
Bacterial Communities Associated with the Cycling of Non-Starch Polysaccharides and Phytate in Aquaponics Systems

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Aquaponics System and Samples Collection
2.2. Prokaryote Diversity through 16S rRNA Gene Sequencing
zOTUs Reported to Carry Phytases and Carbohydrases Enzymes
2.3. Isolation and Identification of Bacterial Strains
2.4. Screening for Carbohydrase-Producing Bacteria
2.4.1. Screening for Non-Starch Polysaccharide-Degrading Bacteria
2.4.2. Screening for Starch-Degrading Bacteria
2.5. Screening for Phytase-Producing Bacteria
2.6. Detection of Phytase Genes by PCR Amplification Using Degenerate Primers
2.7. Statistical Analyses
3. Results
3.1. 16S rRNA Diversity of Bacterial Communities in Fish and Plant Compartments
3.2. Multivariate Correlation Network Clustering of Bacteria Taxa
3.3. Diversity and Abundance of Microbes Reported to Carry Putative Phytases and Carbohydrase Enzyme Genes
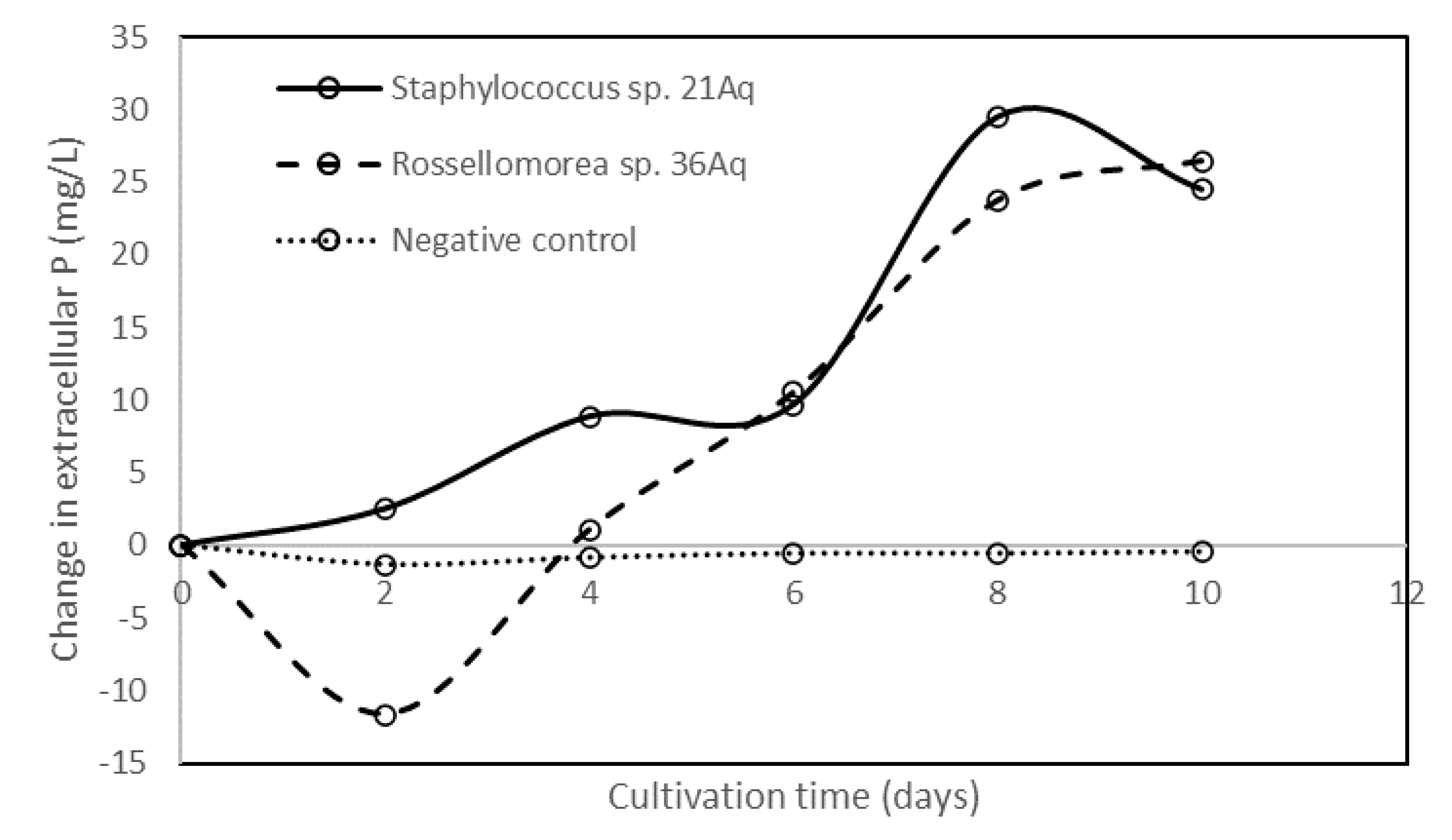
3.4. Phytase-, Xylanase-, β-glucanase-, Cellulase-, and Amylase-Producing Bacterial Isolates
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Delaide, B.; Goddek, S.; Gott, J.; Soyeurt, H.; Jijakli, M.H. Lettuce (Lactuca sativa L. var. Sucrine) growth performance in complemented aquaponic solution outperforms hydroponics. Water 2016, 8, 467. [Google Scholar] [CrossRef]
- Buzby, K.M.; Waterland, N.L.; Semmens, K.J.; Lin, L.-S. Evaluating aquaponic crops in a freshwater flow-through fish culture system. Aquaculture 2016, 460, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Goddek, S.; Espinal, C.A.; Delaide, B.; Jijakli, M.H.; Schmautz, Z.; Wuertz, S.; Keesman, K.J. Navigating towards decoupled aquaponic systems: A system dynamics design approach. Water 2016, 8, 303. [Google Scholar] [CrossRef]
- Rakocy, J.; Masser, M.P.; Losordo, T. Recirculating Aquaculture Tank Production Systems: Aquaponics-Integrating Fish and Plant Culture; Oklahoma State University: Brian, TX, USA, 2016. [Google Scholar]
- Al-Hafedh, Y.S.; Alam, A.; Beltagi, M.S. Food production and water conservation in a recirculating aquaponic system in Saudi Arabia at different ratios of fish feed to plants. J. World Aquac. Soc. 2008, 39, 510–520. [Google Scholar] [CrossRef]
- Coad, M.T.; Petrea, S.M.; Cristea, V.; Dediu, L.; Bandi, C.; Turek-Rahoveanu, M.; Zugravu, A.G.; Rahoveanu, A.T.; Mocuta, D.N. Water Quality in Aquaponic Integrated Systems: An Overview of the Literature Innovation Management and Education Excellence Vision 2020: Regional Development to Global Economic Growth; PUBLONS: London, UK, 2016. [Google Scholar]
- Turkmen, G.; Guner, Y. Aquaponic (integrating fish and plant culture) systems. In International Symposium on Sustainable Development, Science Book; Wiley: London, UK, 2010; pp. 657–666. [Google Scholar]
- Munguia-Fragozo, P.; Alatorre-Jacome, O.; Rico-Garcia, E.; Torres-Pacheco, I.; Cruz-Hernandez, A.; Ocampo-Velazquez, R.V.; Garcia-Trejo, J.F.; Guevara-Gonzalez, R.G. Perspective for aquaponic systems: “omic” technologies for microbial community analysis. BioMed Res. Int. 2015, 2015, 480386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasozi, N.; Kaiser, H.; Wilhelmi, B. Metabarcoding analysis of bacterial communities associated with media grow bed zones in an aquaponic system. Int. J. Microbiol. 2020, 2020, 8884070. [Google Scholar] [CrossRef]
- Rurangwa, E.; Verdegem, M.C. Microorganisms in recirculating aquaculture systems and their management. Rev. Aquac. 2015, 7, 117–130. [Google Scholar] [CrossRef]
- Tyson, R.V.; Treadwell, D.D.; Simonne, E.H. Opportunities and challenges to sustainability in aquaponic systems. HortTechnology 2011, 21, 6–13. [Google Scholar] [CrossRef]
- Leonard, N.; Blancheton, J.; Guiraud, J. Populations of heterotrophic bacteria in an experimental recirculating aquaculture system. Aquac. Eng. 2000, 22, 109–120. [Google Scholar] [CrossRef]
- Schmautz, Z.; Graber, A.; Jaenicke, S.; Goesmann, A.; Junge, R.; Smits, T.H. Microbial diversity in different compartments of an aquaponics system. Arch. Microbiol. 2017, 199, 613–620. [Google Scholar] [CrossRef]
- Francis, G.; Makkar, H.P.; Becker, K. Antinutritional factors present in plant-derived alternate fish feed ingredients and their effects in fish. Aquaculture 2001, 199, 197–227. [Google Scholar] [CrossRef]
- Menezes-Blackburn, D.; Greiner, R. Enzymes Used in Animal Feed: Leading Technologies and Forthcoming Developments; Wiley Online Library: London, UK, 2014. [Google Scholar]
- Council, N.R. Nutrient Requirements of Fish and Shrimp; NRC: Washington, DC, USA, 2011.
- Da Silva Cerozi, B.; Fitzsimmons, K. Effect of dietary phytase on phosphorus use efficiency and dynamics in aquaponics. Aquac. Int. 2017, 25, 1227–1238. [Google Scholar] [CrossRef]
- Liebert, F.; Portz, L. Nutrient utilization of Nile tilapia Oreochromis niloticus fed plant based low phosphorus diets supplemented with graded levels of different sources of microbial phytase. Aquaculture 2005, 248, 111–119. [Google Scholar] [CrossRef]
- Jorquera, M.; Martínez, O.; Maruyama, F.; Marschner, P.; De La Luz Mora, M. Current and future biotechnological applications of bacterial phytases and phytase-producing bacteria. Microbes Environ. 2008, 23, 182–191. [Google Scholar] [CrossRef] [Green Version]
- Hill, J.E.; Kysela, D.; Elimelech, M. Isolation and assessment of phytate-hydrolysing bacteria from the DelMarVa Peninsula. Environ. Microbiol. 2007, 9, 3100–3107. [Google Scholar] [CrossRef]
- Hien, T.T.T.; Be, T.T.; Lee, C.M.; Bengtson, D.A. Development of formulated diets for snakehead (Channa striata and Channa micropeltes): Can phytase and taurine supplementation increase use of soybean meal to replace fish meal? Aquaculture 2015, 448, 334–340. [Google Scholar] [CrossRef]
- Krogdahl, Å.; Hemre, G.I.; Mommsen, T. Carbohydrates in fish nutrition: Digestion and absorption in postlarval stages. Aquac. Nutr. 2005, 11, 103–122. [Google Scholar] [CrossRef]
- MR DNA. Shallowater, TX, USA. 2021. Available online: https://www.mrdnalab.com/ (accessed on 1 November 2020).
- Pardee, A.B.; Jacob, F.; Monod, J. The genetic control and cytoplasmic expression of “inducibility” in the synthesis of β-galactosidase by E. coli. J. Mol. Biol. 1959, 1, 165–178. [Google Scholar] [CrossRef]
- Heuer, H.; Krsek, M.; Baker, P.; Smalla, K.; Wellington, E. Analysis of actinomycete communities by specific amplification of genes encoding 16S rRNA and gel-electrophoretic separation in denaturing gradients. Appl. Environ. Microbiol. 1997, 63, 3233–3241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subajini, M.; Sandrasegarampillai, B.; Vasanthy, A. Screening and identification of a thermophilic and alkalophilic bacterium producing xylanase. Adv. Appl. Sci. Res. 2012, 3, 242–250. [Google Scholar]
- Maravi, P.; Kumar, A. Isolation, Screening and Identification of Cellulolytic Bacteria from Soil. Biotechnol. J. Int. 2020, 24, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Fooladi, J.; Sajjadian, A. Screening the thermophilic and hyperthermophilic bacterial population of three Iranian hot-springs to detect the thermostable α-amylase producing strain. Iran. J. Microbiol. 2010, 2, 46–50. [Google Scholar]
- Ohno, T.; Zibilske, L.M. Determination of low concentrations of phosphorus in soil extracts using malachite green. Soil Sci. Soc. Am. J. 1991, 55, 892–895. [Google Scholar] [CrossRef]
- Huang, H.; Shi, P.; Wang, Y.; Luo, H.; Shao, N.; Wang, G.; Yang, P.; Yao, B. Diversity of beta-propeller phytase genes in the intestinal contents of grass carp provides insight into the release of major phosphorus from phytate in nature. Appl. Environ. Microbiol. 2009, 75, 1508–1516. [Google Scholar] [CrossRef] [Green Version]
- Tye, A.; Siu, F.; Leung, T.; Lim, B. Molecular cloning and the biochemical characterization of two novel phytases from B. subtilis 168 and B. licheniformis. Appl. Microbiol. Biotechnol. 2002, 59, 190–197. [Google Scholar] [PubMed]
- Huang, H.; Zhang, R.; Fu, D.; Luo, J.; Li, Z.; Luo, H.; Shi, P.; Yang, P.; Diao, Q.; Yao, B. Diversity, abundance and characterization of ruminal cysteine phytases suggest their important role in phytate degradation. Environ. Microbiol. 2011, 13, 747–757. [Google Scholar] [CrossRef]
- Ondov, B.D.; Bergman, N.H.; Phillippy, A.M. Interactive metagenomic visualization in a Web browser. BMC Bioinform. 2011, 12, 385. [Google Scholar] [CrossRef] [Green Version]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Köhler, J.; Baumbach, J.; Taubert, J.; Specht, M.; Skusa, A.; Rüegg, A.; Rawlings, C.; Verrier, P.; Philippi, S. Graph-based analysis and visualization of experimental results with ONDEX. Bioinformatics 2006, 22, 1383–1390. [Google Scholar] [CrossRef] [Green Version]
- Joo, H.-S.; Hirai, M.; Shoda, M. Characteristics of ammonium removal by heterotrophic nitrification-aerobic denitrification by Alcaligenes faecalis No. 4. J. Biosci. Bioeng. 2005, 100, 184–191. [Google Scholar] [CrossRef]
- Sharrer, M.J.; Summerfelt, S.T.; Bullock, G.L.; Gleason, L.E.; Taeuber, J. Inactivation of bacteria using ultraviolet irradiation in a recirculating salmonid culture system. Aquac. Eng. 2005, 33, 135–149. [Google Scholar] [CrossRef] [Green Version]
- Sugita, H.; Nakamura, H.; Shimada, T. Microbial communities associated with filter materials in recirculating aquaculture systems of freshwater fish. Aquaculture 2005, 243, 403–409. [Google Scholar] [CrossRef]
- Leonard, N.; Guiraud, J.; Gasset, E.; Cailleres, J.; Blancheton, J. Bacteria and nutrients—Nitrogen and carbon—In a recirculating system for sea bass production. Aquac. Eng. 2002, 26, 111–127. [Google Scholar] [CrossRef]
- Eck, M.; Sare, A.R.; Massart, S.; Schmautz, Z.; Junge, R.; Smits, T.H.; Jijakli, M.H. Exploring bacterial communities in aquaponic systems. Water 2019, 11, 260. [Google Scholar] [CrossRef] [Green Version]
- Nishioka, T.; Elsharkawy, M.M.; Suga, H.; Kageyama, K.; Hyakumachi, M.; Shimizu, M. Development of culture medium for the isolation of Flavobacterium and Chryseobacterium from rhizosphere soil. Microbes Environ. 2016, 31, 104–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ladeira, S.A.; Cruz, E.; Delatorre, A.B.; Barbosa, J.B.; Martins, M.L.L. Cellulase production by thermophilic Bacillus sp: SMIA-2 and its detergent compatibility. Electron. J. Biotechnol. 2015, 18, 110–115. [Google Scholar] [CrossRef] [Green Version]
- Marimuthu, M.; Sorimuthu, A.; Muruganantham, S. Production and Optimization of Xylanase Enzyme from Bacillus subtilis using Agricultural Wastes by Solid State Fermentation. Int. J. Pharm. Investig. 2019, 9, 169–173. [Google Scholar] [CrossRef] [Green Version]
- Subramaniyan, S.; Prema, P. Biotechnology of microbial xylanases: Enzymology, molecular biology, and application. Crit. Rev. Biotechnol. 2002, 22, 33–64. [Google Scholar] [CrossRef] [PubMed]
- Shakir, H.A.; Anwar, A.; Irfan, M.; Khan, M.; Ali, S.; Qazi, J.I. Statistical Optimization of Xylanase from Bacillus licheniformis Using Banana Peels in Submerged Fermentation. Iran. J. Sci. Technol. Trans. A Sci. 2020, 44, 981–991. [Google Scholar] [CrossRef]
- Singh, A.; Sharma, D.; Varghese, L.M.; Mahajan, R. Fast flow rate processes for purification of alkaline xylanase isoforms from Bacillus pumilus AJK and their biochemical characterization for industrial application purposes. Biotechnol. Prog. 2020, 36, e2898. [Google Scholar] [CrossRef]
- Pauly, M.; Keegstra, K. Cell-wall carbohydrates and their modification as a resource for biofuels. Plant J. 2008, 54, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Irfan, M.; Mushtaq, Q.; Tabssum, F.; Shakir, H.A.; Qazi, J.I. Carboxymethyl cellulase production optimization from newly isolated thermophilic Bacillus subtilis K-18 for saccharification using response surface methodology. AMB Express 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elayaraja, S.; Velvizhi, T.; Maharani, V.; Mayavu, P.; Vijayalakshmi, S.; Balasubramanian, T. Thermostable α-amylase production by Bacillus firmus CAS 7 using potato peel as a substrate. Afr. J. Biotechnol. 2011, 10, 11235–11238. [Google Scholar]
- Quesada-Ganuza, A.; Antelo-Varela, M.; Mouritzen, J.C.; Bartel, J.; Becher, D.; Gjermansen, M.; Hallin, P.F.; Appel, K.F.; Kilstrup, M.; Rasmussen, M.D. Identification and optimization of PrsA in Bacillus subtilis for improved yield of amylase. Microb. Cell Factories 2019, 18, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Van Dijl, J.; Hecker, M. Bacillus subtilis: From soil bacterium to super-secreting cell factory. Microb. Cell. Fact. 2013, 12, 3. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.A.; Jilani, G.; Akhtar, M.S.; Naqvi, S.M.S.; Rasheed, M. Phosphorus solubilizing bacteria: Occurrence, mechanisms and their role in crop production. J. Agric. Biol. Sci. 2009, 1, 48–58. [Google Scholar]
- Cheng, C.; Lim, B.L. Beta-propeller phytases in the aquatic environment. Arch. Microbiol. 2006, 185, 1–13. [Google Scholar] [CrossRef]
- Lim, B.L.; Yeung, P.; Cheng, C.; Hill, J.E. Distribution and diversity of phytate-mineralizing bacteria. ISME J. 2007, 1, 321. [Google Scholar] [CrossRef] [PubMed]
- Menezes-Blackburn, D.; Jorquera, M.A.; Greiner, R.; Gianfreda, L.; de la Luz Mora, M. Phytases and phytase-labile organic phosphorus in manures and soils. Crit. Rev. Environ. Sci. Technol. 2013, 43, 916–954. [Google Scholar] [CrossRef]
- Jorquera, M.A.; Gabler, S.; Inostroza, N.G.; Acuña, J.J.; Campos, M.A.; Menezes-Blackburn, D.; Greiner, R. Screening and characterization of phytases from bacteria isolated from Chilean hydrothermal environments. Microb. Ecol. 2018, 75, 387–399. [Google Scholar] [CrossRef]
- Fu, S.; Sun, J.; Qian, L.; Li, Z. Bacillus phytases: Present scenario and future perspectives. Appl. Biochem. Biotechnol. 2008, 151, 1–8. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Compartment | T (°C) | Light | pH | EC | P | K | Ca | Mg | Na | Cu | Zn |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Fish | 29.2 | 0.24 | 6.1 | 2.4 | 25 | 33.9 | 81.8 | 63.1 | 198 | 0.3 | <0.01 |
| Plant | 29.7 | 0.19 | 5.8 | 2.9 | 28 | 34.5 | 83.8 | 65.2 | 183 | 0.5 | <0.01 |
| Isolation ID | Length (bp) | NCBI Genbank Closest Match | Assigned Strain Name | Accession N. | |||
|---|---|---|---|---|---|---|---|
| Seq. ID | Identity | Q.Cover | Species | ||||
| SWAM_2Aq | 1359 | MT539995.1 | 100.00% | 100% | Bacillus subtilis | Bacillus subtilis 2Aq | OK626679 |
| SWAM_6Aq | 1318 | MT229334.1 | 98.79% | 100% | Bacillus velezensis | Bacillus velezensis 6Aq | OK626680 |
| SWAM_7Aq | 1287 | KM922586.1 | 98.45% | 100% | Bacillus subtilis | Bacillus subtilis 7Aq | OK626681 |
| SWAM_8Aq | 1299 | KM922586.1 | 98.00% | 100% | Bacillus subtilis | Bacillus subtilis 8Aq | OK626682 |
| SWAM_10Aq | 1317 | MF521557.1 | 93.06% | 100% | Bacillus cereus | Bacillus cereus 10Aq | OK626683 |
| SWAM_13Aq | 1395 | MN330286.1 | 99.63% | 97% | Bacillus subtilis | Bacillus subtilis 13Aq | OK626684 |
| SWAM_15Aq | 1405 | MK859953.1 | 99.15% | 100% | Bacillus licheniformis | Bacillus licheniformis 15Aq | OK626685 |
| SWAM_16Aq | 1327 | MW578436.1 | 91.40% | 99% | Bacillus licheniformis | Bacillus sp. 16Aq | OK626686 |
| SWAM_17Aq | 1422 | MH005066.1 | 91.82% | 99% | Bacillus infantis | Bacillus sp. 17Aq | OK626687 |
| SWAM_18Aq | 1416 | MT122066.1 | 99.36% | 100% | Bacillus tequilensis | Bacillus tequilensis 18Aq | OK626688 |
| SWAM_21Aq | 1446 | CP035288.1 | 93.05% | 99% | Staphylococcus epidermidis | Staphylococcus sp. 21Aq | OK626689 |
| SWAM_22Aq | 1347 | LS974830.1 | 93.87% | 100% | Rossellomorea marisflavi | Rossellomorea sp. 22Aq | OK626690 |
| SWAM_23Aq | 1380 | MN704425.1 | 98.04% | 99% | Bacillus stercoris | Bacillus subtilis 23Aq | OK626691 |
| SWAM_26Aq | 1414 | MT111029.1 | 98.46% | 100% | Bacillus subtilis | Bacillus tequilensis 26Aq | OK626692 |
| SWAM_28Aq | 1427 | MT326233.1 | 98.95% | 100% | Bacillus licheniformis | Bacillus licheniformis 28Aq | OK626693 |
| SWAM_29Aq | 1371 | MT611946.1 | 98.53% | 100% | Bacillus cereus | Bacillus cereus 29Aq | OK626694 |
| SWAM_33Aq | 1433 | CP053934.1 | 99.02% | 100% | Bacillus thuringiensis | Bacillus thuringiensis 33Aq | OK626695 |
| SWAM_36Aq | 1423 | KF933659.1 | 93.74% | 100% | Rossellomorea aquimaris | Rossellomorea sp. 36Aq | OK626696 |
| SWAM_37Aq | 1423 | MK942526.1 | 89.88% | 100% | Bacillus subtilis | Bacillus sp. 37Aq | OK626697 |
| Fish | Plant | Fish + Plant | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Shannon H | 5.96 | 5.17 | 0.21 | 5.90 | 5.23 | 0.18 | 6.24 | 5.20 | 0.12 |
| Dominance D | 0.01 | 0.01 | 0.00 | 0.01 | 0.01 | 0.00 | 0.00 | 0.01 | 0.00 |
| Evenness EH | 0.24 | 0.25 | 0.06 | 0.26 | 0.27 | 0.02 | 0.26 | 0.26 | 0.03 |
| Fish | % * | Plant | % * | Fish + Plant | % * | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| All | 1655 | 100 | 794 | 245 | 1424 | 100 | 733 | 188 | 1951 | 100 | 763 | 139 |
| BPP | 76 | 6 | 39 | 13 | 68 | 5 | 33 | 12 | 90 | 6 | 36 | 8 |
| HAP | 46 | 3 | 21 | 8 | 31 | 3 | 17 | 4 | 54 | 3 | 19 | 4 |
| PTP | 6 | <1 | 2 | 2 | 10 | <1 | 3 | 2 | 12 | <1 | 3 | 1 |
| All phytases | 98 | 9 | 50 | 17 | 87 | 8 | 41 | 14 | 120 | 8 | 46 | 10 |
| Β-glucanase | 28 | 2 | 15 | 3 | 23 | 1 | 9 | 4 | 31 | 1 | 12 | 3 |
| Xylanase | 108 | 6 | 55 | 18 | 77 | 4 | 34 | 12 | 117 | 5 | 44 | 11 |
| Cellulases | 220 | 18 | 116 | 32 | 220 | 23 | 116 | 32 | 250 | 17 | 116 | 20 |
| All carbohydrases | 232 | 26 | 120 | 34 | 199 | 19 | 103 | 21 | 266 | 23 | 112 | 18 |
| Source | Strain | Phy | XSA | BSA | CSA | ASA |
|---|---|---|---|---|---|---|
| Fish | Bacillus subtilis 2Aq | - | 24 | 14 | 25 | 26 |
| Fish | Bacillus velezensis 6Aq | - | 26 | 24 | 16 | 18 |
| Fish | Bacillus subtilis 7Aq | - | 25 | 27 | 18 | 14 |
| Fish | Bacillus subtilis 8Aq | - | - | 26 | 20 | 14 |
| Fish | Bacillus cereus 10Aq | - | - | - | - | 20 |
| Fish | Bacillus subtilis 13Aq | - | - | 49 | 27 | 20 |
| Fish | Bacillus licheniformis 15Aq | - | 26 | - | 15 | - |
| Fish | Bacillus sp. 16Aq | - | - | - | 12 | - |
| Plant | Bacillus sp. 17Aq | - | - | 33 | - | - |
| Plant | Bacillus tequilensis 18Aq | - | 23 | 46 | 26 | 22 |
| Plant | Staphylococcus sp. 21Aq | + | - | - | - | - |
| Plant | Rossellomorea sp. 22Aq | - | - | - | 25 | - |
| Plant | Bacillus subtilis 23Aq | - | 16 | 25 | - | - |
| Plant | Bacillus tequilensis 26Aq | - | 36 | - | 25 | 30 |
| Plant | Bacillus licheniformis 28Aq | - | - | - | 13 | - |
| Plant | Bacillus cereus 29Aq | - | - | - | 16 | - |
| Plant | Bacillus thuringiensis 33Aq | - | - | - | 16 | 21 |
| Plant | Rossellomorea sp. 36Aq | + | - | - | - | 13 |
| Plant | Bacillus sp. 37Aq | - | - | - | 15 | 10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menezes-Blackburn, D.; Al-Mahrouqi, N.; Al-Siyabi, B.; Al-Kalbani, A.; Greiner, R.; Dobretsov, S. Bacterial Communities Associated with the Cycling of Non-Starch Polysaccharides and Phytate in Aquaponics Systems. Diversity 2021, 13, 631. https://doi.org/10.3390/d13120631
Menezes-Blackburn D, Al-Mahrouqi N, Al-Siyabi B, Al-Kalbani A, Greiner R, Dobretsov S. Bacterial Communities Associated with the Cycling of Non-Starch Polysaccharides and Phytate in Aquaponics Systems. Diversity. 2021; 13(12):631. https://doi.org/10.3390/d13120631
Chicago/Turabian StyleMenezes-Blackburn, Daniel, Nahad Al-Mahrouqi, Buthaina Al-Siyabi, Adhari Al-Kalbani, Ralf Greiner, and Sergey Dobretsov. 2021. "Bacterial Communities Associated with the Cycling of Non-Starch Polysaccharides and Phytate in Aquaponics Systems" Diversity 13, no. 12: 631. https://doi.org/10.3390/d13120631
APA StyleMenezes-Blackburn, D., Al-Mahrouqi, N., Al-Siyabi, B., Al-Kalbani, A., Greiner, R., & Dobretsov, S. (2021). Bacterial Communities Associated with the Cycling of Non-Starch Polysaccharides and Phytate in Aquaponics Systems. Diversity, 13(12), 631. https://doi.org/10.3390/d13120631