
Contrasting Effects of Regional and Local Climate on the Interannual Variability and Phenology of the Scyphozoan, Aurelia coerulea and Nemopilema nomurai in the Korean Peninsula


Abstract
1. Introduction
2. Materials and Methods
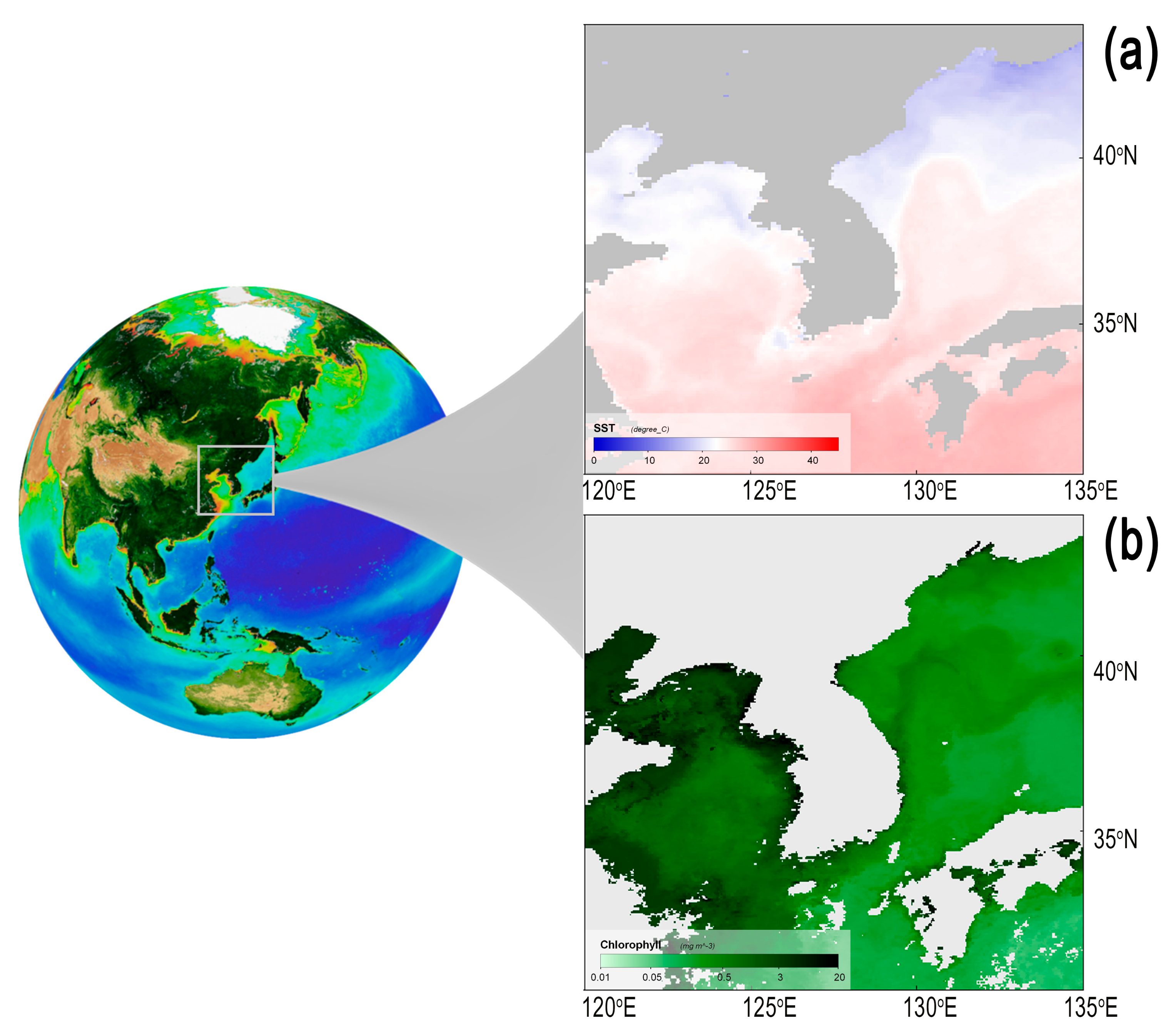
2.1. Hydroclimate Conditions in East Asian Marginal Seas
2.2. Biological Data
2.3. Physical Data
2.4. Statistical Analyses
3. Results and Discussion
3.1. Climate and Hydrographic Patterns in the Korean Peninsula
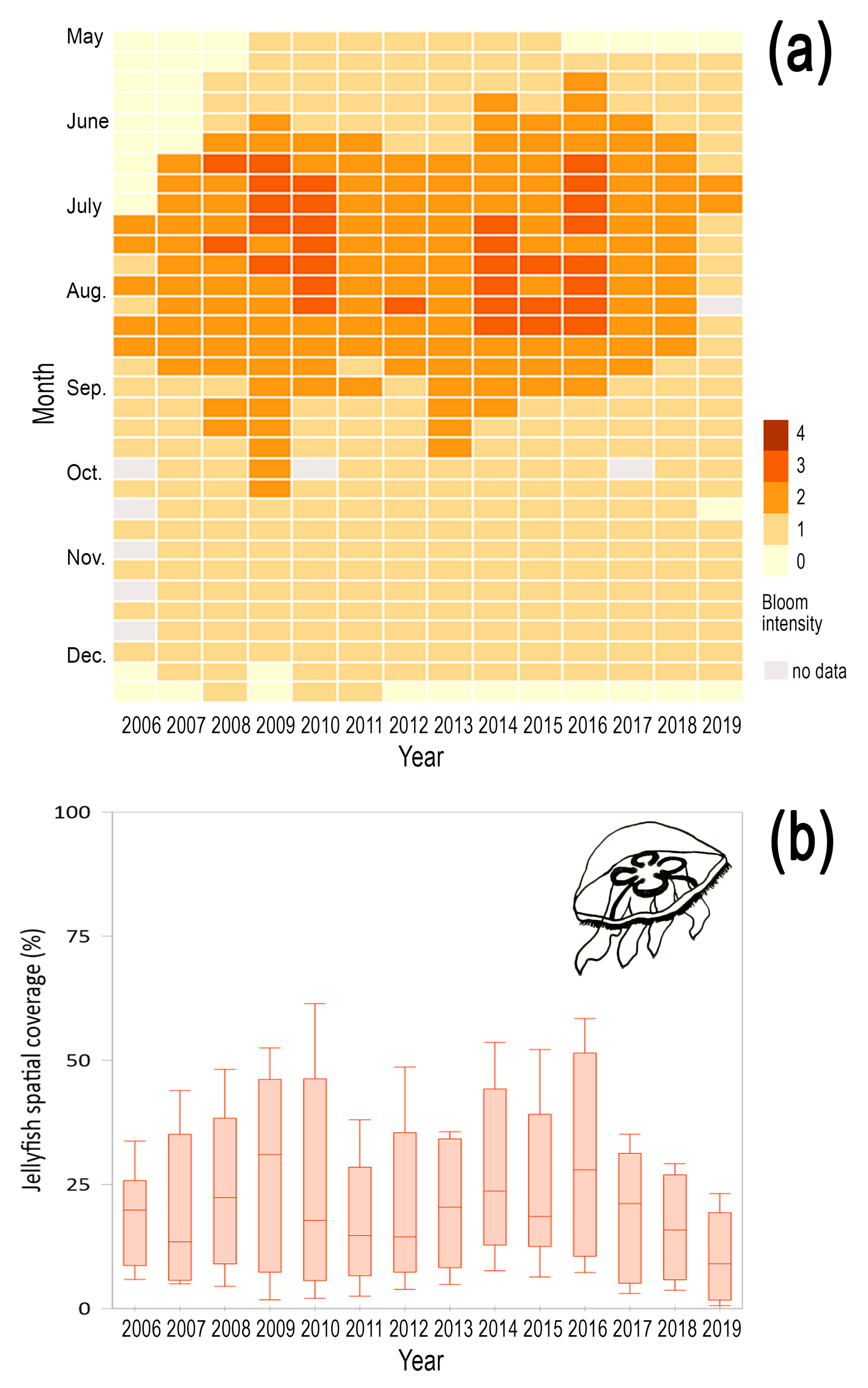
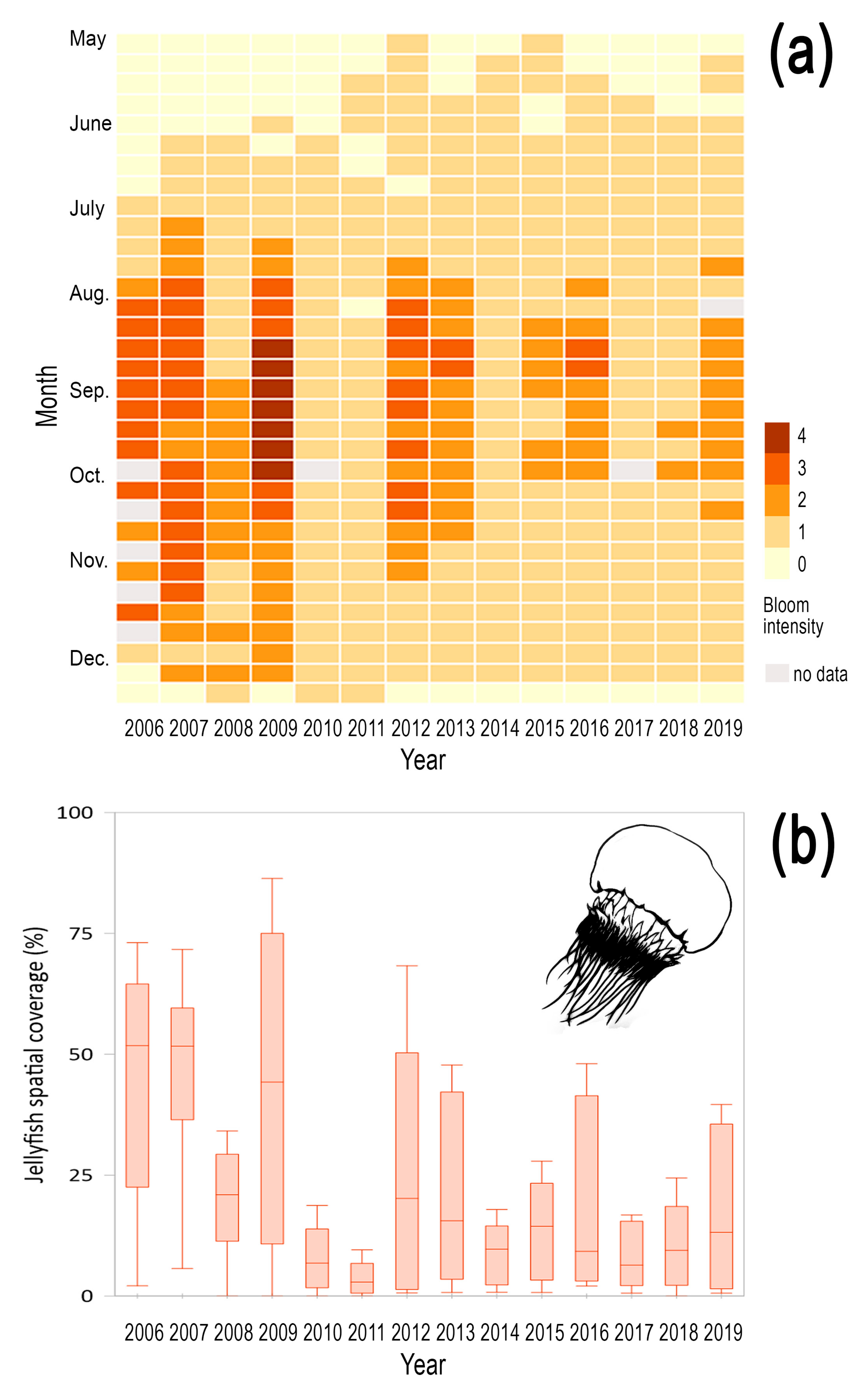
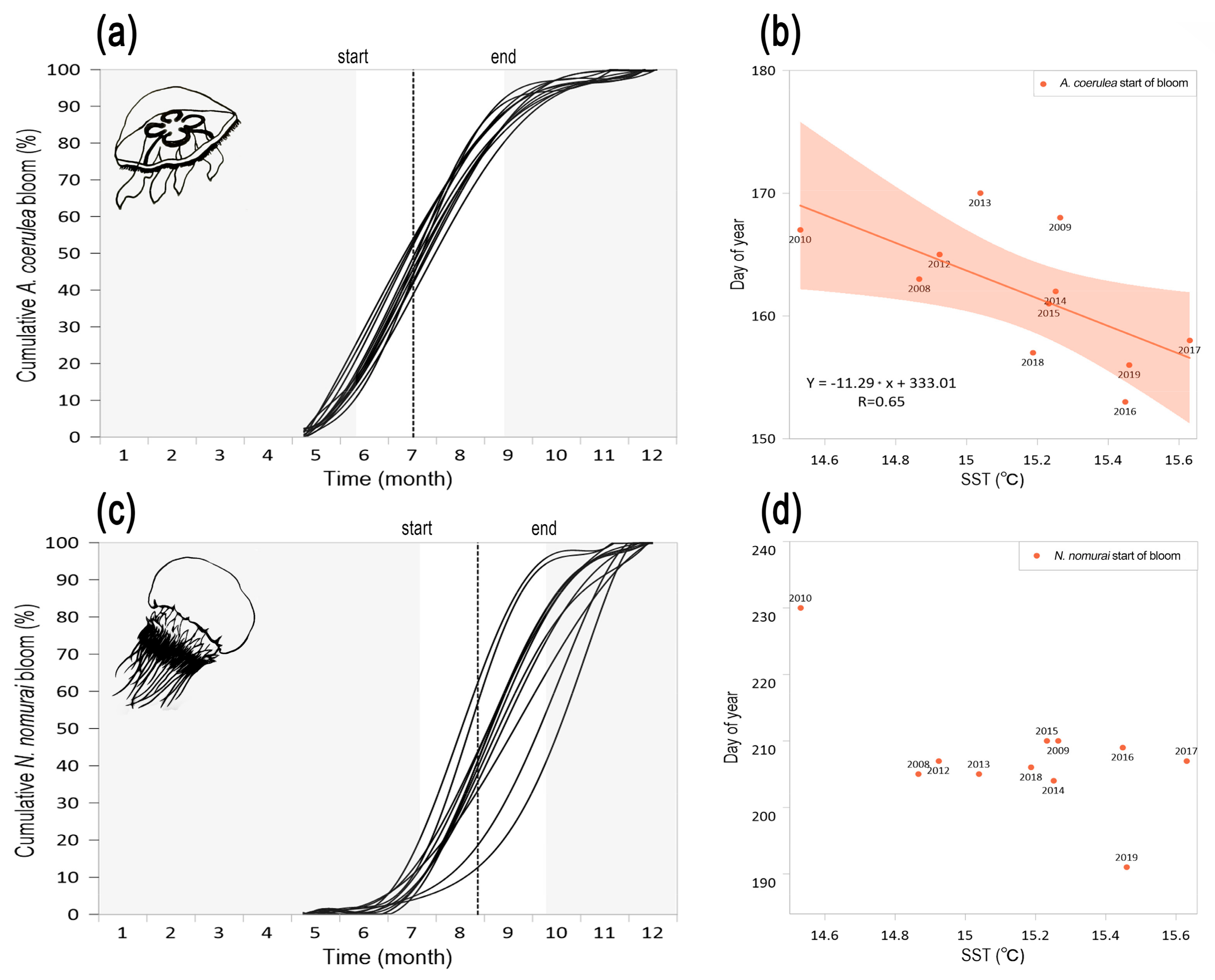
3.2. Interannual Variability of Magnitude and Timing of Jellyfish Blooms in Korean Waters
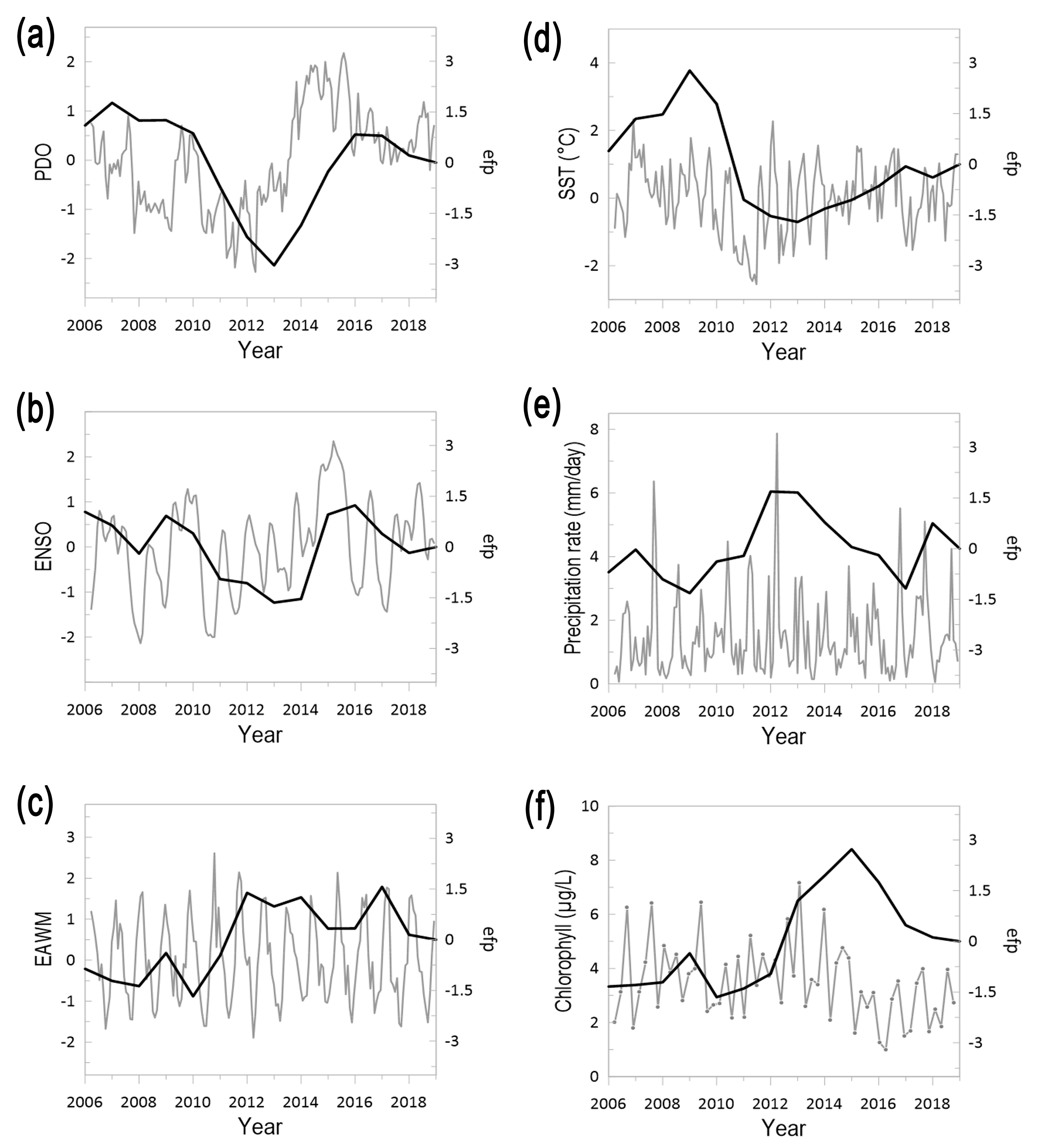
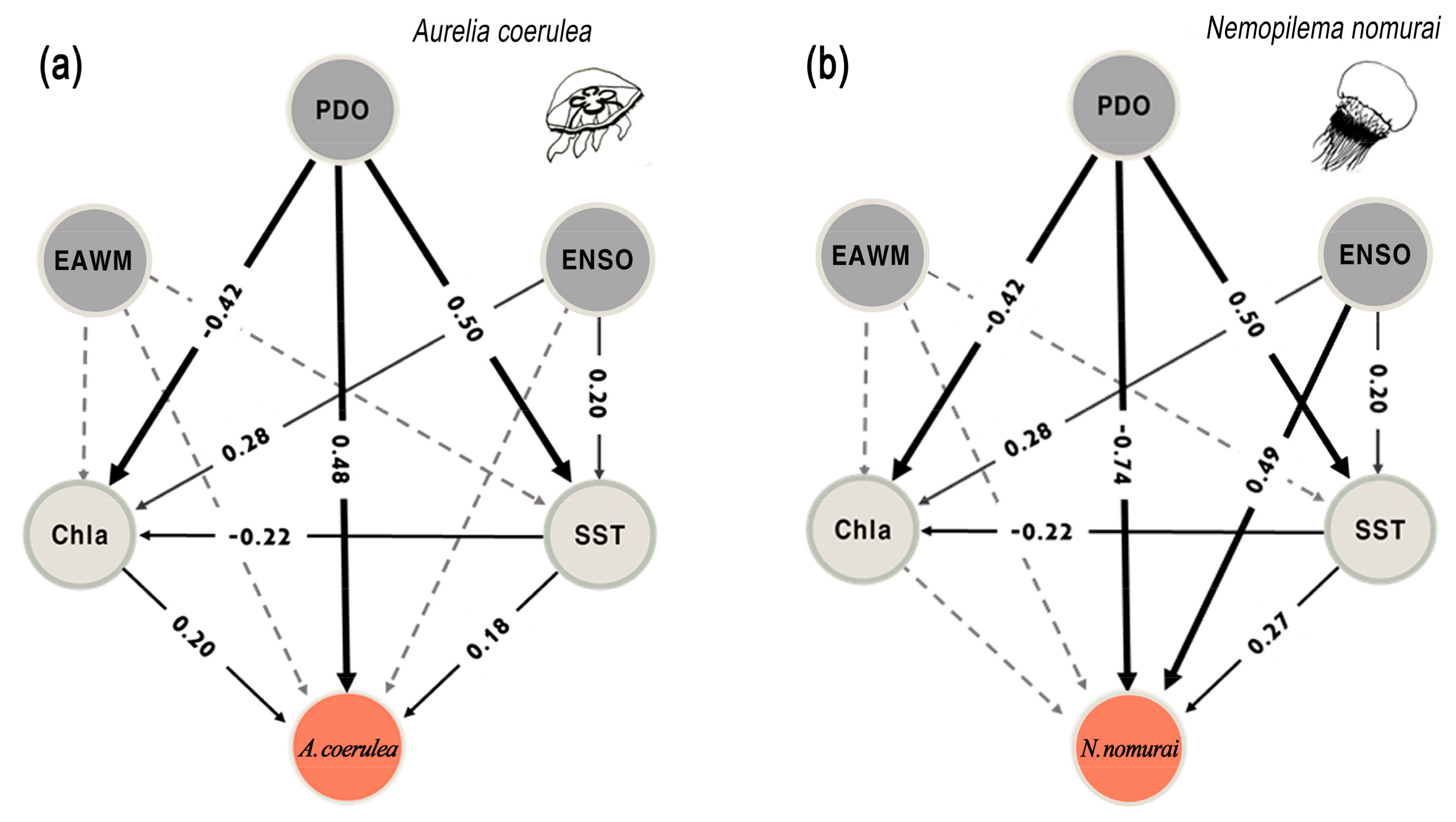
3.3. Climate and Jellyfish Dynamics in the Korean Peninsula
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Molinero, J.C.; Reygondeau, G.; Bonnet, D. Climate variance influence on the non-stationary plankton dynamics. Mar. Environ. Res. 2013, 89, 91–96. [Google Scholar] [CrossRef]
- Taylor, A.H.; Allen, J.I.; Clark, P.A. Extraction of a weak climatic signal by an ecosystem. Nature 2002, 416, 629–632. [Google Scholar] [CrossRef] [PubMed]
- Beaugrand, G.; Reid, P.C. Long-term changes in phytoplankton, zooplankton and salmon related to climate. Glob. Chang. Biol. 2003, 9, 801–817. [Google Scholar] [CrossRef]
- Bindoff, N.L.; Cheung, W.W.L.; Kairo, J.G.; Aristegui, J.; Guinder, V.A.; Hallberg, R.; Hilmi, N.; Jiao, N.; Karim, M.S.; Levin, L.; et al. Changing Ocean, Marine Ecosystems, and Dependent Communities. IPCC Spec. Rep. Ocean Cryosph. Chang. Clim. 2019, 447–588. [Google Scholar]
- Uye, S.I. Blooms of the giant jellyfish Nemopilema nomurai: A threat to the fisheries sustainability of the East Asian Marginal Seas. Plankt. Benthos Res. 2008, 3, 125–131. [Google Scholar] [CrossRef]
- Tseng, H.C.; You, W.L.; Huang, W.; Chung, C.C.; Tsai, A.Y.; Chen, T.Y.; Lan, K.W.; Gong, G.C. Seasonal Variations of Marine Environment and Primary Production in the Taiwan Strait. Front. Mar. Sci. 2020, 7, 1–12. [Google Scholar] [CrossRef]
- Jung, S.; Pang, I.C.; Lee, J.H.; Lee, K. Climate-change driven range shifts of anchovy biomass projected by bio-physical coupling individual based model in the marginal seas of East Asia. Ocean Sci. J. 2016, 51, 563–580. [Google Scholar] [CrossRef]
- Szuwalski, C. Comment on “Impacts of historical warming on marine fisheries production”. Science 2019, 365, 979–983. [Google Scholar] [CrossRef]
- Han, I.-S.; Lee, J.-S. Change the Annual Amplitude of Sea Surface Temperature due to Climate Change in a Recent Decade around the Korean Peninsula. J. Korean Soc. Mar. Environ. Saf. 2020, 26, 233–241. [Google Scholar] [CrossRef]
- Kim, D.-Y.; Lee, J.-S.; Kim, D.-H. A Study on Direction of Industrial Utilization for Jellyfish in Korea. J. Fish. Mar. Sci. Educ. 2014, 26, 587–596. [Google Scholar] [CrossRef][Green Version]
- National Institute of Fisheries Science. Report on the Jellyfish Appearance in Korean Coastal Waters in 2019, E2019-ME-004; National Institute of Fisheries Science Printing Office: Busan, Korea, 2019. [Google Scholar]
- Lee, J.H.; Choi, H.W.; Chae, J.; Kim, D.S.; Lee, S.B. Performance analysis of intake screens in power plants on mass impingement of marine organisms. Ocean Polar Res. 2006, 28, 385–393. [Google Scholar] [CrossRef]
- Yoon, E.-A.; Cha, C.-P.; Hwang, D.-J.; Yoon, Y.-H.; Shin, H.-H.; Gwak, D.-S. Inter-annual occurrence variation of the large jellyfish Nemopilema nomurai due to the changing marine environment in the East China Sea. J. Korean Soc. Fish. Technol. 2012, 48, 242–255. [Google Scholar] [CrossRef]
- Lucas, C.H. Reproduction and life history strategies of the common jellyfish, Aurelia aurita, in relation to its ambient environment. Hydrobiologia 2001, 451, 229–246. [Google Scholar] [CrossRef]
- Dawson, M.N. Macro-morphological variation among cryptic species of the moon jellyfish, Aurelia (Cnidaria: Scyphozoa). Mar. Biol. 2003, 143, 369–379. [Google Scholar] [CrossRef]
- Ki, J.S.; Hwang, D.S.; Shin, K.; Yoon, W.D.; Lim, D.; Kang, Y.S.; Lee, Y.; Lee, J.S. Recent moon jelly (Aurelia sp.1) blooms in Korean coastal waters suggest global expansion: Examples inferred from mitochondrial COI and nuclear ITS-5.8S rDNA sequences. ICES J. Mar. Sci. 2008, 65, 443–452. [Google Scholar] [CrossRef]
- Seo, Y.; Chae, J.; Ki, J.S. The complete mitochondrial genome of the jellyfish Aurelia coerulea (Cnidaria and Scyphozoa) with phylogenetic analysis. Mitochondrial DNA Part B Resour. 2020, 5, 1929–1930. [Google Scholar] [CrossRef]
- Dawson, M.N.; Jacobs, D.K. Molecular evidence for cryptic species of Aurelia aurita (Cnidaria, Scyphozoa). Biol. Bull. 2001, 200, 92–96. [Google Scholar] [CrossRef]
- Scorrano, S.; Aglieri, G.; Boero, F.; Dawson, M.N.; Piraino, S. Unmasking Aurelia species in the Mediterranean Sea: An integrative morphometric and molecular approach. Zool. J. Linn. Soc. 2017, 180, 243–267. [Google Scholar] [CrossRef]
- Kawahara, M.; Uye, S.I.; Ohtsu, K.; Iizumi, H. Unusual population explosion of the giant jellyfish Nemopilema nomurai (Scyphozoa: Rhizostomeae) in East Asian waters. Mar. Ecol. Prog. Ser. 2006, 307, 161–173. [Google Scholar] [CrossRef]
- Yoon, W.D.; Yang, J.Y.; Shim, M.B.; Kang, H.K. Physical processes influencing the occurrence of the giant jellyfish Nemopilema nomurai (Scyphozoa: Rhizostomeae) around Jeju Island, Korea. J. Plankton Res. 2008, 30, 251–260. [Google Scholar] [CrossRef][Green Version]
- Moon, J.H.; Pang, I.C.; Yang, J.Y.; Yoon, W.D. Behavior of the giant jellyfish Nemopilema nomurai in the East China Sea and East/Japan Sea during the summer of 2005: A numerical model approach using a particle-tracking experiment. J. Mar. Syst. 2010, 80, 101–114. [Google Scholar] [CrossRef]
- Kitajima, S.; Hasegawa, T.; Nishiuchi, K.; Kiyomoto, Y.; Taneda, T.; Yamada, H. Temporal fluctuations in abundance and size of the giant jellyfish Nemopilema nomurai medusae in the northern East China Sea, 2006–2017. Mar. Biol. 2020, 167, 1–10. [Google Scholar] [CrossRef]
- Kitajima, S.; Iguchi, N.; Honda, N.; Watanabe, T.; Katoh, O. Distribution of Nemopilema nomurai in the southwestern Sea of Japan related to meandering of the Tsushima Warm Current. J. Oceanogr. 2015, 71, 287–296. [Google Scholar] [CrossRef]
- Dong, Z.; Liu, D.; Keesing, J.K. Jellyfish blooms in China: Dominant species, causes and consequences. Mar. Pollut. Bull. 2010, 60, 954–963. [Google Scholar] [CrossRef] [PubMed]
- Yoon, W.D.; Lee, H.E.; Han, C.; Chang, S.J.; Lee, K. Abundance and distribution of Nemopilema nomurai (Scyphozoa, Rhizostomeae) in Korean waters in 2005–2013. Ocean Sci. J. 2014, 49, 183–192. [Google Scholar] [CrossRef]
- Park, Y.H.; Yoon, J.H.; Youn, Y.H.; Vivier, F. Recent warming in the western north pacific in relation to rapid changes in the atmospheric circulation of the Siberian high and aleutian low systems. J. Clim. 2012, 25, 3476–3493. [Google Scholar] [CrossRef]
- Park, W.S. Interannual and interdecadal variations of sea surface temperature in the East Asian Marginal Seas. Prog. Oceanogr. 2000, 47, 191–204. [Google Scholar] [CrossRef]
- Toda, M.; Watanabe, M. Mechanisms of enhanced ocean surface warming in the Kuroshio region for 1951–2010. Clim. Dyn. 2020, 54, 4129–4145. [Google Scholar] [CrossRef]
- Mantua, N.J.; Hare, S.R.; Zhang, Y.; Wallace, J.M.; Francis, R.C. A Pacific Interdecadal Climate Oscillation with Impacts on Salmon Production. Bull. Am. Meteorol. Soc. 1997, 78, 1069–1079. [Google Scholar] [CrossRef]
- Trenberth, K.E.; Stepaniak, D.P. Indices of El Niño evolution. J. Clim. 2001, 14, 1697–1701. [Google Scholar] [CrossRef]
- Jhun, J.G.; Lee, E.J. A new East Asian winter monsoon index and associated characteristics of the winter monsoon. J. Clim. 2004, 17, 711–726. [Google Scholar] [CrossRef]
- Zeileis, A.; Leisch, F.; Hornik, K.; Kleiber, C. Strucchange: An R package for testing for structural change in linear regression models. J. Stat. Softw. 2002, 7, 1–38. [Google Scholar] [CrossRef]
- Fernández de Puelles, M.L.; Molinero, J.C. Increasing zooplankton variance in the late 1990s unveils hydroclimate modifications in the Balearic Sea, Western Mediterranean. Mar. Environ. Res. 2013, 86, 76–80. [Google Scholar] [CrossRef]
- Greve, W.; Prinage, S.; Zidowitz, H.; Nast, J.; Reiners, F. On the phenology of North Sea ichthyoplankton. ICES J. Mar. Sci. 2005, 62, 1216–1223. [Google Scholar] [CrossRef]
- Alsterberg, C.; Eklöf, J.S.; Gamfeldt, L.; Havenhand, J.N.; Sundbäck, K. Consumers mediate the effects of experimental ocean acidification and warming on primary producers. Proc. Natl. Acad. Sci. USA 2013, 110, 8603–8608. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing, R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Borchers, H.W. Practical Numerical Math Functions. R Package Version 2.2.2. 2018. Available online: https://CRAN.R-project.org/package=pracma (accessed on 17 January 2021).
- Kolde, R. Pheatmap: Pretty Heatmaps. R Package Version 1.0.12. 2019. Available online: https://CRAN.R-project.org/package=pheatmap (accessed on 21 January 2021).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Kim, J.W.; Yeh, S.W.; Chang, E.C. Combined effect of El Niño-Southern Oscillation and Pacific Decadal Oscillation on the East Asian winter monsoon. Clim. Dyn. 2014, 42, 957–971. [Google Scholar] [CrossRef]
- D’Arrigo, R.; Wilson, R.; Panagiotopoulos, F.; Wu, B. On the long-term interannual variability of the east Asian winter monsoon. Geophys. Res. Lett. 2005, 32, 1–4. [Google Scholar] [CrossRef]
- Mantua, N.J. Pacific–Decadal Oscillation (PDO). Encycl. Glob. Environ. Chang. 2002, 1, 592–594. [Google Scholar]
- D’orgeville, M.; Peltier, W.R. On the Pacific Decadal Oscillation and the Atlantic Multidecadal Oscillation: Might they be related? Geophys. Res. Lett. 2007, 34, 1–5. [Google Scholar] [CrossRef]
- Mackas, D.L.; Greve, W.; Edwards, M.; Chiba, S.; Tadokoro, K.; Eloire, D.; Mazzocchi, M.G.; Batten, S.; Richardson, A.J.; Johnson, C.; et al. Changing zooplankton seasonality in a changing ocean: Comparing time series of zooplankton phenology. Prog. Oceanogr. 2012, 97–100, 31–62. [Google Scholar] [CrossRef]
- Chiba, S.; Batten, S.; Sasaoka, K.; Sasai, Y.; Sugisaki, H. Influence of the Pacific Decadal Oscillation on phytoplankton phenology and community structure in the western North Pacific. Geophys. Res. Lett. 2012, 39, 2–7. [Google Scholar] [CrossRef]
- Zhou, X.; Sun, Y.; Huang, W.; Smol, J.P.; Tang, Q.; Sun, L. The Pacific decadal oscillation and changes in anchovy populations in the Northwest Pacific. J. Asian Earth Sci. 2015, 114, 504–511. [Google Scholar] [CrossRef]
- Shin, A.; Yoon, S.C.; Lee, S.I.; Park, H.W.; Kim, S. The relationship between fishing characteristics of Pacific bluefin tuna (Thunnus orientalis) and ocean conditions around Jeju Island. Fish. Aquat. Sci. 2018, 21, 1–12. [Google Scholar] [CrossRef]
- Molinero, J.C.; Tseng, L.C.; Lopez-Lopez, L.L.; Sommer, U.; Souissi, S.; Hwang, J.S. Climate-driven winter variations of Calanus sinicus abundance in the East China Sea. Fish. Oceanogr. 2016, 25, 555–564. [Google Scholar] [CrossRef]
- Molinero, J.C.; Tseng, L.C.; Abbate, C.L.; Ramirez-Romero, E.; Hwang, J.S. Interannual changes in zooplankton echo subtropical and high latitude climate effects in the southern East China Sea. PLoS ONE 2018, 13, e0197382. [Google Scholar] [CrossRef]
- Goldstein, J.; Augustin, C.B.; Bleich, S.; Holst, S. A matter of tolerance: Distribution potential of scyphozoan polyps in a changing environment. Mar. Ecol. 2017, 38, 1–10. [Google Scholar] [CrossRef]
- Loveridge, A.; Lucas, C.H.; Pitt, K.A. Shorter, warmer winters may inhibit production of ephyrae in a population of the moon jellyfish Aurelia aurita. Hydrobiologia 2021, 848, 739–749. [Google Scholar] [CrossRef]
- Wang, Y.T.; Zheng, S.; Sun, S.; Zhang, F. Effect of temperature and food type on asexual reproduction in Aurelia sp.1 polyps. Hydrobiologia 2015, 754, 169–178. [Google Scholar] [CrossRef]
- Dong, Z.; Wang, L.; Liu, Q.; Sun, T. Effects of salinity and temperature on the recruitment of Aurelia coerulea planulae. Mar. Biol. Res. 2018, 14, 454–461. [Google Scholar] [CrossRef]
- Kim, B. Distribution and Removal Effects of Polyps of the Moon Jellyfish, Aurelia coerulea (Cnidaria: Scyphozoa) in Korean coastal area. Ph.D. Thesis, Department of Fisheries Science Graduate School, Kunsan National University, Gunsan, Korea, 2018. [Google Scholar]
- Yoon, W.; Chae, J.; Koh, B.S.; Han, C. Polyp Removal of a Bloom Forming Jellyfish, Aurelia coerulea, in Korean Waters and Its Value Evaluation. Ocean Sci. J. 2018, 53, 499–507. [Google Scholar] [CrossRef]
- Zhang, F.; Sun, S.; Jin, X.; Li, C. Associations of large jellyfish distributions with temperature and salinity in the Yellow Sea and East China Sea. Hydrobiologia 2012, 690, 81–96. [Google Scholar] [CrossRef]
- Feng, S.; Zhang, F.; Sun, S.; Wang, S.; Li, C. Effects of duration at low temperature on asexual reproduction in polyps of the scyphozoan Nemopilema nomurai (Scyphozoa: Rhizostomeae). Hydrobiologia 2015, 754, 97–111. [Google Scholar] [CrossRef]
- Lee, H.E.; Han, C.H.; Kim, B.H.; Yoon, W.D. Effects of temperature and salinity on the asexual reproduction of Nemopilema nomurai (Scyphozoa: Rhizostomeae). Ocean Sci. J. 2017, 52, 573–579. [Google Scholar] [CrossRef]
- Xu, Y.; Ishizaka, J.; Yamaguchi, H.; Siswanto, E.; Wang, S. Relationships of interannual variability in SST and phytoplankton blooms with giantjellyfish (Nemopilema nomurai) outbreaks in the Yellow Sea and East China Sea. J. Oceanogr. 2013, 69, 511–526. [Google Scholar] [CrossRef]
- Hong, H.P.; Han, C.H.; Yoo, J.K. Population dynamics of jellyfish Aurelia aurita (s.l.) in Sihwa Lake. Ocean Polar Res. 2013, 35, 205–217. [Google Scholar] [CrossRef]
- Kim, B.-T.; Eom, K.-H.; Han, I.-S.; Park, H.-J. An Analysis of the Impact of Climatic Elements on the Jellyfish Blooms. J. Fish. Mar. Sci. Educ. 2015, 27, 1755–1763. [Google Scholar] [CrossRef][Green Version]
- Choi, J.G.; Jo, Y.H.; Moon, I.J.; Park, J.; Kim, D.W.; Lippmann, T.C. Physical forces determine the annual bloom intensity of the giant jellyfish Nemopilema nomurai off the coast of Korea. Reg. Stud. Mar. Sci. 2018, 24, 55–65. [Google Scholar] [CrossRef]
- Kang, Y.S.; Jung, S.; Zuenko, Y.; Choi, I.; Dolganova, N. Regional differences in the response of mesozooplankton to oceanographic regime shifts in the northeast Asian marginal seas. Prog. Oceanogr. 2012, 97–100, 120–134. [Google Scholar] [CrossRef]
- Jahan, R.; Choi, J.K. Climate Regime Shift and Phytoplankton Phenology in a Macrotidal Estuary: Long-Term Surveys in Gyeonggi Bay, Korea. Estuaries Coasts 2014, 37, 1169–1187. [Google Scholar] [CrossRef]
- Kim, D.H.; Seo, J.N.; Yoon, W.D.; Suh, Y.S. Estimating the economic damage caused by jellyfish to fisheries in Korea. Fish. Sci. 2012, 78, 1147–1152. [Google Scholar] [CrossRef]
- Kim, H.; Song, S.H.; Lee, S.; Kim, J.-B.; Yoo, J.-T.; Jang, D.-S. Dominant causes on the catch fluctuation of a set net fishery in the mid-south sea of Korea. J. Korean Soc. Fish. Technol. 2013, 49, 250–260. [Google Scholar] [CrossRef]
- Health Insurance Review and Assessment Service Korea, HIRAsabo Vol.141. Available online: http://www.hirasabo.or.kr (accessed on 11 December 2020).
- Ministry of Oceans and Fisheries. Jellyfish Envenomation to Bather in Korea Beaches; Marine Leisure Tourism Division: Sejong, Korea, 2019. [Google Scholar]
- Kim, J.H.; Han, S.B.; Durey, A. Fatal Pulmonary Edema in a Child after Jellyfish Stings in Korea. Wilderness Environ. Med. 2018, 29, 527–530. [Google Scholar] [CrossRef] [PubMed]
- Abukhalifeh, A.N.; Chandran, K. Understanding the Perception of Safety and Security of Tourists at Jeju Island, South Korea. Rev. Integr. Bus. Econ. Res. 2020, 9, 107–114. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
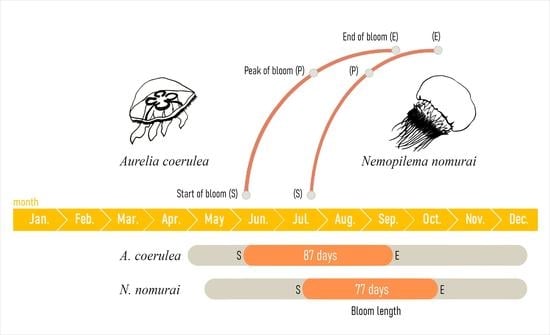
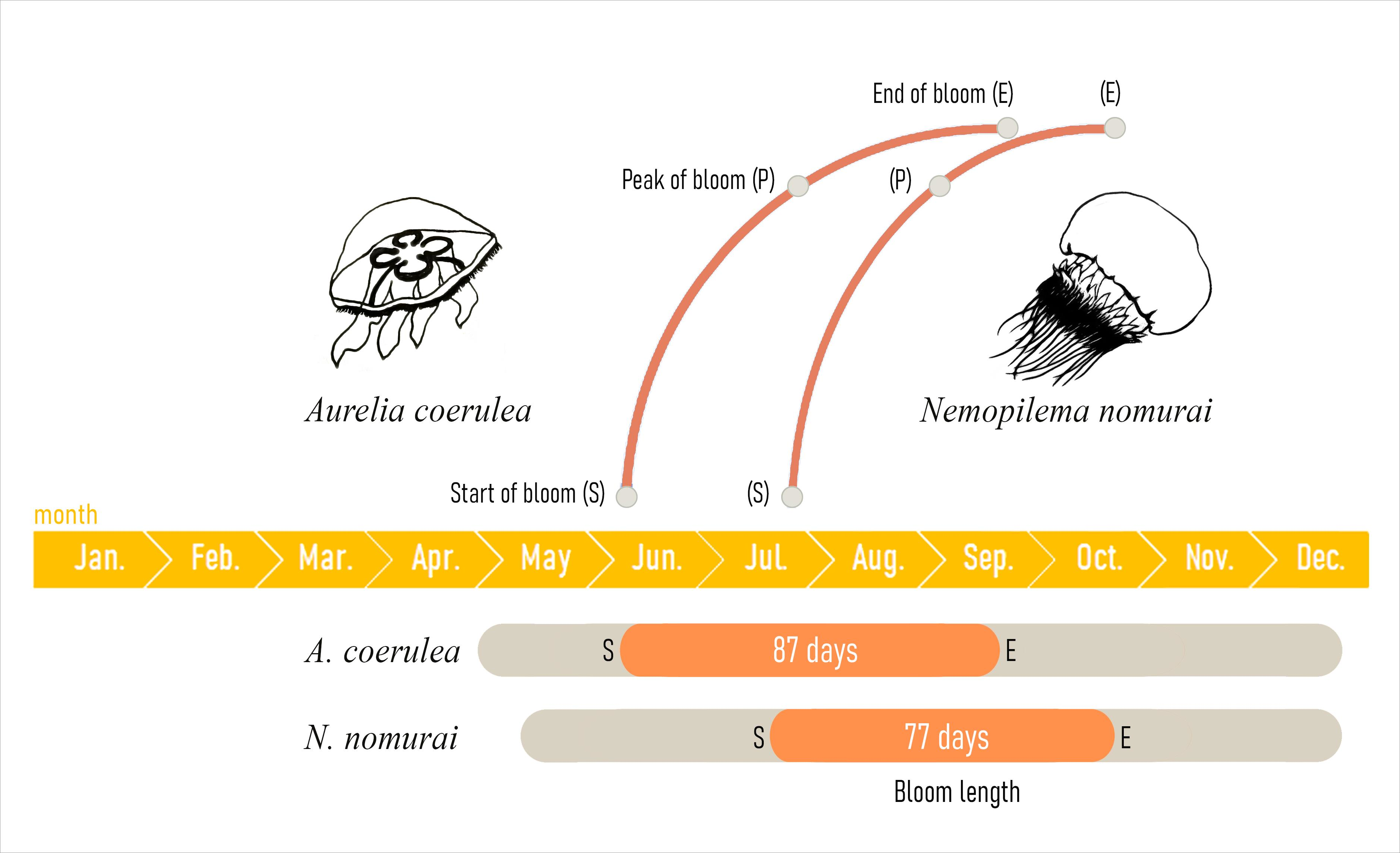
| Species | Start of Bloom | Peak of Bloom | End of Bloom | Bloom Duration (Days) | |||
|---|---|---|---|---|---|---|---|
| Day of Year | Driver | Day of Year | Driver | Day of Year | Driver | ||
| Aurelia coerulea | 163 | SST (0.65) | 198 | Prec. (−0.64) | 250 | SST (−0.60) | 77 |
| Nemopilema nomurai | 205 | -- | 247 | Reg. Temp. (−0.60) | 282 | -- | 87 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-H.; Hwang, J.-S.; Kim, K.-Y.; Molinero, J.C. Contrasting Effects of Regional and Local Climate on the Interannual Variability and Phenology of the Scyphozoan, Aurelia coerulea and Nemopilema nomurai in the Korean Peninsula. Diversity 2021, 13, 214. https://doi.org/10.3390/d13050214
Lee S-H, Hwang J-S, Kim K-Y, Molinero JC. Contrasting Effects of Regional and Local Climate on the Interannual Variability and Phenology of the Scyphozoan, Aurelia coerulea and Nemopilema nomurai in the Korean Peninsula. Diversity. 2021; 13(5):214. https://doi.org/10.3390/d13050214
Chicago/Turabian StyleLee, Sun-Hee, Jiang-Shiou Hwang, Kyoung-Yeon Kim, and Juan Carlos Molinero. 2021. "Contrasting Effects of Regional and Local Climate on the Interannual Variability and Phenology of the Scyphozoan, Aurelia coerulea and Nemopilema nomurai in the Korean Peninsula" Diversity 13, no. 5: 214. https://doi.org/10.3390/d13050214
APA StyleLee, S.-H., Hwang, J.-S., Kim, K.-Y., & Molinero, J. C. (2021). Contrasting Effects of Regional and Local Climate on the Interannual Variability and Phenology of the Scyphozoan, Aurelia coerulea and Nemopilema nomurai in the Korean Peninsula. Diversity, 13(5), 214. https://doi.org/10.3390/d13050214