
Global Characterization of XRN 5′-3′ Exoribonucleases and Their Responses to Environmental Stresses in Plants




Abstract
:1. Introduction
2. Materials and Methods
2.1. Genome-Wide Identification and Characterization of XRN Proteins
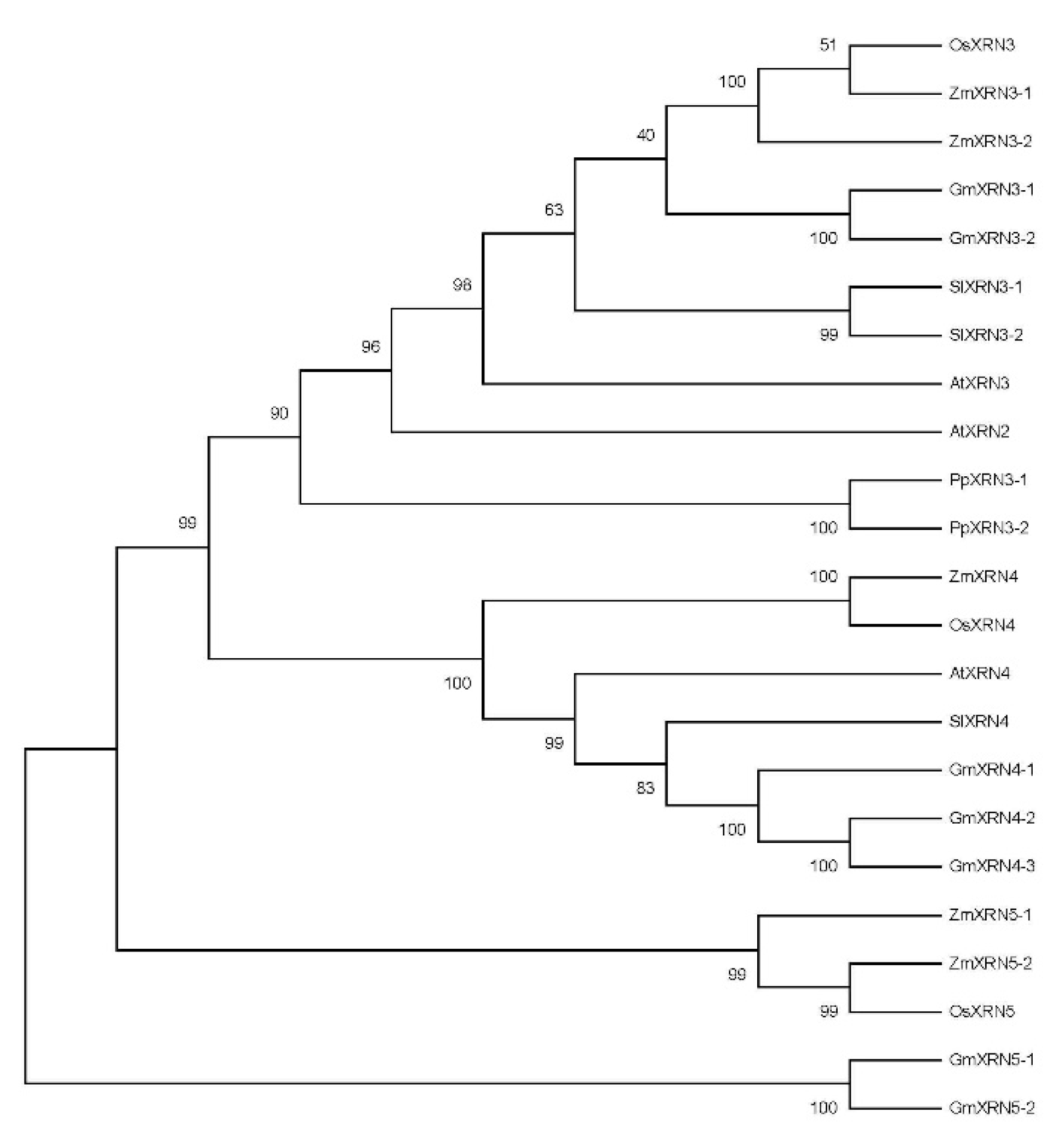
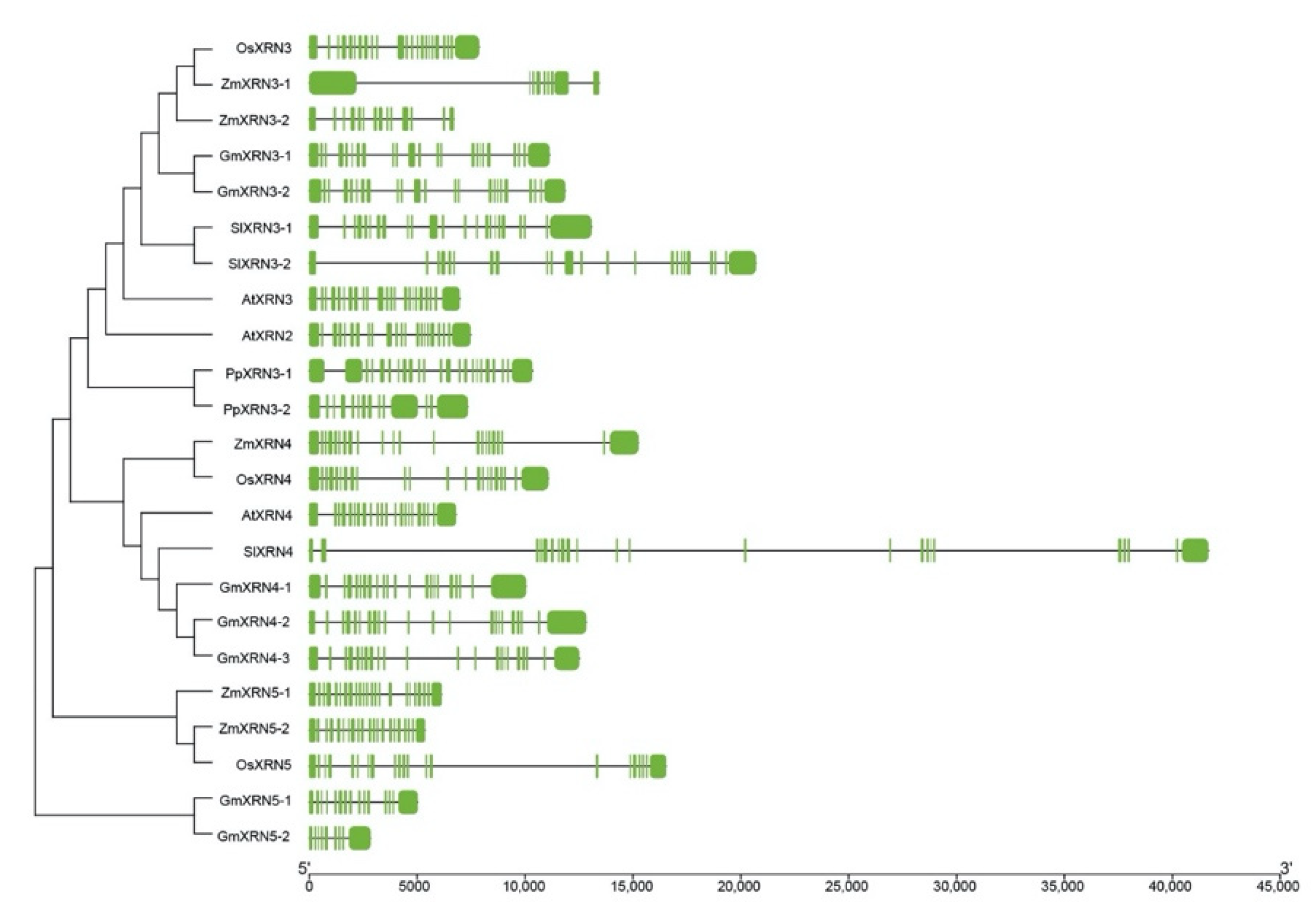
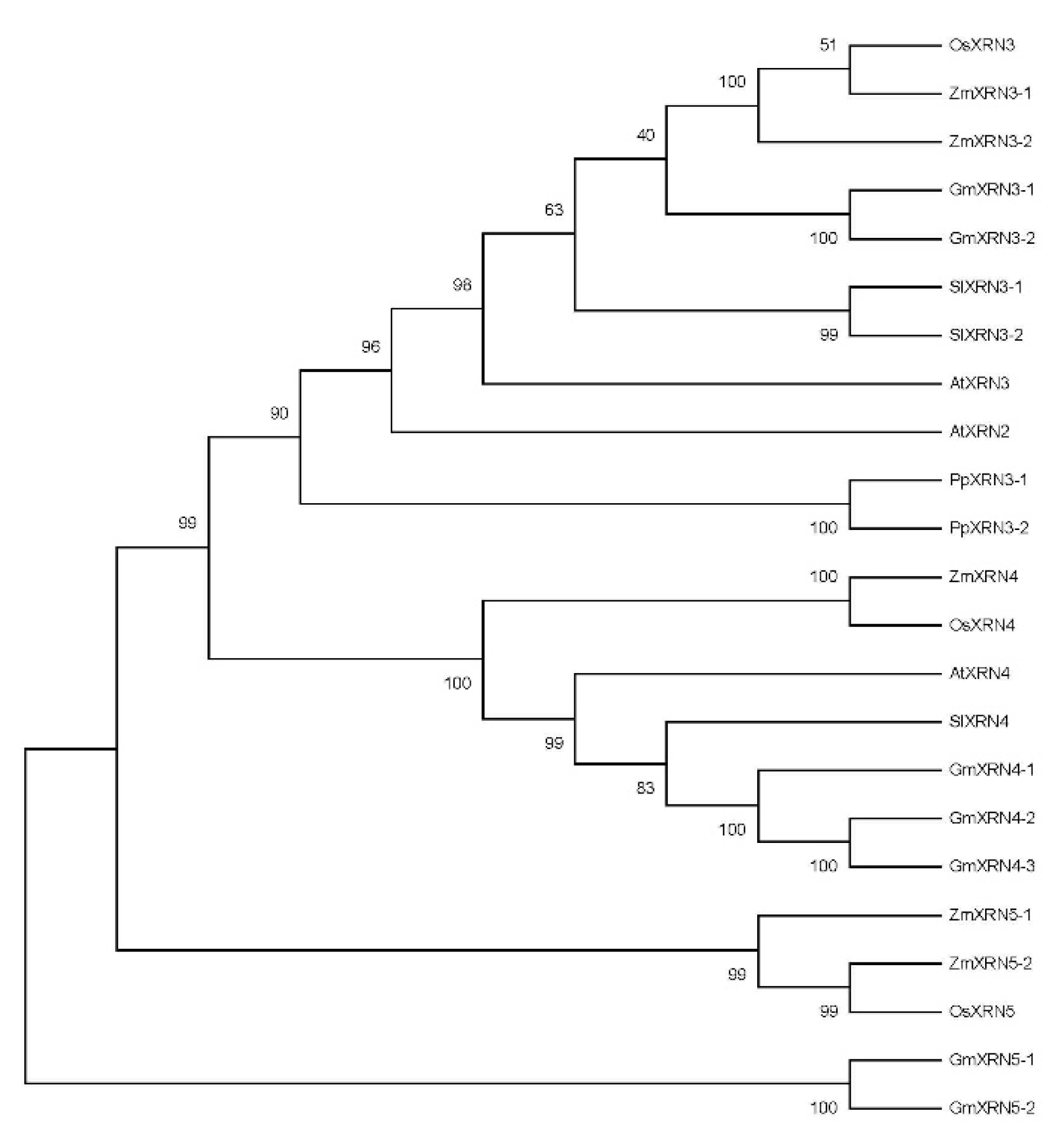
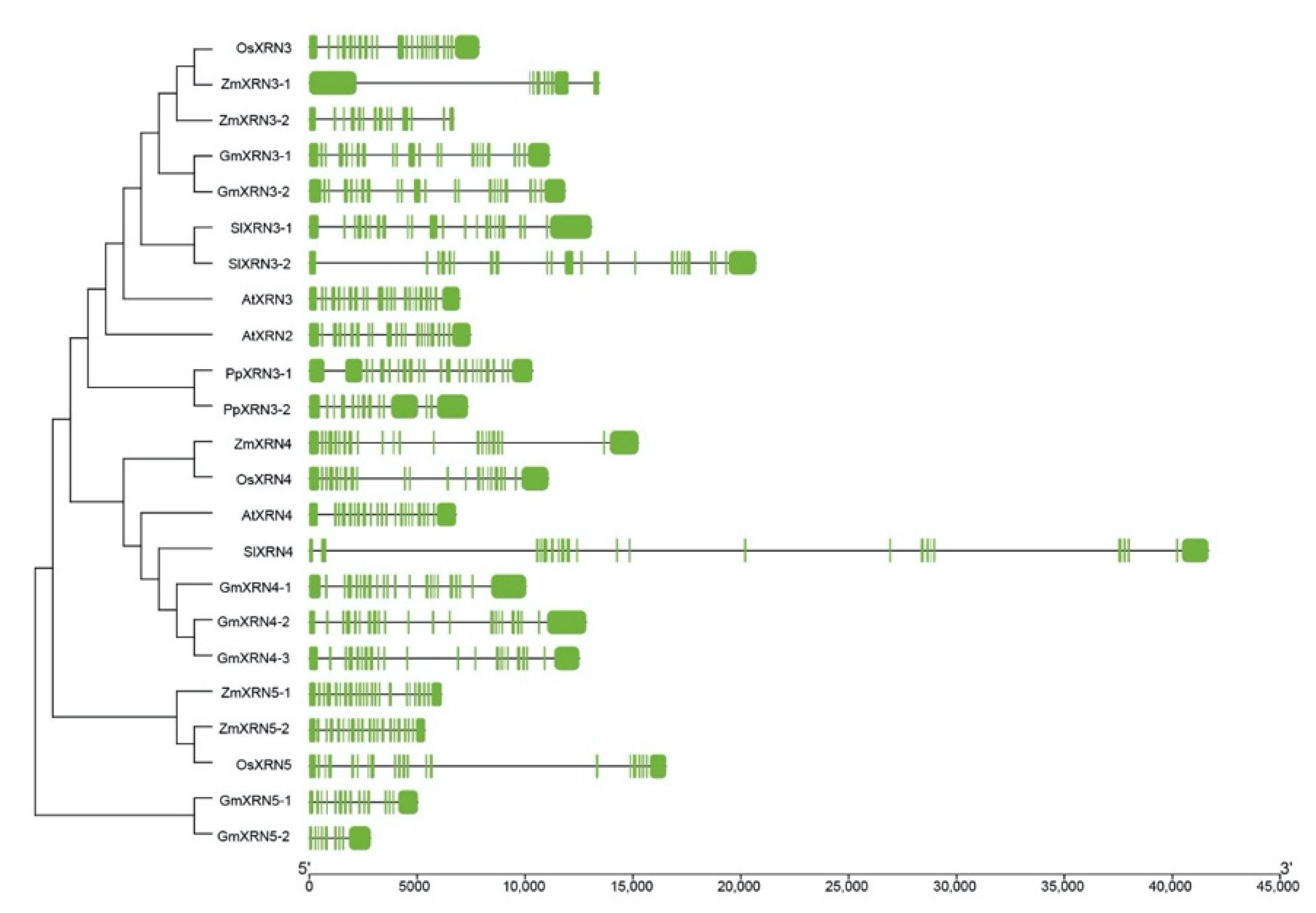
2.2. Construction of Polygenetic Tree and Gene Structure Visualization
2.3. Calculation of Biochemical Properties of XRN Proteins
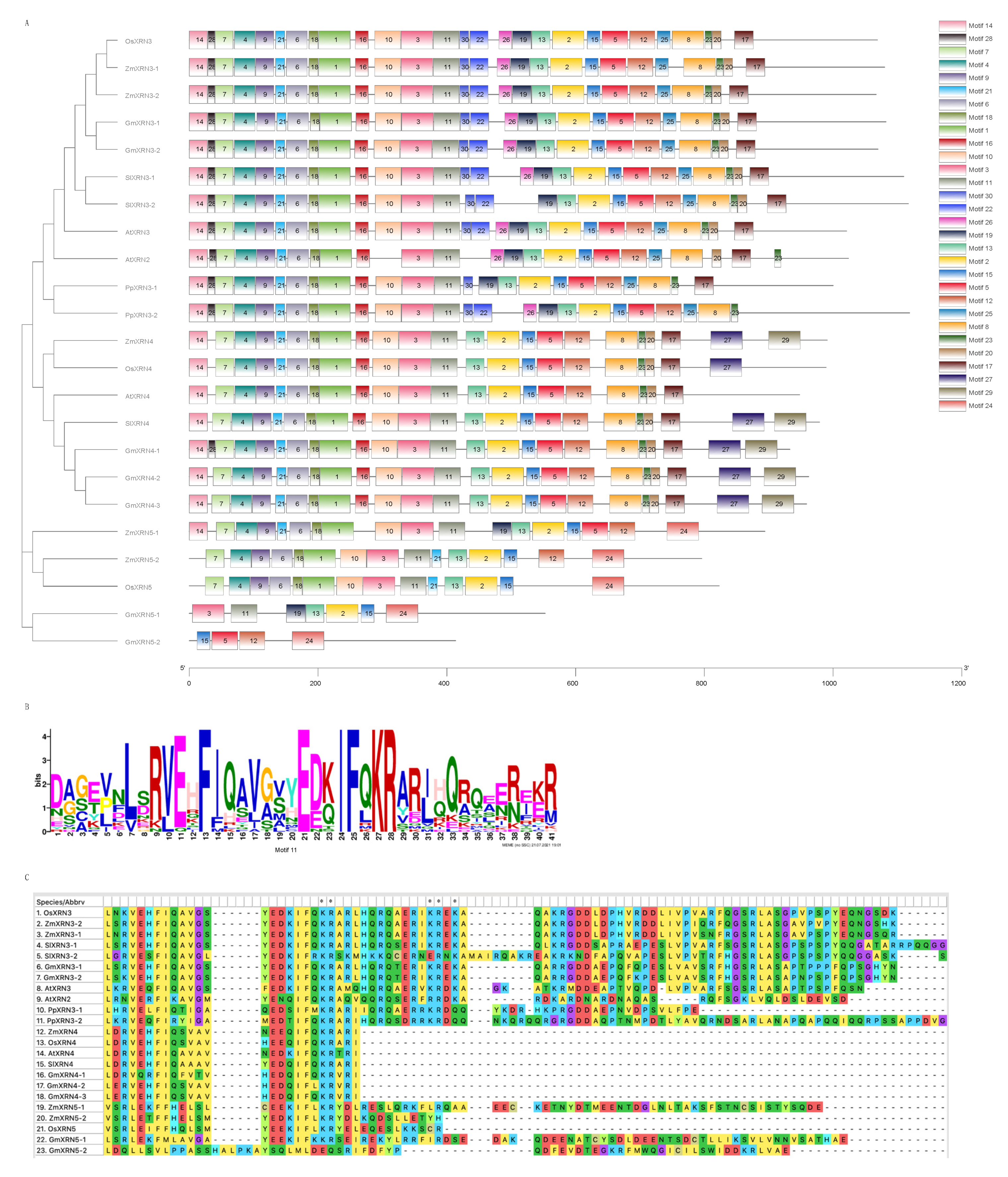
2.4. Conserved Protein Motif and Domain Analysis
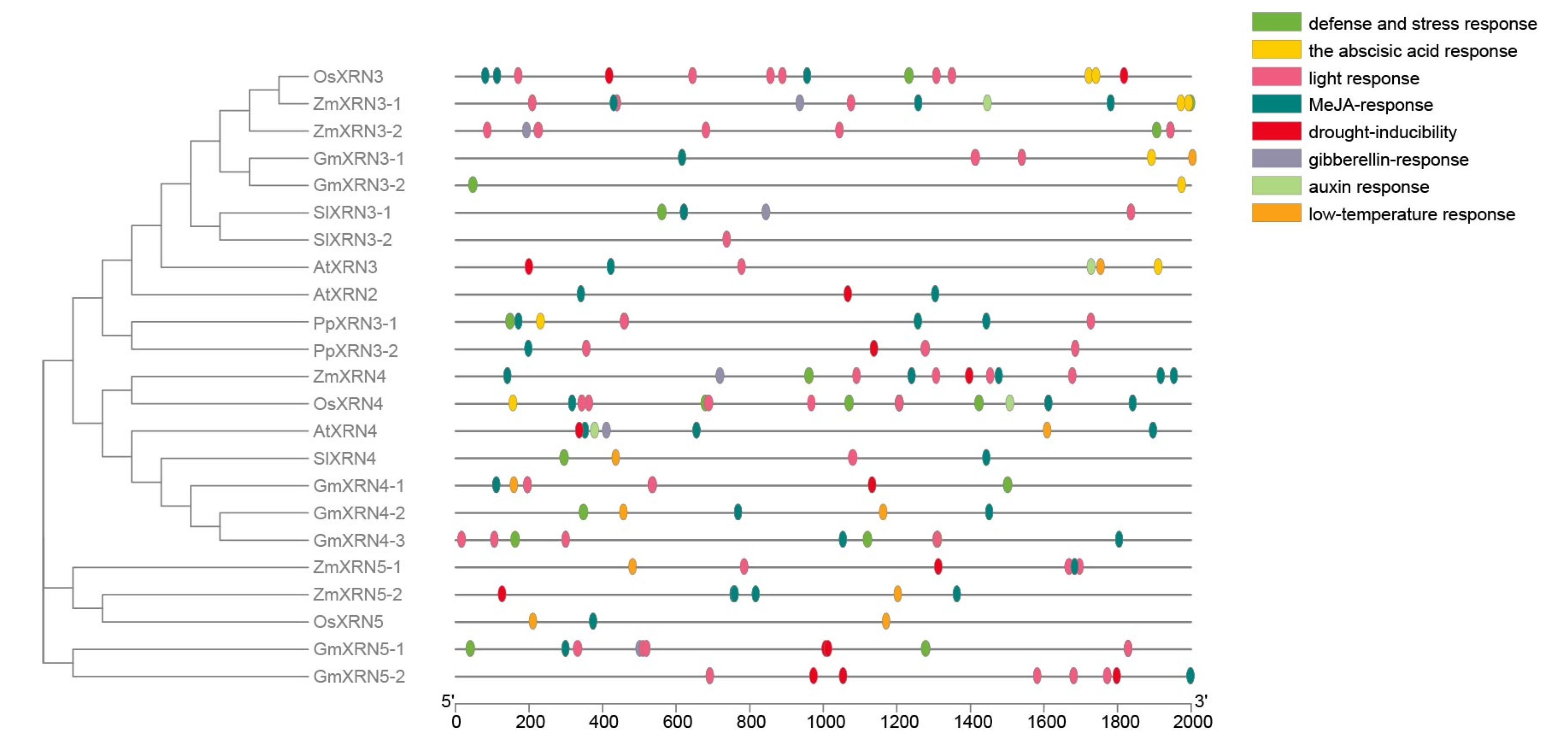
2.5. Promoter Cis-Element Analysis
2.6. Transcriptome Analysis for XRNs
2.7. XRN Protein Structure Prediction and Pairwise Structure Alignment
3. Results
3.1. Identification and Characterization of XRNs in Plant Species
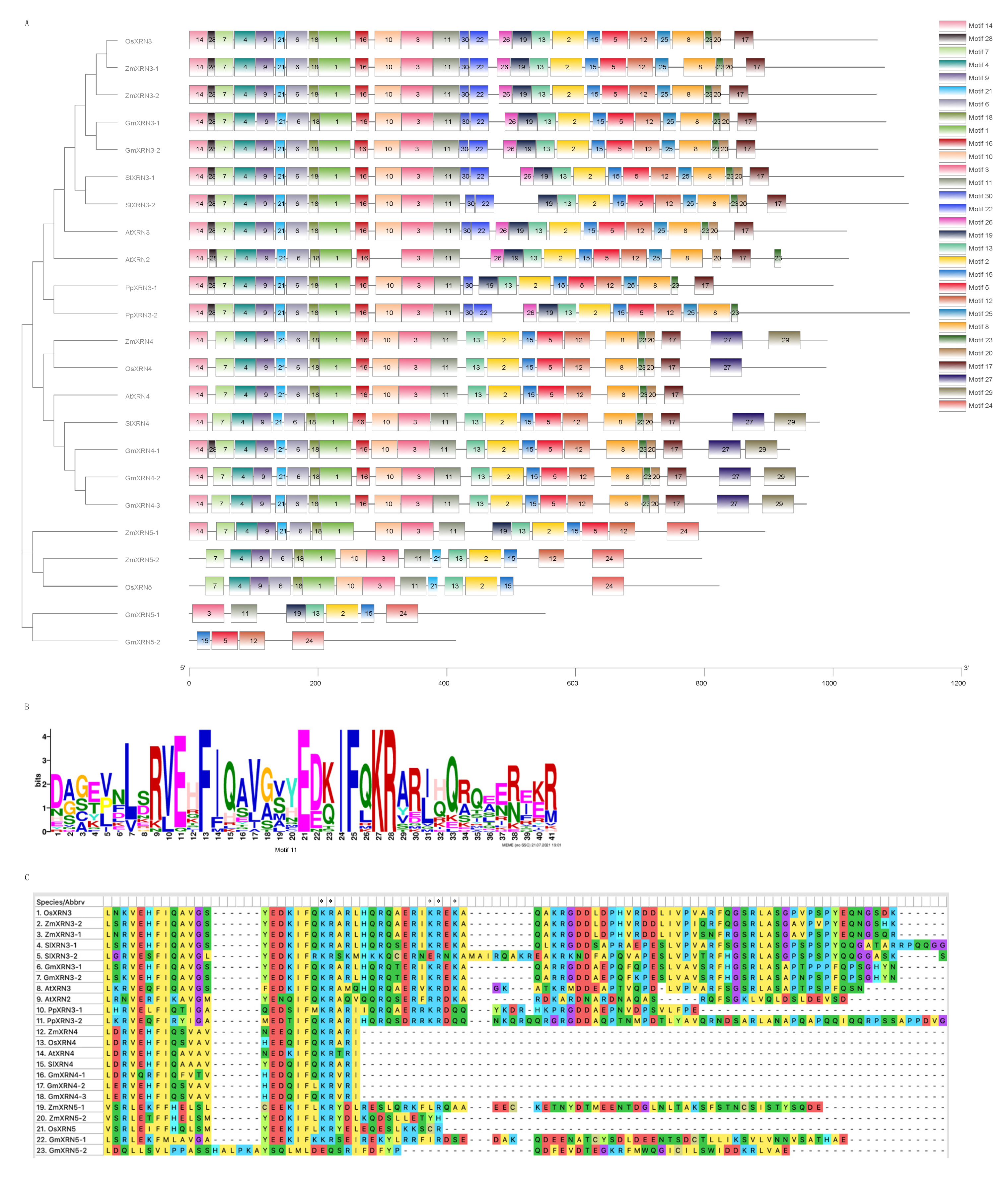
3.2. Gene Structure Variations in XRN Proteins
3.3. Characterization of Conserved Motifs and NLS in XRNs
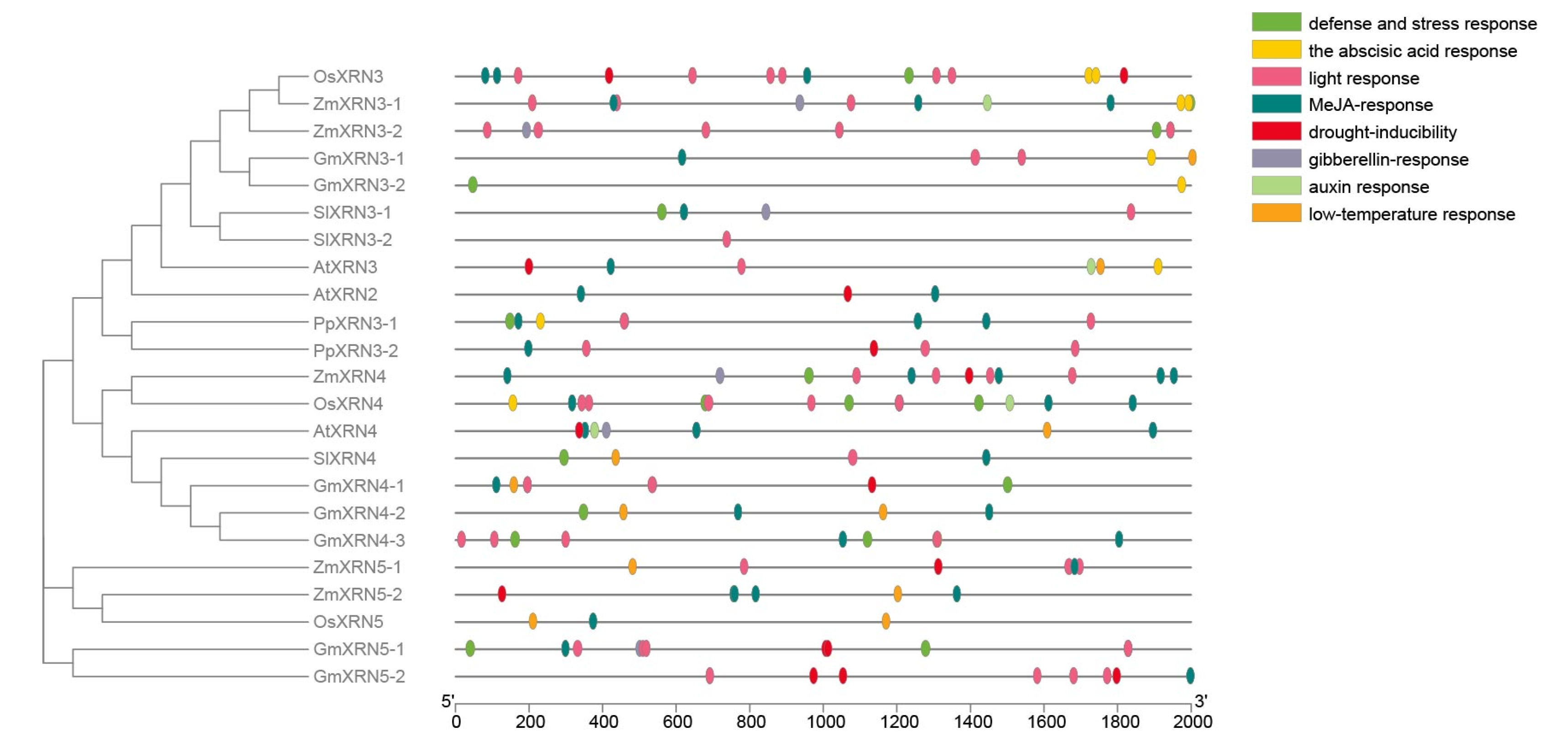
3.4. The Cis-Element Regulators in XRN Promoters
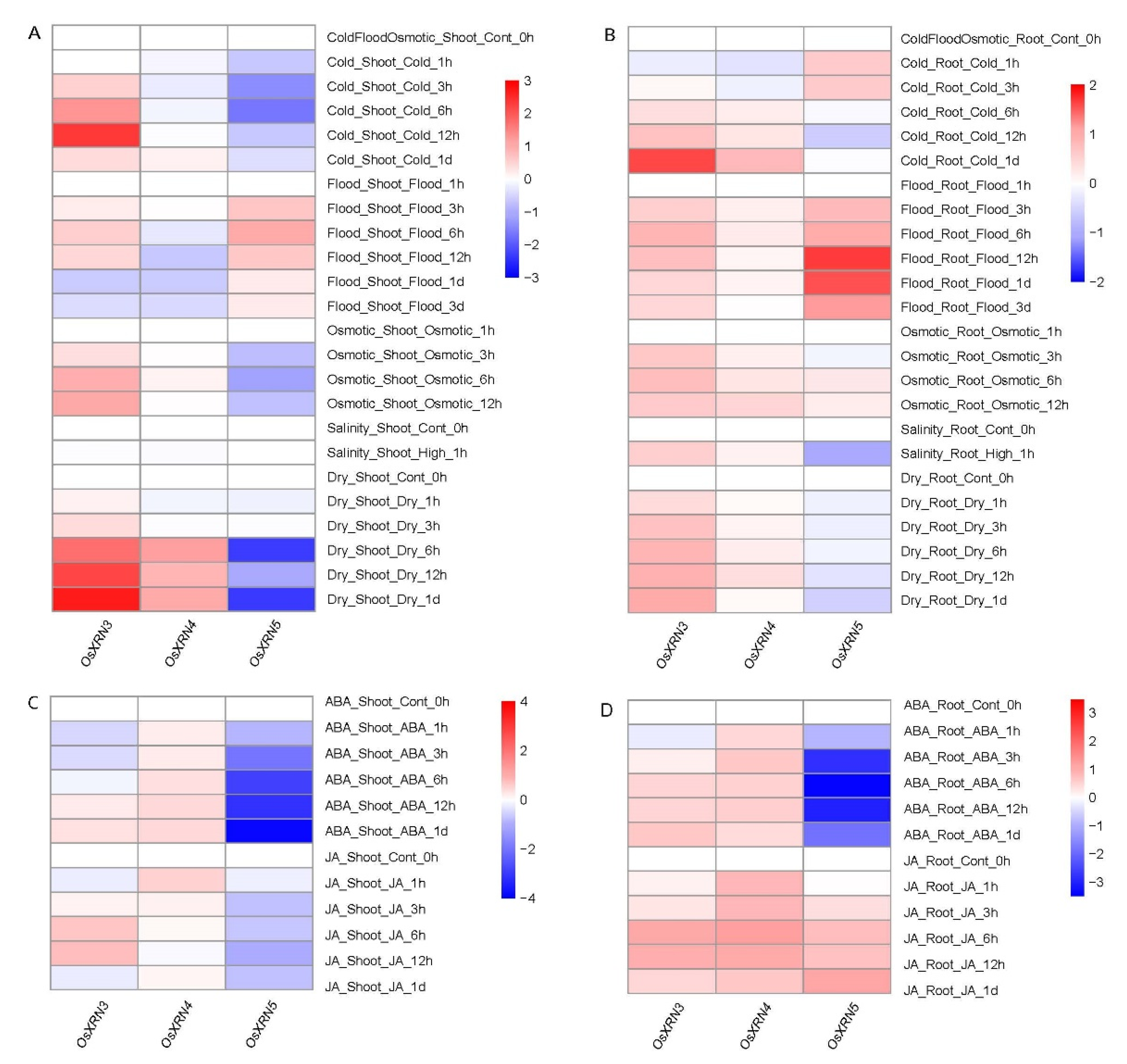
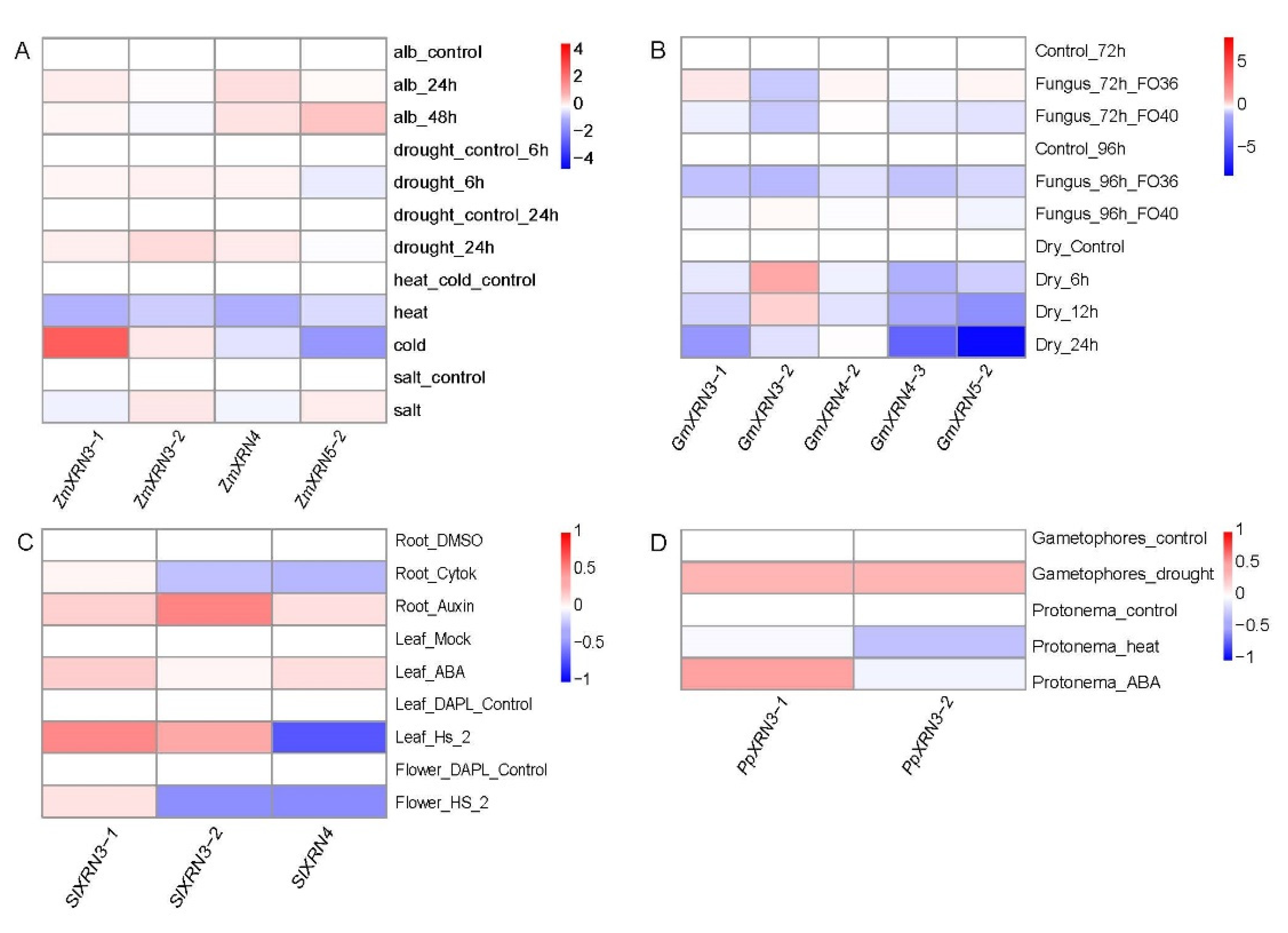
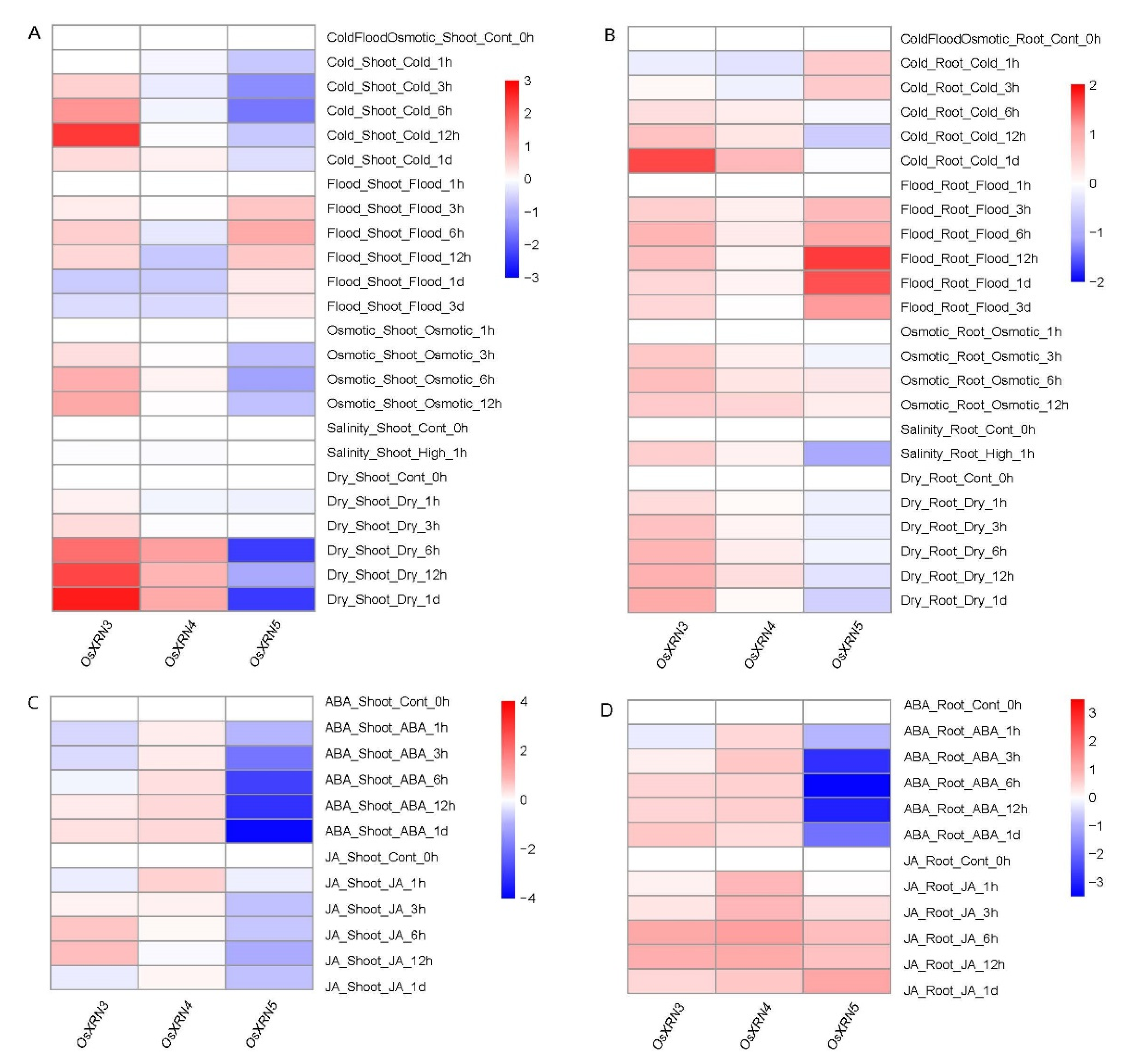
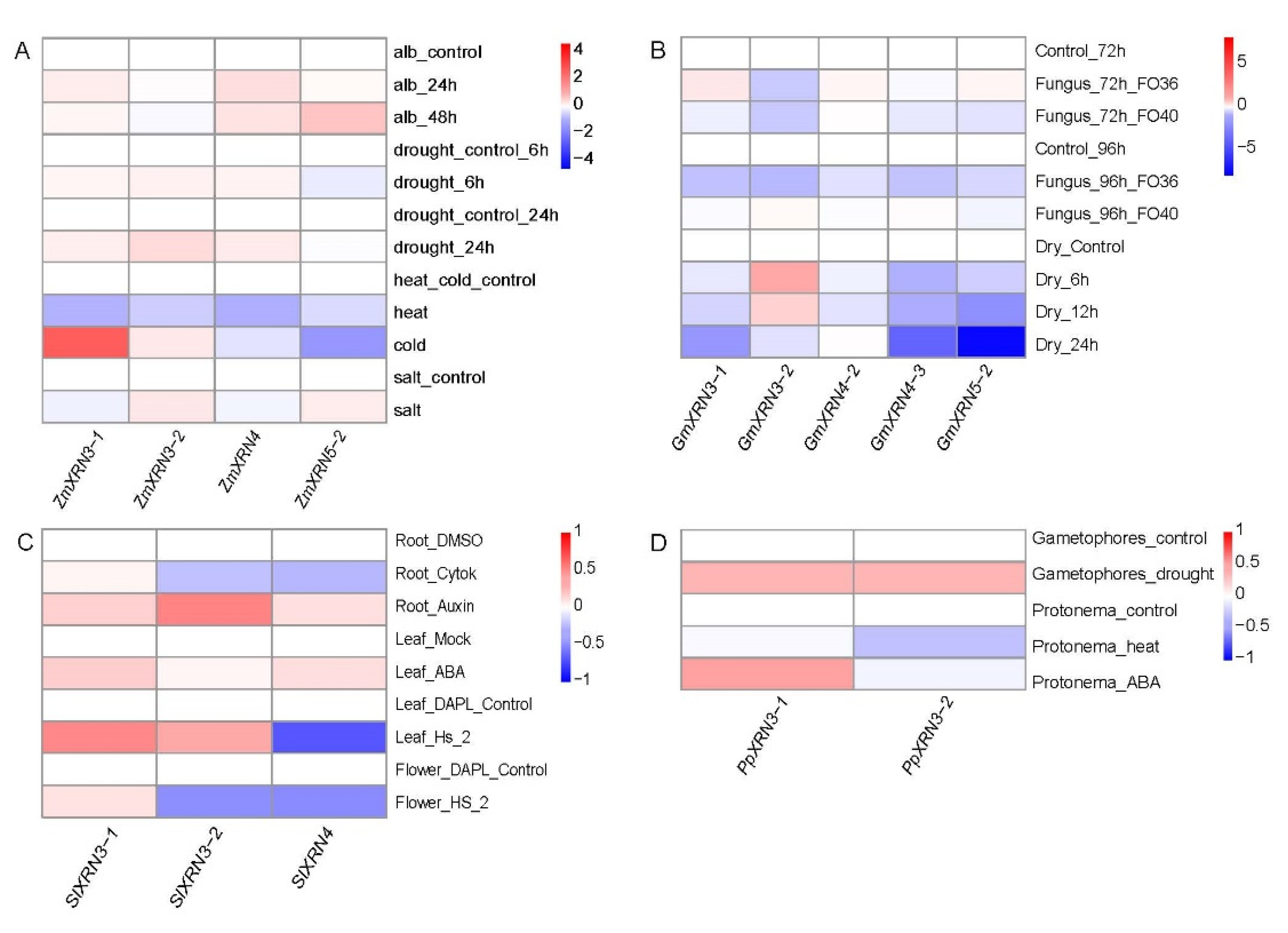
3.5. The Diverse Response of XRNs under Various Environment Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Nagarajan, V.K.; Jones, C.I.; Newbury, S.F.; Green, P.J. XRN 5′→3′ Exoribonucleases: Structure, Mechanisms and Functions. Biochim. Biophys. Acta (BBA)-Gene Regul. Mech. 2013, 1829, 590–603. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.H.; Xiang, S.; Tong, L. Structures of 5′–3′ Exoribonucleases. In The Enzymes; Elsevier: Amsterdam, The Netherlands, 2012; Volume 31, pp. 115–129. ISBN 978-0-12-404740-2. [Google Scholar]
- Langeberg, C.J.; Welch, W.R.W.; McGuire, J.V.; Ashby, A.; Jackson, A.D.; Chapman, E.G. Biochemical Characterization of Yeast Xrn1. Biochemistry 2020, 59, 1493–1507. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.; Cooper-Morgan, A.; Jiao, X.; Kiledjian, M.; Manley, J.L.; Tong, L. Structure and Function of the 5′→3′ Exoribonuclease Rat1 and Its Activating Partner Rai1. Nature 2009, 458, 784–788. [Google Scholar] [CrossRef] [Green Version]
- Pelechano, V.; Wei, W.; Steinmetz, L.M. Widespread Co-Translational RNA Decay Reveals Ribosome Dynamics. Cell 2015, 161, 1400–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kastenmayer, J.P.; Green, P.J. Novel Features of the XRN-Family in Arabidopsis: Evidence That AtXRN4, One of Several Orthologs of Nuclear Xrn2p/Rat1p, Functions in the Cytoplasm. Proc. Natl. Acad. Sci. USA 2000, 97, 13985–13990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hicks, G.R.; Raikhel, N.V. Protein import into the nucleus: An integrated view. Annu. Rev. Cell Dev. Biol. 1995, 11, 155–188. [Google Scholar] [CrossRef] [PubMed]
- Zakrzewska-Placzek, M.; Souret, F.F.; Sobczyk, G.J.; Green, P.J.; Kufel, J. Arabidopsis Thaliana XRN2 Is Required for Primary Cleavage in the Pre-Ribosomal RNA. Nucleic Acids Res. 2010, 38, 4487–4502. [Google Scholar] [CrossRef] [Green Version]
- Merret, R.; Nagarajan, V.K.; Carpentier, M.-C.; Park, S.; Favory, J.-J.; Descombin, J.; Picart, C.; Charng, Y.; Green, P.J.; Deragon, J.-M.; et al. Heat-Induced Ribosome Pausing Triggers MRNA Co-Translational Decay in Arabidopsis Thaliana. Nucleic Acids Res. 2015, 43, 4121–4132. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Willmann, M.R.; Anderson, S.J.; Gregory, B.D. Genome-Wide Mapping of Uncapped and Cleaved Transcripts Reveals a Role for the Nuclear MRNA Cap-Binding Complex in Cotranslational RNA Decay in Arabidopsis. Plant Cell 2016, 28, 2385–2397. [Google Scholar] [CrossRef]
- Carpentier, M.-C.; Deragon, J.-M.; Jean, V.; Be, S.H.V.; Bousquet-Antonelli, C.; Merret, R. Monitoring of XRN4 Targets Reveals the Importance of Cotranslational Decay during Arabidopsis Development. Plant Physiol. 2020, 184, 1251–1262. [Google Scholar] [CrossRef]
- Krzyszton, M.; Zakrzewska-Placzek, M.; Kwasnik, A.; Dojer, N.; Karlowski, W.; Kufel, J. Defective XRN 3-mediated Transcription Termination in Arabidopsis Affects the Expression of Protein-coding Genes. Plant J. 2018, 93, 1017–1031. [Google Scholar] [CrossRef] [Green Version]
- Kurihara, Y. Activity and Roles of Arabidopsis Thaliana XRN Family Exoribonucleases in Noncoding RNA Pathways. J. Plant Res. 2017, 130, 25–31. [Google Scholar] [CrossRef]
- Zhang, X.; Zhu, Y.; Liu, X.; Hong, X.; Xu, Y.; Zhu, P.; Shen, Y.; Wu, H.; Ji, Y.; Wen, X.; et al. Suppression of Endogenous Gene Silencing by Bidirectional Cytoplasmic RNA Decay in Arabidopsis. Science 2015, 348, 120–123. [Google Scholar] [CrossRef]
- Souret, F.F.; Kastenmayer, J.P.; Green, P.J. AtXRN4 Degrades MRNA in Arabidopsis and Its Substrates Include Selected MiRNA Targets. Mol. Cell 2004, 15, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Gazzani, S. A Link between MRNA Turnover and RNA Interference in Arabidopsis. Science 2004, 306, 1046–1048. [Google Scholar] [CrossRef] [Green Version]
- Gregory, B.D.; O’Malley, R.C.; Lister, R.; Urich, M.A.; Tonti-Filippini, J.; Chen, H.; Millar, A.H.; Ecker, J.R. A Link between RNA Metabolism and Silencing Affecting Arabidopsis Development. Dev. Cell 2008, 14, 854–866. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Gao, W.; Wu, S.; Lu, L.; Chen, Y.; Guo, J.; Men, S.; Zhang, X. AtXRN4 Affects the Turnover of Chosen MiRNA*s in Arabidopsis. Plants 2020, 9, 362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurihara, Y.; Schmitz, R.J.; Nery, J.R.; Schultz, M.D.; Okubo-Kurihara, E.; Morosawa, T.; Tanaka, M.; Toyoda, T.; Seki, M.; Ecker, J.R. Surveillance of 3′ Noncoding Transcripts Requires FIERY1 and XRN3 in Arabidopsis. G3 Genes Genomes Genet. 2012, 2, 487–498. [Google Scholar] [CrossRef] [Green Version]
- Gy, I.; Gasciolli, V.; Lauressergues, D.; Morel, J.-B.; Gombert, J.; Proux, F.; Proux, C.; Vaucheret, H.; Mallory, A.C. Arabidopsis FIERY1, XRN2, and XRN3 Are Endogenous RNA Silencing Suppressors. Plant Cell 2007, 19, 3451–3461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwasnik, A.; Wang, V.Y.-F.; Krzyszton, M.; Gozdek, A.; Zakrzewska-Placzek, M.; Stepniak, K.; Poznanski, J.; Tong, L.; Kufel, J. Arabidopsis DXO1 Links RNA Turnover and Chloroplast Function Independently of Its Enzymatic Activity. Nucleic Acids Res. 2019, 47, 4751–4764. [Google Scholar] [CrossRef]
- Yu, X.; Willmann, M.R.; Vandivier, L.E.; Trefely, S.; Kramer, M.C.; Shapiro, J.; Guo, R.; Lyons, E.; Snyder, N.W.; Gregory, B.D. Messenger RNA 5′ NAD+ Capping Is a Dynamic Regulatory Epitranscriptome Mark That Is Required for Proper Response to Abscisic Acid in Arabidopsis. Dev. Cell 2021, 56, 125–140. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Yang, J.; Grudzien-Nogalska, E.; Kiledjian, M. Xrn1 Is a DeNADding Enzyme Modulating Mitochondrial NAD Levels. Mol. Biol. 2021. [Google Scholar] [CrossRef]
- Roman, G.; Lubarsky, B.; Kieber, J.J.; Rothenberg, M.; Ecker, J.R. Genetic analysis of ethylene signal transduction in Arabidopsis thaliana: Five novel mutant loci integrated into a stress response pathway. Genetics 1995, 139, 1393–1409. [Google Scholar] [CrossRef]
- Wawer, I.; Golisz, A.; Sulkowska, A.; Kawa, D.; Kulik, A.; Kufel, J. MRNA Decapping and 5′-3′ Decay Contribute to the Regulation of ABA Signaling in Arabidopsis Thaliana. Front. Plant Sci. 2018, 9, 312. [Google Scholar] [CrossRef]
- Merret, R.; Descombin, J.; Juan, Y.; Favory, J.-J.; Carpentier, M.-C.; Chaparro, C.; Charng, Y.; Deragon, J.-M.; Bousquet-Antonelli, C. XRN4 and LARP1 Are Required for a Heat-Triggered MRNA Decay Pathway Involved in Plant Acclimation and Survival during Thermal Stress. Cell Rep. 2013, 5, 1279–1293. [Google Scholar] [CrossRef] [Green Version]
- Nagarajan, V.K.; Kukulich, P.M.; von Hagel, B.; Green, P.J. RNA Degradomes Reveal Substrates and Importance for Dark and Nitrogen Stress Responses of Arabidopsis XRN4. Nucleic Acids Res. 2019, 47, 9216–9230. [Google Scholar] [CrossRef] [Green Version]
- Stecher, G.; Tamura, K.; Kumar, S. Molecular Evolutionary Genetics Analysis (MEGA) for MacOS. Mol. Biol. Evol. 2020, 37, 1237–1239. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Lescot, M. PlantCARE, a Database of Plant Cis-Acting Regulatory Elements and a Portal to Tools for in Silico Analysis of Promoter Sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Perroud, P.; Haas, F.B.; Hiss, M.; Ullrich, K.K.; Alboresi, A.; Amirebrahimi, M.; Barry, K.; Bassi, R.; Bonhomme, S.; Chen, H.; et al. The Physcomitrella Patens Gene Atlas Project: Large-scale RNA -seq Based Expression Data. Plant J. 2018, 95, 168–182. [Google Scholar] [CrossRef] [Green Version]
- Zouine, M.; Maza, E.; Djari, A.; Lauvernier, M.; Frasse, P.; Smouni, A.; Pirrello, J.; Bouzayen, M. TomExpress, a Unified Tomato RNA-Seq Platform for Visualization of Expression Data, Clustering and Correlation Networks. Plant J. 2017, 92, 727–735. [Google Scholar] [CrossRef] [Green Version]
- Machado, F.B.; Moharana, K.C.; Almeida-Silva, F.; Gazara, R.K.; Pedrosa-Silva, F.; Coelho, F.S.; Grativol, C.; Venancio, T.M. Systematic Analysis of 1298 RNA-Seq Samples and Construction of a Comprehensive Soybean (Glycine Max) Expression Atlas. Plant J. 2020, 103, 1894–1909. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, Y.; Oono, Y.; Wakimoto, H.; Ogata, J.; Kanamori, H.; Sasaki, H.; Mori, S.; Matsumoto, T.; Itoh, T. TENOR: Database for Comprehensive MRNA-Seq Experiments in Rice. Plant Cell Physiol. 2016, 57, e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoopes, G.M.; Hamilton, J.P.; Wood, J.C.; Esteban, E.; Pasha, A.; Vaillancourt, B.; Provart, N.J.; Buell, C.R. An Updated Gene Atlas for Maize Reveals Organ-specific and Stress-induced Genes. Plant J. 2019, 97, 1154–1167. [Google Scholar] [CrossRef] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | Exon Number | Gene Length (bp) | CDS Length (bp) | Protein Length (aa) | pI | MW(kDa) | |
|---|---|---|---|---|---|---|---|---|
| 1 | LOC_Os01g65220 | OsXRN3 | 23 | 7877 | 3207 | 1068 | 6.81 | 120.83 |
| 2 | GRMZM2G064868 | ZmXRN3-1 | 9 | 13,437 | 3240 | 1079 | 6.94 | 122.50 |
| 3 | GRMZM2G121404 | ZmXRN3-2 | 14 | 6709 | 3201 | 1066 | 6.83 | 120.70 |
| 4 | Glyma.14G218000 | GmXRN3-1 | 23 | 11,147 | 3246 | 1081 | 6.85 | 123.09 |
| 5 | Glyma.17G257100 | GmXRN3-2 | 23 | 11,870 | 3210 | 1069 | 7.02 | 121.71 |
| 6 | Solyc04g081280.3 | SlXRN3-1 | 23 | 13,091 | 3330 | 1109 | 5.97 | 126.46 |
| 7 | Solyc12g089280.2 | SlXRN3-2 | 23 | 20,707 | 3351 | 1116 | 8.85 | 126.98 |
| 8 | AT1G75660 | AtXRN3 | 23 | 6985 | 3063 | 1020 | 6.53 | 116.83 |
| 9 | AT5G42540 | AtXRN2 | 22 | 7481 | 3072 | 1023 | 8.99 | 117.69 |
| 10 | Pp3c14_10280 | PpXRN3-1 | 23 | 10,353 | 3000 | 999 | 6.38 | 112.98 |
| 11 | Pp3c24_2160 | PpXRN3-2 | 14 | 7360 | 3357 | 1118 | 6.24 | 126.21 |
| 12 | GRMZM2G099630 | ZmXRN4 | 22 | 15,261 | 2973 | 990 | 7.39 | 113.31 |
| 13 | LOC_Os03g58060 | OsXRN4 | 22 | 11,089 | 2967 | 988 | 7.88 | 112.64 |
| 14 | AT1G54490 | AtXRN4 | 22 | 6808 | 2844 | 947 | 6.37 | 107.78 |
| 15 | Solyc04g049010.3 | SlXRN4 | 23 | 41,681 | 2937 | 978 | 7.09 | 111.57 |
| 16 | Glyma.20G228700 | GmXRN4-1 | 22 | 10,058 | 2799 | 932 | 6.46 | 107.14 |
| 17 | Glyma.03G245000 | GmXRN4-2 | 22 | 12,844 | 2886 | 961 | 6.37 | 109.90 |
| 18 | Glyma.19G242400 | GmXRN4-3 | 22 | 12,523 | 2877 | 958 | 6.48 | 109.94 |
| 19 | GRMZM2G458401 | ZmXRN5-1 | 23 | 6136 | 2682 | 893 | 7.77 | 101.60 |
| 20 | GRMZM2G046755 | ZmXRN5-2 | 21 | 5370 | 2388 | 795 | 5.78 | 90.67 |
| 21 | LOC_Os02g28074 | OsXRN5 | 21 | 16,540 | 2469 | 822 | 6.17 | 93.70 |
| 22 | Glyma.11G180000 | GmXRN5-1 | 15 | 5039 | 1659 | 552 | 8.68 | 63.20 |
| 23 | Glyma.12G093500 | GmXRN5-2 | 9 | 2848 | 1242 | 413 | 8.45 | 46.62 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, W.; Li, Y.; Niu, Y.; Wu, Y.; Bao, Y.; Yu, X. Global Characterization of XRN 5′-3′ Exoribonucleases and Their Responses to Environmental Stresses in Plants. Diversity 2021, 13, 612. https://doi.org/10.3390/d13120612
Song W, Li Y, Niu Y, Wu Y, Bao Y, Yu X. Global Characterization of XRN 5′-3′ Exoribonucleases and Their Responses to Environmental Stresses in Plants. Diversity. 2021; 13(12):612. https://doi.org/10.3390/d13120612
Chicago/Turabian StyleSong, Weimeng, Yanjie Li, Yue Niu, You Wu, Yan Bao, and Xiang Yu. 2021. "Global Characterization of XRN 5′-3′ Exoribonucleases and Their Responses to Environmental Stresses in Plants" Diversity 13, no. 12: 612. https://doi.org/10.3390/d13120612
APA StyleSong, W., Li, Y., Niu, Y., Wu, Y., Bao, Y., & Yu, X. (2021). Global Characterization of XRN 5′-3′ Exoribonucleases and Their Responses to Environmental Stresses in Plants. Diversity, 13(12), 612. https://doi.org/10.3390/d13120612