
Potential Distribution of Colonizing Nine-Banded Armadillos at Their Northern Range Edge

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
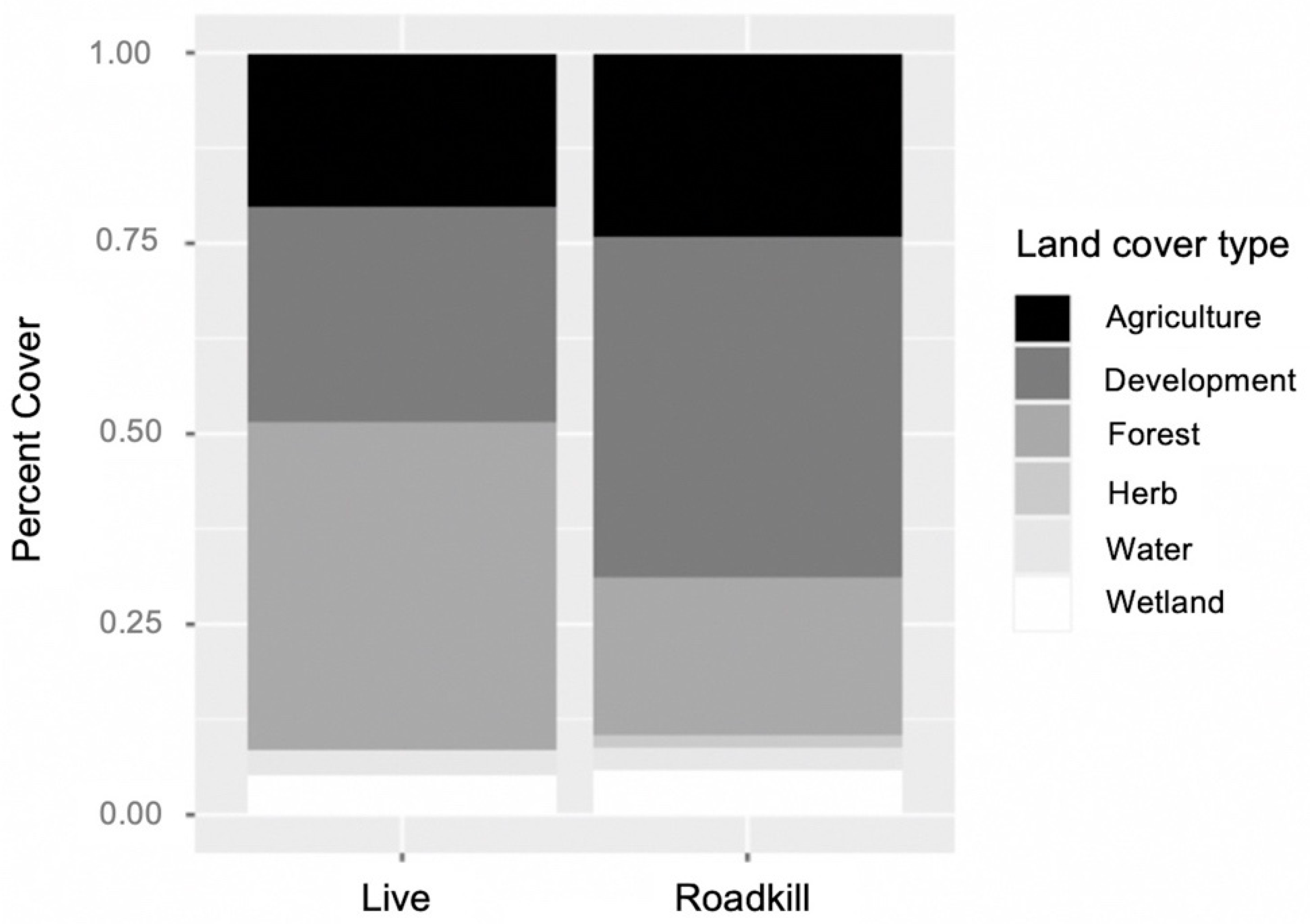
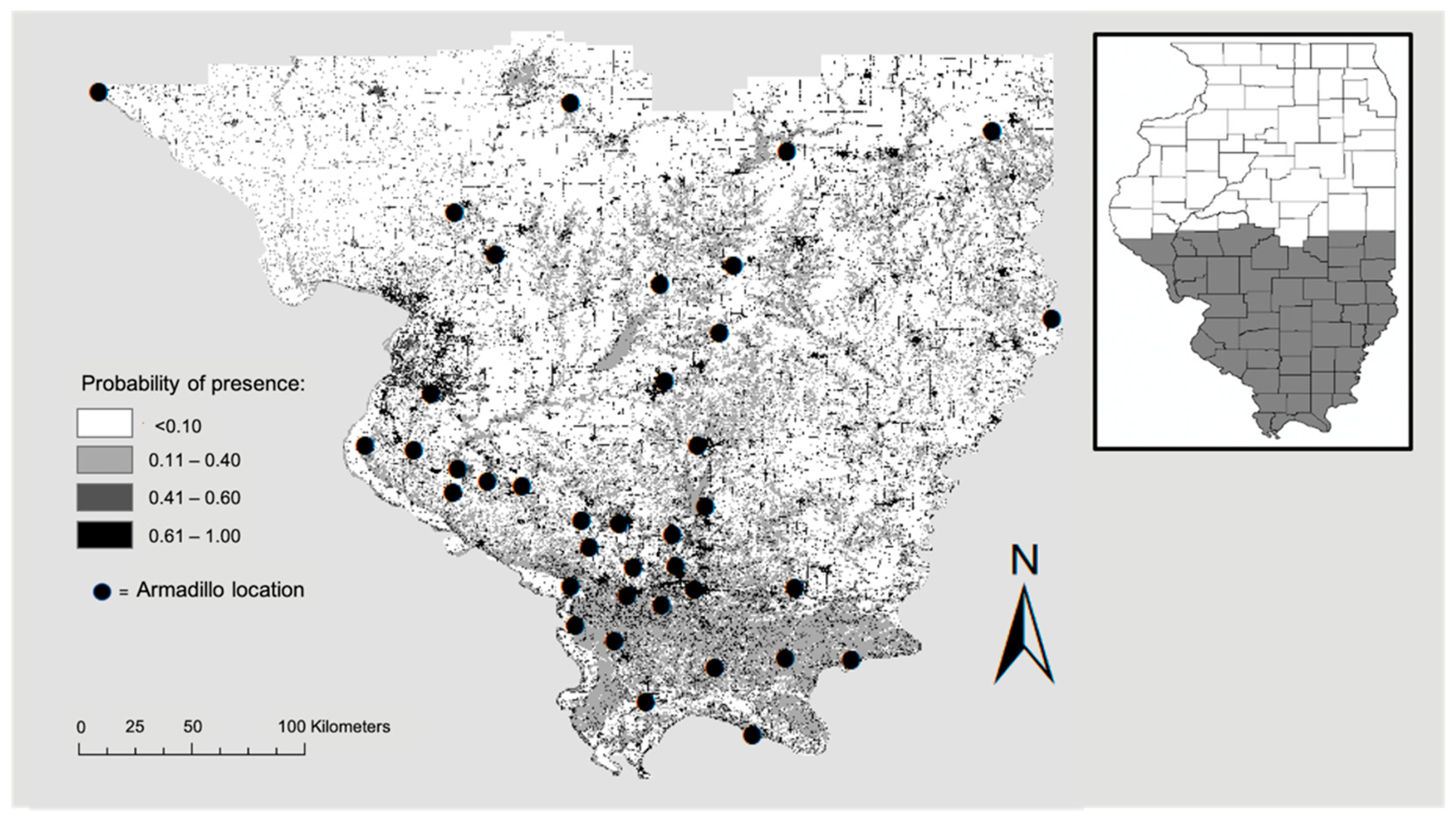
2.2. Armadillo Presence
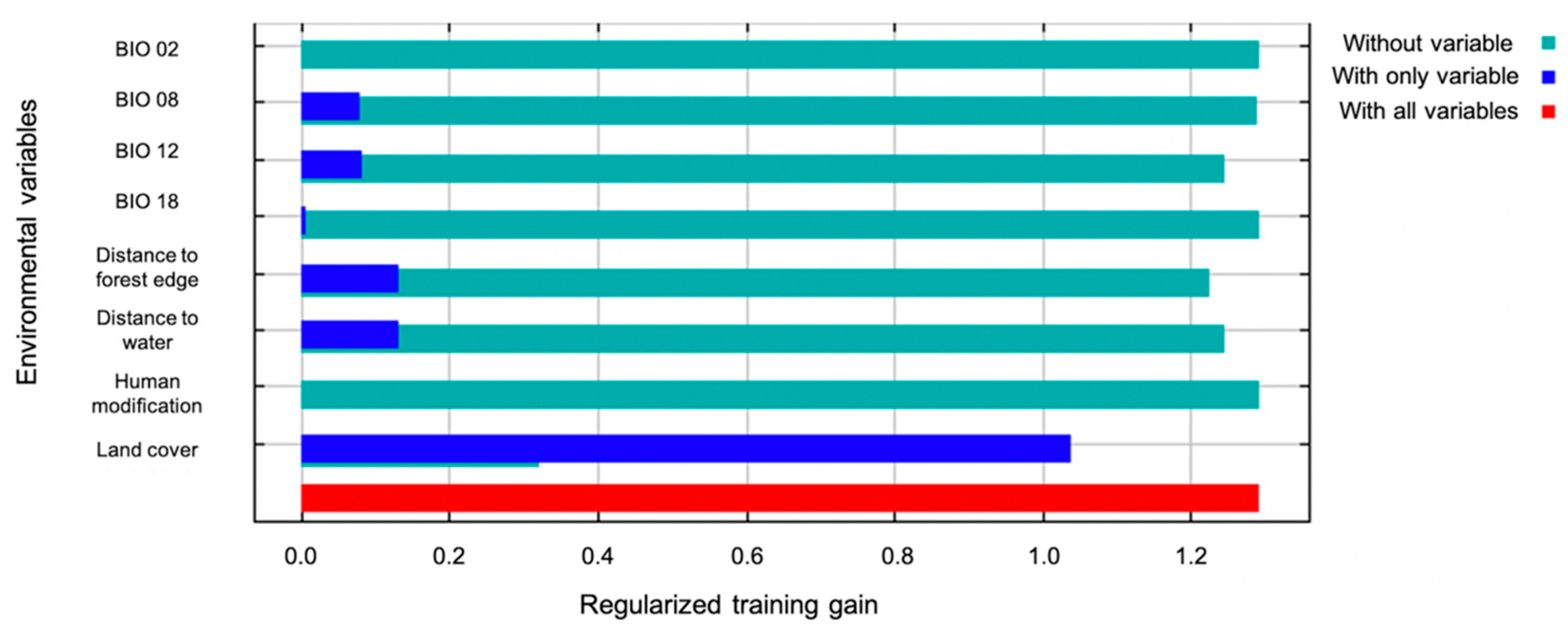
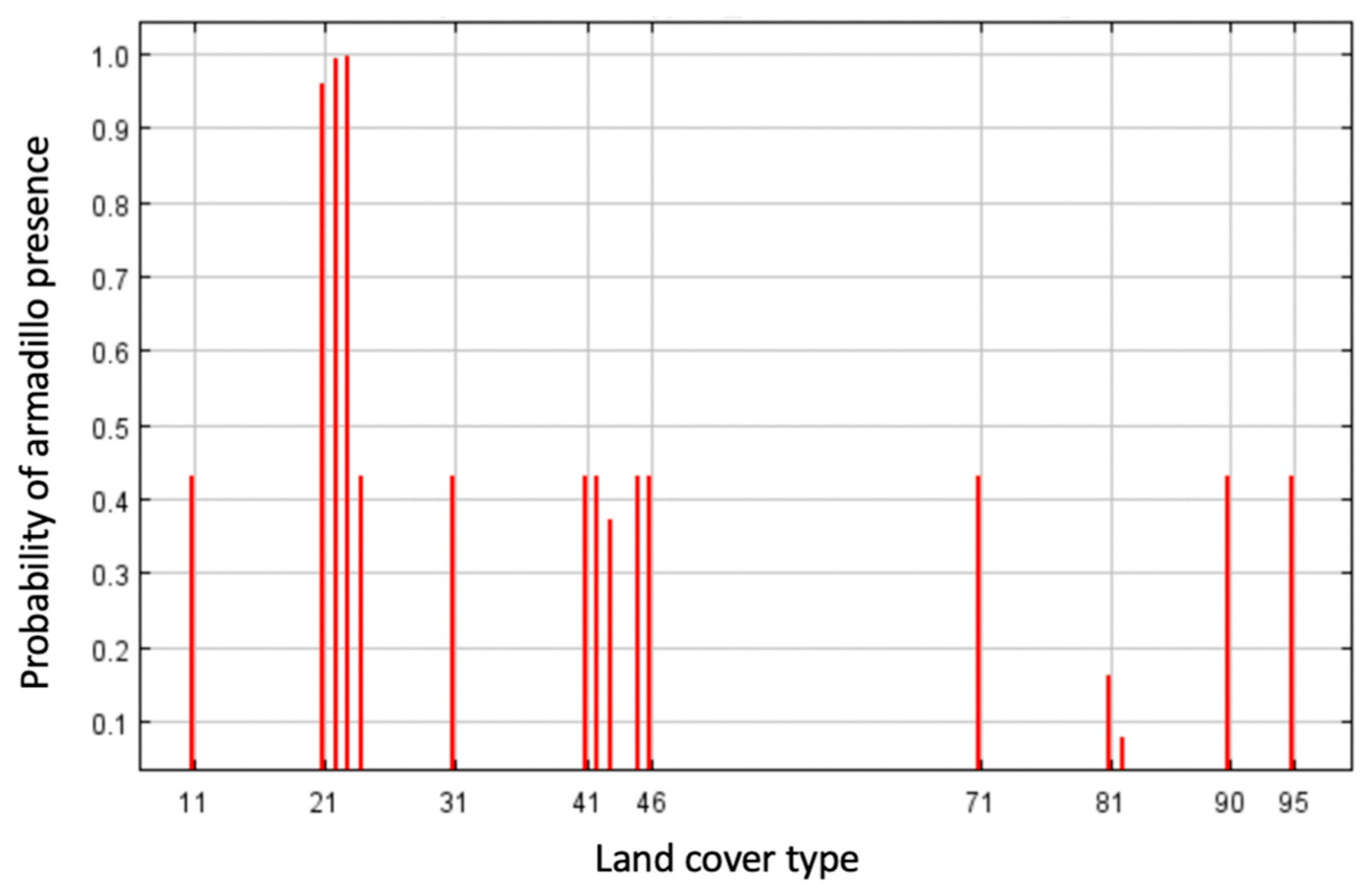
2.3. Environmental Variables
2.4. Potential Distribution
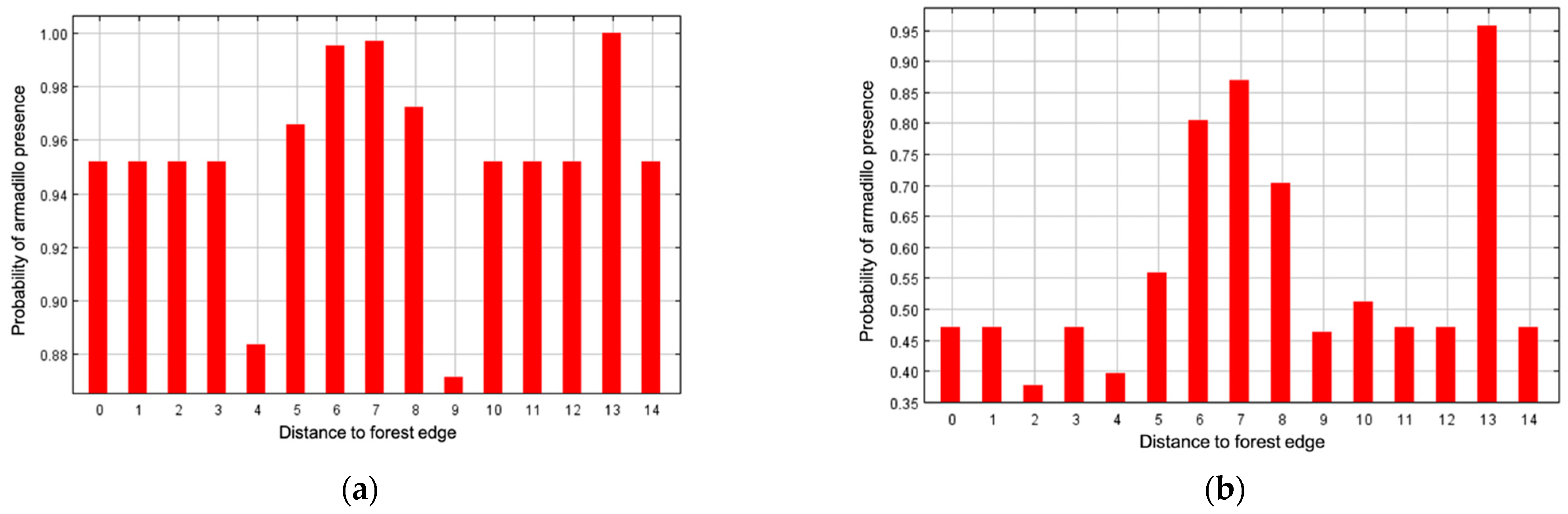
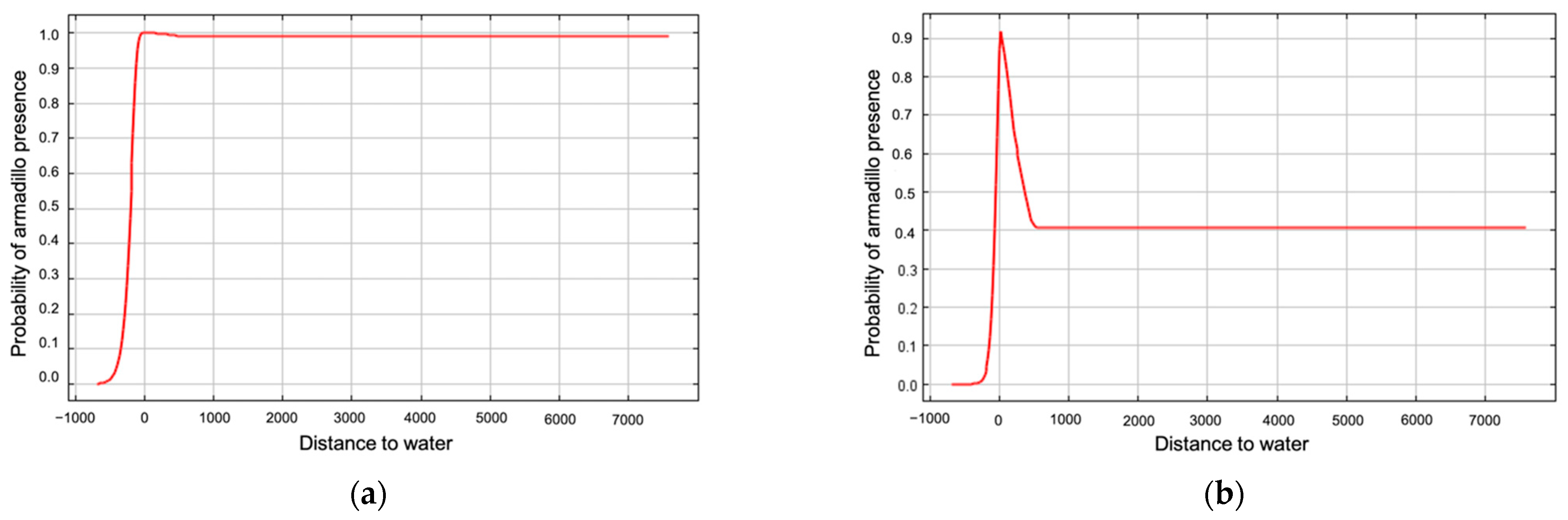
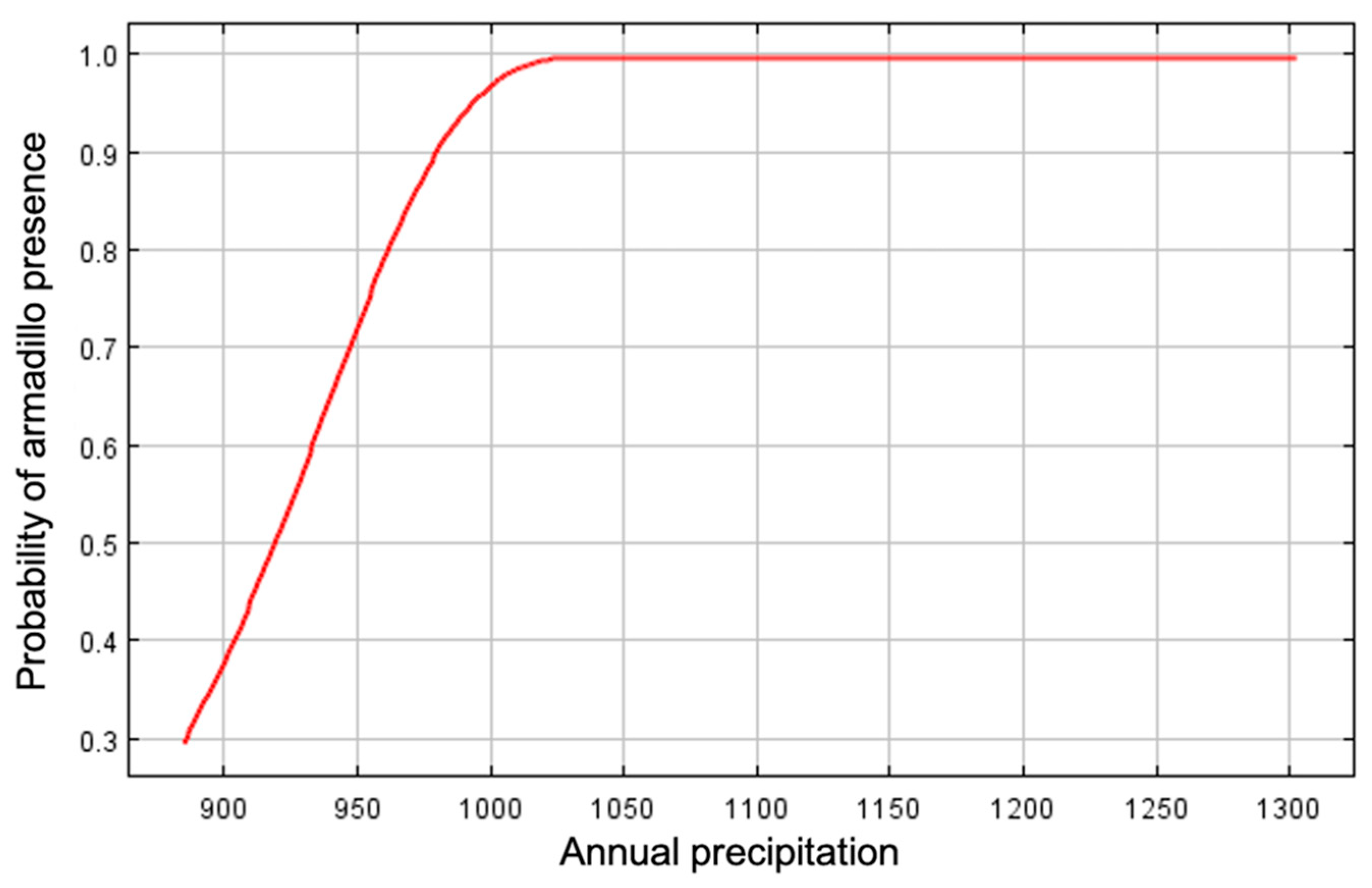
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Loughry, W.J.; McDonough, C.M. The Nine-Banded Armadillo: A Natural History; University of Oklahoma Press: Norman, Oklahoma, 2013. [Google Scholar]
- Feng, X.; Papeş, M. Ecological niche modelling confirms potential north-east range expansion of the nine-banded armadillo (Dasypus novemcinctus) in the USA. J. Biogeogr. 2015, 42, 803–807. [Google Scholar] [CrossRef]
- Taulman, J.F.; Robbins, L.W. Range expansion and distributional limits of the nine-banded armadillo in the United States: An update of Taulman & Robbins (1996). J. Biogeogr. 2014, 41, 1626–1630. [Google Scholar]
- Feijó, A.; Patterson, B.D.; Cordeiro-Estrela, P. Phenotypic variability and environmental tolerance shed light on nine-banded armadillo Nearctic invation. Biol. Invasions 2019, 22, 255–269. [Google Scholar] [CrossRef]
- Genoways, H.H.; Freeman, P.W. Recent northern records of the nine-banded armadillo (Dasypodidae) in Nebraska. Southwest. Nat. 1998, 43, 491–495. [Google Scholar]
- Hofmann, J.E. Records of nine-banded armadillos, Dasypus novemcinctus, in Illinois. Trans. Illinois State Acad. Sci. 2009, 102, 95–106. [Google Scholar]
- Van Deelen, T.R.; Parrish, J.D.; Heske, E.J. A nine-banded armadillo (Dasypus novemcinctus) from central Illinois. Southwest. Nat. 2002, 47, 489–491. [Google Scholar] [CrossRef]
- Mcbee, K.; Baker, R.J. Dasypus novemcinctus. Mamm. Species 1982, 162, 1–9. [Google Scholar] [CrossRef]
- Rodrigues, T.F.; Mantellatto, A.M.B.; Superina, M.; Chiarello, A.G. Ecosystem services provided by armadillos. Biol. Rev. 2020, 95, 1–21. [Google Scholar] [CrossRef]
- Yeo, M.; Acosta, N.; Llewellyn, M.; Sánchez, H.; Adamson, S.; Miles, G.A.J.; López, E.; González, N.; Patterson, J.S.; Gaunt, M.W.; et al. Origins of Chagas disease: Didelphis species are natural hosts of Trypanosoma cruzi I and armadillos hosts of Trypanosoma cruzi II, including hybrids. Int. J. Parasitol. 2005, 35, 225–233. [Google Scholar] [CrossRef]
- Hiestand, S.; Nielsen, C.K.; Jimenez, A.F. Modelling potential presence of metazoan endoparasites of bobcats (Lynx rufus) using verified records. Folia Parasitol. 2014, 61, 401–410. [Google Scholar] [CrossRef]
- Truman, R. Leprosy in wild armadillos. Lepr. Rev. 2005, 76, 198–208. [Google Scholar] [CrossRef]
- Fitch, H.S.; Goodrum, P.; Newman, C. The armadillo in the Southeastern United States. J. Mammal. 1952, 33, 21–37. [Google Scholar] [CrossRef]
- Loughry, W.J.; Truman, R.W.; McDonough, C.M.; Tilak, M.-K.; Garnier, S.; Delsuc, F. Is leprosy spreading among nine-banded armadillos in the southeastern United States? J. Wildl. Dis. 2009, 45, 144–152. [Google Scholar] [CrossRef]
- Staller, E.L.; Warnell, D.B.; Palmer, W.E.; Carroll, J.P.; Thornton, R.P.; Sisson, D.C. Identifying predators at northern bobwhite nests. J. Wildl. Manag. 2005, 69, 124–132. [Google Scholar] [CrossRef]
- Williams, M. Northern Bobwhite. In Endangered and Disappearing Birds of the Midwest; Indiana University Press: Bloomington, Indiana, 2018; pp. 4–7. [Google Scholar]
- Berkman, L.K.; Nielsen, C.K.; Roy, C.L.; Heist, E.J. Population genetic structure among bobwhite in an agriculturally modified landscape. J. Wildl. Manag. 2013, 77, 1472–1481. [Google Scholar] [CrossRef]
- Berkman, L.K.; Nielsen, C.K.; Roy, C.L.; Heist, E.J. Resistance is futile: Effects of landscape features on gene flow of the northern bobwhite. Conserv. Genet. 2013, 14, 323–332. [Google Scholar] [CrossRef]
- Rollins, D.; Carroll, J.P. Impacts of predation on northern bobwhite and scaled quail. Wildl. Soc. Bull. 2001, 29, 39–51. [Google Scholar]
- McDonough, C.M.; Delaney, M.A.; Le, P.Q.; Blackmore, M.S.; Loughry, W.J. Burrow characteristics and habitat associations of armadillos in Brazil and the United States of America. Rev. Biol. Trop. 2000, 48, 109–120. [Google Scholar]
- McDonough, C.M.; Loughry, W.J. Impacts of land management practices on a population of nine-banded armadillos in northern Florida. Wildl. Soc. Bull. 2005, 33, 1198–1209. [Google Scholar] [CrossRef]
- Gammons, D.J.; Mengak, M.T.; Conner, L.M. Armadillo habitat selection in southwestern Georgia. J. Mammal. 2009, 90, 356–362. [Google Scholar] [CrossRef]
- Inbar, M.; Mayer, R.T. Spatio-temporal trends in armadillo diurnal activity and road-kills in Central Florida. Wildl. Soc. Bull. 1999, 27, 865–872. [Google Scholar]
- Humphrey, S.R. Zoogeography of the nine-banded armadillo (Dasypus novemcinctus) in the United States. Bioscience 1974, 24, 457–462. [Google Scholar] [CrossRef]
- Taulman, J.F.; Robbins, L.W. Recent range expansion and distributional limits of the nine- banded armadillo (Dasypus novemcinctus) in the United States. J. Biogeogr. 1996, 23, 635–648. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Pandey, R.; Papes, M. Changes in future potential distributions of apex predator and mesopredator mammals in North America. Reg. Environ. Chang. 2017, 18, 1223–1233. [Google Scholar] [CrossRef]
- Greenspan, E.; Nielsen, C.K.; Cassel, K.W. Potential distribution of coyotes (Canis latrans), Virginia opossums (Didelphis virginiana), striped skunks (Mephitis mephitis), and raccoons (Procyon lotor) in the Chicago Metropolitan Area. Urban Ecosyst. 2018, 21, 983–997. [Google Scholar] [CrossRef]
- Peterson, A.T.; Robins, C.R. Using ecological-niche modeling to predict barred owl invasions with implications for spotted owl conservation. Conserv. Biol. 2003, 17, 1161–1165. [Google Scholar] [CrossRef]
- Schank, C.J.; Cove, M.V.; Kelly, M.J.; Mendoza, E.; O’Farrill, G.; Reyna-Hurtado, R.; Meyer, N.; Jordan, C.A.; González-Maya, J.F.; Lizcano, D.J.; et al. Using a novel model approach to assess the distribution and conservation status of the endangered Baird’s tapir. Divers. Distrib. 2017, 23, 1459–1471. [Google Scholar] [CrossRef]
- Abba, A.M.; Tognelli, M.F.; Seitz, V.P.; Bender, J.B.; Vizcaíno, S.F. Distribution of extant xenarthrans (Mammalia: Xenarthra) in Argentina using species distribution models. Mammalia 2012, 76, 123–136. [Google Scholar] [CrossRef]
- Pietrek, A.G.; González-Roglich, M. Post-establishment changes in habitat selection by an invasive species: Beavers in the Patagonian steppe. Biol. Invasions 2015, 17, 3225–3235. [Google Scholar] [CrossRef]
- Perez-Barberia, F.J.; Hooper, R.J.; Gordon, I.J. Long-term density-dependent changes in habitat selection in red deer (Cervus elaphus). Oecologia 2013, 173, 837–847. [Google Scholar] [CrossRef]
- Robson, A.S.; van Aarde, R.J. Changes in elephant conservation management promote density-dependent habitat selection in the Kruger National Park. Anim. Conserv. 2018, 21, 302–312. [Google Scholar] [CrossRef]
- Pearson, R.G.; Dawson, T.P.; Berry, P.A. SPECIES: A spatial evaluation of climate impact on the envelope on a species. Ecol. Model. 2002, 154, 289–300. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Mackey, B.G.; Lindenmayer, D.B. Towards a hierarchical framework for modelling the spatial distribution of animals. J. Biogeogr. 2001, 28, 1147–1166. [Google Scholar] [CrossRef]
- Pearson, R.G.; Dawson, T.P.; Liu, C. Modelling species distributions in Britain: A hierarchical integration of climate and land-cover data. Ecography 2004, 27, 285–298. [Google Scholar] [CrossRef]
- Ferreira, T.; Garcia, A. Native forests within and outside protected areas are key for nine-banded armadillo (Dasypus novemcinctus) occupancy in agricultural landscapes. Agric. Ecosyst. Environ. 2018, 266, 133–141. [Google Scholar]
- Loughry, W.J.; Perez-Heydrich, C.; McDonough, C.M.; Oli, M.K. Population dynamics and range expansion in nine-banded armadillos. PLoS ONE 2013, 8, e68311. [Google Scholar] [CrossRef]
- Changnon, S.A.; Angel, J.R.; Kunkel, K.E.; Lehmann, C.M. Climate Atlas of Illinois; Illinois State Water Survey: Champaign, IL, USA, 2004. [Google Scholar]
- Illinois State Geological Survey. Illinois State Geological Survey Illinois High and Low. Available online: https://www.isgs.illinois.edu/outreach/geology-resources/illinois-high-and-low (accessed on 8 April 2020).
- Illinois State Water Survey. Illinois Climate Normals–Illinois State Climatologist. Available online: https://stateclimatologist.web.illinois.edu/data/climate-data/ (accessed on 15 January 2020).
- Yang, L.; Jin, S.; Danielson, P.; Homer, C.; Gass, L.; Bender, S.M.; Case, A.; Costello, C.; Dewitz, J.; Fry, J.; et al. A new generation of the United States National Land Cover Database: Requirements, research priorities, design, and implementation strategies. ISPRS J. Photogramm. Remote Sens. 2018, 146, 108–123. [Google Scholar] [CrossRef]
- United States Census Bureau QuickFacts. Available online: https://www.census.gov/programs-surveys/sis/resources/data-tools/quickfacts.html (accessed on 8 April 2020).
- Homer, C.; Dewitz, J.; Jin, S.; Xian, G.; Costello, C.; Danielson, P.; Gass, L.; Funk, M.; Wickham, J.; Stehman, S.; et al. Conterminous United States land cover change patterns 2001–2016 from the 2016 National Land Cover Database. ISPRS J. Photogramm. Remote Sens. 2020, 162, 184–199. [Google Scholar] [CrossRef]
- Kennedy, C.M.; Oakleaf, J.R.; Theobald, D.M.; Baruch-Mordo, S.; Kiesecker, J. Managing the middle: A shift in conservation priorities based on the global human modification gradient. Glob. Chang. Biol. 2019, 25, 1–16. [Google Scholar] [CrossRef]
- United States Geological Survey. Core Science Analytics, Synthesis, and Library-GAP Analysis Project. Available online: https://www.usgs.gov/core-science-systems/science-analytics-and-synthesis/gap/data-tools (accessed on 21 August 2020).
- United States Department of Agriculture National Resources Conservation Service USDA NRCS Geospatial Data Gateway. Available online: https://datagateway.nrcs.usda.gov/ (accessed on 21 August 2020).
- Baldwin, R.A. Use of maximum entropy modeling in wildlife research. Entropy 2009, 11, 854–866. [Google Scholar] [CrossRef]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Townsend Peterson, A. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Brown, J.L.; Bennett, J.R.; French, C.M. SDMtoolbox 2.0: The next generation Python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. PeerJ 2017, 5, e4095. [Google Scholar] [CrossRef] [PubMed]
- Radosavljevic, A.; Anderson, R.P. Making better Maxent models of species distributions: Complexity, overfitting and evaluation. J. Biogeogr. 2013, 41, 629–643. [Google Scholar] [CrossRef]
- Anderson, R.P.; Gonzalez, I. Species-specific tuning increases robustness to sampling bias in models of species distributions: An implementation with Maxent. Ecol. Model. 2011, 222, 2796–2811. [Google Scholar] [CrossRef]
- Brown, J.L. SDMtoolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberón, J.; Pearson, R.G.; Anderson, R.P.; Martínez-Meyer, E.; Nakamura, M.; Araújo, M.B. Ecological Niches and Geographic Distributions; Princeton University Press: Princeton, NJ, USA, 2011. [Google Scholar]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef]
- Shcheglovitova, M.; Anderson, R.P. Estimating optimal complexity for ecological niche models: A jackknife approach for species with small sample sizes. Ecol. Model. 2013, 269, 9–17. [Google Scholar] [CrossRef]
- McDonough, C.M.; Loughry, W.J. Patterns of mortality in a population of nine-banded armadillos, Dasypus novemcinctus. Am. Midl. Nat. 1997, 138, 299–305. [Google Scholar] [CrossRef]
- Talmage, R.; Buchanan, G. The Armadillo (Dasypus novemcinctus). Rice Inst. Pam. 1954, 41, 1–135. [Google Scholar]
- Tomasek, B.J.; Williams, M.M.; Davis, A.S. Changes in field workability and drought risk from projected climate change drive spatially variable risks in Illinois cropping systems. PLoS ONE 2017, 12, e0172301. [Google Scholar] [CrossRef] [PubMed]
- Crane, A.L.; McGrane, C.E.; Mathis, A. Behavioral and physiological responses of Ozark zigzag salamanders to stimuli from an invasive predator: The armadillo. Int. J. Ecol. 2012, 2012, 1–7. [Google Scholar] [CrossRef]
- Mullin, S.J.; Klueh, S. Demographics of a geographically isolated population of a threatened salamander (Caudata: Ambystomatidae) in central Illinois. Herpetol. Conserv. Biol. 2009, 4, 261–269. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Climatic Factor Code | Title | Description |
|---|---|---|
| BIO 02 | Mean Diurnal Range | Mean of the monthly difference between maximum and minimum temperatures |
| BIO 08 | Mean Temperature of the Wettest Quarter | Mean temperature of the three-month period receiving the greatest amount of precipitation |
| BIO 12 | Annual Precipitation | Total precipitation in a year |
| BIO 18 | Precipitation of the Warmest Quarter | Total precipitation during the three-month period with the highest temperatures |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haywood, C.J.; Nielsen, C.K.; Jiménez, F.A. Potential Distribution of Colonizing Nine-Banded Armadillos at Their Northern Range Edge. Diversity 2021, 13, 266. https://doi.org/10.3390/d13060266
Haywood CJ, Nielsen CK, Jiménez FA. Potential Distribution of Colonizing Nine-Banded Armadillos at Their Northern Range Edge. Diversity. 2021; 13(6):266. https://doi.org/10.3390/d13060266
Chicago/Turabian StyleHaywood, Carly J., Clayton K. Nielsen, and F. Agustín Jiménez. 2021. "Potential Distribution of Colonizing Nine-Banded Armadillos at Their Northern Range Edge" Diversity 13, no. 6: 266. https://doi.org/10.3390/d13060266
APA StyleHaywood, C. J., Nielsen, C. K., & Jiménez, F. A. (2021). Potential Distribution of Colonizing Nine-Banded Armadillos at Their Northern Range Edge. Diversity, 13(6), 266. https://doi.org/10.3390/d13060266