
Can We Avoid Tacit Trade-Offs between Flexibility and Efficiency in Systematic Conservation Planning? The Mediterranean Sea as a Case Study

 , ,
, , 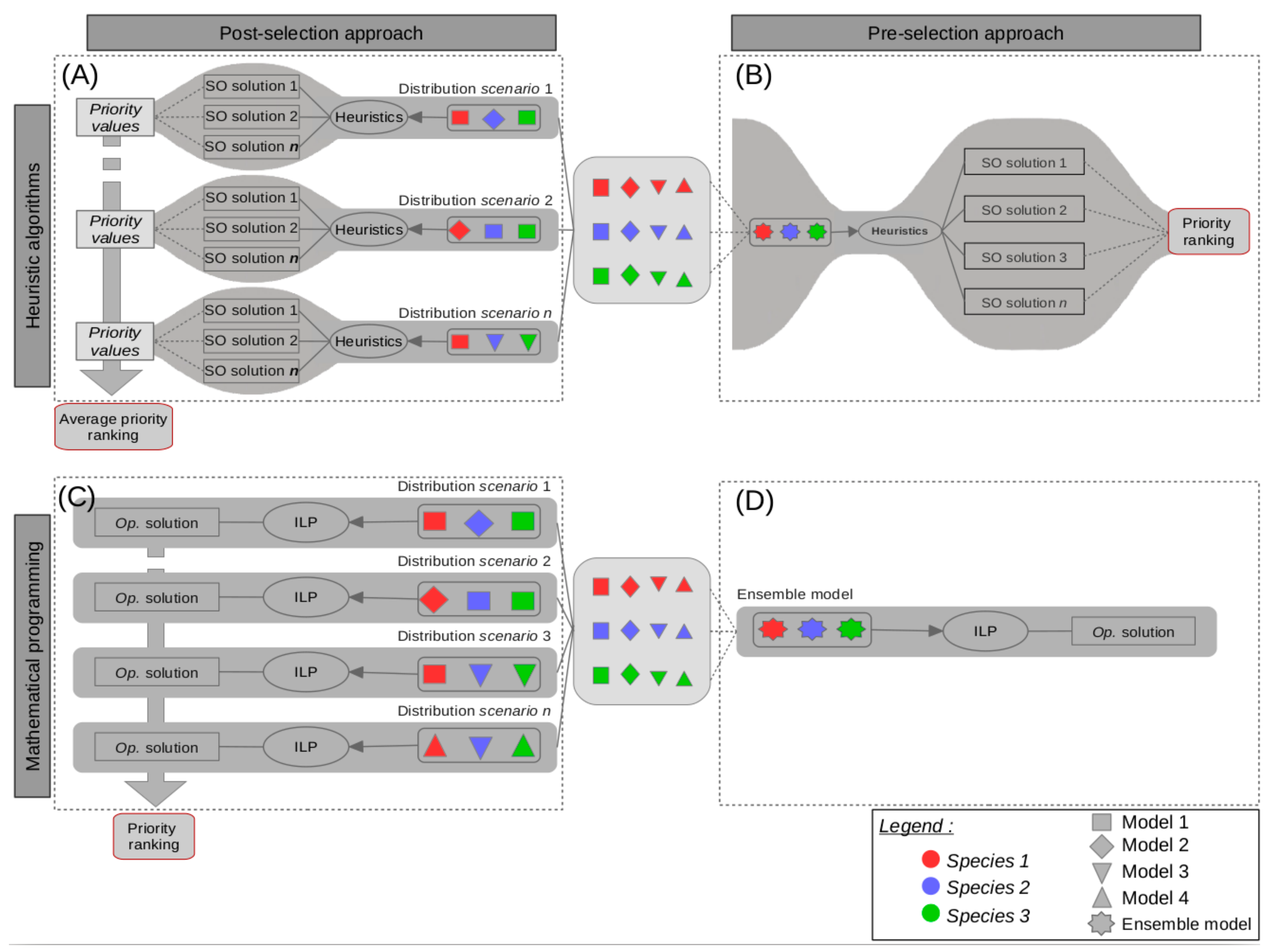{kind=link}
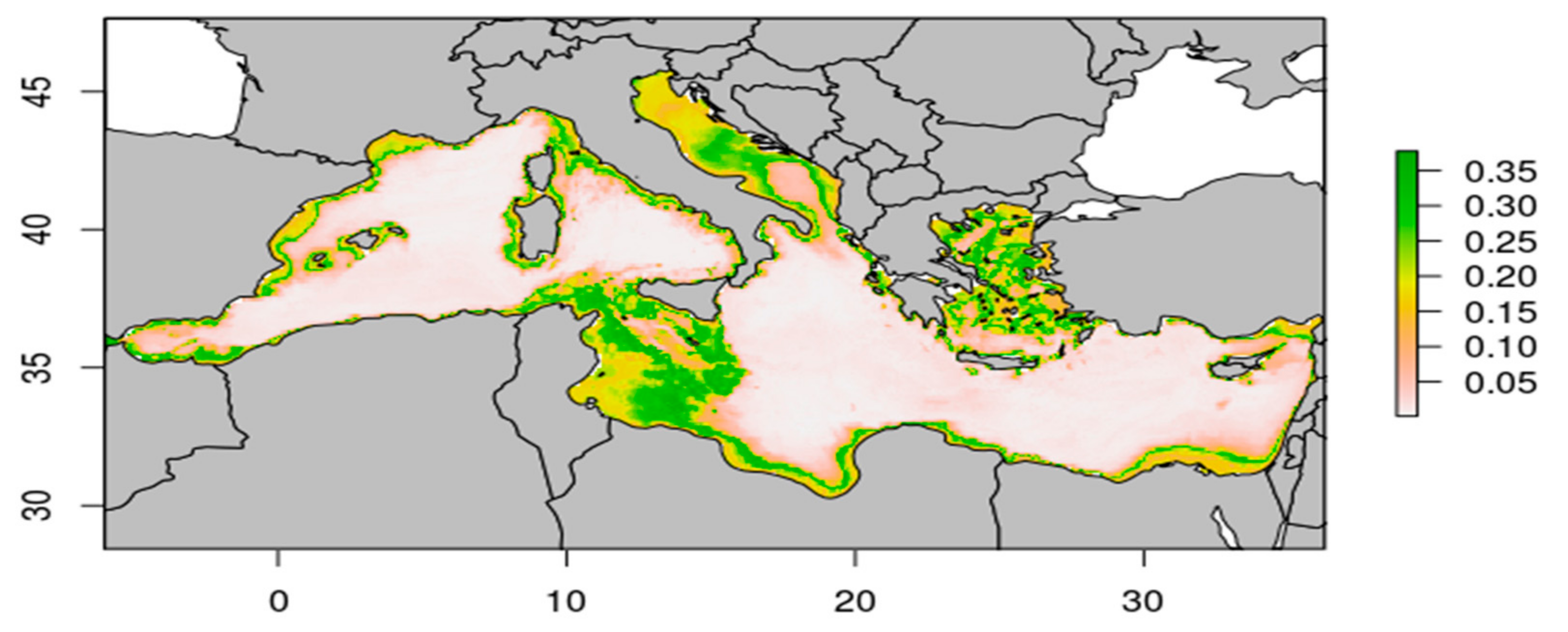{kind=link}
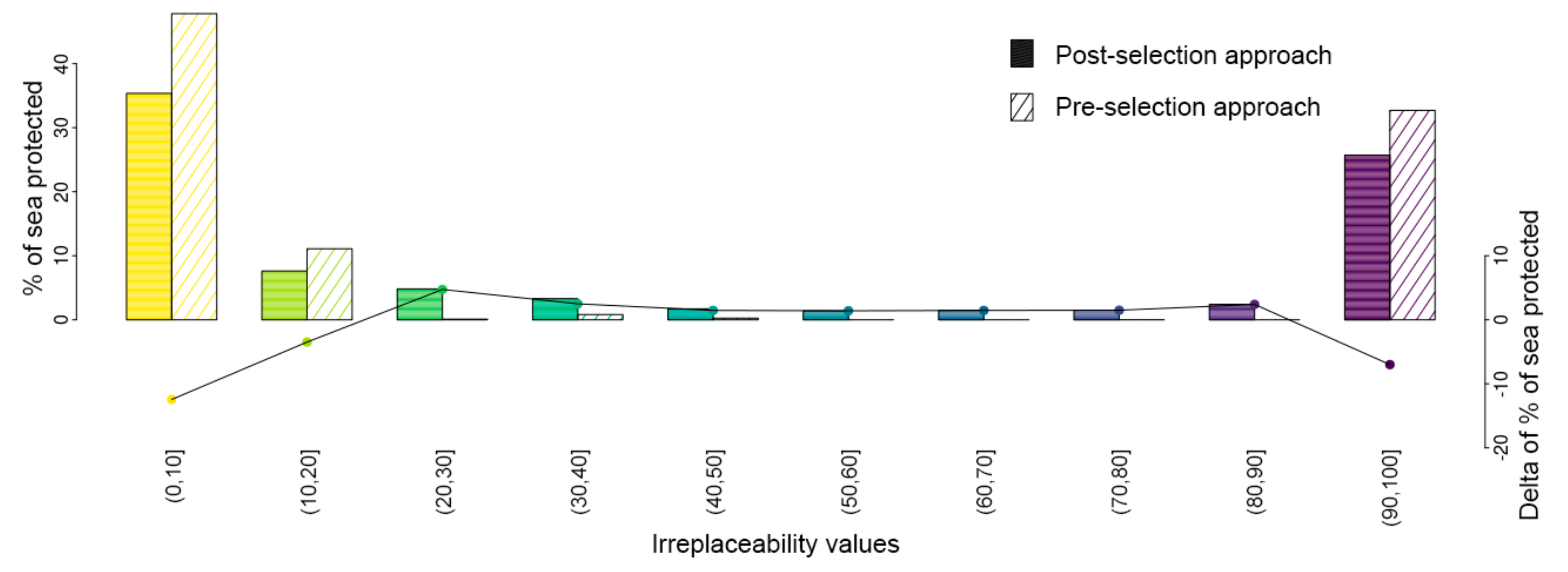{kind=link}
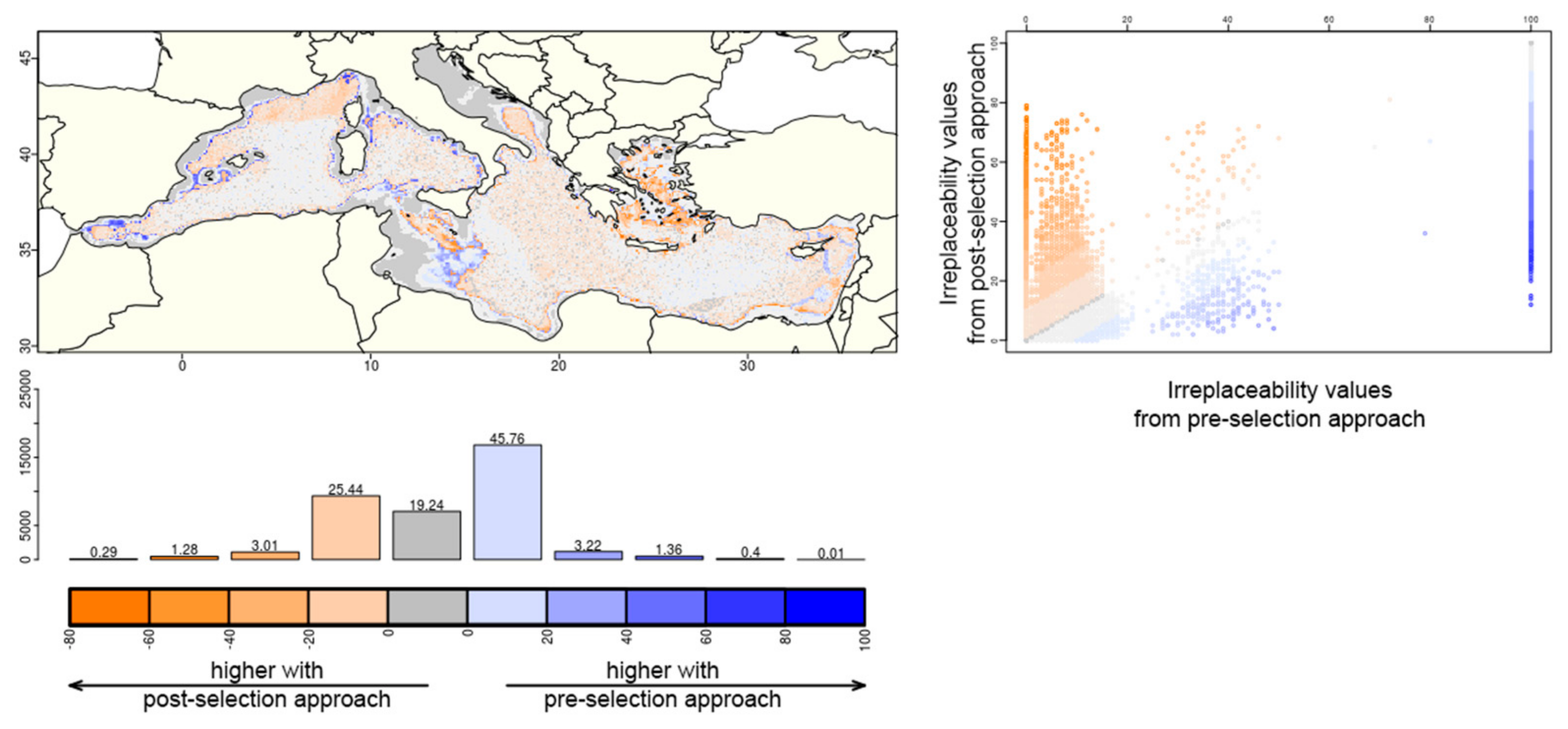{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Species Distribution Modeling
2.2. Species Distribution Scenarios
2.3. Conservation Targets, Reserve Selection Algorithms, and Priority Rankings
2.4. Comparative Analysis
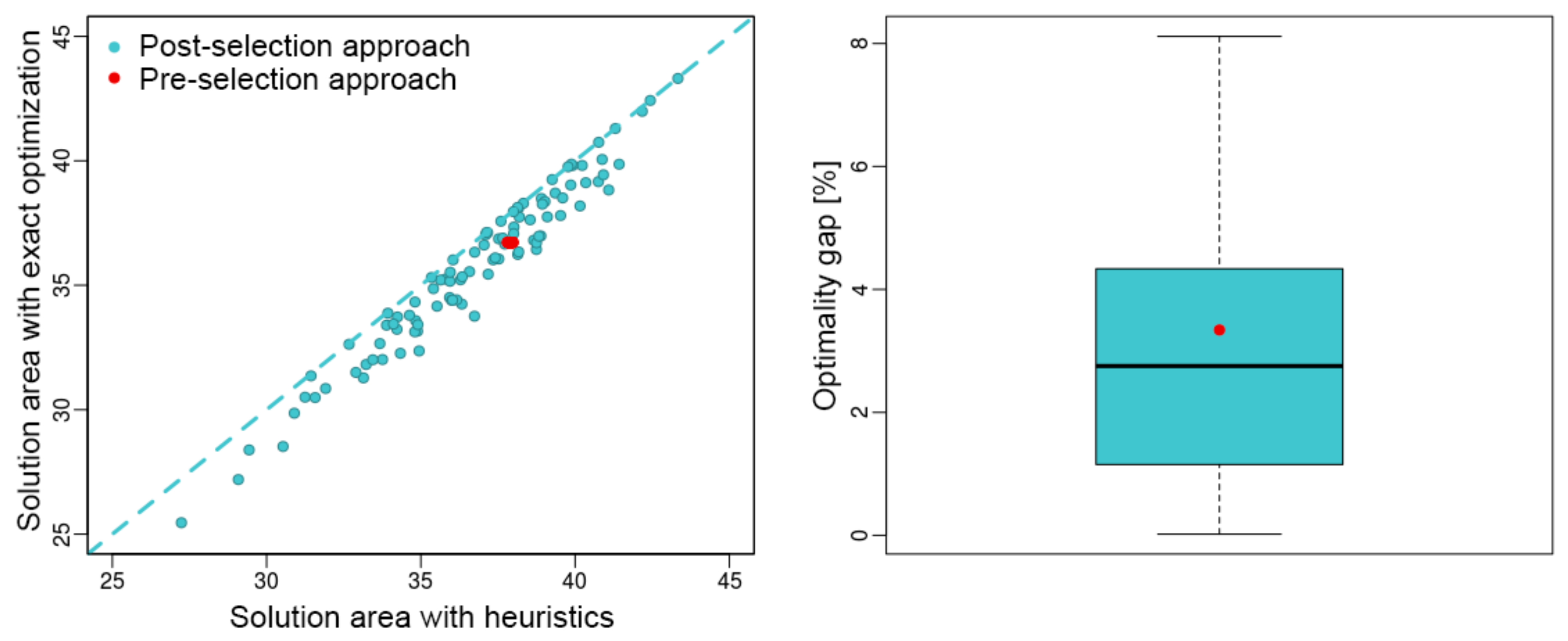
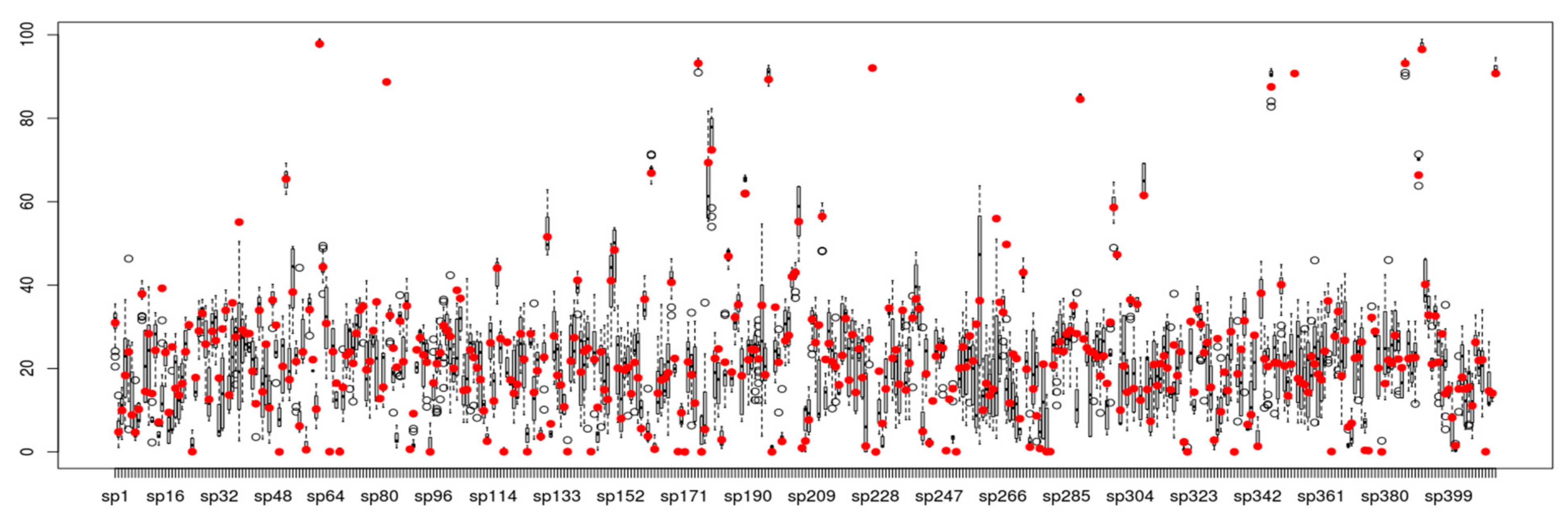
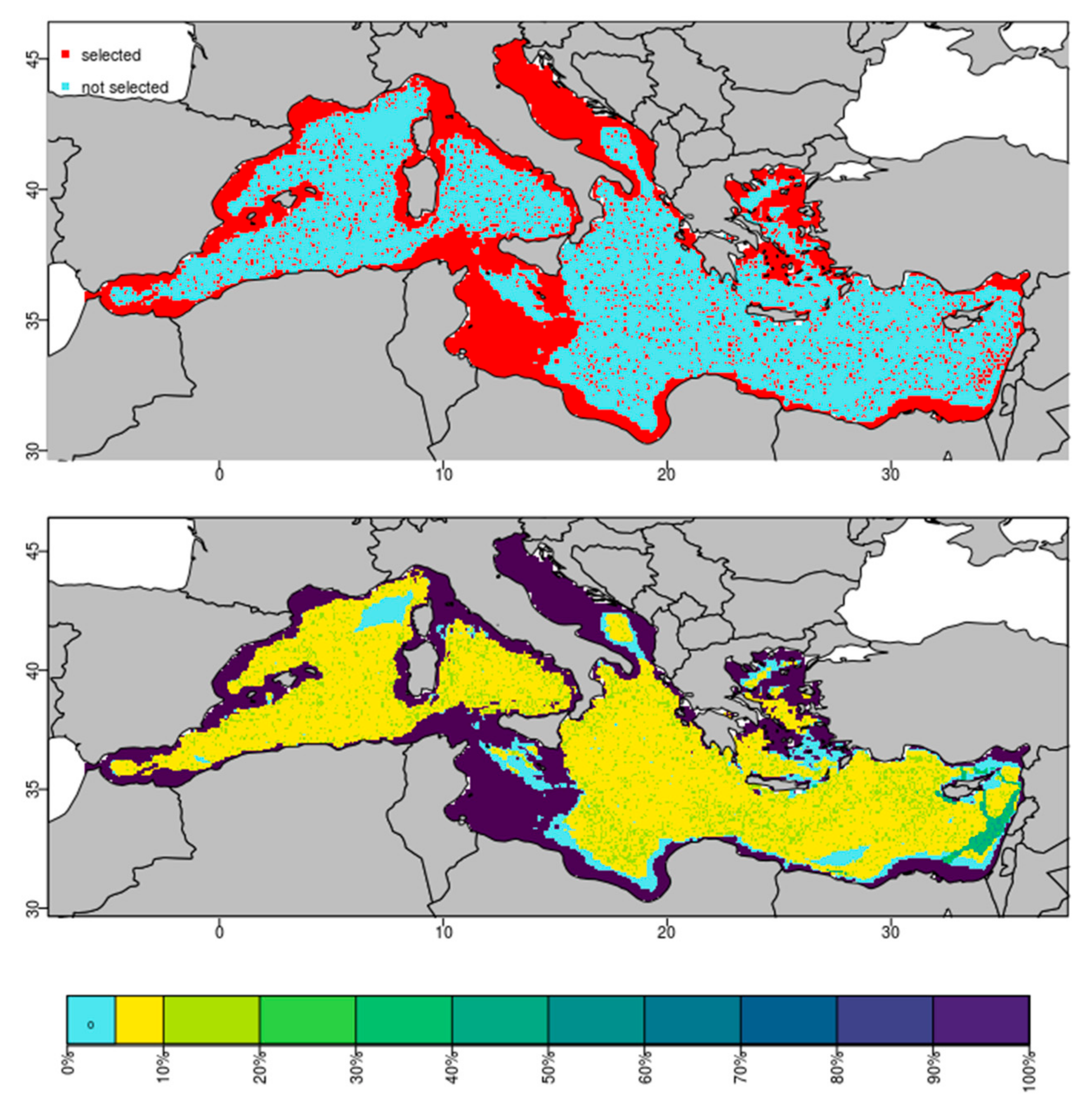
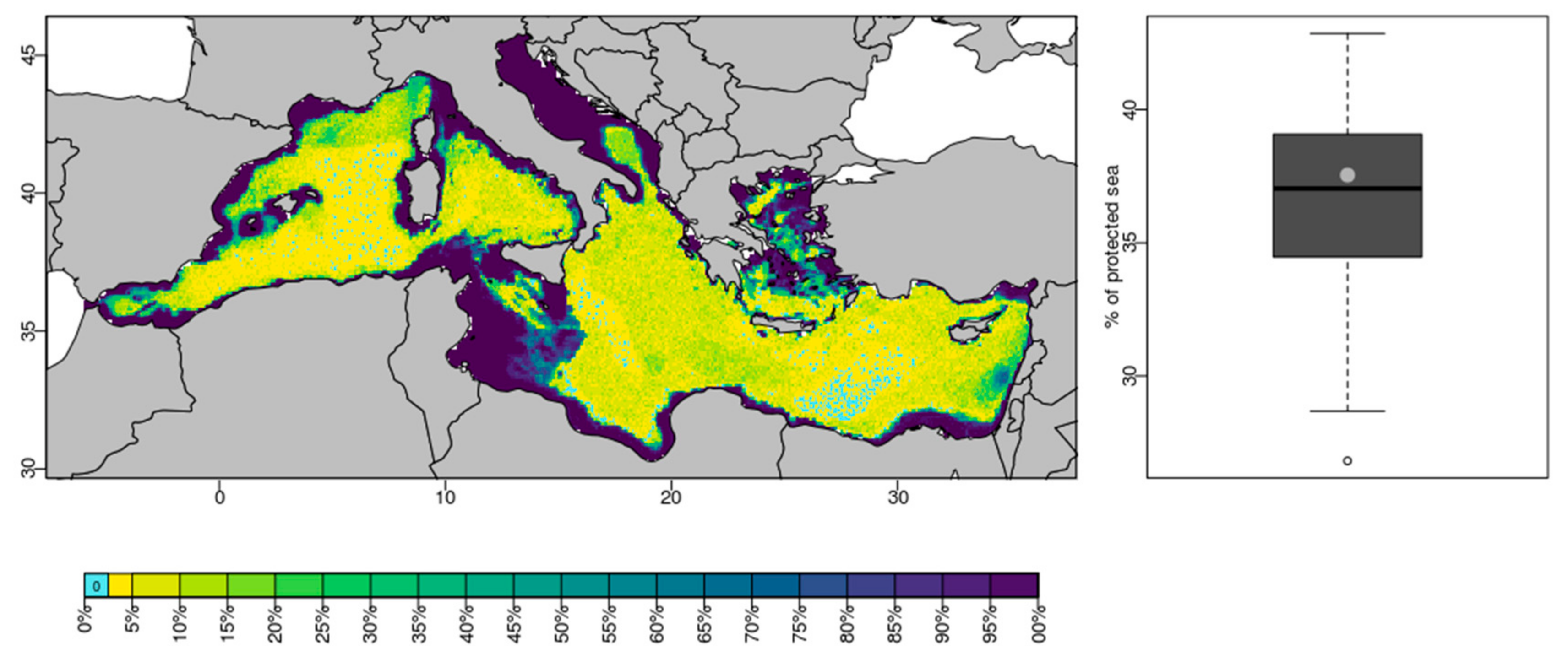
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A




References
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59. [Google Scholar] [CrossRef]
- Worm, B.; Barbier, E.B.; Beaumont, N.; Duffy, J.E.; Folke, C.; Halpern, B.S.; Jackson, J.B.C.; Lotze, H.K.; Micheli, F.; Palumbi, S.R.; et al. Impacts of Biodiversity Loss on Ocean Ecosystem Services. Science 2006, 314, 787–790. [Google Scholar] [CrossRef] [PubMed]
- Margules, C.R.; Pressey, R.L. Systematic conservation planning. Nature 2000, 405, 243–253. [Google Scholar] [CrossRef]
- Possingham, H.; Wilson, K.; Andelman, S.; Vynne, C. Protected areas: Goals, limitations, and design. In Principles of Conservation Biology, 3rd ed; Sinauer Associates: Sunderland, MA, USA, 2006; pp. 507–549. [Google Scholar]
- Magris, R.A.; Pressey, R.L.; Mills, M.; Vila-Nova, D.A.; Floeter, S. Integrated conservation planning for coral reefs: Designing conservation zones for multiple conservation objectives in spatial prioritisation. Global Ecol. Conserv. 2017, 11, 53–68. [Google Scholar] [CrossRef]
- Davies, T.E.; Maxwell, S.M.; Kaschner, K.; Garilao, C.; Ban, N.C. Large marine protected areas represent biodiversity now and under climate change. Sci. Rep. 2017, 7, 1–7. [Google Scholar] [CrossRef]
- Drira, S.; Ben Rais Lasram, F.; Ben Rejeb Jenhani, A.; Shin, Y.J.; Guilhaumon, F. Species-area uncertainties impact the setting of habitat conservation targets and propagate across conservation solutions. Biol. Conserv. 2019, 235, 279–289. [Google Scholar] [CrossRef]
- Araújo, M.B.; Williams, P.H. Selecting areas for species persistence using occurrence data. Biol. Conserv. 2000, 96, 331–345. [Google Scholar] [CrossRef]
- Araújo, M.B.; Williams, P.H.; Turner, A. A sequential approach to minimise threats within selected conservation areas. Biodivers. Conserv. 2002, 11, 1011–1024. [Google Scholar] [CrossRef]
- Harnik, P.G.; Lotze, H.K.; Anderson, S.C.; Finkel, Z.V.; Finnegan, S.; Lindberg, D.R.; Liow, L.H.; Lockwood, R.; McClain, C.R.; McGuire, J.L.; et al. Extinctions in ancient and modern seas. Trends Ecol. Evol. 2012, 27, 608–617. [Google Scholar] [CrossRef]
- Rodrigues, A.S.L.; Andelman, S.J.; Bakarr, M.I.; Boitani, L.; Brooks, T.M.; Cowling, R.M.; Fishpool, L.D.C.; da Fonseca, G.A.B.; Gaston, K.J.; Hoffmann, M.; et al. Effectiveness of the global protected area network in representing species diversity. Nature 2004, 428, 640–643. [Google Scholar] [CrossRef]
- Bini, L.M.; Diniz-Filho, J.A.F.; Rangel, T.F.L.V.B.; Bastos, R.P.; Pinto, M.P. Challenging Wallacean and Linnean shortfalls: Knowledge gradients and conservation planning in a biodiversity hotspot. Divers. Distrib. 2006, 12, 475–482. [Google Scholar] [CrossRef]
- Lomolino, M. Conservation biogeography. In Frontiers of Biogeography: New Directions in the Geography of Nature, 293; Sinauer Associates, Inc.: Sunderland, MA, USA, 2004. [Google Scholar]
- Terribile, L.C.; Lima-Ribeiro, M.S.; Araujo, M.B.; Bizao, N.; Collevatt, R.G.; Dobrovolski, R.; Diniz Filho, J.A.F. Areas of climate stability of species ranges in the Brazilian Cerrado: Disentangling uncertainties through time. Nat. Conserv. 2012, 10, 152–159. [Google Scholar] [CrossRef]
- Whittaker, R.J.; Araújo, M.B.; Jepson, P.; Ladle, R.J.; Watson, J.E.; Willis, K.J. Conservation biogeography: Assessment and prospect. Divers. Distrib. 2005, 11, 3–23. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species Distribution Models: Ecological Explanation and Prediction Across Space and Time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Marmion, M.; Parviainen, M.; Luoto, M.; Heikkinen, R.K.; Thuiller, W. Evaluation of consensus methods in predictive species distribution modelling. Divers. Distrib. 2009, 15, 59–69. [Google Scholar] [CrossRef]
- Wisz, M.S.; Hijmans, R.J.; Li, J.; Peterson, A.T.; Graham, C.H.; Guisan, A. NCEAS Predicting Species Distributions Working Group†. Effects of sample size on the performance of species distribution models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- Elith, J.H.; Graham, C.P.; Anderson, R.; Dudík, M.; Ferrier, S.; Guisan, A.J.; Hijmans, R.; Huettmann, F.R.; Leathwick, J.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.E.; Elith, J.; Graham, C.H.; Phillips, S.; Peterson, A.T. What matters for predicting the occurrences of trees: Techniques, data, or species characteristics? Ecol. Monogr. 2007, 77, 615–630. [Google Scholar] [CrossRef]
- Roura-Pascual, N.; Brotons, L.; Peterson, A.T.; Thuiller, W. Consensual predictions of potential distributional areas for invasive species: A case study of Argentine ants in the Iberian Peninsula. Biol. Invasions 2009, 11, 1017–1031. [Google Scholar] [CrossRef]
- Lentini, P.E.; Wintle, B.A. Spatial conservation priorities are highly sensitive to choice of biodiversity surrogates and species distribution model type. Ecography 2015, 38, 1101–1111. [Google Scholar] [CrossRef]
- Loiselle, B.A.; Howell, C.A.; Graham, C.H.; Goerck, J.M.; Brooks, T.; Smith, K.G.; Williams, P.H. Avoiding Pitfalls of Using Species Distribution Models in Conservation Planning. Conserv. Biol. 2003, 17, 1591–1600. [Google Scholar] [CrossRef]
- Haight, R.G.; Revelle, C.S.; Snyder, S.A. An Integer Optimization Approach to a Probabilistic Reserve Site Selection Problem. Oper. Res. 2000, 48, 697–708. [Google Scholar] [CrossRef]
- Wilson, K.A.; Cabeza, M.; Klein, C.J. Fundamental concepts of spatial conservation prioritization. In Spatial Conservation Prioritization: Quantitative Methods and Computational Tools; Oxford University Press: New York, NY, USA, 2009; pp. 16–27. [Google Scholar]
- Sarkar, S. Ecological Diversity and Biodiversity as Concepts for Conservation Planning: Comments on Ricotta. Acta Biotheor. 2006, 54, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Camm, J.D.; Polasky, S.; Solow, A.; Csuti, B. A note on optimal algorithms for reserve site selection. Biol. Conserv. 1996, 78, 353–355. [Google Scholar] [CrossRef]
- Önal, H. First-best, second-best, and heuristic solutions in conservation reserve site selection. Biol. Conserv. 2004, 115, 55–62. [Google Scholar] [CrossRef]
- Underhill, L.G. Optimal and suboptimal reserve selection algorithms. Biol. Conserv. 1994, 70, 85–87. [Google Scholar] [CrossRef]
- Jantke, K.; Jones, K.R.; Allan, J.R.; Chauvenet, A.L.; Watson, J.E.; Possingham, H.P. Poor ecological representation by an expensive reserve system: Evaluating 35 years of marine protected area expansion. Conserv. Lett. 2018, 11, e12584. [Google Scholar] [CrossRef]
- Hanson, J.O.; Rhodes, J.R.; Butchart, S.H.; Buchanan, G.M.; Rondinini, C.; Ficetola, G.F.; Fuller, R.A. Global conservation of species’ niches. Nature 2020, 580, 232–234. [Google Scholar] [CrossRef] [PubMed]
- Wilhere, G.F.; Goering, M.; Wang, H. Average optimacity: An index to guide site prioritization for biodiversity conservation. Biol. Conserv. 2008, 141, 770–781. [Google Scholar] [CrossRef]
- Ball, I.R.; Possingham, H.P.; Watts, M. Marxan and relatives: Software for spatial conservation prioritisation. In Spatial Conservation Prioritisation: Quantitative Methods and Computational Tools; Oxford University Press: Oxford, UK, 2009; pp. 185–195. [Google Scholar]
- Moilanen, A. Two paths to a suboptimal solution—once more about optimality in reserve selection. Biol. Conserv. 2008, 141, 1919–1923. [Google Scholar] [CrossRef]
- Vanderkam, R.P.D.; Wiersma, Y.F.; King, D.J. Heuristic algorithms vs. linear programs for designing efficient conservation reserve networks: Evaluation of solution optimality and processing time. Biol. Conserv. 2007, 137, 349–358. [Google Scholar] [CrossRef]
- Bailey, H.; Thompson, P. Using marine mammal habitat modelling to identify priority conservation zones within a marine protected area. Mar. Ecol. Prog. Ser. 2009, 378, 279–287. [Google Scholar] [CrossRef]
- Leach, K.; Zalat, S.; Gilbert, F. Egypt’s Protected Area network under future climate change. Biol. Conserv. 2013, 159, 490–500. [Google Scholar] [CrossRef]
- Passoni, G.; Rowcliffe, J.M.; Whiteman, A.; Huber, D.; Kusak, J. Framework for strategic wind farm site prioritisation based on modelled wolf reproduction habitat in Croatia. Eur. J. Wildl. Res. 2017, 63, 38. [Google Scholar] [CrossRef]
- Walther, B.A.; Pirsig, L.H. Determining conservation priority areas for Palearctic passerine migrant birds in sub-Saharan Africa. Avian Conserv. Ecol. 2017, 12, 2. [Google Scholar] [CrossRef][Green Version]
- Zhang, L.; Liu, S.; Sun, P.; Wang, T.; Wang, G.; Zhang, X.; Wang, L. Consensus Forecasting of Species Distributions: The Effects of Niche Model Performance and Niche Properties. PLoS ONE 2015, 10, e0120056. [Google Scholar] [CrossRef] [PubMed]
- Bush, A.; Hermoso, V.; Linke, S.; Nipperess, D.; Turak, E.; Hughes, L. Freshwater conservation planning under climate change: Demonstrating proactive approaches for Australian Odonata. J. Appl. Ecol. 2014, 51, 1273–1281. [Google Scholar] [CrossRef]
- Meller, L.; Cabeza, M.; Pironon, S.; Barbet-Massin, M.; Maiorano, L.; Georges, D.; Thuiller, W. Ensemble distribution models in conservation prioritization: From consensus predictions to consensus reserve networks. Divers. Distrib. 2014, 20, 309–321. [Google Scholar] [CrossRef] [PubMed]
- Ben Rais Lasram, F.; Guilhaumon, F.; Mouillot, D. Fish diversity patterns in the Mediterranean Sea: Deviations from a mid-domain model. Mar. Ecol. Prog. Ser. 2009, 376, 253–267. [Google Scholar] [CrossRef]
- Ben Rais Lasram, F.; Guilhaumon, F.; Albouy, C.; Somot, S.; Thuiller, W.; Mouillot, D. The Mediterranean Sea as a ‘cul-de-sac’ for endemic fishes facing climate change. Glob. Change Biol. 2010, 16, 3233–3245. [Google Scholar] [CrossRef]
- Albouy, C.; Guilhaumon, F.; Leprieur, F.; Ben Rais Lasram, F.; Somot, S.; Aznar, R.; Velez, L.; Le Loc’h, F.; Mouillot, D. Projected climate change and the changing biogeography of coastal Mediterranean fishes. J. Biogeogr. 2013, 40, 534–547. [Google Scholar] [CrossRef]
- Albouy, C.; Ben Rais Lasram, F.; Velez, L.; Guilhaumon, F.; Meynard, C.N.; Boyer, S.; Mouillot, D. FishMed: Traits, phylogeny, current and projected species distribution of Mediterranean fishes, and environmental data: Ecological Archives E096-203. Ecology 2015, 96, 2312–2313. [Google Scholar] [CrossRef]
- Thuiller, W.; Georges, D.; Engler, R.; Breiner, F.; Georges, M.D.; Thuiller, C.W. Package ‘biomod2′. Species distribution modeling within an ensemble forecasting framework. Ecography 2016, 32, 369–373. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS): Assessing the accuracy of distribution models. J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Guilhaumon, F.; Albouy, C.; Claudet, J.; Velez, L.; Ben Rais Lasram, F.; Tomasini, J.-A.; Douzery, E.J.; Meynard, C.N.; Mouquet, N.; Troussellier, M.; et al. Representing taxonomic, phylogenetic and functional diversity: New challenges for Mediterranean marine-protected areas. Divers. Distrib. 2015, 21, 175–187. [Google Scholar] [CrossRef]
- Moilanen, A.; Ball, I. Heuristic and approximate optimization methods for spatial conservation prioritization. In Spatial Conservation Prioritization: Quantitative Methods and Computational Tools; Moilanen, A., Wilson, K.A., Possingham, H.P., Eds.; Oxford University Press: New York, NY, USA, 2009; pp. 58–69. [Google Scholar]
- Possingham, H.; Ball, I.; Andelman, S. Mathematical methods for identifying representative reserve networks. In Quantitative Methods for Conservation Biology; Springer: New York, NY, USA, 2000; pp. 291–306. Available online: https://link.springer.com/book/10.1007/b97704 (accessed on 24 December 2021).
- Gurobi Optimization; Gurobi Optimization. Inc.: Houston, TX, USA, 2012.
- Andelman, S.J.; Willig, M.R. Alternative configurations of conservation reserves for Paraguayan bats: Considerations of spatial scale. Conserv. Biol. 2002, 16, 1352–1363. [Google Scholar] [CrossRef]
- Tulloch, A.I.T.; Sutcliffe, P.; Naujokaitis-Lewis, I.; Tingley, R.; Brotons, L.; Ferraz, K.M.; Possingham, H.; Guisan, A.; Rhodes, J.R. Conservation planners tend to ignore improved accuracy of modelled species distributions to focus on multiple threats and ecological processes. Biol. Conserv. 2016, 199, 157–171. [Google Scholar] [CrossRef]
- Guillera-Arroita, G.; Lahoz-Monfort, J.J.; Elith, J.; Gordon, A.; Kujala, H.; Lentini, P.E.; McCarthy, M.A.; Tingley, R.; Wintle, B.A. Is my species distribution model fit for purpose? Matching data and models to applications. Glob. Ecol. Biogeogr. 2015, 24, 276–292. [Google Scholar] [CrossRef]
- Pearson, R.G.; Thuiller, W.; Araújo, M.B.; Martinez-Meyer, E.; Brotons, L.; McClean, C.; Miles, L.; Segurado, P.; Dawson, T.P.; Lees, D.C. Model-based uncertainty in species range prediction. J. Biogeogr. 2006, 33, 1704–1711. [Google Scholar] [CrossRef]
- Porfirio, L.L.; Harris, R.M.B.; Lefroy, E.C.; Hugh, S.; Gould, S.F.; Lee, G.; Bindoff, N.L.; Mackey, B. Improving the Use of Species Distribution Models in Conservation Planning and Management under Climate Change. PLoS ONE 2014, 9, e113749. [Google Scholar] [CrossRef]
- Molloy, S.; Davis, R.; Van Etten, E. An evaluation and comparison of spatial modelling applications for the management of biodiversity: A case study on the fragmented landscapes of south-Western Australia. Pac. Conserv. Biol. 2016, 22, 338–349. [Google Scholar] [CrossRef]
- Pressey, R.L.; Possingham, H.P.; Margules, C.R. Optimality in reserve selection algorithms: When does it matter and how much? Biol. Conserv. 1996, 76, 259–267. [Google Scholar] [CrossRef]
- MAPAMED, the database of MArine Protected Areas in the MEDiterranean. 2019 edition. © 2020 by SPA/RAC and MedPAN, Licensed under CC BY-NC-SA 4.0. Available online: https://www.mapamed.org/ (accessed on 18 June 2019).





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drira, S.; Ben Rais Lasram, F.; Hattab, T.; Shin, Y.-J.; Ben Rejeb Jenhani, A.; Guilhaumon, F. Can We Avoid Tacit Trade-Offs between Flexibility and Efficiency in Systematic Conservation Planning? The Mediterranean Sea as a Case Study. Diversity 2022, 14, 9. https://doi.org/10.3390/d14010009
Drira S, Ben Rais Lasram F, Hattab T, Shin Y-J, Ben Rejeb Jenhani A, Guilhaumon F. Can We Avoid Tacit Trade-Offs between Flexibility and Efficiency in Systematic Conservation Planning? The Mediterranean Sea as a Case Study. Diversity. 2022; 14(1):9. https://doi.org/10.3390/d14010009
Chicago/Turabian StyleDrira, Sabrine, Frida Ben Rais Lasram, Tarek Hattab, Yunne-Jai Shin, Amel Ben Rejeb Jenhani, and François Guilhaumon. 2022. "Can We Avoid Tacit Trade-Offs between Flexibility and Efficiency in Systematic Conservation Planning? The Mediterranean Sea as a Case Study" Diversity 14, no. 1: 9. https://doi.org/10.3390/d14010009
APA StyleDrira, S., Ben Rais Lasram, F., Hattab, T., Shin, Y.-J., Ben Rejeb Jenhani, A., & Guilhaumon, F. (2022). Can We Avoid Tacit Trade-Offs between Flexibility and Efficiency in Systematic Conservation Planning? The Mediterranean Sea as a Case Study. Diversity, 14(1), 9. https://doi.org/10.3390/d14010009