
Diversity of Macrophytes and Environmental Assessment of the Ljubljanica River (Slovenia)




Abstract
:1. Introduction
2. Materials and Methods
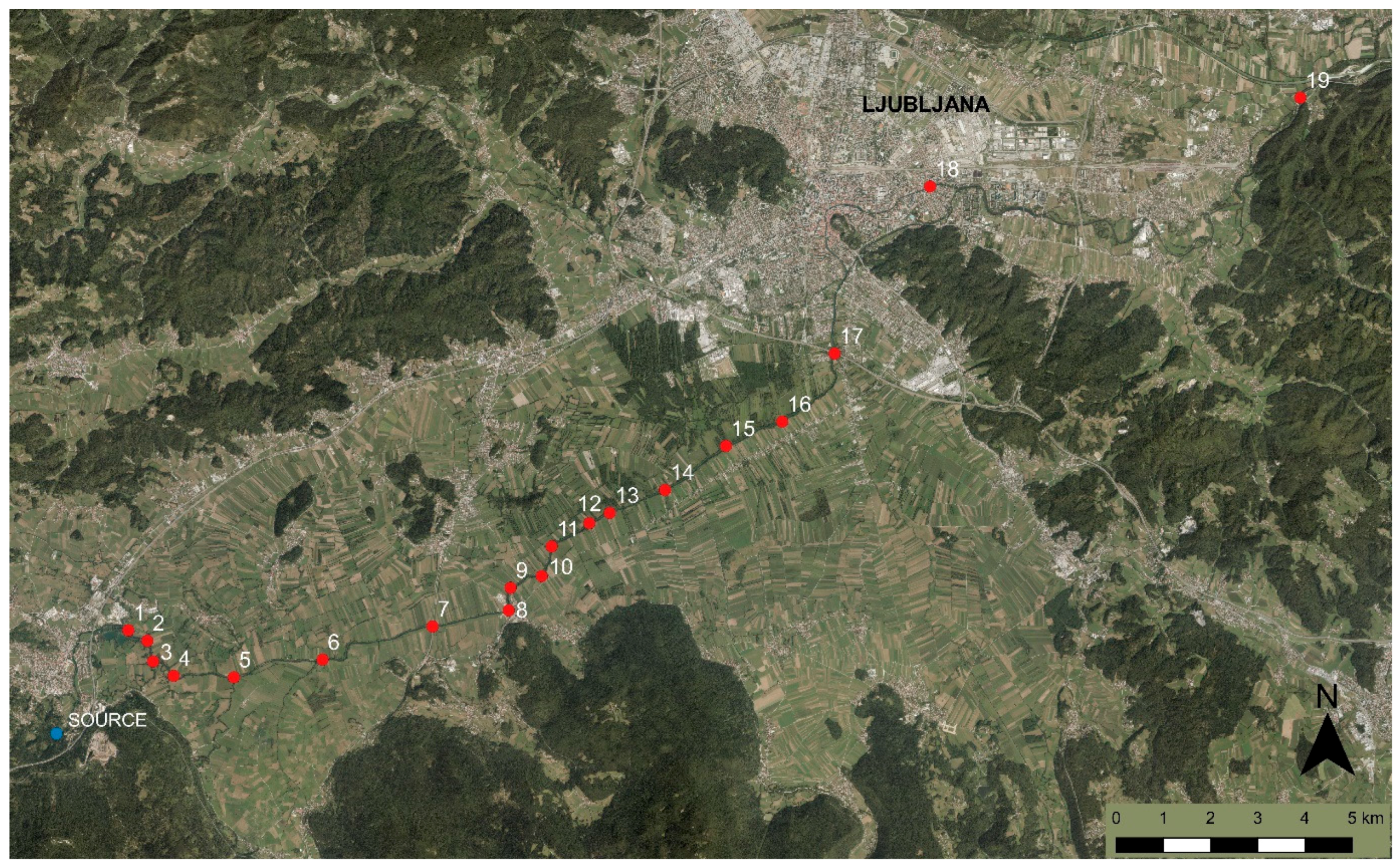
2.1. Study Area
2.2. Macrophyte Survey and Assessment of Environmental Parameters
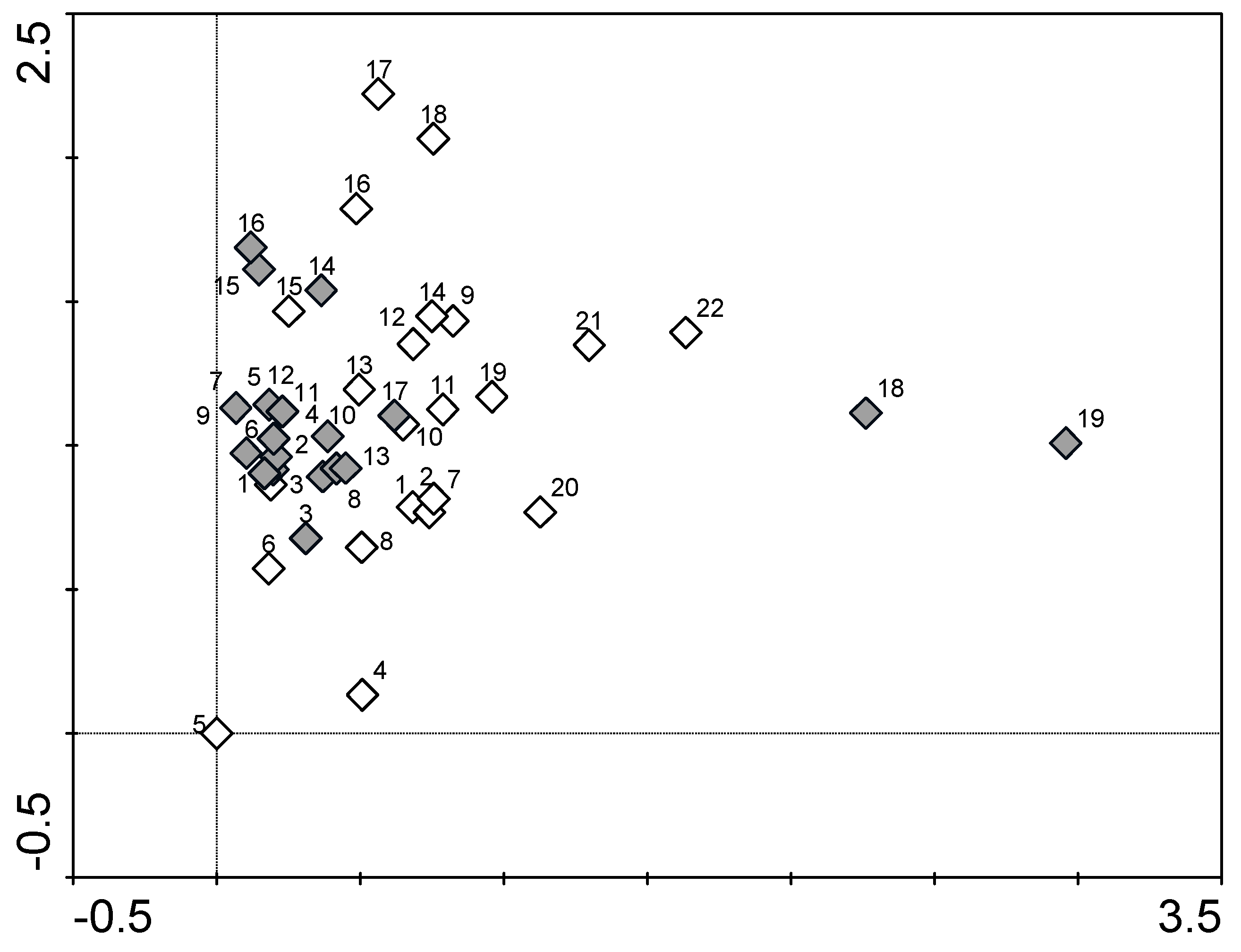
2.3. Data Analyses
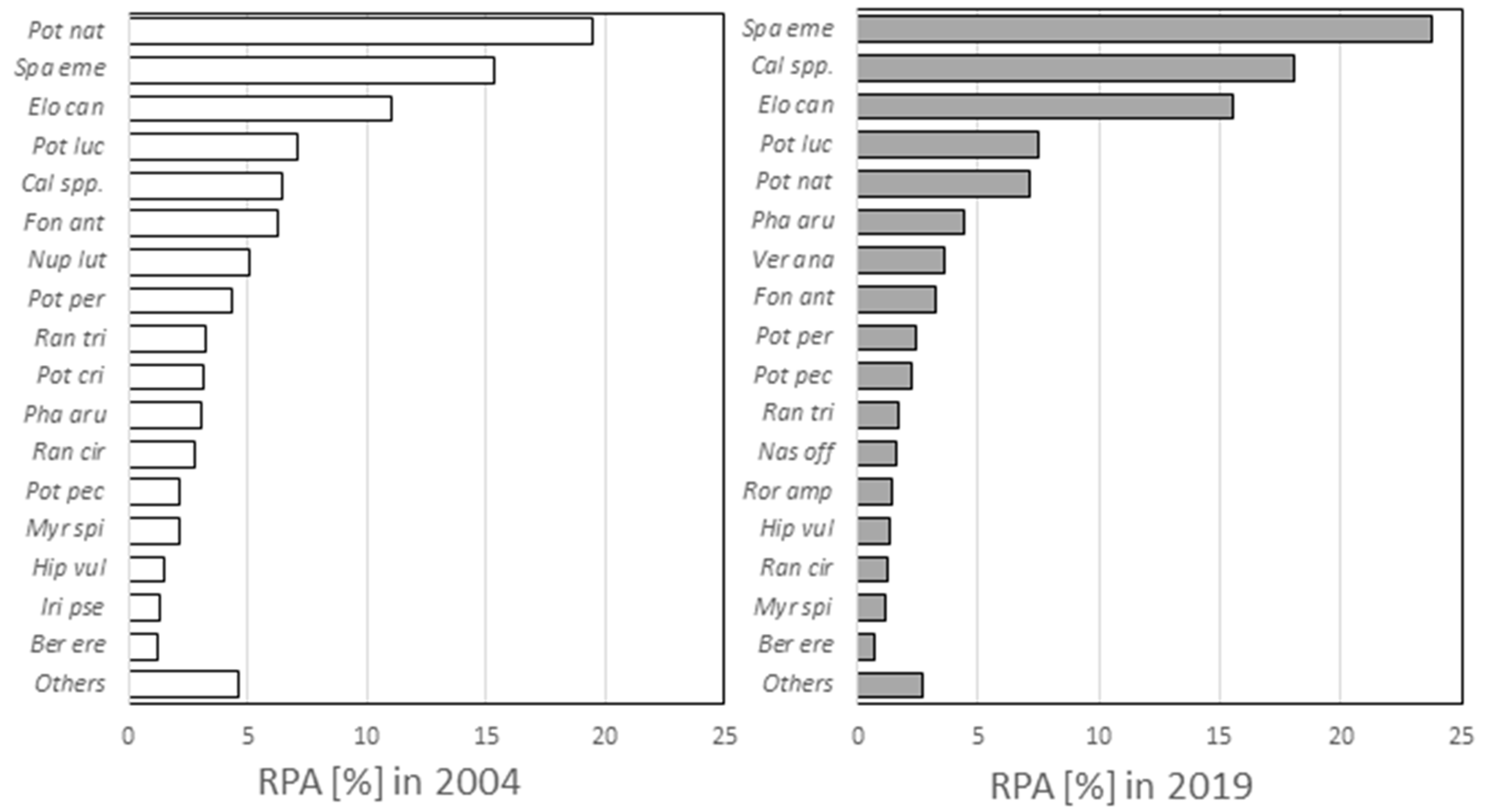
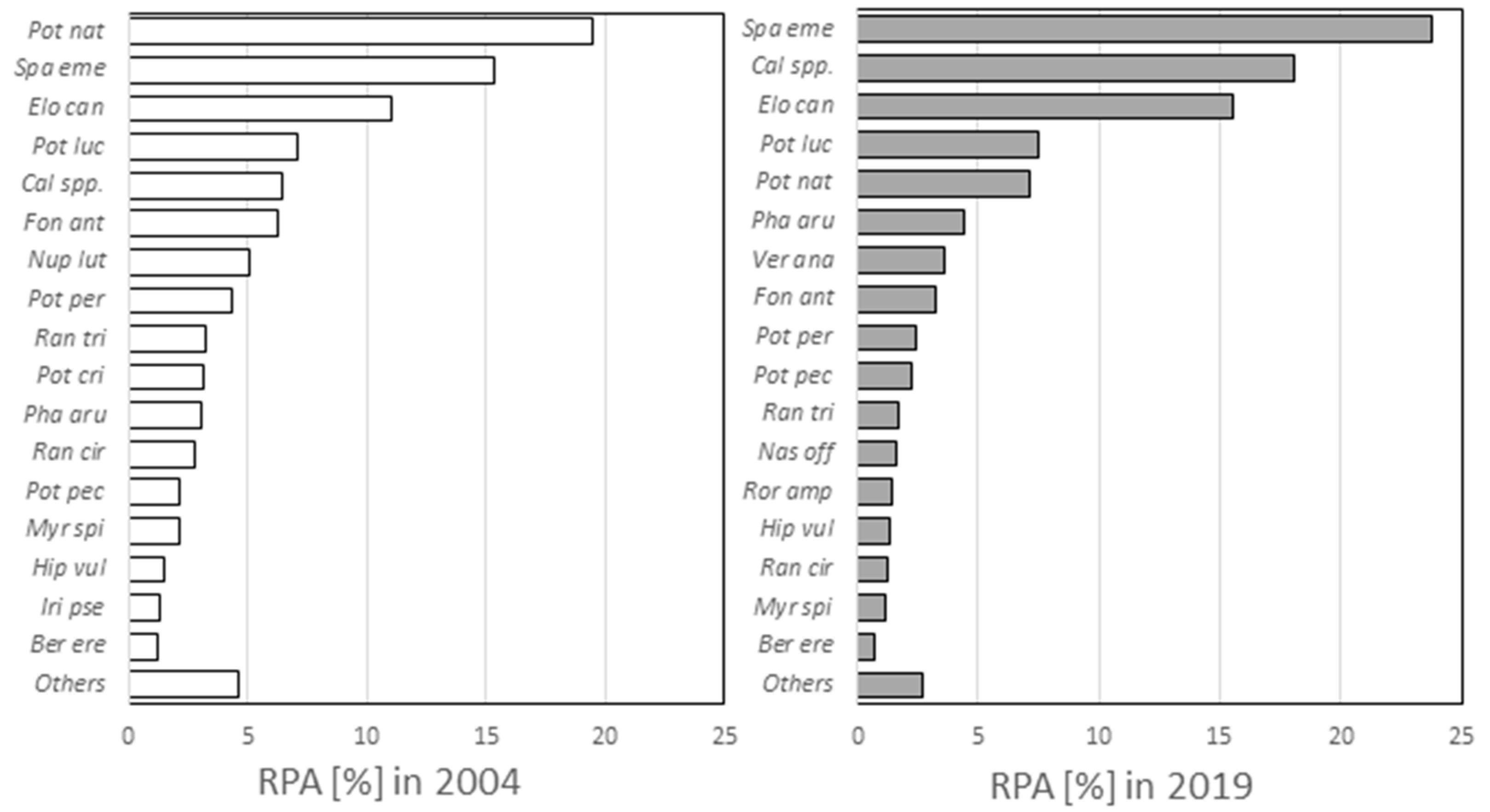
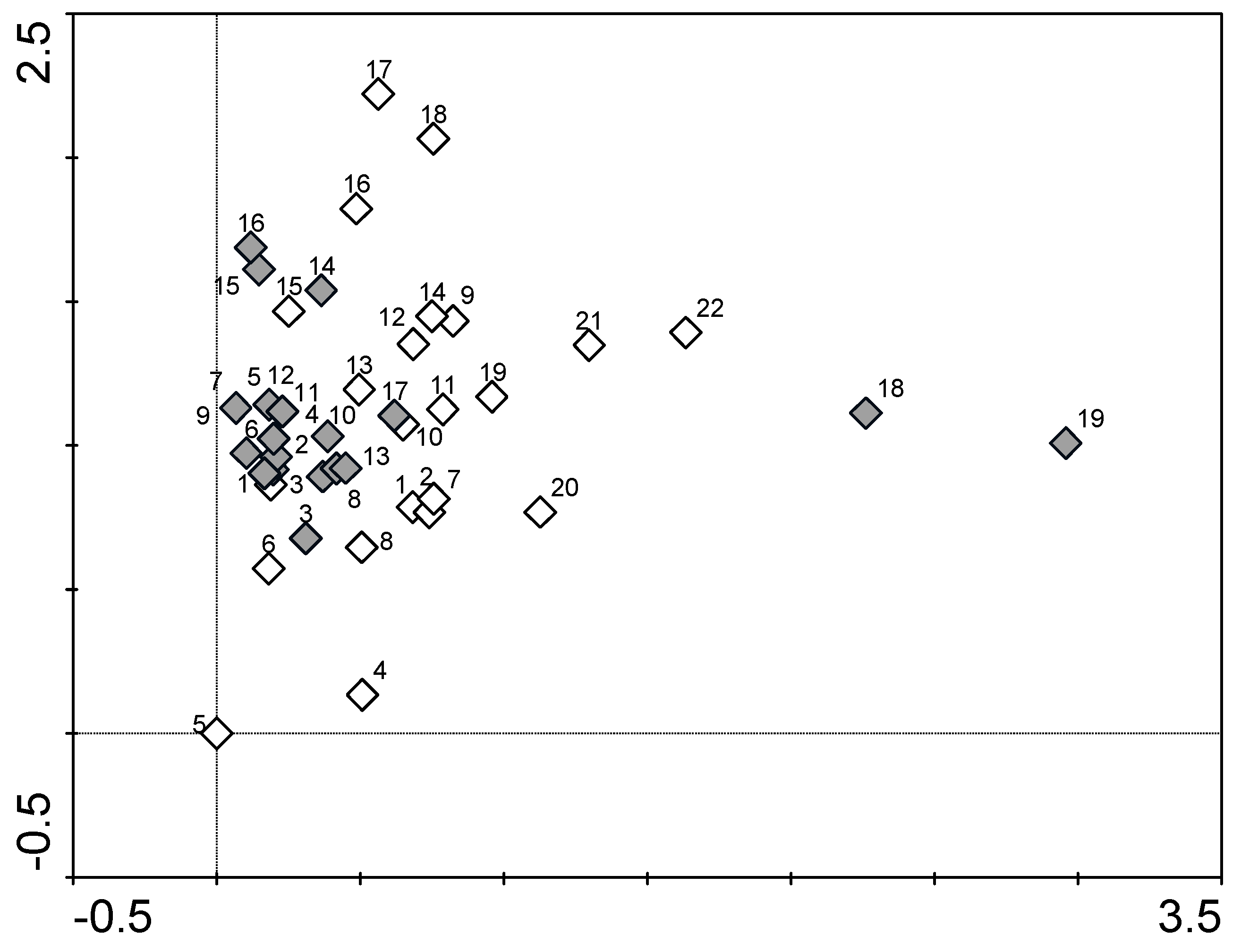
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Borgwardt, F.; Robinson, L.; Trauner, D.; Teixeira, H.; Nogueira, A.J.A.; Lillebø, A.I.; Piet, G.; Kuemmerlen, M.; O’Higgins, T.; McDonaldg, H.; et al. Exploring variability in environmental impact risk fromhuman activities across aquatic ecosystems. Sci. Total Environ. 2019, 652, 1396–1408. [Google Scholar] [CrossRef]
- Grime, J.P. Biodiversity and ecosystem function: The debate deepens. Science 1997, 277, 1260–1261. [Google Scholar] [CrossRef] [Green Version]
- Grime, J. Benefits of plant diversity to ecosystems: Immediate, filter and founder effects. J. Ecol. 1998, 86, 902–910. [Google Scholar] [CrossRef]
- Díaz, S.; Cabido, M. Vive la difference: Plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- Gallardo, B.; Clavero, M.; Sánchez, M.I.; Vilá, M. Global ecological impacts of invasive species in aquatic ecosystems. Glob. Chang. Biol. 2016, 22, 151–163. [Google Scholar] [CrossRef]
- Vilá, M.; Hulme, P.E. Non-native Species, Ecosystem Services, and Human Well-Being. In Impact of Biological Invasions on Ecosystem Services; Springer: Berlin/Heidelberg, Germany, 2017; pp. 1–14. [Google Scholar]
- Franklin, P.; Dunbar, M.; Whitehead, P. Flow controls on lowland river macrophytes: A review. Sci. Total Environ. 2008, 400, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Allan, J.D. Landscapes and riverscapes: The influence of land use on stream ecosystems. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 257–284. [Google Scholar] [CrossRef] [Green Version]
- Vörösmarty, C.J.; Mclntyre, P.B.; Gessner, M.O.; Dudgeon, D.; Prusevich, A.; Green, P.; Glidden, S.; Bunn, S.E.; Sullivan, C.A.; Liermann, C.R. Global threats to humanwater security and river biodiversity. Nature 2010, 467, 555. [Google Scholar] [CrossRef]
- Szoszkiewicz, K.; Ferreira, T.; Korte, T.; Baattrup-Pedersen, A.; Davy-Bowke, J.; O’Hare, M. European River Plant Communities: The Importance of Organic Pollution and the Usefulness of Existing Macrophyte Metrics. Hydrobiologia 2006, 566, 211–234. [Google Scholar] [CrossRef]
- Zelnik, I.; Kuhar, U.; Holcar, M.; Germ, M.; Gaberščik, A. Distribution of vascular plant communities in Slovenian watercourses. Water 2021, 13, 1071. [Google Scholar] [CrossRef]
- Fennessy, M.S.; Geho, R.; Elifritz, B.; Lopez, R.D. Testing the Floristic Quality Assessment Index as an Indicator of Riparian Wetland Disturbance; Ohio Environmental Protection Agency, Division of Surface Water: Columbus, OH, USA, 1998.
- Mack, J.J.; Micacchion, M.; Augusta, L.D.; Sablack, G.R. Vegetation Indices of Biotic Integrity (Vibi) for Wetlands and Calibration of the Ohio Rapid Assessment Method for Wetlands; Grant CD95276; Final Report to US Environmental Protection Agency; Ohio Environmental Protection Agency: Columbus, OH, USA, 2000; p. 98.
- Aznar, J.; Dervieux, A.; Grillas, P. Association between aquatic vegetation and landscape indicators of human pressure. Wetlands 2002, 23, 149–160. [Google Scholar] [CrossRef]
- Bornette, G.; Amoros, C.; Chessel, D. Effect of allogenic processes on successional rates in former river channels. J. Veg. Sci. 1994, 5, 237–246. [Google Scholar] [CrossRef]
- O’Hare, M.T.; Baattrup-Pedersen, A.; Nijboer, R.; Szoszkiewicz, K.; Ferreira, T. Macrophyte communities of European streams with altered physical habitat. Hydrobiologia 2006, 566, 197–210. [Google Scholar] [CrossRef] [Green Version]
- Yang, N.; Zhang, Y.; Duan, K. Effect of Hydrologic Alteration on the Community Succession of Macrophytes at Xiangyang Site, Hanjiang River, China. Scientifica 2017, 2017, 4083696. [Google Scholar] [CrossRef] [PubMed]
- Mackay, S.J.; Arthington, A.H.; Kennard, M.J.; Pusey, B.J. Spatial variation in the distribution and abundance of submerged macrophytes in an Australian subtropical river. Aquat. Bot. 2003, 77, 169–186. [Google Scholar] [CrossRef] [Green Version]
- Kuhar, U.; Germ, M.; Gaberščik, A.; Urbanič, G. Development of a River Macrophyte Index (RMI) for assessing river ecological status. Limnologica 2011, 41, 235–243. [Google Scholar] [CrossRef]
- Hatton-Ellis, T.W.; Grieve, N.; Newman, J. Ecology of watercourses characterised by Ranunculion fluitantis and Callitricho-Batrachion vegetation. Conserving Natura 2000 rivers ecology series. Engl. Nat. 2003, 11, 67. [Google Scholar]
- Halabowski, D.; Lewin, I. Impact of anthropogenic transformations on the vegetation of selected abiotic types of rivers in two ecoregions (Southern Poland). Knowl. Manag. Aquat. Ecosyst. 2020, 421, 35. [Google Scholar] [CrossRef]
- Lindholm, M.; Alahuhta, J.; Heino, J.; Hjort, J.; Toivonen, H. Changes in the functional features of macrophyte communities and driving factors across a 70-year period. Hydrobiologia 2020, 847, 3811–3827. [Google Scholar] [CrossRef] [Green Version]
- Sand-Jensen, K.; Jeppesen, E.; Nielsen, K.; Van Der Bijl, L.; Hjermind, L.; Nielsen, L.W.; Iversen, T.M. Growth of macrophytes and ecosystem consequences in a lowland Danish stream. Freshw. Biol. 1989, 22, 15–32. [Google Scholar] [CrossRef]
- Milner, A.M. System recovery. In The Rivers Handbook: Hydrological and Ecological Principles; Calow, P., Petts, G.E., Eds.; Blackwell Science Ltd.: Oxford, UK, 1994; Volume 2, pp. 76–97. [Google Scholar]
- Underwood, A.J. Spatial and temporal problems with monitoring. In The Rivers Handbook: Hydrological and Ecological Principles; Calow, P., Petts, G.E., Eds.; Blackwell Science Ltd.: Oxford, UK, 1994; Volume 2, pp. 101–123. [Google Scholar]
- Viciani, D.; Vidali, M.; Gigante, D.; Bolpagni, R.; Villani, M.; Acosta, A.T.R.; Adorni, M.; Aleffi, M.; Allegrezza, M.; Angiolini, C.; et al. A first checklist of the alien-dominated vegetation in Italy. Plant Sociol. 2020, 57, 29–54. [Google Scholar] [CrossRef]
- Zelnik, I.; Mavrič-Klenovšek, V.; Gaberščik, A. Complex undisturbed riparian zones are resistant to colonisation by invasive alien plant species. Water 2020, 12, 345. [Google Scholar] [CrossRef] [Green Version]
- Directive 2000/60/EC of the European Parlament and of the Council of 23 October 2000. Available online: https://eur-lex.europa.eu/eli/dir/2000/60/oj (accessed on 8 October 2020).
- Feio, M.J.; Aguiar, F.C.; Almeida, S.F.P.; Ferreira, J.; Ferreira, M.T.; Elias, C.; Serra, S.R.Q.; Buffagni, A.; Cambra, J.; Chauvin, C.; et al. Least Disturbed Condition for European Mediterranean rivers. Sci. Total Environ. 2014, 476–477, 745–756. [Google Scholar] [CrossRef]
- Urbanc-Berčič, O.; Germ, M.; Povž, M.; Šumer, S. Evaluation of Ecological Status of the River Ljubljanica: Macrophytes and Fishes; Nacionalni Inštitut za Biologijo: Ljubljana, Slovenia, 2007; 47p. [Google Scholar]
- Pavšič, J. The Ljubljansko Barje Marshes; Društvo Slovenska Matica: Ljubljana, Slovenia, 2008. [Google Scholar]
- Kohler, A. Methoden der kartierung von flora und vegetation von sußwasserbiotopen. Landsch. Stadt 1978, 10, 78–85. [Google Scholar]
- Schneider, S.; Melzer, A. The Trophic Index of Macrophytes (TIM)—A New Tool for Indicating the Trophic State of Running Waters. Int. Rev. Hydrobiol. 2003, 88, 49–67. [Google Scholar] [CrossRef]
- Petersen, R.C. The RCE: A riparian, channel, and environmental inventory for small streams in the agricultural landscape. Freshw. Biol. 1992, 27, 295–306. [Google Scholar] [CrossRef]
- Pall, K.; Janauer, G.A. Die Makrophytenvegetationvon Flußstauen am Beispiel der Donau zwischen Fluß-km2552,0 und 2511,8 in der Bundesrepublik Deutschland. Arch. Hydrobiol. Suppl. Large Rivers 1995, 9, 91–109. [Google Scholar]
- Braun-Blanquet, J. Pflanzensoziologie, Grundzüge der Vegetationskunde, 3rd ed.; Springer: Berlin, Germany, 1964; p. 631. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- ter Braak, C.J.F.; Smilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5); Microcomputer Power: Ithaca, NY, USA, 2002. [Google Scholar]
- ter Braak, C.J.F.; Verdonschot, P.F.M. Canonical correspondence analysis and related multivariate methods in aquatic ecology. Aquat. Sci. 1995, 57, 255–289. [Google Scholar] [CrossRef]
- Šraj-Kržič, N.; Germ, M.; Urbanc-Berčič, O.; Kuhar, U.; Janauer, G.A.; Gaberščik, A. The quality of the aquatic environment and macrophytes of karstic watercourses. Plant Ecol. 2007, 192, 107–118. [Google Scholar] [CrossRef]
- Lacoul, P.; Freedman, B. Environmental influences on aquatic plants in freshwater ecosystems. Environ. Rev. 2006, 14, 89–136. [Google Scholar] [CrossRef]
- Ljevnaić-Mašić, B.; Džigurski, D.; Nikolić, L.; Brdar-Jokanović, M.; Čabilovski, R.; Ćirić, V.; Petrović, A. Assessment of the habitat conditions of a rare and endangered inland saline wetland community with Bolboschoenus maritimus (L.) Palla dominance in Southeastern Europe: The effects of physical–chemical water and soil properties. Wetl. Ecol. Manag. 2020, 28, 421–438. [Google Scholar] [CrossRef]
- Svitok, M.; Hrivnák, R.; Kochjarová, J.; Oťaheľová, H.; Paľove-Balang, P. Environmental thresholds and predictors of macrophytespecies richness in aquatic habitats in central Europe. Folia Geobot. 2016, 51, 227–238. [Google Scholar] [CrossRef]
- Rosso, J.J.; Fernández Cirelli, A. Effects of land use on environmental conditions and macrophytes in prairie lotic ecosystems. Limnologica 2013, 43, 18–26. [Google Scholar] [CrossRef]
- Alahuhta, J.; Kanninen, A.; Hellsten, S.; Vuori, K.-M.; Kuoppala, M.; Hämäläinen, H. Variable response of functional macrophyte groups to lake characteristics, land use, and space: Implications for bioassessment. Hydrobiologia 2014, 737, 201–214. [Google Scholar] [CrossRef]
- Murphy, K.; Efremov, A.; Davidson, T.A.; Molina-Navarro, K.E.; Fidanza, T.C.; Crivelari Betiol, P. Chambers, Julissa Tapia Grimaldo, Sara Varandas Martins, Irina Springuel, Michael Kennedy, Roger Paulo Mormul, Eric Dibble, Deborah Hofstra, Balázs András Lukács, Daniel Gebler, Lars Baastrup-Spohr, Jonathan Urrutia-Estrada. World distribution, diversity and endemism of aquatic macrophytes. Aquat. Bot. 2019, 158, 103127. [Google Scholar]
- Preston, C. Pondweeds of Great Britain and Ireland; Botanical Society of the British Isles: London, UK, 1995; p. 352. [Google Scholar]
- Haslam, S.M. River plants of western europe. In The Macrophytic Vegetation of Watercourses of the European Economic Community; Cambridge University Press: Cambridge, UK, 1987; p. 511. [Google Scholar]
- Schneider, S. Macrophyte trophic indicator values from a European perspective. Limnol. Ecol. Manag. Inland Waters 2007, 37, 281–289. [Google Scholar] [CrossRef] [Green Version]
- Patten, B.C., Jr. The status of some American species of Myriophyllum as revealed by the discovery of intergrade material between M. exalbescens Fern. and M. spicatum L. in New Jersey. Rhodora 1954, 56, 213–225. [Google Scholar]
- Aiken, S.G.; Newroth, P.R.; Wile, I. The biology of Canadian weeds: 34. Myriophyllum spicatum L. Can. J. Plant Sci. 1979, 59, 201–215. [Google Scholar] [CrossRef]
- Hussner, A. Alien aquatic plants in European countries. Weed Res. 2012, 52, 297–306. [Google Scholar] [CrossRef]
- Hussner, A.; Hoelken, H.P.; Jahns, P. Low Light acclimated submerged freshwater plants show a pronounced sensitivity to increasing irradiances. Aquat. Bot. 2010, 93, 17–24. [Google Scholar] [CrossRef]
- Mjelde, M.; Lombardo, P.; Berge, D.; Johansen, W.S. Mass invasion of non-native Elodea canadensis Michx. In a large, clear-water, species-rich Norwegian lake—Impact on macrophyte biodiversity. Ann. Limnol. Int. J. Limnol. 2012, 48, 225–240. [Google Scholar] [CrossRef] [Green Version]
- Josefsson, M.; Andersson, B. The environmental consequences of alien species in the Swedish lakes Mälaren, Hjälmaren, Vänern and Vättern. Ambio 2001, 30, 514–521. [Google Scholar] [CrossRef]
- Kuhar, U.; Germ, M.; Gaberščik, A. Habitat characteristic of an alien species Elodea canadensis in Slovenian watercourses. Hydrobiologia 2010, 656, 205–212. [Google Scholar] [CrossRef]
- Kłosowski, G.; Kłosowski, S. Flora Polski. Rośliny Wodne i Bagienne; Multico: Warszawa, Poland, 2015; p. 336. [Google Scholar]
- Szoszkiewicz, K.; Zbierska, J.; Jusik, S.; Zgoła, T. Macrophyte Method for River Assessment. A Methodological Manual on the Assessment and Classification of the Ecological Status of Flowing Waters Based on Aquatic Plants; Bogucki Wydawnictwo Naukowe: Poznań, Poland, 2010; pp. 1–78. [Google Scholar]
- Haury, J.; Peter, M.C.; Termolieres, M.; Barbe, J.; Thiebaut, G.; Bernez, I.; Daniel, H.; Chatenet, P.; Haane-Archipof, G.; Muller, S.; et al. A new method to access water trophy and organic pollution—The Macrophite Biological Index for Rivers (IBMR): Its application to different types of river and pollution. Hydrobiologia 2006, 1, 153–158. [Google Scholar]
- Lazzaro, L.; Bolpagni, R.; Buffa, G.; Gentili, R.; Lonati, M.; Stinca, A.; Acosta, A.T.R.; Adorni, M.; Aleffi, M.; Allegrezza, M.; et al. Impact of invasive alien plants on native plant communities and Natura 2000 habitats: State of the art, gap analysis and perspectives in Italy. J. Environ. Manag. 2020, 274, 111140. [Google Scholar] [CrossRef]
- Cook, C.D.; Nicholls, M.S. A monographic study of the genus Sparganium (Sparganiaceae). Part 1. Subgenus Xanthosparganium Holmberg. Bot. Helv. 1986, 96, 213–267. [Google Scholar]
- Ceska, O.; Ceska, A. Myriophyllum haloragaceae L. species in British Columbia: Problems with identification. In Proceedings of the First International Symposium on Watermilfoil, Vancouver, BC, Canada, 23–24 July 1985. [Google Scholar]
- Nikolić, L.; Pajević, S.; Ljevnaić, B. Primary production dynamics of dominant hydrophytes in Lake Provala (Serbia). Cent. Eur. J. Biol. 2009, 4, 250–257. [Google Scholar] [CrossRef]
- Brundu, G.; Stinca, A.; Angius, L.; Bonanomi, G.; Celesti-Grapow, L.; D’Auria, G.; Griffo, R.; Migliozzi, A.; Motti, R.; Spigno, P. Pistia stratiotes L. and Eichhornia crassipes (Mart.) Solms.: Emerging invasive alien hydrophytes in Campania and Sardinia (Italy). Bull. OEPP/EPPO Bull. 2012, 42, 568–579. [Google Scholar] [CrossRef]
- Ali, M.M.; Hassan, S.A.; Shaheen, A.-S.M. Impact of riparian trees shade on aquatic plant abundance in conservation islands. Acta Bot. Croat. 2011, 70, 245–258. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Recorded in 2019 | Species Recorded in 2004 [40] | |
|---|---|---|
| 1 | Alisma plantago-aquatica | Alisma plantago-aquatica |
| 2 | Alisma sp. | |
| 3 | Berula erecta | Berula erecta |
| 4 | Callitriche sp. | Callitriche sp. |
| 5 | Carex sp. | Carex sp. |
| Ceratophyllum demersum | ||
| Chara sp. | ||
| 6 | Elodea canadensis | Elodea canadensis |
| 7 | Equisetum palustre | |
| 8 | Fontinalis antipyretica | Fontinalis antipyretica |
| 9 | Hippuris vulgaris | Hippuris vulgaris |
| Hottonia palustris | ||
| 10 | Iris pseudacorus | Iris pseudacorus |
| 11 | Juncus effusus | |
| Lemna minor | ||
| Lemna trisulca | ||
| 12 | Lycopus europaeus | Lycopus europaeus |
| Lysimachia vulgaris | ||
| 13 | Lythrum salicaria | Lythrum salicaria |
| 14 | Mentha aquatica | Mentha aquatica |
| 15 | Mentha longifolia | |
| 16 | Myriophyllum spicatum | Myriophyllum spicatum |
| Myriophyllum verticillatum | ||
| 17 | Myosotis scorpioides | Myosotis scorpioides |
| 18 | Nasturtium officinale | Nasturtium officinale |
| 19 | Nuphar luteum | Nuphar luteum |
| 20 | Phalaris arundinacea | Phalaris arundinacea |
| 21 | Phragmites australis | Phragmites australis |
| 22 | Polygonum amphibium | |
| 23 | Potamogeton crispus | Potamogeton crispus |
| 24 | Potamogeton lucens | Potamogeton lucens |
| 25 | Potamogeton natans | Potamogeton natans |
| 26 | Potamogeton pectinatus | Potamogeton pectinatus |
| 27 | Potamogeton perfoliatus | Potamogeton perfoliatus |
| 28 | Ranunculus circinatus | Ranunculus circinatus |
| 29 | Ranunculus trichophyllus | Ranunculus trichophyllus |
| Potamogeton filiformis | ||
| 30 | Rorippa sylvestris | Rorippa amphibia |
| 31 | Sagittaria sagittifolia | Sagittaria sagittifolia |
| 32 | Senecio paludosus | |
| 33 | Sparganium emersum | Sparganium emersum |
| Typha angustifolia | ||
| 34 | Veronica anagallis-aquatica | Veronica anagallis-aquatica |
| 34 | 37 |
| Taxa_S | Shannon_H | |
|---|---|---|
| pH | −0.3325 * | ns |
| conductivity | ns | ns |
| [O2] | ns | ns |
| water temperature | ns | ns |
| saturation (O2) | ns | ns |
| distance from the source | −0.3306 * | ns |
| land use | 0.3940 * | 0.3428 * |
| width of RZ | ns | ns |
| connectivity of RZ | ns | ns |
| vegetation structure of RZ | ns | ns |
| retention structures | ns | ns |
| shape of the channel | ns | ns |
| sediments in the channnel | ns | ns |
| bank structure | ns | ns |
| bottom of river | ns | ns |
| riffles, pools | 0.3411 * | 0.3774 * |
| detritus | ns | ns |
| Variable | LambdaA | p | % of TVE |
|---|---|---|---|
| retention structures | 0.55 | 0.002 | 34.9 |
| conectivity of RZ | 0.13 | 0.006 | 8.2 |
| distance from source | 0.13 | 0.004 | 8.2 |
| channel shape | 0.11 | 0.022 | 7.0 |
| sediments in the channnel | 0.1 | 0.036 | 6.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Germ, M.; Janež, V.; Gaberščik, A.; Zelnik, I. Diversity of Macrophytes and Environmental Assessment of the Ljubljanica River (Slovenia). Diversity 2021, 13, 278. https://doi.org/10.3390/d13060278
Germ M, Janež V, Gaberščik A, Zelnik I. Diversity of Macrophytes and Environmental Assessment of the Ljubljanica River (Slovenia). Diversity. 2021; 13(6):278. https://doi.org/10.3390/d13060278
Chicago/Turabian StyleGerm, Mateja, Vanja Janež, Alenka Gaberščik, and Igor Zelnik. 2021. "Diversity of Macrophytes and Environmental Assessment of the Ljubljanica River (Slovenia)" Diversity 13, no. 6: 278. https://doi.org/10.3390/d13060278
APA StyleGerm, M., Janež, V., Gaberščik, A., & Zelnik, I. (2021). Diversity of Macrophytes and Environmental Assessment of the Ljubljanica River (Slovenia). Diversity, 13(6), 278. https://doi.org/10.3390/d13060278