
Spatial Dynamics of Two Host-Parasite Relationships on Intertidal Oyster Reefs


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites and Sampling Methods
2.2. Statistical Analysis
3. Results
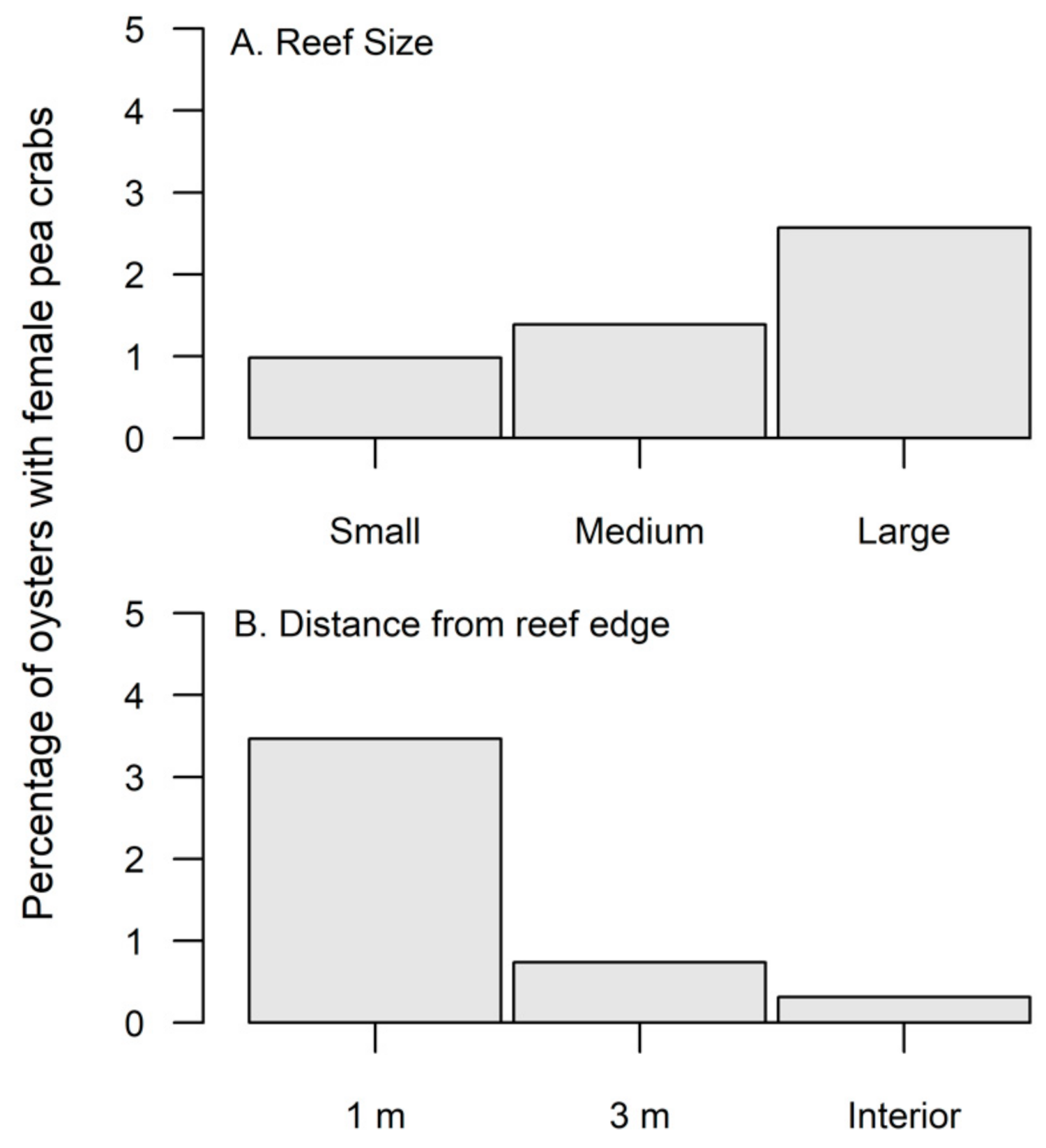
3.1. Oysters and Pea Crabs
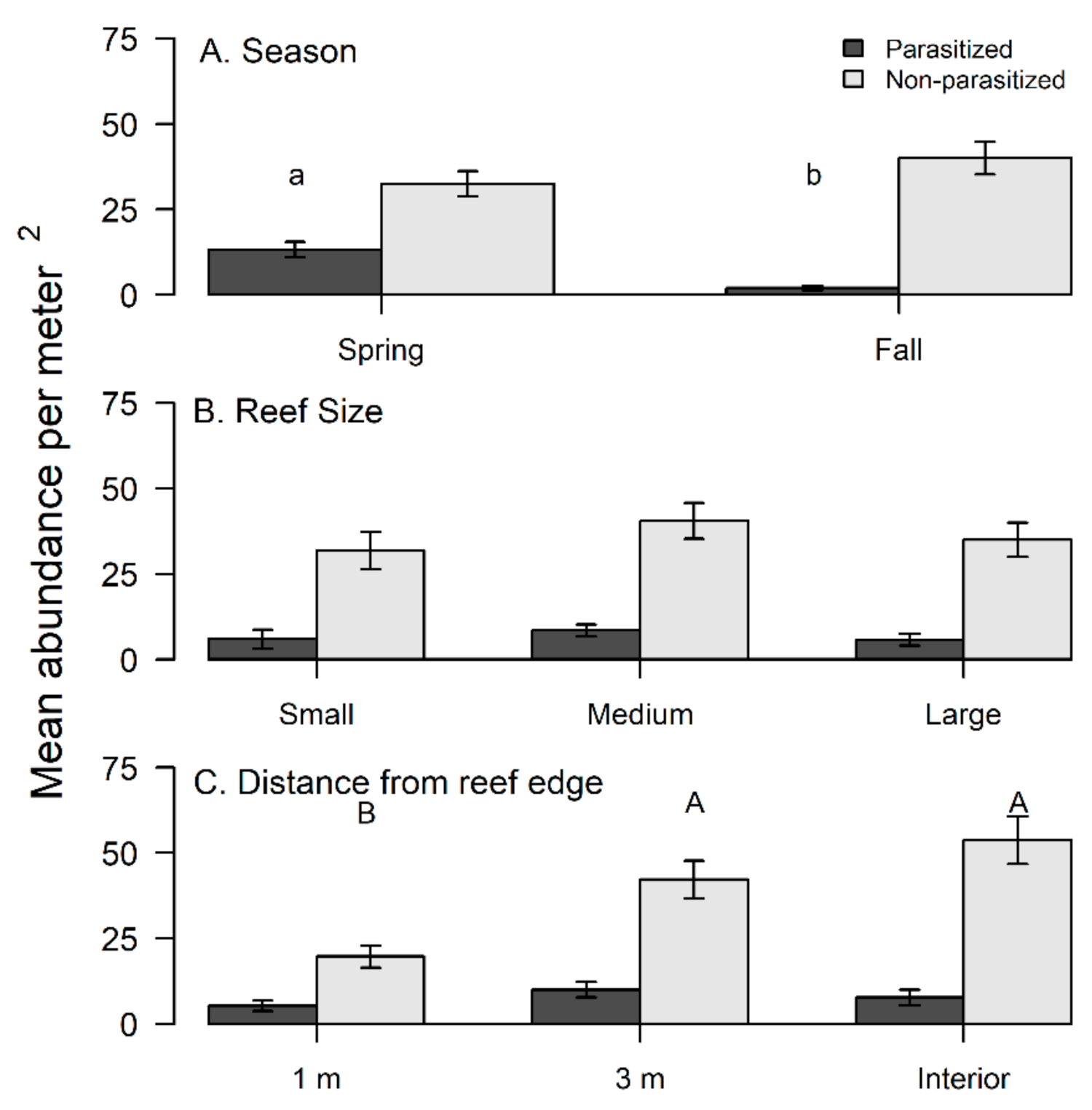
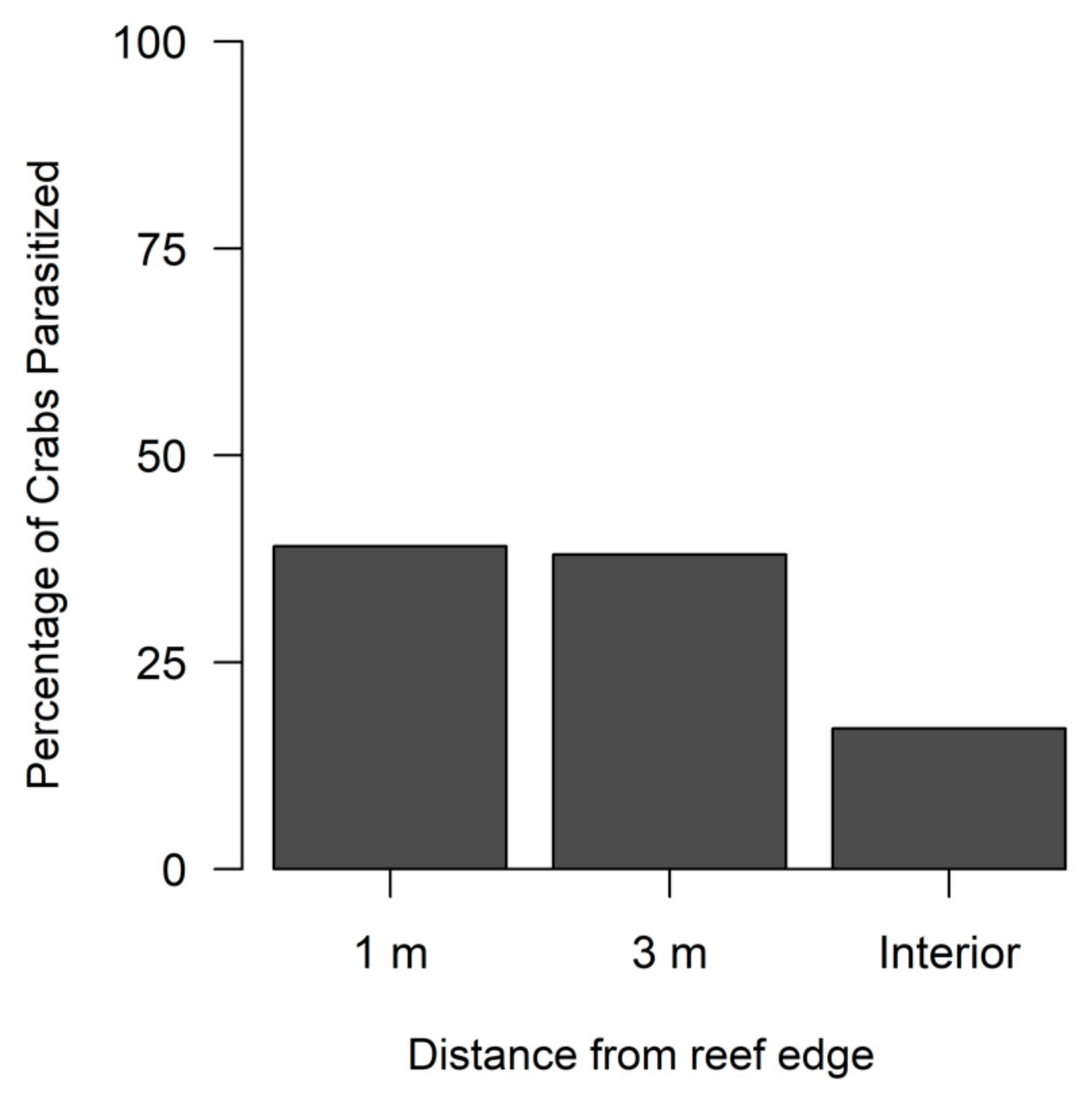
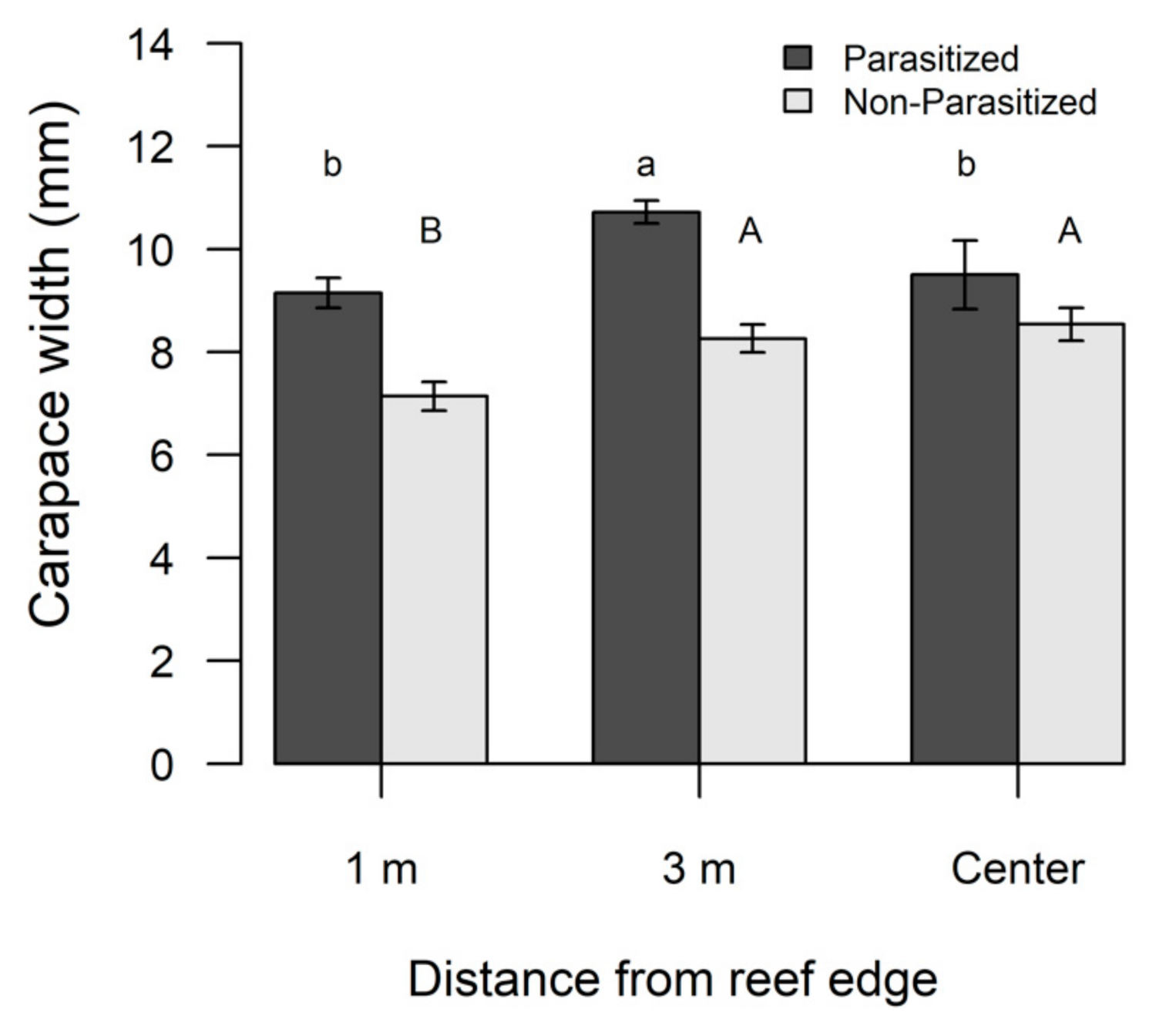
3.2. Xanthid Crabs and Rhizocephalans
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fagan, W.F.; Cantrell, R.S.; Cosner, C. How habitat edges change species interactions. Am. Nat. 1999, 153, 165–182. [Google Scholar] [CrossRef] [PubMed]
- Tews, J.; Brose, U.; Grimm, V.; Tielbörger, K.; Wichmann, M.C.; Schwager, M.; Jeltsch, F. Animal species diversity driven by habitat heterogeneity/diversity: The importance of keystone structures. J. Biogeogr. 2004, 31, 79–92. [Google Scholar] [CrossRef]
- Loreau, M.; Naeem, S.; Inchausti, P.; Bengtsson, J.; Grime, J.P.; Hector, A.; Hooper, D.U.; Huston, M.; Raffaelli, D.; Schmid, B.; et al. Biodiversity and ecosystem functioning: Current knowledge and future challenges. Science 2001, 294, 804–808. [Google Scholar] [CrossRef] [PubMed]
- Tscharntke, T.; Tylianakis, J. Landscape moderation of biodiversity patterns and processes-eight hypotheses. Biol. Rev. 2012, 87, 661–685. [Google Scholar] [CrossRef]
- Fahrig, L. Rethinking patch size and isolation effects: The habitat amount hypothesis. J. Biogeogr. 2013, 40, 1649–1663. [Google Scholar] [CrossRef]
- Holyoak, M. Habitat patch arrangement and metapopulation persistence of predators and prey. Am. Nat. 2000, 156, 378–389. [Google Scholar] [CrossRef]
- Martinson, H.; Fagan, W.; Denno, R. Critical patch sizes for food-web modules. Ecology 2012, 93, 1779–1786. [Google Scholar] [CrossRef]
- Ewers, R.; Thorpe, S.; Didham, R. Synergistic interactions between edge and area effects in a heavily fragmented landscape. Ecology 2007, 88, 96–106. [Google Scholar] [CrossRef]
- Thomas, C.D. Dispersal and extinction in fragmented landscapes. Proc. R. Soc. B Biol. Sci. 2000, 267, 139–145. [Google Scholar] [CrossRef]
- Haila, Y. A conceptual genealogoy of fragmentation research: From island biogeography to landscape ecology. Ecol. Appl. 2002, 12, 321–334. [Google Scholar]
- Cronin, J.T. Habitat edges, within-patch dispersion of hosts, and parasitoid oviposition behavior. Ecology 2009, 90, 196–207. [Google Scholar] [CrossRef]
- Hussain, S.; Ram, M.S.; Kumar, A.; Shivaji, S.; Umapathy, G. Human presence increases parasitic load in endangered lion-tailed macaques (Macaca silenus) in its fragmented rainforest habitats in Southern India. PLoS ONE 2013, 8, e63685. [Google Scholar] [CrossRef]
- Pullan, R.L.; Sturrock, H.J.; Soares Magalhães, R.J.; Clements, A.C.; Brooker, S.J. Spatial parasite ecology and epidemiology: A review of methods and applications. Parasitology 2012, 139, 1870–1887. [Google Scholar] [CrossRef]
- Arneberg, P.; Skorping, A.; Grenfell, B.; Read, A.F. Host densities as determinant of abundance in parasite communities. Proc. R. Soc. B Biological Sci. 1998, 265, 1283–1289. [Google Scholar] [CrossRef]
- Ryder, J.J.; Miller, M.R.; White, A.; Knell, R.J.; Boots, M. Host-parasite population dynamics under combined frequency- and density-dependent transmission. Oikos 2007, 116, 2017–2026. [Google Scholar] [CrossRef]
- Stiling, P.D. The frequency of density dependence in insect host-parasitoid systems. Ecology 1987, 68, 844–856. [Google Scholar] [CrossRef]
- Mbora, D.N.M.; McPeek, M.A. Host density and human activities mediate increased parasite prevalence and richness in primates threatened by habitat loss and fragmentation. J. Anim. Ecol. 2009, 78, 210–218. [Google Scholar] [CrossRef]
- Fahrig, L.; Paloheimo, J. Effect of Spatial Arrangement of Habitat Patches on Local Population Size. Ecology 1988, 69, 468–475. [Google Scholar] [CrossRef]
- Fahrig, L. Effects of habitat fragmentaion on biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef]
- Sullivan, L.L.; Johnson, B.L.; Brudvig, L.A.; Haddad, N.M. Can dispersal mode predict corridor effects on plant parasites? Ecology 2011, 92, 1559–1564. [Google Scholar] [CrossRef]
- Bordes, F.; Morand, S.; Pilosof, S.; Claude, J.; Krasnov, B.R.; Cosson, J.F.; Chaval, Y.; Ribas, A.; Chaisiri, K.; Blasdell, K.; et al. Habitat fragmentation alters the properties of a host-parasite network: Rodents and their helminths in South-East Asia. J. Anim. Ecol. 2015, 84, 1253–1263. [Google Scholar] [CrossRef] [PubMed]
- Lopez, J.E.; Gallinot, L.P.; Wade, M.J. Spread of parasites in metapopulations: An experimental study of the effects of host migration rate and local host population size. Parasitology 2005, 130, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Renwick, A.R.; Lambin, X. Host-parasite interactions in a fragmented landscape. Int. J. Parasitol. 2013, 43, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Lloyd-Smith, J.O.; Cross, P.C.; Briggs, C.J.; Daugherty, M.; Getz, W.M.; Latto, J.; Sanchez, M.S.; Smith, A.B.; Swei, A. Should we expect population thresholds for wildlife disease? Trends Ecol. Evol. 2005, 20, 511–519. [Google Scholar] [CrossRef]
- Ries, L.; Fletcher Jr, R.; Battin, J.; Sisk, T.D. Ecological response to habitat edges: Mechanisms, models, and variability explained. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 491–522. [Google Scholar] [CrossRef]
- Gates, J.E.; Gysel, L.W. Avian nest dispersion and fledging success in field-forest ecotones. Ecology 1978, 59, 871–883. [Google Scholar] [CrossRef]
- Patten, M.; Shochat, E.; Reinking, D.L.; Wolfe, D.H.; Sherrod, S.K. Habitat edge, land management, and rates of brood parasitism in tallgrass prairie. Ecol. Appl. 2006, 16, 687–695. [Google Scholar] [CrossRef]
- Donovan, T.; Jones, P.; Annand, E.; Thompson, F.T., III. Variation in local-scale edge effects: Mechanisms and landscape context. Ecology 1997, 78, 2064–2075. [Google Scholar] [CrossRef]
- Ries, L.; Sisk, T. A predictive model of edge effects. Ecology 2004, 85, 2917–2926. [Google Scholar] [CrossRef]
- Peterson, M.S.; Andres, M.J. Progress on Research Regarding Ecology and Biodiversity of Coastal Fisheries and Nektonic Species and Their Habitats within Coastal Landscapes. Diversity 2021, 13, 168. [Google Scholar] [CrossRef]
- Meyer, D.L.; Townsend, E.C. Faunal utilization of created intertidal eastern oyster (Crassostrea virginica) reefs in the southeastern United States. Estuaries 2000, 23, 34–45. [Google Scholar] [CrossRef]
- Grabowski, J.; Powers, S. Habitat complexity mitigates trophic transfer on oyster reefs. Mar. Ecol. Prog. Ser. 2004, 277, 291–295. [Google Scholar] [CrossRef]
- Peterson, C.; Grabowski, J.; Powers, S. Estimated enhancement of fish production resulting from restoring oyster reef habitat: Quantitative valuation. Mar. Ecol. Prog. Ser. 2003, 264, 249–264. [Google Scholar] [CrossRef]
- Coen, L.; Brumbaugh, R.; Bushek, D.; Grizzle, R.; Luckenbach, M.; Posey, M.; Powers, S.; Tolley, S. Ecosystem services related to oyster restoration. Mar. Ecol. Prog. Ser. 2007, 341, 303–307. [Google Scholar] [CrossRef]
- Hanke, M.H.; Posey, M.H.; Alphin, T.D. The influence of habitat characteristics on intertidal oyster Crassostrea virginica populations. Mar. Ecol. Prog. Ser. 2017, 571, 121–138. [Google Scholar] [CrossRef]
- Hanke, M.H.; Posey, M.H.; Alphin, T.D. The effects of intertidal oyster reef habitat characteristics on faunal utilization. Mar. Ecol. Prog. Ser. 2017, 581, 57–70. [Google Scholar] [CrossRef]
- Lenihan, H.; Peterson, C. How habitat degradation through fishery disturbance enhances impacts of hypoxia on oyster reefs. Ecol. Appl. 1998, 8, 128–140. [Google Scholar] [CrossRef]
- Coen, L.D.; Luckenbach, M.W. Developing success criteria and goals for evaluating oyster reef restoration: Ecological function or resource exploitation? Ecol. Eng. 2000, 15, 323–343. [Google Scholar] [CrossRef]
- Beck, M.W.; Brumbaugh, R.D.; Airoldi, L.; Carranza, A.; Coen, L.D.; Crawford, C.; Defeo, O.; Edgar, G.J.; Hancock, B.; Kay, M.C.; et al. Oyster reefs at risk and recommendations for conservation, restoration, and management. Bioscience 2011, 61, 107–116. [Google Scholar] [CrossRef]
- Harwell, H.D.; Posey, M.H.; Alphin, T.D. Landscape aspects of oyster reefs: Effects of fragmentation on habitat utilization. J. Exp. Mar. Bio. Ecol. 2011, 409, 30–41. [Google Scholar] [CrossRef]
- Macreadie, P.; Geraldi, N.; Peterson, C. Preference for feeding at habitat edges declines among juvenile blue crabs as oyster reef patchiness increases and predation risk grows. Mar. Ecol. Prog. Ser. 2012, 466, 145–153. [Google Scholar] [CrossRef]
- Eggleston, D.B.; Etherington, L.L.; Elis, W.E. Organism response to habitat patchiness: Species and habitat-dependent recruitment of decapod crustaceans. J. Exp. Mar. Bio. Ecol. 1998, 223, 111–132. [Google Scholar] [CrossRef]
- Eggleston, D.B.; Elis, W.E.; Etherington, L.L.; Dahlgren, C.P.; Posey, M.H. Organism responses to habitat fragmentation and diversity: Habitat colonization by estuarine macrofauna. J. Exp. Mar. Bio. Ecol. 1999, 236, 107–132. [Google Scholar] [CrossRef]
- Griffitt, J.; Posey, M.H.; Alphin, T.D. Effects of edge fragmentation on oyster reef utilization by transient nekton. J. Elisha Mitchell Sci. Soc. 1999, 115, 98–103. [Google Scholar]
- Stauber, L. Pinnotheres ostreum, parasitic on the American oyster, Ostrea (Gryphaea) virginica. Biol. Bull. 1945, 88, 269–291. [Google Scholar] [CrossRef]
- Christensen, A.; McDermott, J. Life-history and biology of the oyster crab, Pinnotheres ostreum Say. Biol. Bull. 1958, 114, 146–179. [Google Scholar] [CrossRef]
- Hanke, M.H.; Hargrove, J.M.; Alphin, T.D.; Posey, M.H. Oyster utilization and host variation of the oyster pea crab (Zaops ostreum). J. Shellfish Res. 2015, 34, 281–287. [Google Scholar] [CrossRef]
- Haven, D. The pea crab Pinnotheres ostreum as a parasite of the oyster. Va. J. Sci. 1958, 8, 301–302. [Google Scholar]
- Mercado-Silva, N. Condition index of the eastern oyster, Crassostrea virginica (Gmelin, 1791) in Sapelo Island Georgia-Effects of site, position on bed and pea crab parasitism. J. Shellfish Res. 2005, 24, 121–126. [Google Scholar]
- Byers, J.E.; Rogers, T.L.; Grabowski, J.H.; Hughes, R.A.; Piehler, M.F.; Kimbro, D.L. Host and parasite recruitment correlated at a regional scale. Oecologia 2014, 174, 731–738. [Google Scholar] [CrossRef]
- Hines, A.; Alvarez, F.; Reed, S. Introduced and native populations of a marine parasitic castrator: Variation in prevalence of the rhizocephalan Loxothylacus panopaei in xanthid crabs. Bull. Mar. Sci. 1997, 61, 197–214. [Google Scholar]
- Tolley, S.G.; Winstead, J.T.; Haynes, L.; Volety, A.K. Influence of salinity on prevalence of the parasite Loxothylacus panopaei in the xanthid Panopeus obesus in SW Florida. Dis. Aquat. Organ. 2006, 70, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Freeman, A.; Blakeslee, A.; Fowler, A. Northward expansion of the rhizocephalan Loxothylacus panopaei (Gissler, 1884) in the northwest Atlantic. Aquat. Invasions 2013, 8, 347–353. [Google Scholar] [CrossRef]
- Kruse, I.; Hare, M.P.; Hines, A.H. Genetic relationships of the marine invasive crab parasite Loxothylacus panopaei: An analysis of DNA sequence variation, host specificity, and distributional range. Biol. Invasions 2011, 14, 701–715. [Google Scholar] [CrossRef]
- Alvarez, F.; Hines, A.H.; Reaka-kudla, M.L. The effects of parasitism by the barnacle Loxothylacus panopaei (Gissler) (Cirripedia: Rhizocephala) on growth and survival of the host crab Rhithropanopeus harrisii (Gould) (Brachyura: Xanthidae). J. Exp. Mar. Bio. Ecol. 1995, 192, 221–232. [Google Scholar] [CrossRef]
- Brothers, C.A.; Blakeslee, A.M.H. Alien vs predator play hide and seek: How habitat complexity alters parasite mediated host survival. J. Exp. Mar. Bio. Ecol. 2021, 535, 151488. [Google Scholar] [CrossRef]
- O’Shaughnessy, K.A.; Harding, J.M.; Burge, E.J. Ecological effects of the invasive parasite Loxothylacus panopaei on the flatback mud crab Eurypanopeus depressus with implications for estuarine communities. Bull. Mar. Sci. 2014, 90, 611–621. [Google Scholar] [CrossRef]
- Grosholz, E.D.; Ruiz, G.M. Does spatial heterogeneity and genetic variation in populations of the xanthid crab Rhithropanopeus harrisii (Gould) influence the prevalence of an introduced parasitic castrator? J. Exp. Mar. Bio. Ecol. 1995, 187, 129–145. [Google Scholar] [CrossRef]
- Kulp, R.; Politano, V.; Lane, H. Predation of juvenile Crassostrea virginica by two species of mud crabs found in the Chesapeake Bay. J. Shellfish Res. 2011, 30, 1–6. [Google Scholar] [CrossRef]
- Toscano, B.J.; Newsome, B.; Griffen, B.D. Parasite modification of predator functional response. Oecologia 2014, 175, 345–352. [Google Scholar] [CrossRef]
- Meyer, D.L. Habitat partitioning between the Xanthid crabs Panopeus herbstii and Eurypanopeus depressus on intertidal oyster reefs (Crassostrea virginica) in southeastern North Carolina. Estuaries 1994, 17, 674–679. [Google Scholar] [CrossRef]
- Silliman, B.R.; Layman, C.A.; Geyer, K.; Zieman, J.C. Predation by the black-clawed mud crab, Panopeus herbstii, in Mid-Atlantic salt marshes: Further evidence for top-down control of marsh grass production. Estuaries 2004, 27, 188–196. [Google Scholar] [CrossRef]
- Robinson, E.; Lunt, J.; Marshall, C.; Smee, D. Eastern oysters Crassostrea virginica deter crab predators by altering their morphology in response to crab cues. Aquat. Biol. 2014, 20, 111–118. [Google Scholar] [CrossRef]
- Toscano, B.; Griffen, B. Predatory crab size diversity and bivalve consumption in oyster reefs. Mar. Ecol. Prog. Ser. 2012, 445, 65–74. [Google Scholar] [CrossRef]
- Grabowski, J. Habitat complexity disrupts predator-prey interactions but not the trophic cascade on oyster reefs. Ecology 2004, 85, 995–1004. [Google Scholar] [CrossRef]
- Kruse, I.; Hare, M.P. Genetic diversity and expanding nonindigenous range of the rhizocephalan Loxothylacus panopaei parasitizing mud crabs in the western north Atlantic. J. Parasitol. 2007, 93, 575–582. [Google Scholar] [CrossRef]
- Glancy, T.; Frazer, T.; Cichra, C.; Lindberg, W. Comparative patterns of occupancy by decapod crustaceans in seagrass, oyster, and marsh-edge habitats in a northeast Gulf of Mexico estuary. Estuaries 2003, 26, 1291–1301. [Google Scholar] [CrossRef]
- Sloan, L.M.; Anderson, S.V.; Pernet, B. Kilometer-scale spatial variation in revalence of the rhizocephalan Lernaeodiscus porcellanae on the Porcelain Crab Petrolisthes cabrilloi. J. Crustac. Biol. 2010, 30, 159–166. [Google Scholar] [CrossRef][Green Version]
- Baggett, L.P.; Powers, S.P.; Brumbaugh, R.; Coen, L.D.; DeAngelis, B.; Greene, J.; Hancock, B.; Morlock, S. Oyster Habitat Restoration Monitoring and Assessment Handbook; The Nature Conservancy: Arlington, VA, USA, 2014; 96p. [Google Scholar]
- Kimbro, D.L.; Byers, J.E.; Grabowski, J.H.; Hughes, A.R.; Piehler, M.F. The biogeography of trophic cascades on US oyster reefs. Ecol. Lett. 2014, 17, 845–854. [Google Scholar] [CrossRef]
- McDonald, J. Divergent life history patterns in the po-occuring intertidal crabs Panopeus herbstii and Eurypanopeus depressus (Crustacea: Brachyura: Xanthidae). Mar. Ecol. Prog. Ser. 1982, 8, 173–180. [Google Scholar] [CrossRef]
- Carroll, J.M.; Dashiell, R.; Watts, J.C.; Hunter, E.A. Tidal level affects the prevalence and impacts of pests and parasites on oysters (Crassostrea virginica) on intertidal reefs in Georgia, USA. Mar. Biol. 2021, 168, 1–11. [Google Scholar] [CrossRef]
- Luckenbach, M.W.; Orth, R. A chemical defense in Crustacea? J. Exp. Mar. Bio. Ecol. 1990, 137, 79–87. [Google Scholar] [CrossRef]
- Hill, J.; Weissburg, M. Habitat complexity and predator size mediate interactions between intraguild blue crab predators and mud crab prey in oyster reefs. Mar. Ecol. Prog. Ser. 2013, 488, 209–219. [Google Scholar] [CrossRef]
- Gehman, A.L.M.; Byers, J.E. Non-native parasite enhances susceptibility of host to native predators. Oecologia 2017, 183, 919–926. [Google Scholar] [CrossRef]
- Takahashi, T.; Matsuura, S. Laboratory studies on molting and growth of the shore crab, Hemigrapsus sanguineus de Haan, parasitized by a rhizocephalan barnacle. Biol. Bull. 1994, 300–308. [Google Scholar] [CrossRef]
- Kennedy, V.S.; Breisch, L.L. Sixteen decades of polical management of the oyster fishery in Maryland’s Chesapeake Bay. J. Environ. Manag. 1983, 164, 153–171. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effects | df | F Value | p Value |
|---|---|---|---|
| Season | 1,220 | 6.05 | 0.01 |
| Size | 2,220 | 4.36 | 0.01 |
| Distance | 2,220 | 29.63 | <0.0001 |
| Season × Size | 2,220 | 1.97 | 0.14 |
| Season × Distance | 2,220 | 0.58 | 0.56 |
| Size × Distance | 3,220 | 0.57 | 0.63 |
| Season × Size × Distance | 3,220 | 0.77 | 0.51 |
| Effects | df | F Value | p Value |
|---|---|---|---|
| Season | 1,63 | 1.36 | 0.24 |
| Size | 2,63 | 0.97 | 0.38 |
| Distance | 2,63 | 3.32 | 0.07 |
| Season × Size | 2,63 | 6.02 | 0.004 |
| Season × Distance | 2,63 | 0.76 | 0.38 |
| Size × Distance | 3,63 | 0.73 | 0.39 |
| Season × Size × Distance | 3,63 | 1.76 | 0.18 |
| Effects | df | F Value | p Value |
|---|---|---|---|
| A. Non-parasitized | |||
| Season | 1,219 | 0.70 | 0.40 |
| Size | 2,219 | 0.32 | 0.72 |
| Distance | 2,219 | 9.34 | 0.0001 |
| Season × Size | 2,219 | 2.37 | 0.09 |
| Season × Distance | 2,219 | 0.63 | 0.53 |
| Size × Distance | 3,219 | 0.50 | 0.68 |
| Season × Size × Distance | 3,219 | 0.55 | 0.64 |
| B. Parasitized | |||
| Season | 1,219 | 20.38 | <0.0001 |
| Size | 2,219 | 0.53 | 0.58 |
| Distance | 2,219 | 1.17 | 0.31 |
| Season × Size | 2,219 | 0.42 | 0.65 |
| Season × Distance | 2,219 | 0.68 | 0.50 |
| Size × Distance | 3,219 | 0.20 | 0.89 |
| Season × Size × Distance | 3,219 | 0.18 | 0.90 |
| Effects | df | F Value | p Value |
|---|---|---|---|
| A. Non-parasitized | |||
| Season | 1,312 | 0.01 | 0.92 |
| Size | 2,312 | 1.47 | 0.23 |
| Distance | 2,312 | 3.22 | 0.04 |
| Season × Size | 2,312 | 0.98 | 0.37 |
| Season × Distance | 2,312 | 0.02 | 0.98 |
| Size × Distance | 2,312 | 0.32 | 0.80 |
| Season × Size × Distance | 3,312 | 1.01 | 0.39 |
| B. Parasitized | |||
| Season | 1,89 | 0.18 | 0.67 |
| Size | 2,89 | 0.03 | 0.97 |
| Distance | 2,89 | 6.49 | 0.002 |
| Season × Size | 2,89 | 1.52 | 0.22 |
| Season × Distance | 2,89 | 0.51 | 0.60 |
| Size × Distance | 2,89 | 1.50 | 0.22 |
| Season × Size × Distance | 3,89 | 0.47 | 0.70 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanke, M.H.; Posey, M.H.; Alphin, T.D. Spatial Dynamics of Two Host-Parasite Relationships on Intertidal Oyster Reefs. Diversity 2021, 13, 260. https://doi.org/10.3390/d13060260
Hanke MH, Posey MH, Alphin TD. Spatial Dynamics of Two Host-Parasite Relationships on Intertidal Oyster Reefs. Diversity. 2021; 13(6):260. https://doi.org/10.3390/d13060260
Chicago/Turabian StyleHanke, Marc H., Martin H. Posey, and Troy D. Alphin. 2021. "Spatial Dynamics of Two Host-Parasite Relationships on Intertidal Oyster Reefs" Diversity 13, no. 6: 260. https://doi.org/10.3390/d13060260
APA StyleHanke, M. H., Posey, M. H., & Alphin, T. D. (2021). Spatial Dynamics of Two Host-Parasite Relationships on Intertidal Oyster Reefs. Diversity, 13(6), 260. https://doi.org/10.3390/d13060260