 568, 387–390. [Google Scholar] [CrossRef]
- Hurlbut, C.J. Community recruitment: Settlement and juvenile survival of seven co-occurring species of sessile marine invertebrates. Mar. Biol. 1991, 109, 507–515. [Google Scholar] [CrossRef]
- Heery, E.C.; Hoeksema, B.W.; Browne, N.K.; Reimer, J.D.; Ang, P.O.; Huang, D.; Friess, D.A.; Chou, L.M.; Loke, L.H.; Saksena-Taylor, P.; et al. Urban coral reefs: Degradation and resilience of hard coral assemblages in coastal cities of East and Southeast Asia. Mar. Pollut. Bull. 2018, 135, 654–681. [Google Scholar] [CrossRef]
- Burt, J.A.; Bartholomew, A. Towards more sustainable coastal development in the Arabian Gulf: Opportunities for ecological engineering in an urbanized seascape. Mar. Pollut. Bull. 2019, 142, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Burt, J.A.; Camp, E.F.; Enochs, I.C.; Johansen, J.L.; Morgan, K.M.; Riegl, B.; Hoey, A.S. Insights from extreme coral reefs in a changing world. Coral Reefs 2020, 39, 495–507. [Google Scholar] [CrossRef]
- Todd, P.A.; Heery, E.C.; Loke, L.H.; Thurstan, R.H.; Kotze, D.J.; Swan, C. Towards an urban marine ecology: Characterizing the drivers, patterns and processes of marine ecosystems in coastal cities. Oikos 2019, 128, 1215–1242. [Google Scholar] [CrossRef]
- Mumby, P.; Steneck, R. The resilience of coral reefs and its implications for reef management. In Coral Reefs: An Ecosystem in Transition; Dubinsky, Z., Sambler, N., Eds.; Springer Science: New York, NY, USA, 2011; pp. 509–519. [Google Scholar]
- Birrell, C.L.; McCook, L.J.; Willis, B.L. Effects of algal turfs and sediment on coral Settlement. Mar. Pollut. Bull. 2005, 51, 408–414. [Google Scholar] [CrossRef]
- Hughes, T.P.; Tanner, J.E. Recruitment failure, life histories, and long-term decline of Caribbean corals. Ecology 2000, 81, 2250–2263. [Google Scholar] [CrossRef]
- Connell, J.H. Disturbance and recovery of coral assemblages. Coral Reefs 1997, 16, 101–113. [Google Scholar] [CrossRef]
- Rogers, C.S. Responses of coral reefs and reef organisms to sedimentation. Mar. Ecol. Prog. Ser. 1990, 62, 185–202. [Google Scholar] [CrossRef]
- Hughes, T.P. Catastrophes, phase shifts, and large-scale degradation of a Caribbean coral reef. Science 1994, 265, 1547–1551. [Google Scholar] [CrossRef]
- Delgado, E.A.H.; Rosado-Matías, B.J. Long-lasting impacts of beach renourishment on nearshore urban coral reefs: A glimpse on future impacts of shoreline erosion, sea level rise and climate change. Ann. Mar. Biol. Res. 2017, 4, 1021. [Google Scholar]
- Erftemeijer, P.L.; Riegl, B.; Hoeksema, B.W.; Todd, P.A. Environmental impacts of dredging and other sediment disturbances on corals: A review. Mar. Pollut. Bull. 2012, 64, 1737–1765. [Google Scholar] [CrossRef]
- Cloern, J.E. Our evolving conceptual model of the coastal eutrophication problem. Mar. Ecol. Prog. Ser. 2001, 210, 223–253. [Google Scholar] [CrossRef]
- Roth, F.; Saalmann, F.; Thomson, T.; Coker, D.J.; Villalobos, R.; Jones, B.H.; Wild, C.; Carvalho, S. Coral reef degradation affects the potential for reef recovery after disturbance. Mar. Env. Res. 2018, 142, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Lapointe, B.E. Chasing nutrients and algal blooms in Gulf and Caribbean waters: A personal story. Gulf Caribb. Res. 2019, 30, xvi-xxx. [Google Scholar] [CrossRef]
- Fabricius, K.E. Effects of terrestrial runoff on the ecology of corals and coral reefs: Review and synthesis. Mar. Poll. Bull. 2005, 50, 125–146. [Google Scholar] [CrossRef]
- Nelson, H.R.; Altieri, A.H. Oxygen: The universal currency on coral reefs. Coral Reefs 2019, 38, 177–198. [Google Scholar] [CrossRef]
- Pawlik, J.R.; Burkepile, D.E.; Thurber, R.V. A vicious circle? Altered carbon and nutrient cycling may explain the low resilience of Caribbean coral reefs. BioScience 2016, 66, 470–476. [Google Scholar] [CrossRef]
- Kleypas, J.A.; McManus, J.W.; Meñez, L.A.B. Environmental limits to coral reef development: Where do we draw the line? Am. Zool. 1999, 39, 146–159. [Google Scholar] [CrossRef]
- Guinotte, J.M.; Buddemeier, R.W.; Kleypas, J.A. Future coral reef habitat marginality: Temporal and spatial effects of climate change in the Pacific basin. Coral Reefs 2003, 22, 551–558. [Google Scholar] [CrossRef]
- De Oliveira Soares, M. Marginal reef paradox: A possible refuge from environmental changes? Ocean. Coast. Mgmt. 2020, 185, 105063. [Google Scholar] [CrossRef]
- Skirving, W.J.; Heron, S.F.; Marsh, B.L.; Liu, G.; De La Cour, J.L.; Geiger, E.F.; Eakin, C.M. The relentless march of mass coral bleaching: A global perspective of changing heat stress. Coral Reefs 2019, 38, 547–557. [Google Scholar] [CrossRef]
- UNEP Projections of Future Coral Bleaching Conditions Using IPCC CMIP6 Models: Climate Policy Implications, Management Applica-tions, and Regional Seas Summaries; United Nations Environment Programme: Nairobi, Kenya, 2020; pp. 1–102.
- UNEP Coral Bleaching Futures—Downscaled Projections of Bleaching Conditions for the World’s Coral Reefs, Implications of Climate Policy and Management Responses; United Nations Environment Programme: Nairobi, Kenya, 2017; pp. 1–69.
- Hernández-Pacheco, R.; Hernández-Delgado, E.A.; Sabat, A.M. Demographics of bleaching in the Caribbean reef-building coral Montastraea annularis. Ecosphere 2011, 2, 1–13. [Google Scholar] [CrossRef]
- Levas, S.; Schoepf, V.; Warner, M.E.; Aschaffenburg, M.; Baumann, J.; Grottoli, A.G. Long-term recovery of Caribbean corals from bleaching. J. Exp. Mar. Biol. Ecol. 2018, 506, 124–134. [Google Scholar] [CrossRef]
- Edmunds, P.J. Recruitment hotspots and bottlenecks mediate the distribution of corals on a Caribbean reef. Biol. Lett. 2021, 17, 20210149. [Google Scholar] [CrossRef] [PubMed]
- Edmunds, P.J.; Didden, C.; Frank, K. Over three decades, a classic winner starts to lose in a Caribbean coral community. Ecosphere 2021, 12, e03517. [Google Scholar] [CrossRef]
- Weil, E.; Urreiztieta, I.; Garzón-Ferreira, J. Geographic variability in the incidence of coral and octocoral diseases in the wider Caribbean. Proc. 9th Int. Coral Reef Symp. 2000, 2, 1231–1237. [Google Scholar]
- Cróquer, A.; Weil, E. Changes in Caribbean coral disease prevalence after the 2005 bleaching event. Dis. Aquat. Orgs. 2009, 87, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Weil, E.; Cróquer, A. Spatial variability in distribution and prevalence of Caribbean scleractinian coral and octocoral diseases. I. Community-level analysis. Dis. Aquat. Orgs. 2009, 83, 195–208. [Google Scholar] [CrossRef]
- Weil, E.; Rogers, C.S. Coral reef diseases in the Atlantic-Caribbean. In Coral Reefs: An Ecosystem in Transition; Springer: Dordrecht, The Netherlands, 2011; pp. 465–491. [Google Scholar]
- Weil, E.; Rogers, C.S.; Croquer, A. Octocoral diseases in a changing ocean. In Marine Animal Forests; Springer: Cham, Germany, 2016; pp. 1–55. [Google Scholar]
- Pratchett, M.S.; Wilson, S.K.; Baird, A.H. Declines in the abundance of Chaetodon butterflyfishes following extensive coral depletion. J. Fish. Biol. 2006, 69, 1269–1280. [Google Scholar] [CrossRef]
- Graham, N.A.; Wilson, S.K.; Jennings, S.; Polunin, N.V.; Robinson, J.A.N.; Bijoux, J.P.; Daw, T.M. Lag effects in the impacts of mass coral bleaching on coral reef fish, fisheries, and ecosystems. Conserv. Biol. 2007, 21, 1291–1300. [Google Scholar] [CrossRef]
- Munday, P.L.; Jones, G.P.; Sheaves, M.; Williams, A.J.; Goby, G. Vulnerability of fishes of the Great Barrier Reef to climate change. In Climate Change and the Great Barrier Reef; Johnson, J.E., Marshall, P.A., Eds.; Great Barrier Reef Marine Park Authority and Australian Greenhouse Office: Townsville, Australia, 2007; pp. 357–391. [Google Scholar]
- Munday, P.L.; Jones, G.P.; Pratchett, M.S.; Williams, A.J. Climate change and the future for coral reef fishes. Fish Fish. 2008, 9, 261–285. [Google Scholar] [CrossRef]
- Pratchett, M.S.; Munday, P.L.; Wilson, S.K.; Graham, N.A.; Cinner, J.E.; Bellwood, D.R.; Jones, G.P.; Polunin, N.V.; McClanahan, T.R. Effects of climate-induced coral bleaching on coral-reef fishes—ecological and economic consequences. Oceanogr. Mar. Biol. Ann. Rev. 2008, 46, 251–296. [Google Scholar]
- Pratchett, M.S.; Wilson, S.K.; Graham, N.A.J.; Munday, P.L.; Jones, G.P.; Polunin, N.V. Coral bleaching and consequences for motile reef organisms: Past, present and uncertain future effects. In Coral Bleaching, Ecological Studies 205; van Oppen, M.J.H., Lough, J.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 139–158. [Google Scholar]
- Pratchett, M.S.; Hoey, A.S.; Wilson, S.K.; Messmer, V.; Graham, N.A. Changes in biodiversity and functioning of reef fish assemblages following coral bleaching and coral loss. Diversity 2011, 3, 424–452. [Google Scholar] [CrossRef]
- Alvarez-Filip, L.; Paddack, M.J.; Collen, B.; Robertson, D.R.; Côté, I.M. Simplification of Caribbean reef-fish assemblages over decades of coral reef degradation. PLoS ONE 2015, 10, e0126004. [Google Scholar]
- Uthicke, S.; Furnas, M.; Lønborg, C. Coral reefs on the edge? Carbon chemistry on inshore reefs of the Great Barrier Reef. PLoS ONE 2014, 9, e109092. [Google Scholar] [CrossRef]
- Perry, C.T.; Murphy, G.N.; Kench, P.S.; Smithers, S.G.; Edinger, E.N.; Steneck, R.S.; Mumby, P.J. Caribbean-wide decline in carbonate production threatens coral reef growth. Nat. Commun. 2013, 4, 1402–1407. [Google Scholar] [CrossRef]
- Anthony, K.R.; Maynard, J.A.; Díaz-Pulido, G.; Mumby, P.J.; Marshall, P.A.; Cao, L.; Hoegh-Guldberg, O. Ocean acidification and warming will lower coral reef resilience. Glob. Chang. Biol. 2011, 17, 1798–1808. [Google Scholar] [CrossRef]
- Siegel, K.J.; Cabral, R.B.; McHenry, J.; Ojea, E.; Owashi, B.; Lester, S.E. Sovereign states in the Caribbean have lower social-ecological vulnerability to coral bleaching than overseas territories. Proc. R Soc. B 2019, 286, 20182365. [Google Scholar] [CrossRef]
- Hughes, T.P.; Rodrigues, M.J.; Bellwood, D.R.; Ceccarelli, D.; Hoegh-Guldberg, O.; McCook, L.; Moltschaniwskyj, N.; Pratchett, M.S.; Steneck, R.S.; Willis, B. Phase shifts, herbivory, and the resilience of coral reefs to climate change. Curr. Biol. 2007, 17, 360–365. [Google Scholar] [CrossRef]
- Rogers, C.R.; Fitz, C.; Gilnack, M.; Beets, J.; Hardin, J. Scleractinian coral recruitment patterns at Salt River Submarine Canyon, St. Croix, U.S. Virgin Islands. Coral Reefs 1984, 3, 69–76. [Google Scholar] [CrossRef]
- Connell, J.H.; Hughes, T.P.; Wallace, C.C. A 30-year study of coral abundance, recruitment, and disturbance at several scales in space and time. Ecol. Monogr. 1997, 67, 461–488. [Google Scholar] [CrossRef]
- Carlon, D.B. Depth-related patterns of coral recruitment and cryptic suspension-feeding invertebrates on Guan Island, British Virgin Islands. Bull. Mar. Sci. 2001, 68, 525–541. [Google Scholar]
- Darling, E.S.; Côté, I. Seeking resilience in marine ecosystems. Science 2018, 359, 986–987. [Google Scholar] [CrossRef] [PubMed]
- Scheffer, M.; Carpenter, S.; Foley, J.A.; Folke, C.; Walker, B. Catastrophic shifts in ecosystems. Nature 2001, 413, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Bellwood, D.R.; Hughes, T.P.; Folke, C.; Nyström, M. Confronting the coral reef crisis. Nature 2004, 429, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Folke, C.; Carpenter, S.; Walker, B.; Scheffer, M.; Elmqvist, T.; Gunderson, L.; Holling, C.S. Regime shifts, resilience, and biodiversity in ecosystem management. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 557–581. [Google Scholar] [CrossRef]
- Jackson, J.B.C.; Donovan, M.K.; Cramer, K.L.; Lam, V.V. Status and Trends of Caribbean Coral Reefs: 1970–2012; Global Coral Reef Monitoring Network, IUCN: Gland, Switzerland, 2014; pp. 1–304. [Google Scholar]
- O’Cain, E.D.; Frischer, M.E.; Harrison, J.S.; Walters, T.L.; Thompson, M.E.; Fogarty, N.D.; Ruzicka, R.; Gleason, D.F. Identification of newly settled Caribbean coral recruits by ITS-targeted single-step nested multiplex PCR. Coral Reefs 2019, 38, 79–92. [Google Scholar] [CrossRef]
- Irizarry-Soto, E.; Weil, E. Spatial and temporal variability in juvenile coral densities, survivorship and recruitment in La Parguera, southwestern Puerto Rico. Caribb. J. Sci. 2009, 45, 269–281. [Google Scholar] [CrossRef]
- Hernández-Delgado, E.A.; González-Ramos, C.M.; Alejandro-Camis, P.J. Large-scale coral recruitment patterns on Mona Island, Puerto Rico: Evidence of a transitional community trajectory after massive coral bleaching and mortality. Rev. Biol. Trop 2014, 62, 283–298. [Google Scholar]
- Edmunds, P.J. The demography of hurricane effects on two coral populations differing in dynamics. Ecosphere 2019, 10, e02836. [Google Scholar] [CrossRef]
- IPCC. Summary for Policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021; pp. 3–32. [Google Scholar] [CrossRef]
- Fong, P.; Glynn, P.W. A dynamic size-structure population model: Does disturbance control size structure of a population of the massive coral Gardineroseris planulata in the Eastern Pacific? Mar. Biol. 1998, 130, 663–674. [Google Scholar] [CrossRef]
- Bonkosky, M.; Hernández-Delgado, E.A.; Sandoz, B.; Robledo, I.E.; Norat-Ramírez, J.; Mattei, H. Detection of spatial fluctuations of non-point source fecal pollution in coral reef surrounding waters in southwestern Puerto Rico using PCR-based assays. Mar. Poll. Bull. 2009, 58, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Delgado, E.A.; Sandoz, B.; Bonkosky, M.; Mattei, H.; Norat, J. Impacts of non-point source sewage pollution in Elkhorn coral, Acropora palmata (Lamarck), assemblages of the southwestern Puerto Rico shelf. In Proceedings of the 11th International Coral Reef Symposium, Ft. Lauderdale, FL, USA, 7–11 July 2008; pp. 747–751. [Google Scholar]
- Díaz-Ortega, G.; Hernández-Delgado, E.A. Land-based source pollution in a climate of change: A roadblock to the conservation and recovery of Elkhorn coral Acropora palmata (Lamarck 1816). Nat. Resour. 2014, 5, 561–581. [Google Scholar]
- Rogers, C.S.; Garrison, G.; Gtober, R.; Hillis, Z.M.; Franke, M.A. Coral Reef Monitoring Manual for the Caribbean and Western Atlantic; National Park Service: St. John, NY, USA, 1994. [Google Scholar]
- Anderson, M.; Gorley, R.; Clarke, K. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Clarke, K.; Gorley, R.; Somerfield, P.; Warwick, R. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 3rd ed.; PRIMER-E: Plymouth, UK, 2014. [Google Scholar]
- Cangialosi, J.P.; Latto, A.S.; Berg, R. Hurricane Irma (AL112017) 30 August–12 September 2017; National Hurricane Center Tropical Cyclone Report; NOAA: Washington, DC, USA, 2018; pp. 1–111. [Google Scholar]
- Pasch, R.J.; Penny, A.B.; Berg, R. Hurricane María (AL152017) 16–30 September 2017; National Hurricane Center Tropical Cyclone Report; NOAA: Washington, DC, USA, 2018; pp. 1–48. [Google Scholar]
- Lapointe, B.E.; Clark, M.W. Nutrient inputs from the watershed and coastal eutrophication in the Florida Keys. Estuaries 1992, 15, 465–476. [Google Scholar] [CrossRef]
- Szmant, A.M. Nutrient enrichment on coral reefs: Is it a major cause of coral reef decline? Estuaries 2002, 25, 743–766. [Google Scholar] [CrossRef]
- Norat-Ramírez, J.; Méndez-Lázaro, P.; Hernández-Delgado, E.A.; Mattei-Torres, H.; Cordero-Rivera, L. A septic weight index model to measure the impact of septic tanks on coastal water quality and coral reef communities in Rincón, Puerto Rico. Ocean Coast Manag. 2019, 169, 201–213. [Google Scholar] [CrossRef]
- Díaz, J.R.; Jordan, D.G. Water Resources of the Río Grande de Añasco-Lower Valley, Puerto Rico. U.S Geological Survey Water Resources Investigation Report 85-4237. 1987. Available online: https://pubs.er.usgs.gov/publication/wri854237 (accessed on 20 November 2021).
- Hernández-Delgado, E.A.; Rosado-Matías, B.J.; Sabat, A.M. Management failures and coral decline threatens fish functional groups recovery patterns in the Luis Peña Channel No-Take Natural Reserve, Culebra Island, PR. Proc. Gulf Caribb. Fish. Inst. 2006, 57, 577–605. [Google Scholar]
- Hernández-Delgado, E.A.; Medina-Muñiz, J.L.; Mattei, H.; Norat-Ramírez, J. Unsustainable land use, sediment-laden runoff, and chronic raw sewage offset the benefits of coral reef ecosystems in a no-take marine protected area. Env. Mgmt. Sust. Dev. 2017, 6, 292–333. [Google Scholar] [CrossRef]
- Alhajjar, B.J.; Chesters, G.; Harkin, J.M. Indicators of chemical pollution from septic systems. Groundwater 1990, 28, 559–568. [Google Scholar] [CrossRef]
- Fay, S.R.; Spong, R.C.; Alexander, S.C.; Alexander, E.C., Jr. Optical brighteners: Sorption behavior, detection, septic system tracer applications. In Proceedings of the International Association of Hydrogeologists XXVI International Congress, Edmonton, AB, Canada, June 1995. [Google Scholar]
- Tavares, M.E.; Spivey, M.I.; McIver, M.R.; Mallin, M.A. Testing for optical brighteners and fecal bacteria to detect sewage leaks in tidal creeks. J. North Car. Acad. Sci. 2008, 124, 91–97. [Google Scholar]
- Cao, Y.; Griffith, J.F.; Weisberg, S.B. Evaluation of optical brightener photodecay characteristics for detection of human fecal contamination. Water Res. 2009, 43, 2273–2279. [Google Scholar] [CrossRef]
- Hagedorn, C.; Weisberg, S.B. Chemical-based fecal source tracking methods: Current status and guidelines for evaluation. Rev. Env. Sci. Bio. Technol. 2009, 8, 275–287. [Google Scholar] [CrossRef]
- Chandler, D.M.; Lerner, D.N. A low cost method to detect polluted surface water outfalls and misconnected drainage. Water Environ. J. 2015, 29, 202–206. [Google Scholar] [CrossRef]
- González-Figueroa, M.C.; Hernández-Delgado, E.A. Variación espacial en los patrones de recuperación natural de los arrecifes de coral someros urbanos en Puerto Rico. Perspect. Asun. Ambient. 2021, 9, 90–111. Available online: https://uagm.edu/es/v9-perspectivas (accessed on 1 July 2022).
- Fabricius, K.E. Factors determining the resilience of coral reefs to eutrophication: A review and conceptual model. In Coral Reefs: An Ecosystem in Transition; Dubinsky, Z., Sambler, N., Eds.; Springer Science: New York, NY, USA, 2011; pp. 493–505. [Google Scholar]
- Babcock, R.D.P. Effects of sedimentation on settlement of Acropora millepora. Coral Reefs 1991, 9, 205–208. [Google Scholar] [CrossRef]
- Kuffner, I.R.; Walters, L.J.; Becerro, M.A.; Paul, V.J.; Williams, R.R.; Beach, K.S. Inhibition of coral recruitment by macroalgae and cyanobacteria. Mar. Ecol. Prog. Ser. 2006, 323, 107–117. [Google Scholar] [CrossRef]
- Hoey, A.S.; Pratchett, M.S.; Cvitanovic, C. High macroalgal cover and low coral recruitment undermines the potential resilience of the world’s southernmost coral reef assemblages. PLoS ONE 2011, 6, e25824. [Google Scholar] [CrossRef]
- Patterson, K.L.; Porter, J.W.; Ritchie, K.B.; Polson, S.W.; Mueller, E.; Peters, E.C.; Santavy, D.L.; Smith, G.W. The etiology of white pox, a lethal disease of the Caribbean elkhorn coral, Acropora palmata. Proc. Natl. Acad. Sci. USA 2002, 99, 8725–8730. [Google Scholar] [CrossRef]
- Sutherland, K.P.; Porter, J.W.; Torres, C. Disease and immunity in Caribbean and Indo-Pacific zooxanthellate corals. Mar. Ecol. Progr. Ser. 2004, 266, 273–302. [Google Scholar] [CrossRef]
- Kaczmarsky, L.T.; Draud, M.; Williams, E.H. Is there a relationship between proximity to sewage effluent and the prevalence of coral disease. Caribb. J. Sci. 2005, 41, 124–137. [Google Scholar]
- Voss, J.D.; Richardson, L.L. Nutrient enrichment enhances black band disease progression in corals. Coral Reefs 2006, 25, 569–576. [Google Scholar] [CrossRef]
- Sutherland, K.P.; Porter, J.W.; Turner, J.W.; Thomas, B.J.; Looney, E.E.; Luna, T.P.; Meyers, M.K.; Futch, J.C.; Lipp, E.K. Human sewage identified as likely source of white pox disease of the threatened Caribbean elkhorn coral, Acropora palmata. Environ. Microbiol. 2010, 12, 1122–1131. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Delgado, E.A.; Hutchinson-Delgado, Y.M.; Laureano, R.; Hernández-Pacheco, R.; Ruiz-Maldonado, T.M.; Oms, J.; Díaz, P.L. Sediment stress, water turbidity and sewage impacts on threatened Elkhorn coral (Acropora palmata) stands at Vega Baja, Puerto Rico. Proc. Gulf Caribb. Fish. Inst. 2011, 63, 83–92. [Google Scholar]
- Kaczmarsky, L.; Richardson, L.L. Do elevated nutrients and organic carbon on Philippine reefs increase the prevalence of coral disease? Coral Reefs 2011, 30, 253–257. [Google Scholar] [CrossRef]
- Sutherland, K.P.; Shaban, S.; Joyner, J.L.; Porter, J.W.; Lipp, E.K. Human Pathogen Shown to Cause Disease in the Threatened Eklhorn Coral Acropora palmata. PLoS ONE 2011, 6, e23468. [Google Scholar] [CrossRef] [PubMed]
- Redding, J.E.; Myers-Miller, R.L.; Baker, D.M.; Fogel, M.; Raymundo, L.J.; Kim, K. Link between sewage-derived nitrogen pollution and coral disease severity in Guam. Mar. Poll. Bull. 2013, 73, 57–63. [Google Scholar] [CrossRef]
- Yoshioka, R.M.; Kim, C.J.; Tracy, A.M.; Most, R.; Harvell, C.D. Linking sewage pollution and water quality to spatial patterns of Porites lobata growth anomalies in Puako, Hawaii. Mar. Poll. Bull. 2016, 104, 313–321. [Google Scholar] [CrossRef]
- Wear, S.L.; Thurber, R.V. Sewage pollution: Mitigation is key for coral reef stewardship. Ann. N. Y. Acad. Sci. 2015, 1355, 15–30. [Google Scholar] [CrossRef]
- Wakwella, A.; Mumby, P.J.; Roff, G. Sedimentation and overfishing drive changes in early succession and coral recruitment. Proc. R. Soc. B 2020, 287, 20202575. [Google Scholar] [CrossRef]
- Kennedy, E.V.; Perry, C.T.; Halloran, P.R.; Iglesias-Prieto, R.; Schönberg, C.H.; Wisshak, M.; Form, A.U.; Carricart-Gavinet, J.P.; Fine, M.; Eakin, C.M.; et al. Avoiding coral reef functional collapse requires local and global action. Curr. Biol. 2013, 23, 912–918. [Google Scholar] [CrossRef]
- Hernández-Delgado, E.A. The Emerging Threats of Climate Change on Tropical Coastal Ecosystem Services, Public Health, Local Economies and Livelihood Sustainability of Small Islands: Cumulative Impacts and Synergies. Mar. Poll. Bull. 2015, 101, 5–28. Available online: http://www.sciencedirect.com/science/article/pii/S0025326X15300357 (accessed on 20 November 2021). [CrossRef]
- Rogers, C.S.; Ramos-Scharrón, C.E. Assessing effects of sediment delivery to coral reefs: A Caribbean watershed perspective. Front. Mar. Sci. 2022, 8, 773968. [Google Scholar] [CrossRef]
- Otaño-Cruz, A.; Montañez-Acuña, A.A.; Torres-López, V.I.; Hernández-Figueroa, E.M.; Hernández-Delgado, E.A. Effects of changing weather, oceanographic conditions, and land uses on spatio-temporal variation of sedimentation dynamics along near-shore coral reefs. Front. Mar. Sci 2017, 4, 249. [Google Scholar] [CrossRef]
- Otaño-Cruz, A.; Montañez-Acuña, A.A.; Benson, E.; Cuevas, E.P.; Ortiz, J.; Hernández-Delgado, E.A. Response of near-shore coral reefs benthic communities to changes of sedimentation dynamics and environmental conditions. Front. Mar. Sci 2019, 6, 551. [Google Scholar] [CrossRef]
- Gómez Andújar, N.X.; Hernández-Delgado, E.A. Spatial benthic community analysis of shallow coral reefs to support coastal management in Culebra Island, Puerto Rico. PeerJ 2020, 8, 10080. [Google Scholar] [CrossRef] [PubMed]
- Woodley, J.D.; Chornesky, E.A.; Clifford, P.A.; Jackson, J.B.C.; Kaufman, L.S.; Knowlton, N.; Lang, J.C.; Pearson, M.P.; Porter, J.W.; Rooney, M.C.; et al. Hurricane Allen’s impact on Jamaican coral reefs. Science 1981, 214, 749–755. [Google Scholar] [CrossRef] [PubMed]
- Harmelin-Vivien, M.L.; Laboute, P. Catastrophic impact of hurricanes on atoll outer reef slopes in the Tuamotu (French Polynesia). Coral Reefs 1986, 5, 55–62. [Google Scholar] [CrossRef]
- Fenner, D.P. Effects of Hurricane Gilbert on coral reefs, fishes and sponges at Cozumel, Mexico. Bull. Mar. Sci. 1991, 48, 719–730. [Google Scholar]
- Lugo, A.E.; Rogers, C.S.; Nixon, S.W. Hurricanes, coral reefs and rainforests: Resistance, ruin and recovery in the Caribbean. AMBIO 2000, 29, 106–114. [Google Scholar] [CrossRef]
- Stoddart, D.R. Three Caribbean atolls: Turneffe Islands, Lighthouse Reef, and Glover’s Reef, British Honduras. Atoll Res. Bull. 1962, 87, 1–161. [Google Scholar] [CrossRef]
- Stoddart, D.R. Effects of Hurricane Hattie on the British Honduras reefs and cays, 30–31 October 1961. Atoll Res. Bull. 1963, 95, 1–142. [Google Scholar] [CrossRef]
- Rogers, C.S. A matter of scale: Damage from Hurricane Hugo (1989) to US Virgin Islands reefs at the colony, community and whole reef level. In Proceedings of the 7th International Coral Reef Symposium, Guam, Micronesia, 22 June 1992; Volume 1, pp. 127–133. [Google Scholar]
- Zimmerman, J.K.; Willig, M.X.; Hernández-Delgado, E.A. Resistance, resilience and vulnerability of socio-ecological systems to hurricanes in Puerto Rico. Ecosphere 2020, 11, e03159. [Google Scholar] [CrossRef]
- Nieto, R.R.; Carrara, X.C.; Sheppard, C. Efectos de los huracanes sobre la estabilidad de paisajes asociados con arrecifes coralinos. Cien. Mar. 2012, 38, 47–55. [Google Scholar]
- Lirman, D.; Fong, P. Patterns of damage to the branching coral Acropora palmata following Hurricane Andrew: Damage and survivorship ofhurricane-generated asexual recruits. J. Coast. Res. 1997, 13, 67–72. [Google Scholar]
- Hubbard, D.K. Sedimentation as a control of reef development: St. Croix. USVI. Coral Reefs 1986, 5, 117–125. [Google Scholar] [CrossRef]
- Ritson-Williams, R.; Arnold, S.N.; Fogarty, N.D.; Steneck, R.S.; Vermeij, M.J.; Paul, V.J. New perspectives on ecological mechanisms affecting coral recruitment on reefs. Smith Contrib. Mar. Sci. 2009, 38, 437–457. [Google Scholar] [CrossRef]
- Doropoulos, C.; Ward, S.; Díaz-Pulido, G.; Hoegh-Guldberg, O.; Mumby, P.J. Ocean acidification reduces coral recruitment by disrupting intimate larval-algal settlement interactions. Ecol. Lett. 2012, 15, 338–346. [Google Scholar] [CrossRef]
- Harrington, L.; Fabricius, K.; De’ath, G.; Negri, A. Recognition and selection of settlement substrata determine post-settlement survival in corals. Ecology 2004, 85, 3428–3437. [Google Scholar] [CrossRef]
- Day, R.W. Effects of benthic algae on sessile animals: Observational evidence from coral reef habitats. Bull. Mar. Sci. 1983, 33, 597–605. [Google Scholar]
- Wolf, A.T.; Nugues, M.M. Predation on coral settlers by the corallivorous fireworm Hermodice carunculata. Coral Reefs 2013, 32, 227–231. [Google Scholar] [CrossRef]
- Arnold, S.N.; Steneck, R.S.; Mumby, P.J. Running the gauntlet: Inhibitory effects of algal turfs on the processes of coral recruitment. Mar. Ecol. Prog. Ser. 2010, 414, 91–105. [Google Scholar] [CrossRef]
- Roff, G.; Chollett, I.; Doropoulos, C.; Golbuu, Y.; Steneck, R.S.; Isechal, A.L.; van Woesik, R.; Mumby, P.J. Exposure-driven macroalgal phase shift following catastrophic disturbance on coral reefs. Coral Reefs 2015, 34, 715–725. [Google Scholar] [CrossRef]
- Maida, M.; Coll, J.C.; Sammarco, P.W. Shedding new light on scleractinian coral recruitment. J. Exp. Mar. Biol. Ecol. 1994, 180, 189–202. [Google Scholar] [CrossRef]
- Baird, A.H.; Babcock, R.C.; Mundy, C.P. Habitat selection by larvae influences the depth distribution of six common coral species. Mar. Ecol. Progr. Ser. 2003, 252, 289–293. [Google Scholar] [CrossRef]
- Edmunds, P.J.; Leichter, J.J.; Adjeroud, M. Landscape-scale variation in coral recruitment in Moorea, French Polynesia. Mar. Ecol. Prog. Ser. 2010, 414, 75–89. [Google Scholar] [CrossRef][Green Version]
- Salinas-de-León, P.; Dryden, C.; Smith, D.J.; Bell, J.J. Temporal and spatial variability in coral recruitment on two Indonesian coral reefs: Consistently lower recruitment to a degraded reef. Mar. Biol. 2013, 160, 97–105. [Google Scholar] [CrossRef]
- Fox, H.E. Coral recruitment in blasted and unblasted sites in Indonesia: Assessing rehabilitation potential. Mar. Ecol. Progr. Ser. 2004, 269, 131–139. [Google Scholar] [CrossRef]
- Fox, H.E.; Mous, P.J.; Pet, J.S.; Muljadi, A.H.; Caldwell, R.L. Experimental assessment of coral reef rehabilitation following blast fishing. Conserv. Biol. 2005, 19, 98–107. [Google Scholar] [CrossRef]
- Raymundo, L.J.; Maypa, A.P.; Gomez, E.D.; Cadiz, P. Can dynamite-blasted reefs recover? A novel, low-tech approach to stimulating natural recovery in fish and coral populations. Mar. Poll. Bull. 2007, 54, 1009–1019. [Google Scholar] [CrossRef]
- Harmelin-Vivien, M.L. The effects of storms and cyclones on coral reefs: A review. J. Coast. Res. 1994, 211–231. [Google Scholar]
- Dajka, J.C.; Wilson, S.K.; Robinson, J.P.; Chong-Seng, K.M.; Harris, A.; Graham, N.A. Uncovering drivers of juvenile coral density following mass bleaching. Coral Reefs 2019, 38, 637–649. [Google Scholar] [CrossRef]
- Mallela, J.; Crabbe, M.J.C. Hurricanes and coral bleaching linked to changes in coral recruitment in Tobago. Mar. Environ. Res. 2009, 68, 158–162. [Google Scholar] [CrossRef] [PubMed]
- Mumby, P.J.; Harborne, A.R.; Williams, J.; Kappel, C.V.; Brumbaugh, D.R.; Micheli, F.; Holmes, K.E.; Dahlgren, C.P.; Paris, C.B.; Blackwell, P.G. Trophic cascade facilitates coral recruitment in a marine reserve. Proc. Natl. Acad. Sci. USA 2007, 104, 8362–8367. [Google Scholar] [CrossRef] [PubMed]
- Bozec, Y.M.; Alvarez-Filip, L.; Mumby, P.J. The dynamics of architectural complexity on coral reefs under climate change. Glob. Chang. Biol. 2014, 21, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Edmunds, P.J.; Elahi, R. The demographics of a 15-year decline in cover of the Caribbean reef coral Montastraea annularis. Ecol. Monogr. 2007, 77, 3–18. [Google Scholar] [CrossRef]
- Weil, E.; Hernández-Delgado, E.A.; González, M.; Williams, S.; Suleimán-Ramos, S.; Figuerola, M.; Metz-Estrella, T. Spread of the new coral disease “SCTLD” into the Caribbean: Implications for Puerto Rico. Reef Encount. 2009, 34, 38–43. [Google Scholar]
- Meiling, S.; Smith, T.B.; Muller, E.; Brandt, M.E. Stony coral tissue loss disease (SCTLD) lesion progression slows in association with thermal stress. Front. Mar. Sci. 2020, 7, 1128. [Google Scholar] [CrossRef]
- Miller, J.; Waara, R.; Muller, E.; Rogers, C. Coral bleaching and disease combine to cause extensive mortality on reefs in US Virgin Islands. Coral Reefs 2006, 25, 418. [Google Scholar] [CrossRef]
- Rogers, C.S.; Muller, E.; Spitzack, T.; Miller, J. Extensive coral mortality in the US Virgin Islands in 2005/2006: A review of the evidence for synergy among thermal stress, coral bleaching and disease. Caribb. J. Sci. 2009, 45, 204–214. [Google Scholar] [CrossRef]
- Rogers, C. Coral bleaching and disease should not be underestimated as causes of Caribbean coral reef decline. Proc. R. Soc. B Biol. Sci. 2009, 276, 197–198. [Google Scholar] [CrossRef]
- Aeby, G.; Ushijima, B.; Bartels, E.; Walter, C.; Kuehl, J.; Jones, S.; Paul, V.J. Changing stony coral tissue loss disease dynamics through time in Montastraea cavernosa. Front. Mar. Sci. 2021, 8, 699075. [Google Scholar] [CrossRef]
- Brandt, M.E.; Ennis, R.S.; Meiling, S.S.; Townsend, J.; Cobleigh, K.; Glahn, A.; Quetel, J.; Brandtneris, V.; Henderson, L.M.; Smith, T.B. The emergence and initial impact of stony coral tissue loss disease (SCTLD) in the United States Virgin Islands. Front. Mar. Sci. 2021, 8, 715329. [Google Scholar] [CrossRef]
- Estrada-Saldívar, N.; Quiroga-García, B.A.; Pérez-Cervantes, E.; Rivera-Garibay, O.O.; Alvarez-Filip, L. Effects of the stony coral tissue loss disease outbreak on coral communities and the benthic composition of Cozumel Reefs. Front. Mar. Sci. 2021, 8, 632777. [Google Scholar] [CrossRef]
- Neely, K.L.; Lewis, C.L.; Lunz, K.S.; Kabay, L. Rapid population decline of the pillar coral Dendrogyra cylindrus along the Florida Reef Tract. Front. Mar. Sci. 2021, 8, 656515. [Google Scholar] [CrossRef]
- Spadafore, R.; Fura, R.; Precht, W.F.; Vollmer, S.V. Multi-variate analyses of coral mortality from the 2014–2015 stony coral tissue loss disease outbreak off Miami-Dade County, Florida. Front. Mar. Sci. 2021, 8, 723998. [Google Scholar] [CrossRef]
- Alvarez-Filip, L.; González-Barrios, F.; Pérez-Cervantes, E.; Molina-Hernandez, A.; Estrada-Saldívar, N. An emerging coral disease outbreak decimated Caribbean coral populations and reshaped reef functionality. Res. Sq. 2021, 1–17. [Google Scholar] [CrossRef]
- Perry, C.T.; Murphy, G.N.; Kench, P.S.; Edinger, E.N.; Smithers, S.G.; Steneck, R.S.; Mumby, P.J. Changing dynamics of Caribbean reef carbonate budgets: Emergence of reef bioeroders as critical controls on present and future reef growth potential. Proc. R. Soc. B Biol. Sci. 2014, 281, 20142018. [Google Scholar] [CrossRef]
- Perry, C.T.; Alvarez-Filip, L. Changing geo-ecological functions of coral reefs in the Anthropocene. Funct. Ecol. 2019, 33, 976–988. [Google Scholar] [CrossRef]
- González-Barrios, F.J.; Cabral-Tena, R.A.; Alvarez-Filip, L. Recovery disparity between coral cover and the physical functionality of reefs with impaired coral assemblages. Glob. Chang. Biol. 2021, 27, 640–651. [Google Scholar] [CrossRef]
- Carpenter, R.C.; Edmunds, P.J. Local and regional scale recovery of Diadema promotes recruitment of scleractinian corals. Ecol. Lett. 2006, 9, 271–280. [Google Scholar] [CrossRef]
- Mumby, P.J. Bleaching and hurricane disturbances to populations of coral recruits in Belize. Mar. Ecol. Prog. Ser. 1999, 190, 27–35. [Google Scholar] [CrossRef]
- Miller, M.W.; Weil, E.; Szmant, A.M. Coral recruitment and juvenile mortality as structuring factors for reef benthic communities in Biscayne National Park, USA. Coral Reefs 2000, 19, 115–123. [Google Scholar] [CrossRef]
- Ruiz-Zárate, M.A.; Arias-González, J.E. Spatial study of juvenile corals in the Northern region of the Mesoamerican Barrier Reef System (MBRS). Coral Reefs 2004, 23, 584–594. [Google Scholar]
- Rylaarsdam, K.W. Life histories and abundance patterns of colonial corals on Jamaican reefs. Mar. Ecol. Prog. Ser. 1983, 13, 249–260. [Google Scholar] [CrossRef]
- Bak, R.P.M.; Engel, M.S. Distribution, abundance and survival of juvenile hermatypic corals (Scleractinia) and the importance of life history strategies in the parent coral community. Mar. Biol. 1979, 54, 341–352. [Google Scholar] [CrossRef]
- Mumby, P.J.; Harborne, A.R. Marine reserves enhance the recovery of corals on Caribbean reefs. PLoS ONE 2010, 5, e8657. [Google Scholar] [CrossRef]
- Edmunds, P.J. Juvenile coral population dynamics track rising seawater temperature on a Caribbean reef. Mar. Ecol. Prog. Ser. 2004, 269, 111–119. [Google Scholar] [CrossRef]
- Green, D.H.; Edmunds, P.J.; Carpenter, R.C. Increasing relative abundance of Porites astreoides on Caribbean reefs mediated by an overall decline in coral cover. Mar. Ecol. Progr. Ser. 2008, 359, 1–10. [Google Scholar] [CrossRef]
- Lowe, R.J.; Falter, J.L. Oceanic forcing of coral reefs. Annu. Rev. Mar. Sci. 2015, 7, 43–66. [Google Scholar] [CrossRef]
- Storlazzi, C.; Reguero, B.; Lowe, E.; Shope, J.; Gibbs, A.; Beck, M.; Nickel, B. Rigorously valuing the role of coral reefs in coastal protection: An example from Maui, Hawaii, USA. Coast. Dynam. 2017, 35, 665–674. [Google Scholar]
- Rogers, A.; Blanchard, J.L.; Mumby, P.J. Fisheries productivity under progressive coral reef degradation. J. Appl. Ecol. 2017, 55, 1041–1049. [Google Scholar] [CrossRef]
- Rogers, A.; Blanchard, J.L.; Newman, S.P.; Dryden, C.S.; Mumby, P.J. High refuge availability on coral reefs increases the vulnerability of reef-associated predators to overexploitation. Ecology 2018, 99, 450–463. [Google Scholar] [CrossRef]
- Osorio-Cano, J.D.; Osorio, A.F.; Alcérreca-Huerta, J.C.; Oumeraci, H. Drag and inertia forces on a branched coral colony of Acropora palmata. J. Fluids Struct. 2019, 88, 31–47. [Google Scholar] [CrossRef]
- Reguero, B.G.; Secaira, F.; Toimil, A.; Escudero, M.; Díaz-Simal, P.; Beck, M.W.; Silva, R.; Storlazzi, C.; Losada, I.J. The risk reduction benefits of the Mesoamerican reef in Mexico. Front. Earth Sci. 2019, 7, 125. [Google Scholar] [CrossRef]
- Asher, S.; Shavit, U. The effect of water depth and internal geometry on the turbulent flow inside a coral reef. J. Geophys. Res. Ocean. 2019, 124, 3508–3522. [Google Scholar] [CrossRef]
- Zhu, L.; Chen, Q.; Ding, Y.; Jafari, N.; Rosati, J.D. Semianalytical model of depth-integrated vegetal drag force based on Stokes second-order wave theory. J. Waterw. Port Coast. Ocean. Eng. 2019, 145, 04018041. [Google Scholar] [CrossRef]
- Smithers, S.G.; Hoeke, R.K. Geomorphological impacts of high-latitude storm waves on low-latitude reef islands—Observations of the December 2008 event on Nukutoa, Takuu, Papua New Guinea. Geomorphology 2014, 222, 106–121. [Google Scholar] [CrossRef]
- Tajima, Y.; Shimozono, T.; Gunasekara, K.H.; Cruz, E.C. Study on locally varying inundation characteristics induced by Super Typhoon Haiyan. Part 2: Deformation of storm waves on the beach with fringing reef along the East Coast of Eastern Samar. Coast. Eng. J. 2016, 58, 1640003-1–1640003-24. [Google Scholar] [CrossRef]
- Pomeroy, A.W.; Lowe, R.J.; Ghisalberti, M.; Storlazzi, C.; Symonds, G.; Roelvink, D. Sediment transport in the presence of large reef bottom roughness. J. Geophys. Res. Ocean. 2017, 122, 1347–1368. [Google Scholar] [CrossRef]
- Pomeroy, A.W.; Lowe, R.J.; Ghisalberti, M.; Winter, G.; Storlazzi, C.; Cuttler, M. Spatial variability of sediment transport processes over intratidal and subtidal timescales within a fringing coral reef system. J. Geophys. Res. Earth Surf. 2018, 123, 1013–1034. [Google Scholar] [CrossRef]
- Graham, N.A.J.; Nash, K.L. The importance of structural complexity in coral reef ecosystems. Coral Reefs 2013, 32, 315–326. [Google Scholar] [CrossRef]
- Rogers, A.; Blanchard, J.L.; Mumby, P.J. Vulnerability of coral reef fisheries to a loss of structural complexity. Curr. Biol. 2014, 24, 1000–1005. [Google Scholar] [CrossRef]
- Rogers, A.; Harborne, A.R.; Brown, C.J.; Bozec, Y.M.; Castro, C.; Chollett, I.; Hock, K.; Knowland, C.A.; Marshell, A.; Ortiz, J.C.; et al. Anticipative management for coral reef ecosystem services in the 21st century. Glob. Chang. Biol. 2015, 21, 504–514. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández-Delgado, E.A.; Ortiz-Flores, M.F. The Long and Winding Road of Coral Reef Recovery in the Anthropocene: A Case Study from Puerto Rico. Diversity 2022, 14, 804. https://doi.org/10.3390/d14100804
Hernández-Delgado EA, Ortiz-Flores MF. The Long and Winding Road of Coral Reef Recovery in the Anthropocene: A Case Study from Puerto Rico. Diversity. 2022; 14(10):804. https://doi.org/10.3390/d14100804
Chicago/Turabian StyleHernández-Delgado, Edwin A., and María F. Ortiz-Flores. 2022. "The Long and Winding Road of Coral Reef Recovery in the Anthropocene: A Case Study from Puerto Rico" Diversity 14, no. 10: 804. https://doi.org/10.3390/d14100804
APA StyleHernández-Delgado, E. A., & Ortiz-Flores, M. F. (2022). The Long and Winding Road of Coral Reef Recovery in the Anthropocene: A Case Study from Puerto Rico. Diversity, 14(10), 804. https://doi.org/10.3390/d14100804