
Genetic, but Not Behavioral, Evidence Supports the Distinctiveness of the Mealy Amazon Parrot in the Brazilian Atlantic Forest


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
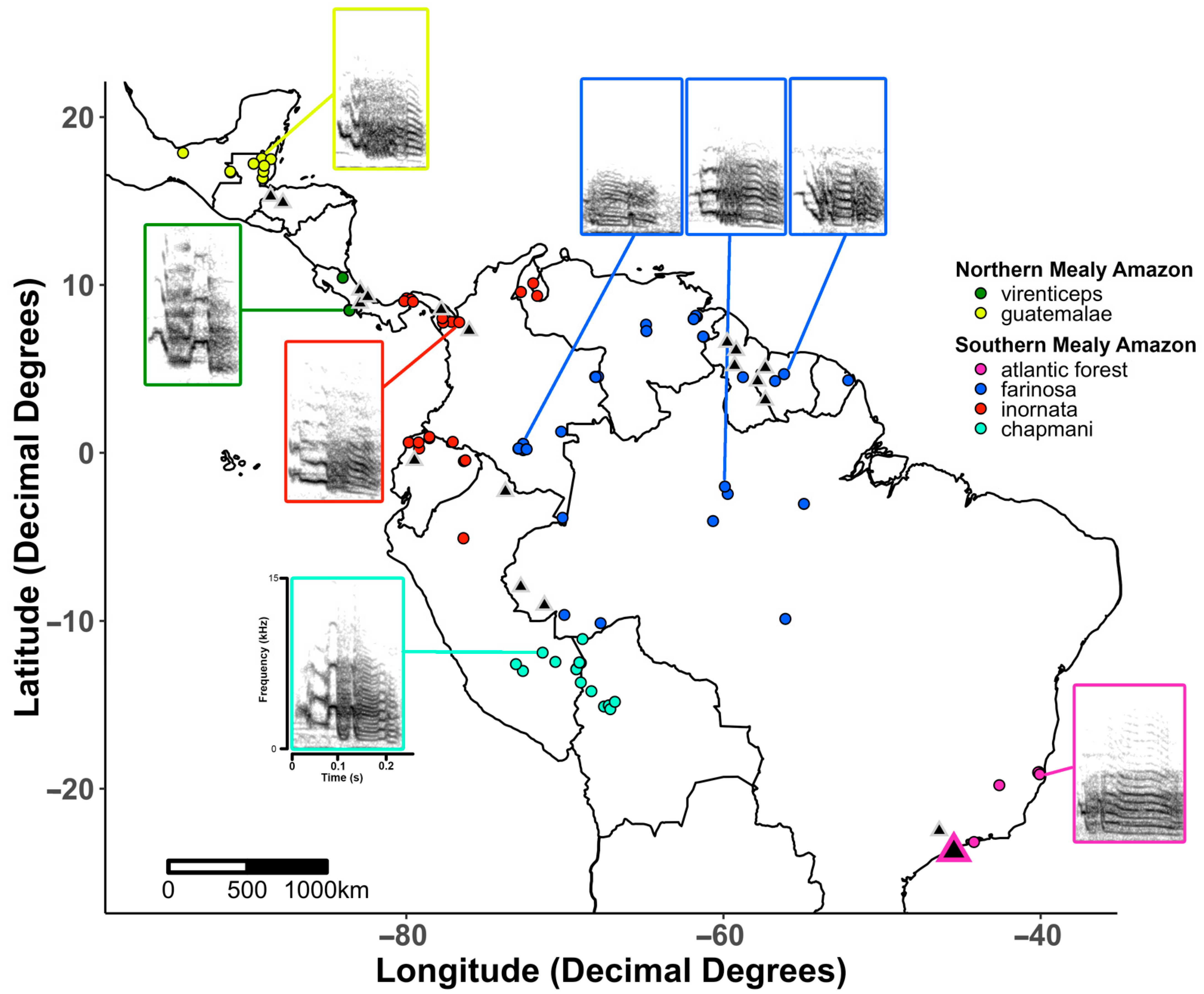
2.1. Study Species and Data Summary
2.2. Phylogenetic Analysis and Haplotype Network
2.3. Genetic Differentiation
2.4. Acoustic Analysis
3. Results
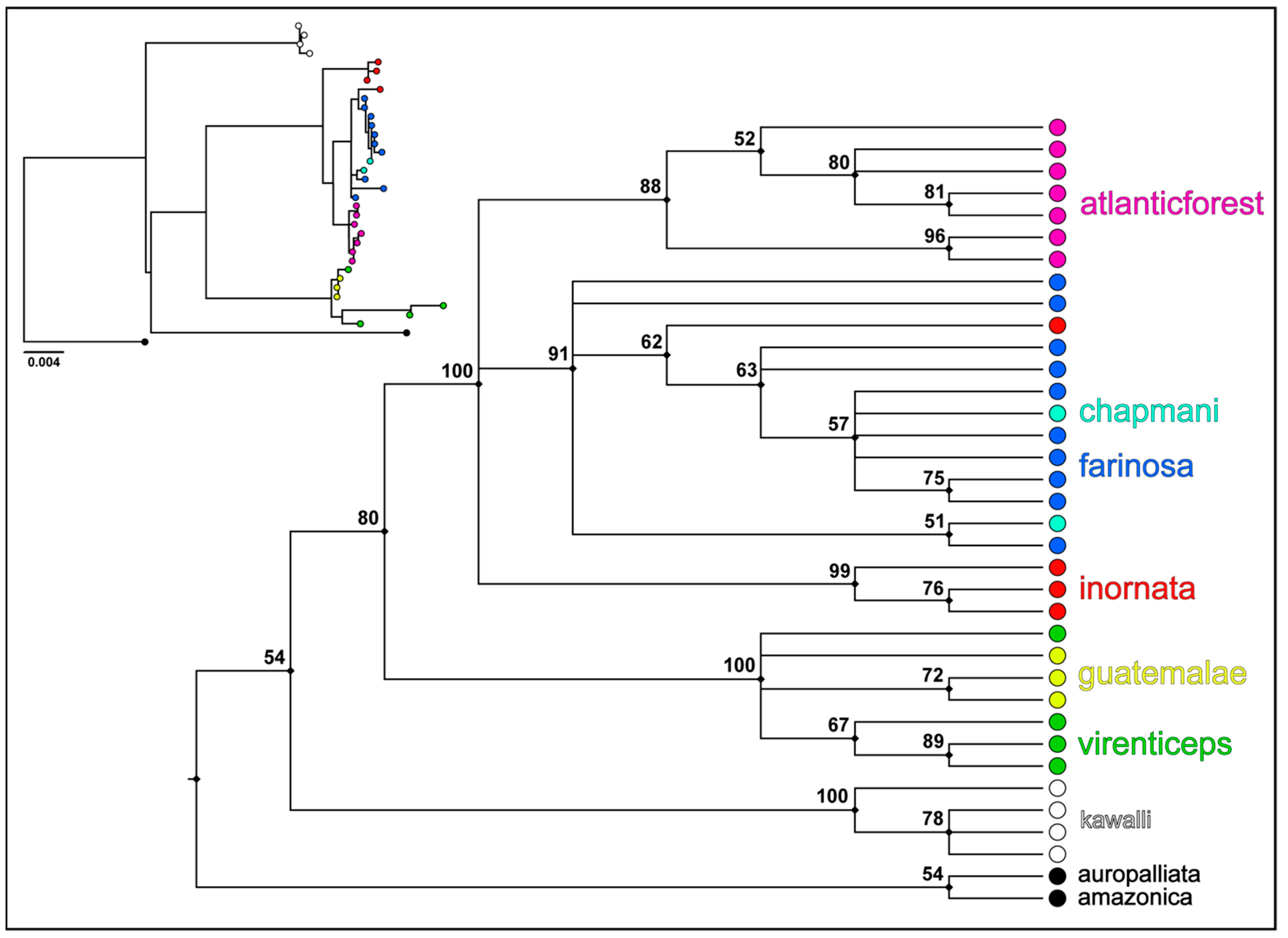
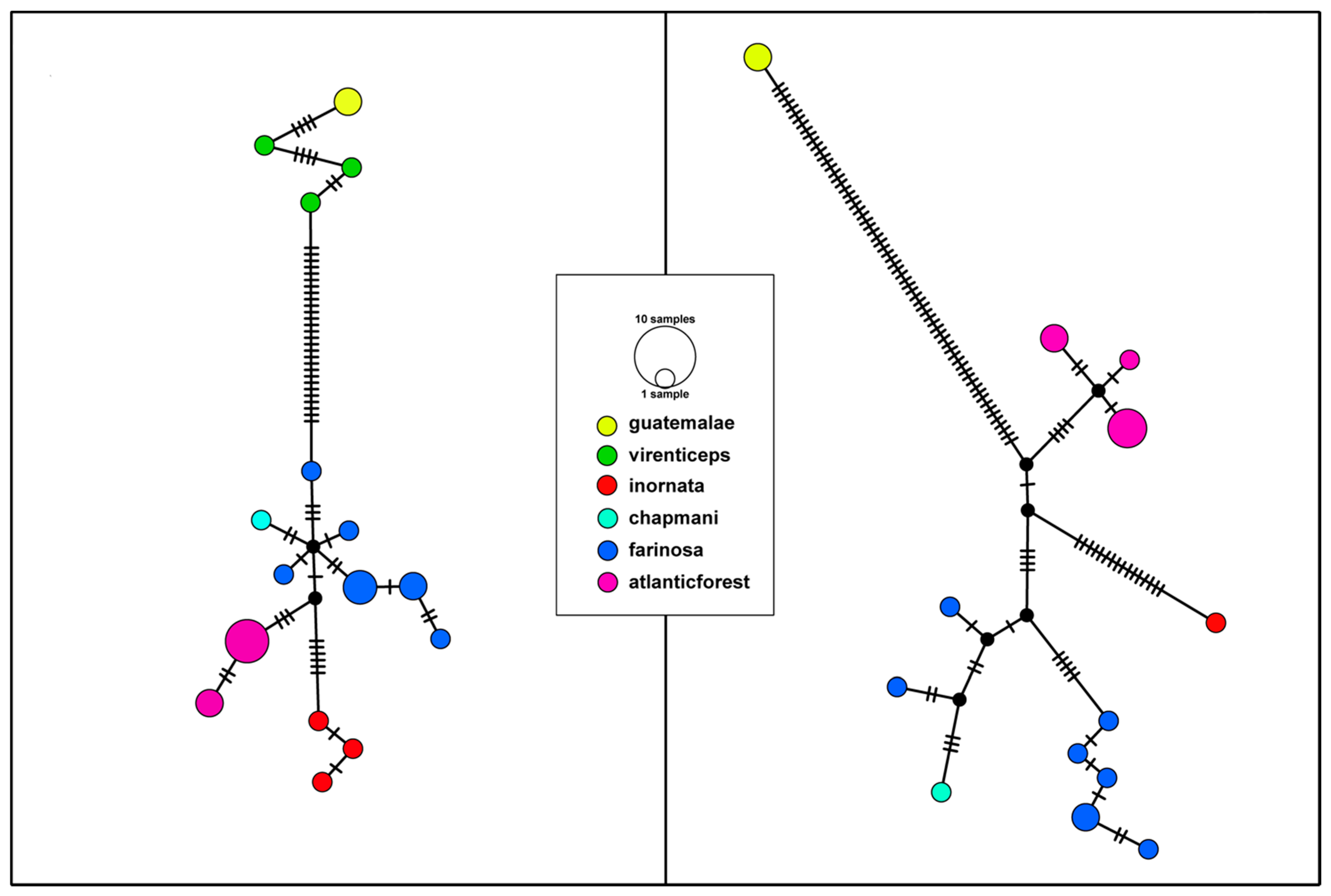
3.1. Phylogenetic Tree and Haplotype Network
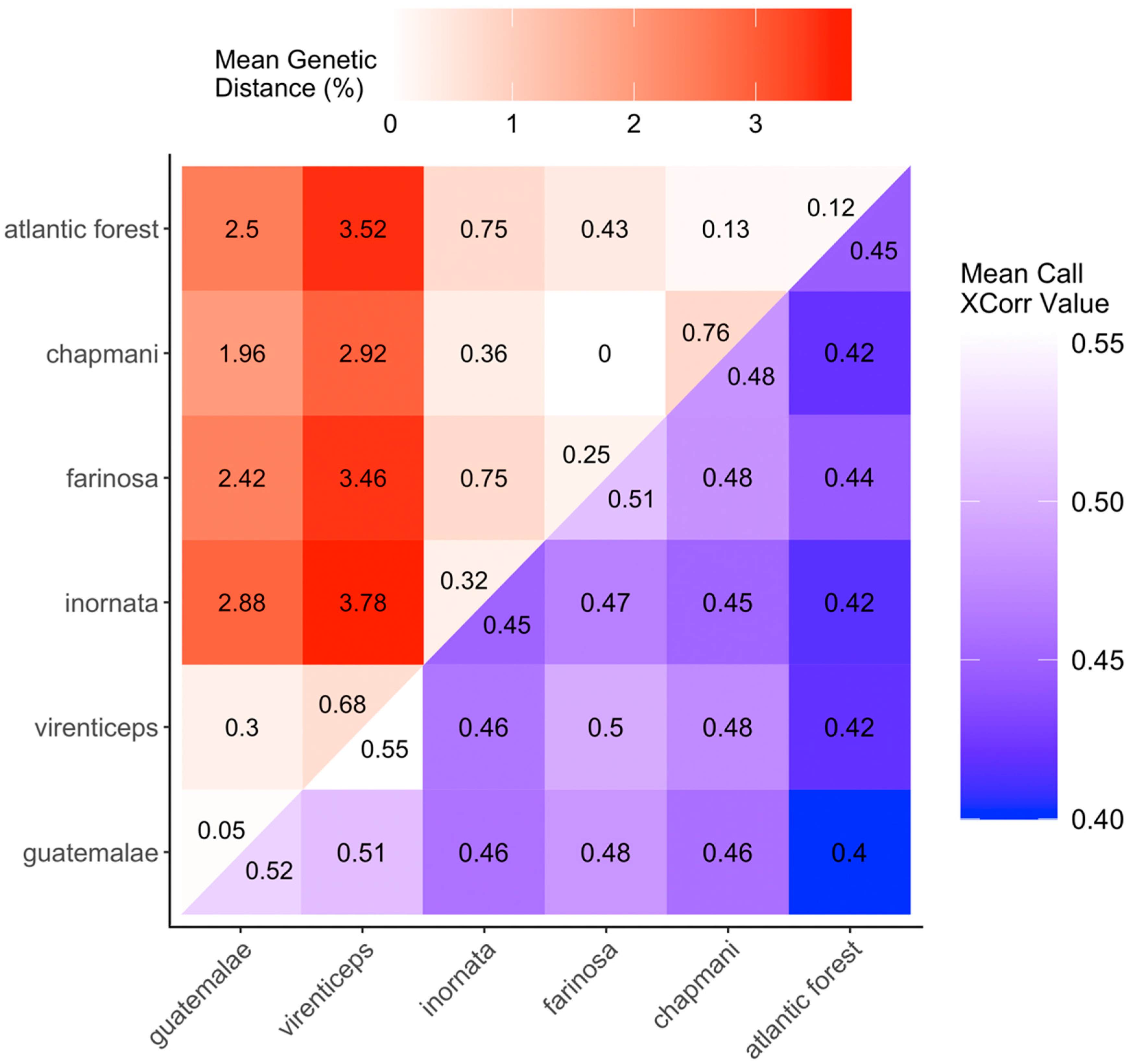
3.2. Genetic Differentiation
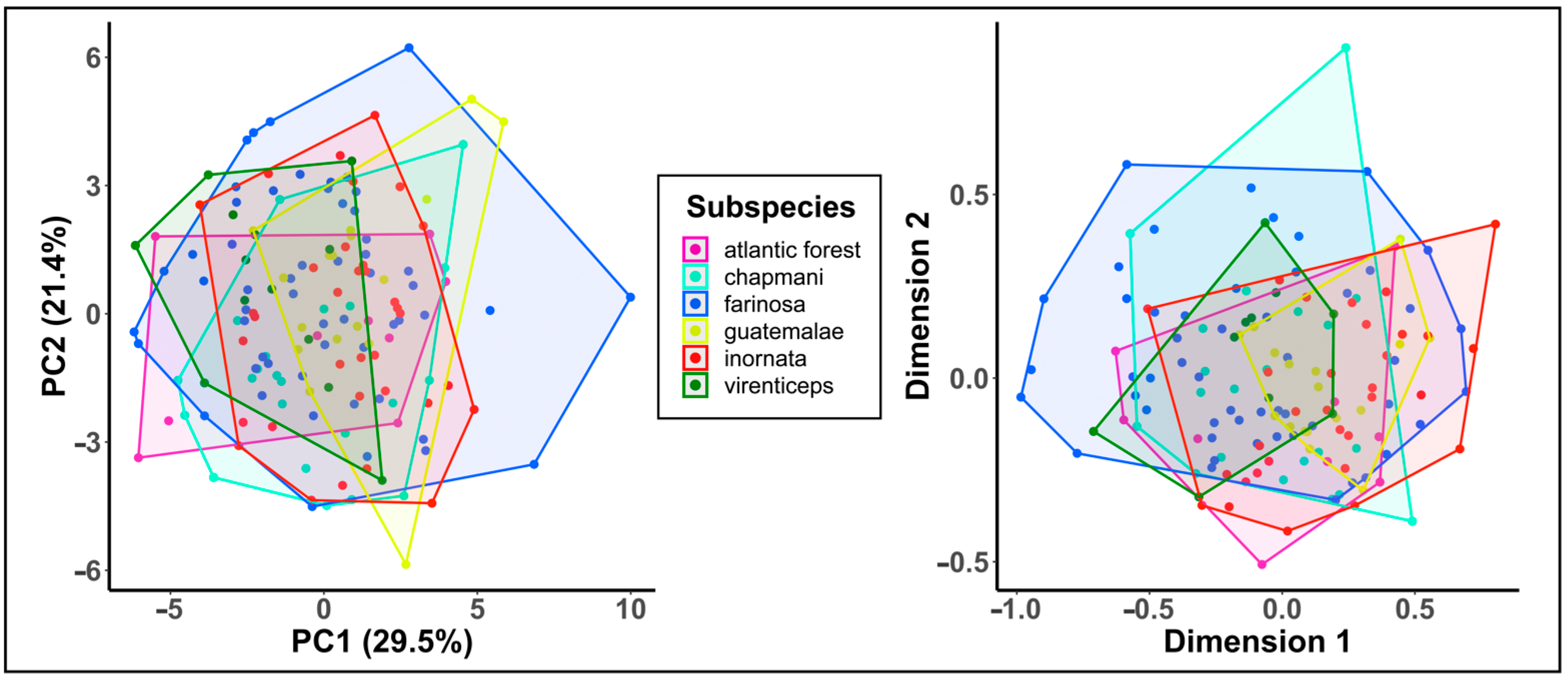
3.3. Call Similarity between Subspecies
3.4. Assessment of Genetic Distance and Vocal Divergence
4. Discussion
4.1. Genetic Relationships of the Mealy Amazon Clades
4.2. Utility of Behavioral Variation for Detecting Cryptic Species
4.3. Conservation Implications
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Struck, T.H.; Feder, J.L.; Bendiksby, M.; Birkeland, S.; Cerca, J.; Gusarov, V.I.; Kistenich, S.; Larsson, K.H.; Liow, L.H.; Nowak, M.D.; et al. Finding evolutionary processes hidden in cryptic species. Trends Ecol. Evol. 2018, 33, 153–163. [Google Scholar] [CrossRef]
- Bickford, D.; Lohman, D.J.; Sodhi, N.S.; Ng, P.K.L.; Meier, R.; Winker, K.; Ingram, K.K.; Das, I. Cryptic species as a window on diversity and conservation. Trends Ecol. Evol. 2007, 22, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Angulo, A.; Icochea, J. Cryptic species complexes, widespread species and conservation: Lessons from Amazonian frogs of the Leptodactylus marmoratus group (Anura: Leptodactylidae). Syst. Biodivers. 2010, 8, 357–370. [Google Scholar] [CrossRef]
- Davidson-Watts, I.; Walls, S.; Jones, G. Differential habitat selection by Pipistrellus pipistrellus and Pipistrellus pygmaeus identifies distinct conservation needs for cryptic species of echolocating bats. Biol. Conserv. 2006, 133, 118–127. [Google Scholar] [CrossRef]
- Delić, T.; Trontelj, P.; Rendoš, M.; Fišer, C. The importance of naming cryptic species and the conservation of endemic subterranean amphipods. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Wilkins, M.R.; Seddon, N.; Safran, R.J. Evolutionary divergence in acoustic signals: Causes and consequences. Trends Ecol. Evol. 2013, 28, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Jones, G. Acoustic signals and speciation: The roles of natural and sexual selection in the evolution of cryptic species. Adv. Study Behav. 1997, 26, 317–354. [Google Scholar] [CrossRef]
- Thabah, A.; Rossiter, S.J.; Kingston, T.; Zhang, S.; Parsons, S.; Mya, K.M.; Akbar, Z.; Jones, G. Genetic divergence and echolocation call frequency in cryptic species of Hipposideros larvatus s.l. (Chiroptera: Hipposideridae) from the Indo-Malayan region. Biol. J. Linn. Soc. 2006, 88, 119–130. [Google Scholar] [CrossRef]
- Kingston, T.; Lara, M.C.; Jones, G.; Akbar, Z.; Kunz, T.H.; Schneider, C.J. Acoustic divergence in two cryptic Hipposideros species: A role for social selection? Proc. R. Soc. B Biol. Sci. 2001, 268, 1381–1386. [Google Scholar] [CrossRef]
- Braune, P.; Schmidt, S.; Zimmermann, E. Acoustic divergence in the communication of cryptic species of nocturnal primates (Microcebus ssp.). BMC Biol. 2008, 6, 9–11. [Google Scholar] [CrossRef]
- Henry, C.S.; Brooks, S.J.; Duelli, P.; Johnson, J.B. Discovering the true Chrysoperla carnea (Insecta: Neuroptera: Chrysopidae) using song analysis, morphology, and ecology. Ann. Entomol. Soc. Am. 2006, 95, 172–191. [Google Scholar] [CrossRef]
- Funk, C.W.; Caminer, M.; Ron, S.R. High levels of cryptic species diversity uncovered in Amazonian frogs. Proc. R. Soc. B Biol. Sci. 2012, 279, 1806–1814. [Google Scholar] [CrossRef] [PubMed]
- Irwin, D.E.; Alström, P.; Olsson, U.; Benowitz-Fredericks, Z.M. Cryptic species in the genus Phylloscopus (Old World leaf warblers). Ibis (Lond. 1859) 2001, 143, 233–247. [Google Scholar] [CrossRef]
- Groth, J.G. Resolution of cryptic species in Appalachian red crossbills. Condor 1988, 90, 745–760. [Google Scholar] [CrossRef]
- Isler, M.L.; Isler, P.R.; Brumfield, R.T. Clinal variation in vocalizations of an antbird (Thamnophilidae) and implications for defining species limits. Auk 2005, 122, 433–444. [Google Scholar] [CrossRef]
- Irwin, D.E.; Thimgan, M.P.; Irwin, J.H. Call divergence is correlated with geographic and genetic distance in greenish warblers (Phylloscopus trochiloides): A strong role for stochasticity in signal evolution? J. Evol. Biol. 2008, 21, 435–448. [Google Scholar] [CrossRef]
- Lachlan, R.F.; Servedio, M.R. Song learning accelerates allopatric speciation. Evolution (N. Y.) 2004, 58, 2049–2063. [Google Scholar] [CrossRef]
- Olofsson, H.; Frame, A.M.; Servedio, M.R. Can reinforcement occur with a learned trait? Evolution (N. Y.) 2011, 65, 1992–2003. [Google Scholar] [CrossRef]
- Verzijden, M.N.; ten Cate, C.; Servedio, M.R.; Kozak, G.M.; Boughman, J.W.; Svensson, E.I. The impact of learning on sexual selection and speciation. Trends Ecol. Evol. 2012, 27, 511–519. [Google Scholar] [CrossRef]
- Mason, N.A.; Burns, K.J.; Tobias, J.A.; Claramunt, S.; Seddon, N.; Derryberry, E.P. Song evolution, speciation, and vocal learning in passerine birds. Evolution (N. Y.) 2017, 71, 786–796. [Google Scholar] [CrossRef]
- Doupe, A.J.; Kuhl, P.K. Birdsong and human speech: Common themes and mechanisms. Annu. Rev. Neurosci. 1999, 22, 567–631. [Google Scholar] [CrossRef]
- Farabaugh, S.M.; Linzenbold, A.; Dooling, R.J. Vocal plasticity in budgerigars (Melopsittacus undulatus): Evidence for social factors in the learning of contact calls. J. Comp. Psychol. 1994, 108, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Genzel, D.; Desai, J.; Paras, E.; Yartsev, M.M. Long-term and persistent vocal plasticity in adult bats. Nat. Commun. 2019, 10, 1–12. [Google Scholar] [CrossRef]
- Sewall, K.B.; Young, A.M.; Wright, T.F. Social calls provide novel insights into the evolution of vocal learning. Anim. Behav. 2016, 120, 163–172. [Google Scholar] [CrossRef]
- IUCN The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org (accessed on 15 May 2020).
- Berkunsky, I.; Quillfeldt, P.; Brightsmith, D.J.; Abbud, M.C.; Aguilar, J.M.R.E.; Alemán-Zelaya, U.; Aramburú, R.M.; Arce Arias, A.; Balas McNab, R.; Balsby, T.J.S.; et al. Current threats faced by Neotropical parrot populations. Biol. Conserv. 2017, 214, 278–287. [Google Scholar] [CrossRef]
- Wright, T.F.; Toft, C.A.; Enkerlin-Hoeflich, E.; Gonzalez-Elizondo, J.; Albornoz, M.; Rodríguez-Ferraro, A.; Rojas-Suárez, F.; Sanz, V.; Trujillo, A.; Beissinger, S.R.; et al. Nest poaching in Neotropical parrots. Conserv. Biol. 2001, 15, 710–720. [Google Scholar] [CrossRef]
- Wright, T.F.; Dahlin, C.R. Vocal dialects in parrots: Patterns and processes of cultural evolution. Emu Austral Ornithol. 2018, 118, 50–66. [Google Scholar] [CrossRef] [PubMed]
- Slabbekoorn, H.; Smith, T.B. Bird song, ecology and speciation. Philos. Trans. R. Soc. B Biol. Sci. 2002, 357, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Rendell, L.; Mesnick, S.L.; Dalebout, M.L.; Burtenshaw, J.; Whitehead, H. Can genetic differences explain vocal dialect variation in sperm whales, Physeter macrocephalus? Behav. Genet. 2012, 42, 332–343. [Google Scholar] [CrossRef]
- Keighley, M.V.; Heinsohn, R.; Langmore, N.E.; Murphy, S.A.; Peñalba, J.V. Genomic population structure aligns with vocal dialects in Palm Cockatoos (Probosciger aterrimus); evidence for refugial late-Quaternary distribution? Emu 2019, 119, 24–37. [Google Scholar] [CrossRef]
- Nevo, E.; Heth, G.; Beiles, A.; Frankenberg, E. Geographic dialects in blind mole rats: Role of vocal communication in active speciation. Proc. Natl. Acad. Sci. USA 1987, 84, 3312–3315. [Google Scholar] [CrossRef]
- Russello, M.A.; Stahala, C.; Lalonde, D.; Schmidt, K.L.; Amato, G. Cryptic diversity and conservation units in the Bahama parrot. Conserv. Genet. 2010, 11, 1809–1821. [Google Scholar] [CrossRef]
- McElroy, K.; Beattie, K.; Symonds, M.R.E.; Joseph, L. Mitogenomic and nuclear diversity in the Mulga Parrot of the Australian arid zone: Cryptic subspecies and tests for selection. Emu Austral Ornithol. 2018, 118, 22–35. [Google Scholar] [CrossRef]
- Murphy, S.A.; Joseph, L.; Burbidge, A.H.; Austin, J. A cryptic and critically endangered species revealed by mitochondrial DNA analyses: The western Ground Parrot. Conserv. Genet. 2011, 12, 595–600. [Google Scholar] [CrossRef]
- Wenner, T.J.; Russello, M.A.; Wright, T.F. Cryptic species in a Neotropical parrot: Genetic variation within the Amazona farinosa species complex and its conservation implications. Conserv. Genet. 2012, 13, 1427–1432. [Google Scholar] [CrossRef]
- Juniper, T.; Parr, M. Parrots: A Guide to Parrots of the World, 1st ed.; Yale University Press: New Haven, CT, USA, 1998; ISBN 978-0300074536. [Google Scholar]
- Collar, N.; del Hoyo, J.; Bonan, A.; Kirwan, G.M.; Boesman, P.F.D. Mealy Parrot (Amazona farinosa); Version 1; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Government of São Paulo. Decree on the Declaration of the Species of Wild Fauna in the State of São Paulo Regionally Extinct, Those Threatened with Extinction, Those Almost Threatened and Those with Insufficient Data for Evaluation. Available online: https://www.al.sp.gov.br/repositorio/legislacao/decreto/2018/decreto-63853-27.11.2018.html (accessed on 27 November 2018).
- Ribeiro, M.C.; Martensen, A.C.; Metzger, J.P.; Tabarelli, M.; Scarano, F.; Fortin, M.-J. The Brazilian Atlantic Forest: A Shrinking Biodiversity Hotspot. Biodiversity Hotspots, 1st ed.; Springer: Berlin, Germany, 2011. [Google Scholar]
- Silva, J.M.C.; Casteleti, C.H.M. Status of the biodiversity of the Atlantic Forest of Brazil. In The Atlantic Forest of South America: Biodiversity Status, Threats, and Outlook; Galindo-Leal, C., de Gusmão Câmara, I., Eds.; Island Press: Washington, DC, USA, 2003; pp. 43–59. [Google Scholar]
- Batalha-Filho, H.; Fjeldså, J.; Fabre, P.H.; Miyaki, C.Y. Connections between the Atlantic and the Amazonian forest avifaunas represent distinct historical events. J. Ornithol. 2013, 154, 41–50. [Google Scholar] [CrossRef]
- Fusinatto, L.A.; Alexandrino, J.; Haddad, C.F.B.; Brunes, T.O.; Rocha, C.F.D.; Sequeira, F. Cryptic genetic diversity is paramount in small-bodied amphibians of the genus Euparkerella (Anura: Craugastoridae) endemic to the Brazilian Atlantic forest. PLoS ONE 2013, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Costa, W.J.E.M.; Amorim, P.F. Delimitation of cryptic species of Notholebias, a genus of seasonal miniature killifishes threatened with extinction from the Atlantic Forest of outh-eastern Brazil (Cyprinodontiformes: Rivulidae). Ichthyol. Explor. Freshwaters 2013, 24, 63–72. [Google Scholar]
- Mata, H.; Fontana, C.S.; Maurício, G.N.; Bornschein, M.R.; de Vasconcelos, M.F.; Bonatto, S.L. Molecular phylogeny and biogeography of the eastern Tapaculos (Aves: Rhinocryptidae: Scytalopus, Eleoscytalopus): Cryptic diversification in Brazilian Atlantic Forest. Mol. Phylogenet. Evol. 2009, 53, 450–462. [Google Scholar] [CrossRef]
- Forlani, M.C.; Tonini, J.F.R.; Cruz, C.A.G.; Zaher, H.; de Sá, R.O. Molecular and morphological data reveal three new cryptic species of Chiasmocleis (Mehely 1904) (Anura, Microhylidae) endemic to the Atlantic Forest, Brazil. PeerJ 2017, 2017, 1–43. [Google Scholar] [CrossRef]
- Nemésio, A.; Engel, M.S. Three new cryptic species of Euglossa from Brazil (Hymenoptera, apidae). Zookeys 2012, 222, 47–68. [Google Scholar] [CrossRef]
- Ribeiro, M.C.; Metzger, J.P.; Martensen, A.C.; Ponzoni, F.J.; Hirota, M.M. The Brazilian Atlantic Forest: How much is left, and how is the remaining forest distributed? Implications for conservation. Biol. Conserv. 2009, 142, 1141–1153. [Google Scholar] [CrossRef]
- Tabarelli, M.; Pinto, L.P.; Silva, J.M.C.; Hirota, M.; Bedê, L. Challenges and opportunities for biodiversity conservation in the Brazilian Atlantic forest. Conserv. Biol. 2005, 19, 695–700. [Google Scholar] [CrossRef]
- Cock, P.J.A.; Antao, T.; Chang, J.T.; Chapman, B.A.; Cox, C.J.; Dalke, A.; Friedberg, I.; Hamelryck, T.; Kauff, F.; Wilczynski, B.; et al. Biopython: Freely available Python tools for computational molecular biology and bioinformatics. Bioinformatics 2009, 25, 1422–1423. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. 2010 Gatew. Comput. Environ. Work. GCE 2010 2010. [Google Scholar] [CrossRef]
- FigTree; Version 1.4.4; A Graphical Viewer of Phylogenetic Trees; Andrew Rambaut: Edinburgh, UK, 2007.
- Bandelt, H.-J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Stecher, G.; Tamura, K.; Kumar, S. Molecular Evolutionary Genetics Analysis (MEGA) for macOS. Mol. Biol. Evol. 2020, 37, 1237–1239. [Google Scholar] [CrossRef]
- Araya-Salas, M.; Smith-Vidaurre, G. warbleR: An R package to streamline analysis of animal acoustic signals. Methods Ecol. Evol. 2016, 184–191. [Google Scholar] [CrossRef]
- Schmidt, K.L.; Aardema, M.L.; Amato, G. Genetic analysis reveals strong phylogeographical divergences within the Scarlet Macaw Ara macao. Ibis 2020, 162, 735–748. [Google Scholar] [CrossRef]
- Eberhard, J.R.; Bermingham, E. Phylogeny and biogeography of the Amazona ochrocephala (Aves: Psittacidae) complex. Auk 2004, 121, 318–332. [Google Scholar] [CrossRef]
- Johns, G.C.; Avise, J.C. A comparative summary of genetic distances in the vertebrates from the mitochondrial cytochrome b gene. Mol. Biol. Evol. 1998, 15, 372–382. [Google Scholar] [CrossRef] [PubMed]
- Bocalini, F.; Bolívar-Leguizamón, S.D.; Silveira, L.F.; Bravo, G.A. Comparative phylogeographic and demographic analyses reveal a congruent pattern of sister relationship between bird populations of the northern and south-central Atlantic Forest. Mol. Phylogenet. Evol. 2020, 154, 106973. [Google Scholar] [CrossRef]
- Capurucho, J.M.G.; Ashley, M.V.; Ribas, C.C.; Bates, J.M. Connecting Amazonian, Cerrado, and Atlantic forest histories: Paraphyly, old divergences, and modern population dynamics in tyrant-manakins (Neopelma/Tyranneutes, Aves: Pipridae). Mol. Phylogenet. Evol. 2018, 127, 696–705. [Google Scholar] [CrossRef]
- Cabanne, G.S.; d’Horta, F.M.; Sari, E.H.R.; Santos, F.R.; Miyaki, C.Y. Nuclear and mitochondrial phylogeography of the Atlantic forest endemic Xiphorhynchus fuscus (Aves: Dendrocolaptidae): Biogeography and systematics implications. Mol. Phylogenet. Evol. 2008, 49, 760–773. [Google Scholar] [CrossRef]
- Costa, L.P. The historical bridge between the Amazon and the Atlantic Forest of Brazil: A study of molecular phylogeography with small mammals. J. Biogeogr. 2003, 30, 71–86. [Google Scholar] [CrossRef]
- Trujillo-Arias, N.; Calderón, L.; Santos, F.R.; Miyaki, C.Y.; Aleixo, A.; Witt, C.C.; Tubaro, P.L.; Cabanne, G.S. Forest corridors between the central Andes and the southern Atlantic Forest enabled dispersal and peripatric diversification without niche divergence in a passerine. Mol. Phylogenet. Evol. 2018, 128, 221–232. [Google Scholar] [CrossRef]
- Lavinia, P.D.; Barreira, A.S.; Campagna, L.; Tubaro, P.L.; Lijtmaer, D.A. Contrasting evolutionary histories in Neotropical birds: Divergence across an environmental barrier in South America. Mol. Ecol. 2019, 28, 1730–1747. [Google Scholar] [CrossRef]
- Carnaval, A.C.; Moritz, C. Historical climate modelling predicts patterns of current biodiversity in the Brazilian Atlantic forest. J. Biogeogr. 2008, 35, 1187–1201. [Google Scholar] [CrossRef]
- Cheng, H.; Sinha, A.; Cruz, F.W.; Wang, X.; Edwards, R.L.; D’Horta, F.M.; Ribas, C.C.; Vuille, M.; Stott, L.D.; Auler, A.S. Climate change patterns in Amazonia and biodiversity. Nat. Commun. 2013, 4. [Google Scholar] [CrossRef]
- Smith-Vidaurre, G.; Araya-Salas, M.; Wright, T.F. Individual signatures outweigh social group identity in contact calls of a communally nesting parrot. Behav. Ecol. 2021, 31, 448–458. [Google Scholar] [CrossRef]
- Wright, T.F.; Wilkinson, G.S. Population genetic structure and vocal dialects in an amazon parrot. Proc. R. Soc. B Biol. Sci. 2001, 268, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Ribot, R.F.H.; Buchanan, K.L.; Endler, J.A.; Joseph, L.; Bennett, A.T.D.; Berg, M.L. Learned vocal variation is associated with abrupt cryptic genetic change in a parrot species complex. PLoS ONE 2012, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- MacDougall-Shackleton, E.A.; MacDougall-Shackleton, S.A. Cultural and genetic evolution in mountain white-crowned sparrows: Song dialects are associated with population structure. Evolution (N. Y.) 2001, 55, 2568–2575. [Google Scholar] [CrossRef]
- Lipshutz, S.E.; Overcast, I.A.; Hickerson, M.J.; Brumfield, R.T.; Derryberry, E.P. Behavioural response to song and genetic divergence in two subspecies of white-crowned sparrows (Zonotrichia leucophrys). Mol. Ecol. 2017, 26, 3011–3027. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hellmich, D.L.; Saidenberg, A.B.S.; Wright, T.F. Genetic, but Not Behavioral, Evidence Supports the Distinctiveness of the Mealy Amazon Parrot in the Brazilian Atlantic Forest. Diversity 2021, 13, 273. https://doi.org/10.3390/d13060273
Hellmich DL, Saidenberg ABS, Wright TF. Genetic, but Not Behavioral, Evidence Supports the Distinctiveness of the Mealy Amazon Parrot in the Brazilian Atlantic Forest. Diversity. 2021; 13(6):273. https://doi.org/10.3390/d13060273
Chicago/Turabian StyleHellmich, Dominique L., Andre B. S. Saidenberg, and Timothy F. Wright. 2021. "Genetic, but Not Behavioral, Evidence Supports the Distinctiveness of the Mealy Amazon Parrot in the Brazilian Atlantic Forest" Diversity 13, no. 6: 273. https://doi.org/10.3390/d13060273
APA StyleHellmich, D. L., Saidenberg, A. B. S., & Wright, T. F. (2021). Genetic, but Not Behavioral, Evidence Supports the Distinctiveness of the Mealy Amazon Parrot in the Brazilian Atlantic Forest. Diversity, 13(6), 273. https://doi.org/10.3390/d13060273