Abstract
Most of our knowledge on the processes structuring parasite communities in amphibians originate from temperate-zone taxa, whereas Afrotropical communities have been neglected so far. We found evidence that ecological fitting of the hosts and, probably, differential immune response may influence the variation in parasite species richness, prevalence, and infestation intensity of East African frogs Hyperolius kivuensis and H. viridiflavus. The most closely related host species share the same macrohabitat (that implies the same pool of potential parasites), but differ in microhabitat preference, so that a comparative analyses of syntopic and allopatric populations is expedient to reveal ecological fitting. We detected 11 parasite species (one annelid, four nematodes, five trematodes, one cestode) and two endocommensal species (protozoans). The component parasite communities included 4–5 helminth species in H. kivuensis and 6–8 in the more aquatic H. viridiflavus, supporting the hypothesis that trematode diversity increases with the amount of time spent in water. Five parasite species (Orneoascaris chrysanthemoides, Clinostomum chabaudi, an undetermined echinostomatid) and two protozoans (Nyctotheroides sp., and Protoopalina sp.) are shared among the syntopic amphibian populations. This finding indicates a similar susceptibility of these amphibians to infestation from the local parasite pool. Yet, the low prevalence of single- and multi-species infestations in H. kivuensis indicates that parasite clearing by its immune response is probably more effective and prominent than in H. viridiflavus. Therefore, H. viridiflavus suffered from significantly reduced short-term survival due to the infection. Thus, we conclude that the processes structuring component parasite communities in amphibians do not differ generally between temperate-zone and Afrotropical host species, but they do in the magnitude of ecological fitting.
1. Introduction
Processes shaping host–parasite communities are complex in many ways. Parasites can infect several alternative host species at a certain stage of their life cycle, and a host can be infected by many different parasite species [1,2]. Parasite impact on host fitness may vary from neutral to detrimental [3]. For example, populations of two congeneric and sympatric host species, infested by cercariae of the same trematode parasite, may differ considerably in parasite-induced mortality [4]. The specific responses of a host to infestation or the resulting loss of fitness is directly dependent on the parasite species [5,6]. Moreover, host–parasite and parasite–parasite interactions are often characterized by seasonal dynamics and distinct context-dependence [1]. Consequently, ecological fitting is a major environmental determinant that structures the susceptibility of hosts to their potential parasites [7].
Amphibians are hosts with an ample diversity of life history patterns, reproductive modes, body sizes, foraging modes, and trophic relations, which permit the exploration of many aspects of processes influencing parasite community organization [3,8,9]. In aquatic and terrestrial food webs, amphibians may serve as intermediate, paratenic, or definite hosts to a variety of endoparasites. Yet, most of our knowledge on the ecological and evolutionary processes determining the parasite-supercommunity structure result from studies in temperate-zone amphibian-parasite systems [10,11]. As a rule, parasitic nematodes are more in species and abundance in terrestrial amphibian hosts, whereas parasitic trematodes dominate the parasite community of semi-aquatic and aquatic amphibians [8]. For example, the diversity of platyhelminth parasites in anuran hosts is increasing with the amount of time the host spends in aquatic habitats [7]. This relationship seems to be widely independent of the phylogenetic relationships among host species [6]. Partitioning variation in community structures shows that among-localities variation explains the largest portion (ecological fitting), followed by that among host species (phylogenetic divergence) and that among host individuals (random processes) [11]. When focusing on the ecological fitting of component parasite communities, it may be stated that the currently known community forming processes in temperate-zone amphibians seem to favor a dominance and higher diversity of parasitic nematode species in more terrestrial hosts, a dominance and higher diversity of parasitic trematodes in more aquatic species, and a habitat-independent, diet-mediated presence of parasitic cestodes and acanthocephalans [6,8,11].
In this study, we tested the hypothesis that the processes structuring the parasite communities in temperate-zone amphibians apply to those in Afrotropical amphibians as well. As a specific model system to study the effects of ecological fitting we used the East African Reed Frogs Hyperolius kivuensis and H. viridiflavus, which are closely related phylogenetically (resulting in a similar parasite susceptibility due to common ancestry), co-occur with high abundance in wetlands at 1287–2143 m above sea level sharing the same larval habitats (common sympatric macrohabitat preference), but differ in the adults’ microhabitat preference and several other features, i.e., the amount of time spent in water [12,13,14,15,16,17]. If host–parasite communities from tropical Africa follow the same assembly rules as in the temperate zone, we would predict that (1) the component parasite community of the more aquatic host species is trematode-dominated and more diverse than that of the more terrestrial host; (2) the prevalence of the infested hosts may increase with the species richness of the component parasite community; (3) syntopic host populations share more parasite species than allopatric ones; and (4) prevalence of shared parasite species, infestation intensity and impact on host life-history may be similar in the two syntopic hosts.
To test these predictions, we first assessed the taxonomic identity (α-diversity) and seasonal dynamics of endoparasite species infesting these frogs. Current knowledge is limited to six identified species [18,19,20]. Consequently, we collected more than 100 specimens per host species from syntopic and allopatric populations at the beginning, the middle and the end of the rainy periods that determine the reproductive behavior and general activity of frogs. We quantified the frequency of infested host specimens and identified the parasites taxonomically by combining morphological and molecular methods [18,19,20]. The resulting data set was conclusive to evaluate the predictions with respect to the ecological fitting of component parasite communities and their impact on the growth and survival of their hosts.
2. Materials and Methods
2.1. Sampling
In total, 89 adult Kivu reed frogs (Hyperolius kivuensis, HK) and 100 common reed frogs (H. viridiflavus, HV) were sampled from syntopic populations inhabiting the cultivated wetlands (“marais”, 2.60° S 29.76° E, 1645m a.s.l.) near Huye (Butare), Rwanda [12]. Sampling dates were 2 October 2015 (n = 18 HK; n = 26 HV), 1 October 2016 (n = 4 HV), 14 October 2018 (n = 27 HV), 10 January 2017 (n = 35 HK; n = 25 HV) and 19 March 2017 (n = 26 HK; n = 18 HV), thus including the complete rainy period at this locality. Complementary samples of an allopatric H. kivuensis population were collected in a wetland near Musanze (Ruhengeri, 1.51° S 29.65° E, 1807m a.s.l.; n = 26) on 6 October 2015. Additional samples of allopatric H. viridiflavus were collected in a wetland near Muhanga (Gitarama, 2.12° S, 29.82° E, 1650 m a.s.l.; n = 29) on 16 October 2018 [16]. The Reed Frogs collected in October 2015 were transported to the laboratory in Koblenz, Germany and kept alive in groups of five conspecific individuals within terraria with food ad libitum until 30 November 2015. They were euthanized by immersion into a 2% solution of tricaine methane-sulfonate (MS 222), except for 10 H. kivuensis individuals kept alive until 6 May 2017. All other host specimens collected after 2015 were euthanized immediately after collection. Each individual was sexed and the snout-vent length (SVL, distance between snout tip and cloaca) was measured to the nearest 0.1 mm using a caliper. The body cavity, digestive tract, lungs, and the urogenital system were subsequently examined macroscopically and (light-) microscopically for the presence of protozoan endocommensals and metazoan endoparasites. Captured prey present in the digestive tract were counted and roughly assigned to taxonomic groups. No attempt was made to identify prey items to generic or species level due to the lack of identification keys for this geographic region. Carcasses of frogs were fixed in 10% buffered formalin, transferred to 70% ethanol for long-term storage and deposited in the collection of the Zoologisches Forschungsmuseum Alexander Koenig, Bonn, Germany (ZFMK). Collection numbers are ZFMK 102849–102962 (H. kivuensis) and ZFMK 102963–103098 (H. viridiflavus).
Representative voucher specimens of metazoan parasites and protozoan commensals from the host frogs were transferred to Eppendorf tubes with 1.5 mL ethanol for molecular identification. All remaining parasites were fixed in 10% buffered formalin for subsequent morphological examination. Metazoan parasites were assessed quantitatively per host individual, whereas the presence of protozoan endocommensals was recorded qualitatively. All applicable international, national and/or institutional guidelines for the care and use of animals were followed, and all procedures performed were in accordance with the ethical standards of the University of Koblenz-Landau (approval # Si 2015/01, ethic committee of FB3).
2.2. Morphological Identification of Parasites and Endocommensals
The macroscopically detected metazoan parasites were first observed live using (1) a digital Keyence microscope VHX-6000 and digitally documented using Keyence VHX-H1M1 software, or (2) an Olympus BX 50 microscope equipped with a high-resolution camera Olympus DP20 for documentation with the Cell Imaging software. Formalin-fixed helminths were placed in a drop of lactophenol on a microscope slide, cover-slipped and examined using the BX 50 microscope.
The trematodes and nematodes were identified to genus level based on the available identification keys. Whenever possible they were determined to the species level by subsequent comparison with the original morphological descriptions [21,22,23,24,25]. Parasite terminology is used in accordance with [8]. Parasitic annelids have been reviewed in [26,27,28]. Cestode taxonomy is still far from being well-understood [29,30,31]. Therefore, we consulted T. Scholz (České Budějovice, Czech Republic) for an expert opinion on the cestodes collected. Studies on intestinal ciliates and opalinids of amphibians are limited to few geographical areas and host species. Nevertheless, identification to genus level was possible [32,33,34]. Vouchers specimens of each taxon were deposited at the Zoologisches Museum Hamburg, LIB-Leibniz Institut für die Analyse des Biodiversitätswandels, Germany. Collection numbers are ZMH N14023–14027, ZMH Ol15490–15491, ZMH Pr318–321, and ZMH Tr13202–13207.
Species richness of the parasite/endocommensal community was assessed using the rarefaction technique combined with a jackknife estimate of potentially overlooked species. A presence/absence data matrix (parasite species in each host specimen) was constructed for each host species and sampling period (October, January, March). Then, matrices were submitted to a Cole-rarefaction analysis and a jackknife richness estimation using the statistical package EstimateS 9.1.0 [35]. The α-diversity of the local helminth parasite community was quantified using the Shannon–Wiener Index and the corresponding evenness [35]. The β-diversity among helminth parasite communities in allopatric, conspecific and in syntopic, heterospecific host populations was estimated as the Sørensen and the Jaccard similarity-coefficients [36].
2.3. Molecular Characterisation of Parasites
The ethanol-fixed samples were incubated in 1.5 mL reaction tubes, with open cap, at 40 °C in a Thermomixer (Eppendorf, Germany) until the ethanol was completely evaporated. The sample was mixed with 180 µL ATL buffer and 20 µL proteinase K, incubated at 56 °C until the specimen was completely lysed, and then further processed following the protocol for DNA purification from tissues of the QIAamp DNA mini kit (Qiagen, Germany). The extracted DNA was used to amplify either 18S, 18S-ITS1–5.8S or 28S fragments of nuclear ribosomal locus or partial mitochondrial cytochrome oxidase subunit 1 (COI1) locus (Table 1). Polymerase chain reaction (PCR) was conducted using Taq PCR core kit as recommended by the manufacturer (Qiagen). The following program was used: 40 cycles of 1 min at 94 °C, 1 min at 48 °C and 1 min at 72 °C. All PCR cycles were initiated with a denaturation step for 3 min at 94 °C and terminated with an extension step of 72 °C for 7 min. Amplicons were separated on a 1% agarose gel, stained with ethidium bromide and visualized on a UV transilluminator. Prior to sequencing, PCR products were purified using QIAquick Purification Kit (Qiagen).
For bidirectional sequencing, the same primers as for the PCR were applied (Table 1). Assembly of DNA sequence files was conducted with DNA Baser (Heracle BioSoft, Romania) and primer sequences were clipped. Sequences were deposited in GenBank (Table 2). Closest matches of sequences were identified by a BLAST search against GenBank entries [37].

Table 2.
Species list of metazoan endoparasites and protozoan endocommensals infesting reed frogs in Rwanda. Prevalence (presented as the absolute number of parasitized frog hosts and as ratio by the total number examined), total number of individuals per parasite species in H. kivuensis (nHK) and H. viridiflavus (nHV), and accession number of sequences uploaded to the GenBank data base.
Table 1.
Primers used for amplification and sequencing.
Table 1.
Primers used for amplification and sequencing.
| Gene | Primer | Sequence | Reference |
|---|---|---|---|
| 18S-ITS1–5.8S | S1 | 5′-ATTCCGATAACGAACGAGACT-3′ | [38] |
| IR8 | 5′-GCTAGCTGCGTTCTTCATCGA-3′ | [39] | |
| 18S | F18 | 5′-ACCTGGTTGATCCTGCCAGTAG-3′ | [40] |
| 18RG | 5′-CTCTCTTAACCATTACTTCGG-3′ | [40] | |
| 18F3 | 5′-GGACGGCATGTTTACTTTGA-3′ | [40] | |
| IR5 | 5′-TACGGAAACCTTGTTACGAC-3′ | [40] | |
| 28S | LSU5′ | 5′-TAGGTCGACCCGCTGAAYTTAAGCA-3′ | [41] |
| IR14 | 5′-CATGTTAAACTCCTTGGTCCG-3′ | [40] | |
| IF15 | 5′-GTCTGTGGCGTAGTGGTAGAC-3′ | [40] | |
| LSU3′ | 5′-TAGAAGCTTCCTGAGGGAAACTTCGG-3′ | [41] | |
| COI | MplatCOX1dF | 5′-TGTAAAACGACGGCCAGTTTWCITTRGATCATAAG-3′ | [42] |
| MplatCOX1dR | 5′-CAGGAAACAGCTATGACTGAAAYAAYAIIGGATCICCACC-3′ | [42] |
2.4. Host Life-History Traits: Age and Size
The carcasses of 174 reed frogs (101 HK, 73 HV) were examined for potential effects of parasite infestation on size and age). Age was estimated following standard laboratory protocols of skeletochronology [43]. In the diaphysis cross sections, we counted the strongly stained lines of arrested growth (LAGs) in the periosteal bone as an estimator of age (1 LAG = 1 year). Life-history traits recorded for each frog were: (1) overall growth as SVL, (2) age as number of LAGs.
3. Results
3.1. Local Host Biology: H. kivuensis and H. viridiflavus
Mediated by the wide tolerance to human disturbance, local macrohabitat preference of the two species was very similar and included flooded natural and artificial grasslands, papyrus swamps, vegetation-rich shores of ditches, streams and fishponds in agricultural areas, and the reed belts of natural lakes. Macrohabitat segregation was limited to an avoidance of areas with closed and tall reed, papyrus, or sedge vegetation by H. viridiflavus because this species preferred areas with smaller herbs and open water surface (Figure 1A,B). Microhabitat choice of reproductive adults did not differ between the species in quality, but significantly in the frequency of use. During night, males of H. kivuensis gave advertisement calls usually from herbs, reed, or papyrus at a height of 30–100 cm, more rarely close to the water surface sitting on sedges (Figure 1D,F). Calling males of H. viridiflavus were found at similar sites, but showed a clear preference of call positions near or at the ground or on the water surface at floating leaves of water lilies (Figure 1C,E). Changing call position included frequently swimming in H. viridiflavus, whereas H. kivuensis usually leapt from branch to branch. Overall, H. kivuensis showed more terrestrial/arboreal habits than H. viridiflavus, i.e., the amount of time spent in water was considerably less in H. kivuensis.

Figure 1.
Macro- and microhabitats of the host frogs H. kivuensis and H. viridiflavus. (A) Flooded rice field with irrigation channels in Huye (HK and HV in syntopy); (B) dense papyrus swamp in Mugesera (only HK); (C,D) calling sites 40–60 cm above ground; (E,F) calling sites close to the water surface on floating leaves. Hyperolius kivuensis is shown on (D,F), H. viridiflavus on (C,E).
The reproductive period of the two species was identical to the rainy period, i.e., from October to March of the following year. Hyperolius kivuensis deposited egg masses at moist shore ground near the water surface, whereas H. viridiflavus placed egg clutches directly into the water. Following embryogenesis in the egg gelatin, tadpoles of H. kivuensis were washed to the nearby water body during the frequent rainfalls. Tadpoles of the two species developed usually in the same waterbodies, reaching up to 54 mm total length at Gosner stages 37–39. Duration of the aquatic development varied between 2–4 months depending on water and food availability and did not differ between species. At the end of the rainy season in March, reproductive behavior of H. kivuensis ceased entirely and males lost the bright yellow coloration of the vocal sac. In contrast, H. viridiflavus continued to advertise at the beginning of the dry period. It is still unknown whether mating and egg deposition continue to take place in habitats providing open water during the dry period.
Diet composition of the two species has not yet been quantified, but we observed frogs feeding opportunistically on moving prey small enough to be swallowed. Food was not a limiting factor for adult frogs because the digestive system of all individuals examined contained 1–6 prey items. Diet of adults included mainly orthopterans and dipterans, rarely odonates, coleopterans and annelids, i.e., the terrestrial life stages of the prey species. We did not find evidence for aquatic prey such as tadpoles, aquatic insects (including larvae) or mollusks.
The two reed frogs were short-lived, with a longevity of 2–3 years in the natural habitat. Average age and corresponding standard deviations of parasite-free H. kivuensis (females and males pooled) did not differ significantly from those of infested individuals (1.1 ± 0.2 LAGs vs. 1.1 ± 0.2 LAGs, t-test, t = −0.34, p = 0.7331). The same counted for H. viridiflavus (1.0 ± 0.2 LAGs vs. 1.1 ± 0.2 LAGs, t-test, t = −0.52, p = 0.6073). Average and maximum male snout-vent length was similar in the two species (HK SVL 28.4 mm, 35.5 mm; HV SVL 27.3 mm, 34.3 mm). Log10-transformed SVL of female H. kivuensis (n = 17) was significantly greater than that of males (n = 98; 2-factor ANCOVA, F1,115 = 21.79, p < 0.0001), if adjusted for age (2-factor ANCOVA, F1,115 = 5.89, p = 0.0167). The presence of one or more endoparasite species (metazoans and protozoans considered) also affected SVL, with infested individuals being significantly smaller than parasite-free specimens (2-factor ANCOVA, F1,115 = 5.33, p = 0.0226). The interaction between sex and state of infestation was highly significant (2-factor ANCOVA, F1,115 = 7.00, p = 0.0092). In H. viridiflavus, log10-transformed SVL did not differ significantly between females and males (2-factor ANCOVA, F1,72 = 0.52, p = 0.4732) or among age classes (2-factor ANCOVA, F1,72 = 0.92, p = 0.3397). In contrast, parasite-free individuals were significantly smaller than infested ones (2-factor ANCOVA, F1,79 = 6.24, p = 0.0149). The low number of available female H. viridiflavus (n = 4) did not allow an estimation of the interaction between sex and state of infestation.
3.2. Parasite-Community Structure: Species Richness, Abundance and Organ Systems Infested
The taxonomic diversity of endoparasites in H. kivuensis and H. viridiflavus included 11 metazoan species, with 1512 specimens and two unicellular eukaryote endocommensal species with unquantified abundance (Table 2; Figure 2). Helminth species richness of the syntopic populations at Huye was five in H. kivuensis (α-diversity: Shannon-Index 1.42, Evenness 0.89), and eight in H. viridiflavus (α-diversity: Shannon-Index 1.04, Evenness 0.50). In the allopatric population of H. kivuensis at Musanze, four helminth species were detected (α-diversity: Shannon-Index 1.25, Evenness 0.91). The nematode Orneoascaris schoutedeni occurred instead of O. chrysanthemoides. In the allopatric population of H. viridiflavus at Muhanga, four helminth species were detected (α-diversity: Shannon-Index 0.17, Evenness 0.13). The high abundance of the unidentified echinostomatid metacercariae in H. viridiflavus, especially in Muhanga, decreased the evenness index considerably compared with those of H. kivuensis. The syntopic populations of H. kivuensis and H. viridiflavus at Huye shared three helminth species (β-diversity: Sørensen–coefficient 0.461, Jaccard similarity coefficient 0.30). The local parasite communities of the two allopatric H. kivuensis host populations shared three species (β-diversity: Sørensen–coefficient 0.667, Jaccard similarity coefficient 0.50), whereas those of H. viridiflavus four species (β-diversity: Sørensen–coefficient 0.667, Jaccard similarity coefficient 0.50). The endocommensal protozoan species were present in both host species at all localities surveyed.
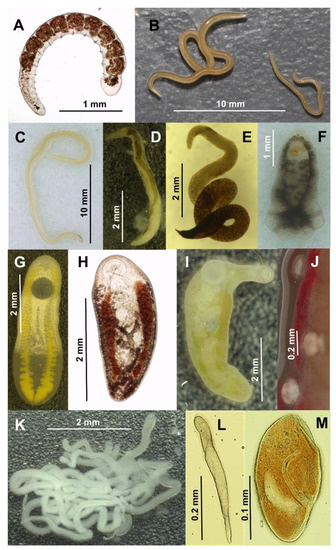
Figure 2.
Endoparasite and endocommensal species detected in H. kivuensis and H. viridiflavus. (A) Dero rwandae (HK); (B) female (left) and male (right) Orneoascaris chrysanthemoides (HK, HV); (C) male O. schoutedeni (HK, exclusively in Musanze); (D) female Aplectana chamaeleonis (HK); (E) Rhabdias collaris (HV); (F) Polystoma sp. (HV); (G) Clinostomum chabaudi (HK, HV); (H) Glypthelmins africana (HV); (I) Gorgoderina africana (HV); (J) three undetermined echinostomatid metacercariae (HK, HV); (K) five Nematotaenia sp. individuals (HV); (L) Protoopalina sp. (HK, HV); (M) Nyctotherioides sp. (HK, HV). Bars represent the scale of each photo.
The rarefaction analyses including all sampled localities showed a saturation plateau (six parasite and two endocommensal species) in H. kivuensis after adding 60 individuals, whereas in H. viridiflavus a saturation plateau was not reached after adding all 129 individuals (Figure 3). The jackknife estimate of the actual total number of parasite and endocommensal species comprising the community was 8.9 ± 0.2 (mean and corresponding 95% confidence interval) in H. kivuensis, and 11.0 ± 0.2 in H. viridiflavus, suggesting the presence of one overlooked species.
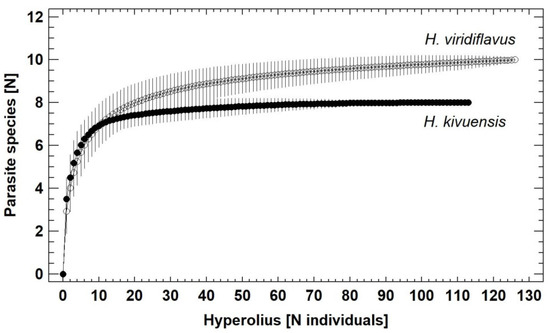
Figure 3.
Rarefaction (species-saturation) diagram of parasite + endocommensal species richness in H. kivuensis and H. viridiflavus. Vertical lines give the standard deviation for each estimate.
Details on the morphological and molecular identification of six species (Dero rwandae Sinsch, Dehling, Scheid & Balczun, 2019, Orneoascaris chrysanthemoides Skrjabin, 1916, Orneoascaris schoutedeni (Baylis, 1940), Aplectana chamaeleonis (Baylis, 1929), Rhabdias collaris Baker, 1987, Clinostomum chabaudi Vercammen-Grandjean, 1960) have been published by our research group elsewhere [17,18,19]. Two digenetic trematodes were identified morphologically as adult Glypthelmins africana Dollfus, 1950 and Gorgoderina africana Meskal, 1970, respectively [44,45]. None of the Gorgoderina specimens had completed development entirely, all had uteri filled with eggs, but testes were absent. Here, we provide the first DNA sequences for the barcoding of these rarely found African trematode species. Another four taxa were morphologically identified to genus level and in part characterized by DNA sequences: the monogenean trematode Polystoma sp., the cyclophyllid cestode Nematotaenia sp., and the unicellular eukaryotes Nyctotheroides sp. and Protoopalina sp. All Polystoma and Nematotaenia specimens were adults carrying eggs. BLAST search based on the DNA sequences obtained were inconclusive for species determination. The most abundant trematode metacercaria belonged to an echinostomatid, that was encysted on the kidney surface. Specific and generic identity remained unknown, but the most similar sequence (accession number AY245709) identified by BLAST search pertained to Petasiger phalacrocoracis (Echinostomatidae) with a deep sequence divergence of 9–10% (18S and ITS1).
The most affected host organ systems were the small and large intestines, harboring a total of seven species, among them three nematodes (O. chrysanthemoides, O. schoutedeni, A. chamaeleonis), the trematode Glypthelmins africana, the cestode Nematotaenia sp., and the two protozoans (Nyctotheroides sp., Protoopalina sp.). The urogenital system was infested by four species, the annelid D. rwandae (bladder and Wolffian ducts) and three trematodes (Gorgoderina africana within the Wolffian ducts, undetermined echinostomatid metacercaria on the surface epithelium of the kidneys, Polystoma sp. within the bladder). In contrast, the respiratory system was parasite-free in all but one host individual, in which the lung contained a single specimen of the nematode R. collaris. Lymphatic sacs and body cavity contained frequently metacercariae of the digenetic trematode C. chabaudi.
3.3. Host-Parasite Interactions
3.3.1. Susceptibility of Hosts to Parasite Infestation
The prevalence of individuals infested with metazoan parasites differed among the host species (Table 3). Of H. kivuensis, 34.8% were infested, while 62.0% of H. viridiflavus showed endoparasitic infections. Parasite load was generally lower in infested H. kivuensis in terms of multi-species infestations and with respect to the number of parasites per host. Multi-species infestations were rare in H. kivuensis (n = 3, 2.6%), while six times more common in H. viridiflavus (n = 20, 15.6%). Based on the prevalence of each parasite species in the host population at Huye, the expected number of two-species infestations was three times greater than that the actually detected number in H. kivuensis (6.0% vs. 1.7%). In contrast, the expected frequency of two-species infestations was slightly lower than the actual one in H. viridiflavus (10.7% vs. 14.0%). Specifically, co-infestations of Clinostomum and echinostomatid metacercaria were more frequent (7.8%) than expected (6.5%). Among the shared parasite species, prevalence numbers of C. chabaudi and O. chrysanthemoides were about the same in the two host species, whereas seven times more H. viridiflavus than H. kivuensis were infested with echinostomatid metacercariae.
Table 3.
Helminth parasite load, parasite association and overall prevalence in host species.
3.3.2. Seasonal Variation of Infestation
Rate of host infestation showed distinct seasonal patterns in the helminth parasites (Table 4). Polystoma sp. was detected exclusively at the beginning of the rainy period, whereas D. rwandae only at its end. The nematode O. chrysanthemoides, the trematode Gorgoderina africana, and the cestode Nematotaenia were confined to the first half of the rainy period. In contrast, the nematode A. chamaeleonis and the trematodes C. chabaudi, Glypthelmins africana, and the unidentified echinostomatid were present in the frog hosts at any time of the rainy season, with random fluctuations in prevalence.
Table 4.
Seasonal variation of infestation by metazoan parasites in the two host species. October marks the beginning of the rainy season, January a period of less rainfall, March the end of the rainy period. Prevalence is expressed as percent of collected individuals infested with the specific parasite. Intensity of infestation is expressed as the number of parasite individuals in a single frog host. Orneoascaris schoutedeni and Rhabdias collaris are not listed because they were found only in a single host frog.
3.3.3. Impact on Host Survival
Hyperolius viridiflavus specimens captured at the beginning of the rainy period and held in captivity for 60 d showed a significantly lowered survival of infested specimens (43.2 ± 3.7 d vs. 55.3 ± 4.1 d), if the intensity of infestation was considered as a co-variable (data log10-normalized, ANCOVA, F1,25 = 4.35, p = 0.0484). Still, the intensity of infestation did not co-vary significantly with survival (ANCOVA, F1,25 = 3.45, p = 0.0762). Analysis of data on specimens infested with a single parasite species did not provide statistically significant differences in survival due to the low case numbers, but there was a tendency that the survival of frogs infested with Polystoma and/or Nematotaenia specimens was lower than that of parasite-free frogs (Table 5).
Table 5.
Effect of host infestation on survival of frogs collected in the wild and kept 60 d in captivity.
In contrast, presence of parasites and parasite load did not affect the short-term survival of captive H. kivuensis (55.4 ± 3.2 d vs. 54.4 ± 3.1 d; ANCOVA, F1,31 = 0.05, p = 0.8233). The presence of A. chamaeleonis and/or echinostomatid metacercariae had no notable effects on short- and long-term survival and growth of six H. kivuensis kept for two years in a terrarium. There was no tendency for an impact of any parasite on short-term survival (Table 5).
4. Discussion
We provide evidence that seasonal ecological fitting and, probably, the immune response to infestation are important processes shaping the component parasite communities in two small-sized East African frog species that inhabit the same locality (ecological context) and are closely related (common ancestry). Previous studies on tropical West African anuran-parasite communities focused exclusively on the supercommunity composition of distantly related anuran hosts and failed to recognize the underlying structuring processes due to low numbers of host individuals surveyed [46,47,48,49,50,51,52]. The α-diversity of the component parasite communities found in the East African Hyperolius spp. (4–8 helminth species) is within the range of West African frog hosts like the aquatic Xenopus muelleri and X. tropicalis (5–10 species), the semi-aquatic Ptychadena spp. and Hoplobatrachus occipitalis (7–14 species), and the terrestrial Arthroleptis poecilonotus (13 species) [46,48,49,50,51,52]. These study results and the jackknife estimation of total species richness presented in our study emphasize that our sampling effort has yielded a data set conclusive for the analysis of the interactions in the Hyperolius-parasite model system. Subsequently, we evaluate the validity of predictions on the assembly rules derived from parasite communities in temperate-zone anurans for Afrotropical Hyperolius spp.
4.1. Prediction 1: The Component Parasite Community of the More Aquatic Host Species Is Trematode-Dominated and More Diverse Than That of the More Terrestrial Host
Hyperolius viridiflavus spending more time in aquatic microhabitats than syntopic H. kivuensis harbored three more trematode species as predicted. This finding agrees with the trematode-species richness of the community infesting the fully aquatic South African Xenopus laevis [53]. In contrast, dominance of trematodes in anuran hosts strongly attached to water does not fit to West African Xenopus, Hoplobatrachus, and Ptychadena spp [46,48,50,52]. Thus, the assembly rule derived from parasite communities in temperate-zone anurans [6] is not generally supported by Afrotropical anurans.
The monogenean Polystoma sp. and the digeneans Glypthelmins africana and Gorgoderina africana were exclusive to H. viridiflavus, representing the definite host. All shared an infestation dynamic confined to the first half of the rainy season. Adult bladder-infesting Polystoma originate from free-swimming larvae (oncomiracidia), that have infested the tadpole of the anuran host during its aquatic development [54,55]. Sexual maturation of Polystoma seems to be synchronized with the onset of reproductive activity of the host frog, like observations in the toad Sclerophrys regularis [56]. Anurans get infected either by skin penetration of cercariae (Glypthelmins) or by feeding on the second intermediate host (Gorgoderina) [57,58]. Surprisingly, all Gorgoderina africana found in the kidneys of H. viridiflavus had uteri full of eggs, but no testes, as observed before in G. attenuata [59]. This syndrome was attributed to a poor nutritional state of the host. Given that we never found Gorgoderina individuals in the bladder (like G. vitelloba in H. arborea), it appears possible that H. viridiflavus is an accidental, but not genuine definite host (i.e., Amietia angolensis [45]), because the same parasite mass may trigger a stronger antigenic response in smaller than in larger hosts, and the kidney may act as a barrier to the completion of the development in the bladder [60]. In conclusion, the presence of the three trematodes exclusive to H. viridiflavus is certainly related to the longer time spent in aquatic habitat. The significant host specificity of Polystoma may prevent H. kivuensis from becoming infested, though they share the larval habitat with H. viridiflavus. The reasons for the absence of Glypthelmins and of Gorgoderina in H. kivuensis remain open, but the latter does not seem to be able to complete development in small-sized frogs.
4.2. Prediction 2: Prevalence of Infested Hosts Increases with the Species Richness of the Component Parasite Community
In the syntopic Hyperolius populations, 62.0% of the H. viridiflavus specimens were infested, whereas only 34.8% of H. kivuensis showed an endoparasitic infection with a less diverse component parasite community. This agrees with the prediction above (Section 4.1), but alternatively may be the result of the greater amount of time that H. viridiflavus spent in water. The prevalence in H. viridiflavus is like that of West African semi-aquatic frogs such as H. occipitalis and Ptychadena spp (67.7–100%) [46,47,52]. Yet, the ecological fitting by microhabitat segregation is probably not the only process controlling prevalence in the Hyperolius-parasite system. As the frequency of multi-species infestation in H. viridiflavus was higher than expected from the one-species infestations, we speculate that some parasite species may facilitate co-infection with other parasites [61,62]. In contrast, the low prevalence of infested H. kivuensis despite a susceptibility to at least six helminth species may be mediated by a stronger host immune response. Since priority effects, i.e., the arrival of the first parasite in the host, have been shown to play an important role in structuring parasite communities within hosts [62], the triggering of the host immune response by the first parasite may cause the very low number of multi-species infestations. The striking difference in prevalence and infestation intensity of the unidentified echinostomatid metacercariae between the two host species supports the assumed successful immune response of H. kivuensis (see Section 4.4).
4.3. Prediction 3: Syntopic Host Populations Share More Parasite Species Than Allopatric Ones
The major source of variability in parasite community of temperate-zone anurans structure appears to be among-localities variation (ecological fitting) [11]. The β-diversity among syntopic and allopatric host populations in the Hyperolius model system does not suggest a corresponding partition of variation in the structure of Afrotropical parasite communities. Allopatric, conspecific host populations harbored more similar parasite communities than syntopic, heterospecific host populations, indicating that the genetic determination of susceptibility outweighed the structuring role of ecological fitting. Yet, composition of local parasite communities in syntopic and allopatric populations of the semi-aquatic frogs Ptychadena pumilo and P. “mascareniensis” from Nigeria seems to follow different rules than those in the Hyperolius-parasite system [48,52]. In syntopy, the parasite communities of the two species are considerably more similar (at Okada: Sørensen–coefficient 0.50; at Rumuji-Emohua: Sørensen–coefficient 0.63) than those of conspecific host frog populations in allopatry (P. “mascareniensis”: Sørensen–coefficient 0.13; P. pumilio: Sørensen–coefficient 0.33). The importance of ecological fitting in the structuring of parasite communities seems to differ among West and East African localities and possibly also depend on frog-host features.
4.4. Prediction 4: Prevalence of Shared Parasite Species, Infestation Intensity and Impact on Host Life-History Are Similar in the Two Host Species
Seasonal variation of infestation and prevalence of infested hosts was, in fact, similar in the two Hyperolius hosts with respect to the nematode O. chrysanthemoides and the trematode C. chabaudi, indicating that susceptibility to these parasites is an ancestral (evolutionary) feature in the host species. In contrast, prevalence of hosts carrying the undetermined echinostomatid was strongly biased towards H. viridiflavus reaching 96.6% in the Muhanga population and showing significantly greater intensities of infestation. Echinostomatid metacercariae are known to cause reduced survival, decreased growth, and inflammations in several temperate-zone anurans [3,63,64]. Little is known about the role of host immunity in regulating amphibian–trematode interactions, but there is experimental evidence that echinostomid metacercariae are sensitive to immune responses of the host and may be eliminated completely by the host [65]. If we assume that infestation probability of tadpoles does not differ between the two syntopic host species, the apparent differences found in the kidneys of adults may result from an effective immune response of H. kivuensis.
The smaller size of trematode-infested H. kivuensis compared to parasite-free individuals, indicates an important energy expenditure as response to infestation to prevent negative effects on the short -and long-term survival of parasitized H. kivuensis adults. In contrast, short-term survival of infested H. viridiflavus was lower than in parasite-free individuals, while no effects on size were observed. The reversed response pattern indicates a fundamentally different physiological reaction to infestation. Taken together with the unusual low prevalence of parasitized H. kivuensis adults and the non-randomly low number of multi-species infestation, we assume that the immune response to parasites is more sufficient in H. kivuensis than in H. viridiflavus.
5. Conclusions
The processes structuring component parasite communities in amphibians do not differ generally between temperate-zone and Afrotropical host species, but they do in detail. The major source of variation in the component community composition seems to be the ecological fitting of the parasite pool present in each host habitat in the two ecoregions [10]. Microhabitat segregation within the same macrohabitat of closely related host frog species may lead to a predictable difference in trematode diversity. Unlike the findings in temperate-zone communities [3], more terrestrial habits of the host did not increase nematode species richness or intensity of infestation. Moreover, the features of syntopic and allopatric Hyperolius populations emphasize that genetically predetermined host susceptibility to certain parasites and probably different degrees of immune response to parasites may substantially influence component community composition and the intensity of infestation. The understudied Afrotropical anuran-parasite communities deserve our increased attention in future to complement the temperate-zone biased bulk of information for better founded ecological insights.
Author Contributions
Conceptualization, U.S., J.M.D., P.S. and C.B.; methodology, C.B. and U.S.; software, U.S.; validation, U.S.; formal analysis, U.S.; investigation, U.S., J.M.D., P.S. and C.B.; resources, U.S. and C.B.; data curation, U.S. and C.B.; writing—original draft preparation, U.S., J.M.D., P.S. and C.B.; writing—review and editing, U.S., J.M.D., P.S. and C.B.; visualization, U.S.; supervision, U.S.; and project administration, U.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
All applicable international, national and/or institutional guidelines for the care and use of animals were followed, and all procedures performed were in accordance with the ethical standards of the University of Koblenz-Landau (approval # Si 2015/01, ethic committee of FB3).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article, host and parasite specimens analyzed in this study are deposited in the natural history Museums of Bonn and Hamburg, Germany (collections numbers within the article), and are available on request to the collection managers.
Acknowledgments
We are indebted to Bonny Dumbo for assistance during the collection of Hyperolius in the field. Collecting and export permits (Nos. 2/ORTPN/V.U/09, 08/RDB-T&C/V.U/12, 18/RDB-T&C/V.U/12 and 10/RDB-T&C/V.U/15) of Hyperolius for parasitological screening were issued by the Rwandan Development Board.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Johnson, P.T.J.; Buller, I.D. Parasite competition hidden by correlated coinfection: Using surveys and experiments to understand parasite interactions. Ecology 2011, 92, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Poulin, R.; Forbes, M.R. Meta-analysis and research on host-parasite interactions: Past and future. Evol. Ecol. 2012, 26, 1169–1185. [Google Scholar] [CrossRef]
- Koprivnikar, J.; Marcogliese, D.J.; Rohr, J.R.; Orlofske, S.A.; Raffel, T.R.; Johnson, P.T.J. Macroparasite Infections of Amphibians: What Can They Tell Us? Ecohealth 2012, 9, 342–360. [Google Scholar] [CrossRef] [PubMed]
- Thomas, F.; Renaud, F.; Rousset, F.; Cezilly, F.; Meeuûs, T.D. Differential mortality of two closely related host species induced by one parasite. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1995, 260, 349–352. [Google Scholar] [CrossRef]
- Rauque, C.; Paterson, R.; Poulin, R.; Tompkins, D. Do different parasite species interact in their effects on host fitness? A case study on parasites of the amphipod Paracalliope fluviatilis. Parasitology 2011, 138, 1176–1182. [Google Scholar] [CrossRef] [PubMed]
- Wuerthner, V.P.; Hua, J.; Hoverman, J.T. The benefits of coinfection: Trematodes alter disease outcomes associated with virus infection. J. Anim. Ecol. 2017, 86, 921–931. [Google Scholar] [CrossRef] [PubMed]
- Brooks, D.R.; Leon-Regagnon, V.; McLennan, D.A.; Zelmer, D. Ecological fitting as a determinant of the community structure of platyhelminth parasites of anurans. Ecology 2006, 87, S76–S85. [Google Scholar] [CrossRef]
- Aho, J.M. Helminth communities of amphibians and reptiles: Comparative approaches to understanding patterns and processes. In Parasite Communities: Patterns and Processes; Esch, G.W., Bush, A.O., Aho, J.M., Eds.; Springer: Dordrecht, The Netherlands, 1990; pp. 157–195. [Google Scholar] [CrossRef]
- Martins, P.M.; Poulin, R.; Gonçalves-Souza, T. Drivers of parasite β-diversity among anuran hosts depend on scale, realm and parasite group. Philos. Trans. R. Soc. B Biol. Sci. 2021, 376, 20200367. [Google Scholar] [CrossRef]
- Moss, W.E.; McDevitt-Galles, T.; Calhoun, D.M.; Johnson, P.T.J. Tracking the assembly of nested parasite communities: Using beta-diversity to understand variation in parasite richness and composition over time and scale. J. Anim. Ecol. 2020, 89, 1532–1542. [Google Scholar] [CrossRef] [PubMed]
- Bush, A.; Lafferty, K.; Lotz, J.; Shostak, A. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- Sinsch, U.; Lümkemann, K.; Rosar, K.; Schwarz, C.; Dehling, J.M. Acoustic niche partitioning in an anuran community inhabiting an Afromontane wetland (Butare, Rwanda). Afr. Zool. 2012, 47, 60–73. [Google Scholar] [CrossRef]
- Dehling, J.M.; Sinsch, U. Partitioning of morphospace in larval and adult reed frogs (Anura: Hyperoliidae: Hyperolius) of the Central African Albertine Rift. Zool. Anz. 2019, 280, 65–77. [Google Scholar] [CrossRef]
- Portik, D.M.; Bell, R.C.; Blackburn, D.C.; Bauer, A.M.; Barratt, C.D.; Branch, W.R.; Burger, M.; Channing, A.; Colston, T.J.; Conradie, W.; et al. Sexual Dichromatism Drives Diversification Within a Major Radiation of African Amphibians. Syst. Biol. 2019, 68, 859–875. [Google Scholar] [CrossRef]
- Mindje, M.; Tumushimire, L.; Sinsch, U. Diversity assessment of anurans in the Mugesera wetland (eastern Rwanda): Impact of habitat disturbance and partial recovery. Salamandra 2020, 56, 27–38. [Google Scholar]
- Tumushimire, L.; Mindje, M.; Sinsch, U.; Dehling, J.M. Anuran diversity of cultivated wetlands in Rwanda: Melting pot of generalists? Salamandra 2020, 56, 99–112. [Google Scholar]
- Dehling, D.M.; Dehling, J.M. Elevated alpha diversity in disturbed sites obscures regional decline and homogenization of amphibian diversity. bioRxiv 2021. [Google Scholar] [CrossRef]
- Sinsch, U.; Dehling, J.M.; Scheid, P.; Balczun, C. A new African species of parasitic Dero (Annelida, Clitellata, Naididae) in the urinary tract of reed frogs. Parasitol. Res. 2019, 118, 3359–3370. [Google Scholar] [CrossRef] [PubMed]
- Sinsch, U.; Dehling, J.M.; Scheid, P.; Balczun, C. Molecular Diversity of Nematode Parasites in Afrotropical Reed Frogs (Hyperolius spp.). Divers.-Basel 2020, 12, 265. [Google Scholar] [CrossRef]
- Sinsch, U.; Dehling, J.M.; Scheid, P.; Balczun, C. Alternative Development Strategies of Clinostomum chabaudi (Digenea) Metacercariae in Frog Hosts (Hyperolius spp.). Diversity 2021, 13, 93. [Google Scholar] [CrossRef]
- Anderson, R.C.; Chabaud, A.G.; Willmott, S. Keys to the Nematode Parasites of Vertebrates: Archival Volume; CABI: Wallingford, UK, 2009. [Google Scholar]
- Gibbons, L.M. Keys to the Nematode Parasites of Vertebrates: Supplementary Volume; CABI: Wallingford, UK, 2010; Volume 10. [Google Scholar]
- Gibson, D.I.; Jones, A.; Bray, R. (Eds.) Keys to Trematoda; CABI: Wallingford, UK, 2002; Volume 1, p. 544. [Google Scholar]
- Jones, A.; Bray, R.A.; Gibson, D.I. (Eds.) Keys to the Trematoda; CABI: Wallingford, UK, 2005; Volume 2, p. 768. [Google Scholar]
- Bray, R.A.; Gibson, D.I.; Jones, A. (Eds.) Keys to the Trematoda; CABI: Wallingford, UK, 2008; Volume 3, p. 805. [Google Scholar]
- Harman, W.J. A Review of the Subgenus Allodero (Oligochaeta: Naididae: Dero) with a Description of D. (A.) floridana n. sp. from Bufo terrestris. Trans. Am. Microsc. Soc. 1971, 90, 225–228. [Google Scholar] [CrossRef]
- Andrews, J.M.; Childress, J.N.; Iakovidis, T.J.; Langford, G.J. Elucidating the Life History and Ecological Aspects of Allodero hylae (Annelida: Clitellata: Naididae), A Parasitic Oligochaete of Invasive Cuban Tree Frogs in Florida. J. Parasitol. 2015, 101, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Oda, F.H.; Petsch, D.K.; Ragonha, F.H.; Batista, V.G.; Takeda, A.M.; Takemoto, R.M. Dero (Allodero) lutzi Michaelsen, 1926 (Oligochaeta: Naididae) associated with Scinax fuscovarius (Lutz, 1925) (Anura: Hylidae) from Semi-deciduous Atlantic Rain Forest, southern Brazil. Braz. J. Biol. 2015, 75, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Douglas, L.T. The Taxonomy of Nematotaeniid Cestodes. J. Parasitol. 1958, 44, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Mariaux, J. Cestode systematics: Any progress? Int. J. Parasitol. 1996, 26, 231–243. [Google Scholar] [CrossRef]
- Scholz, T.; Besprozvannykh, V.V.; Boutorina, T.E.; Choudhury, A.; Cribb, T.H.; Ermolenko, A.V.; Faltynkova, A.; Shedko, M.B.; Shimazu, T.; Smit, N.J. Trematode diversity in freshwater fishes of the Globe I: ‘Old World’. Syst. Parasitol. 2016, 93, 257–269. [Google Scholar] [CrossRef]
- Wilbert, N.; Schmeier, U. Survey of the Intestinal Opalines and Ciliates in Central European Amphibians. Arch. Protistenkd. 1982, 125, 271–285. [Google Scholar] [CrossRef]
- Delvinquier, B.L.J.; Markus, M.B.; Passmore, N.I. Opalinidae in African Anura. IV. Genus Protoopalina. Syst. Parasitol. 1995, 30, 81–120. [Google Scholar] [CrossRef]
- Li, M.; Hu, G.; Li, C.; Zhao, W.S.; Zou, H.; Li, W.X.; Wu, S.G.; Wang, G.T.; Ponce-Gordo, F. Morphological and molecular characterization of a new ciliate Nyctotheroides grimi n. sp. (Armophorea, Clevelandellida) from Chinese frogs. Acta Trop. 2020, 208, 105531. [Google Scholar] [CrossRef] [PubMed]
- Colwell, R.K.; Elsensohn, J.E. EstimateS turns 20: Statistical estimation of species richness and shared species from samples, with non-parametric extrapolation. Ecography 2014, 37, 609–613. [Google Scholar] [CrossRef]
- Sørensen, T.A. A method of establishing groups of equal amplitude in plant sociology based on similarity of species content and its application to analyses of the vegetation on Danish commons. K. Dan. Vidensk. Selskab. 1948, 5, 1–34. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Sinnappah, N.D.; Lim, L.-H.S.; Rohde, K.; Tinsley, R.; Combes, C.; Verneau, O. A paedomorphic parasite associated with a neotenic amphibian host: Phylogenetic evidence suggests a revised systematic position for Sphyranuridae within anuran and turtle Polystomatoineans. Mol. Phyl. Evol. 2001, 18, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Kaci-Chaouch, T.; Verneau, O.; Desdevises, Y. Host specificity is linked to intraspecific variability in the genus Lamellodiscus (Monogenea). Parasitology 2008, 135, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Badets, M.; Verneau, O. Origin and evolution of alternative developmental strategies in amphibious sarcopterygian parasites (Platyhelminthes, Monogenea, Polystomatidae). Org. Divers. Evol. 2009, 9, 155–164. [Google Scholar] [CrossRef]
- Snyder, S.D.; Loker, E.S. Evolutionary relationships among the Schistosomatidae (Platyhelminthes: Digenea) and an Asian origin for Schistosoma. J. Parasitol. 2000, 86, 283–288. [Google Scholar] [CrossRef]
- Moszczynska, A.; Locke, S.A.; McLaughlin, J.D.; Marcogliese, D.J.; Crease, T.J. Development of primers for the mitochondrial cytochrome c oxidase I gene in digenetic trematodes (Platyhelminthes) illustrates the challenge of barcoding parasitic helminths. Mol. Ecol. Resour. 2009, 9, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Sinsch, U. Review: Skeletochronological assessment of demographic life-history traits in amphibians. Herpetol. J. 2015, 25, 5–13. [Google Scholar]
- Dollfus, R.P. Trematodes récoltés au Congo Belge par le Professeur Paul Brien (mai-août 1937). Ann. Mus. Congo Belge. C.-Zool. 1950, 1, 1–135. [Google Scholar]
- Meskal, F.H. Trematodes of anurans from Ethiopia. Arb. Univ. Bergen Mat.-Naturv. 1970, 1, 1–73. [Google Scholar]
- Aisien, S.O.; Ugbo, A.D.; Ilavbare, A.N.; Ogunbor, O. Endoparasites of amphibians from South-Western Nigeria. Acta Parasitol. 2001, 46, 299–305. [Google Scholar]
- Aisien, M.; Sampson, S.A.; Amuzie, C. Anuran parasites from three biotopes in Rivers State, Nigeria. Niger. J. Parasitol. 2017, 38, 129–135. [Google Scholar] [CrossRef]
- Okere, S.; Joseph, F.; Amuzie, C. Endo-parasitic helminths of amphibians, Ptychadena mascareniensis and Ptychadena pumilio at Rumuji-Emohua, Rivers state, Nigeria. Afr. J. Environ. Nat. Sci. Res. 2019, 2, 71–76. [Google Scholar]
- Edo-Taiwo, O.; Aisien, M. Helminth parasitic infections of leaf litter frogs (Arthroleptis and Phrynobatrachus spp.) from cocoa plantations in southern Nigeria. Niger. J. Parasitol. 2020, 41, 93–100. [Google Scholar] [CrossRef]
- Imasuen, A.; Aisien, M. Helminth parasites of Silurana tropicalis from the Okomu National Park, Edo State, Nigeria. Niger. J. Parasitol. 2015, 36, 61–66. [Google Scholar]
- Imasuen, A.; Aisien, M. Helminth parasitofauna of Ptychadena species from altered rainforest. Niger. J. Parasitol. 2019, 40, 193–197. [Google Scholar] [CrossRef]
- Imasuen, A.A.; Ojo, O.O.; Adesina, O.O.; Enabuele, E.E.; Aisien, M. Parasitic endohelminths of tree frogs from two rainforest habitats in Edo State, Nigeria. Zoologist 2020, 17, 13–17. [Google Scholar] [CrossRef]
- Tinsley, R.C. Parasites of Xenopus. In The Biology of Xenopus; Tinsley, R.C., Kobel, H.R., Eds.; Clarendon Press: London, UK, 1996; pp. 233–262. [Google Scholar]
- Paul, A.A. Life History Studies of North American Fresh-Water Polystomes. J. Parasitol. 1938, 24, 489–510. [Google Scholar] [CrossRef]
- Badets, M.; Morrison, C.; Verneau, O. Alternative parasite development in transmission strategies: How time flies! J. Evol. Biol. 2010, 23, 2151–2162. [Google Scholar] [CrossRef] [PubMed]
- Tinsley, R.C. Correlation between life-cycle of Eupolystoma alluaudi (Monogenea) and ecology of its host Bufo regularis. Parasitology 1975, 71, R16–R17. [Google Scholar]
- Hamann, M.I. Seasonal maturation of Glypthelmins vitellinophilum (Trematoda: Digenea) in Lysapsus limellus (Anura: Pseudidae) from an Argentinian subtropical permanent pond. Rev. Bras. Biol. 2006, 66, 85–93. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bolek, M.G.; Snyder, S.D.; Janovy, J., Jr. Alternative Life Cycle Strategies and Colonization of Young Anurans by Gorgoderina attenuata in Nebraska. J. Parasitol. 2009, 95, 604–616. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nollen, P.M.; Alberico, R.A. Absence of testes in Gorgoderina attenuata. J. Parasitol. 1972, 58, 1011–1012. [Google Scholar] [CrossRef] [PubMed]
- Hoole, D.; Mitchell, J.B. Gorgoderina vitelliloba (Digenea: Gorgoderidae) in Hyla arborea. Z. Parasitenkd.-Parasitol. Res. 1984, 70, 829–831. [Google Scholar] [CrossRef]
- Johnson, P.T.J.; Hoverman, J.T. Parasite diversity and coinfection determine pathogen infection success and host fitness. Proc. Natl. Acad. Sci. USA 2012, 109, 9006–9011. [Google Scholar] [CrossRef] [PubMed]
- Hoverman, J.T.; Hoye, B.J.; Johnson, P.T.J. Does timing matter? How priority effects influence the outcome of parasite interactions within hosts. Oecologia 2013, 173, 1471–1480. [Google Scholar] [CrossRef]
- Johnson, P.T.; McKenzie, V.J. Effects of environmental change on helminth infections in amphibians: Exploring the emergence of Ribeiroia and Echinostoma infections in North America. In The Biology of Echinostomes; Toledo, R., Fried, B., Eds.; Springer: New York, NY, USA, 2009; pp. 249–280. [Google Scholar] [CrossRef]
- Orlofske, S.A.; Belden, L.K.; Hopkins, W.A. Effects of Echinostoma trivolvis metacercariae infection during development and metamorphosis of the wood frog (Lithobates sylvaticus). Comp. Biochem. Physiol. Mol. Integr. Physiol. 2017, 203, 40–48. [Google Scholar] [CrossRef] [PubMed]
- LaFonte, B.E.; Johnson, P.T.J. Experimental infection dynamics: Using immunosuppression and in vivo parasite tracking to understand host resistance in an amphibian-trematode system. J. Exp. Biol. 2013, 216, 3700–3708. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).