
Moisture and Salinity Drive the Vegetation Composition of Wadi Hargan, Riyadh, Saudi Arabia

 ,
,  ,
, 
 and
and 
Abstract
1. Introduction
2. Materials and Methods
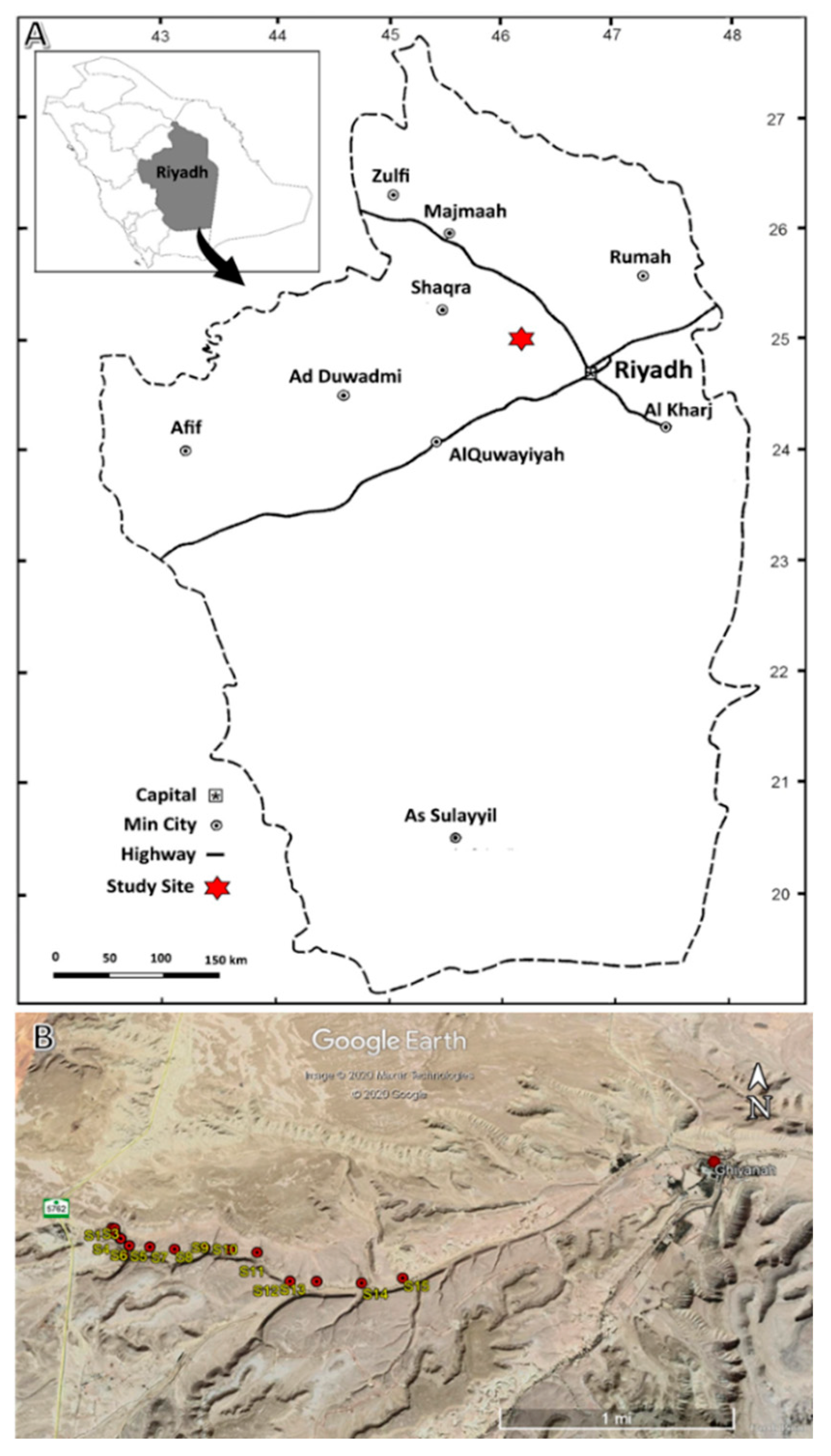
2.1. The Study Area
2.2. Vegetation Analysis
2.3. Soil Analysis
2.4. Data Analysis
3. Results
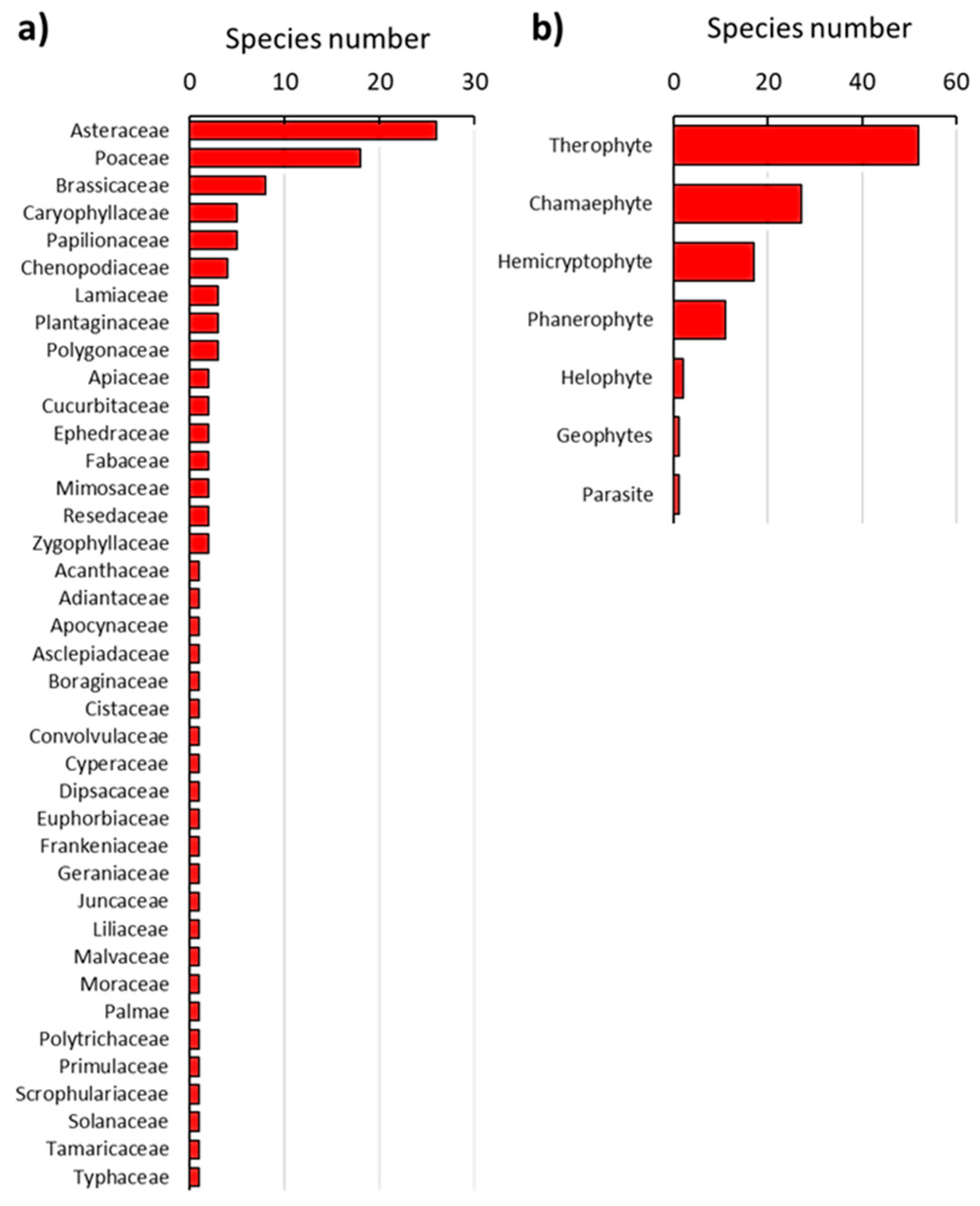
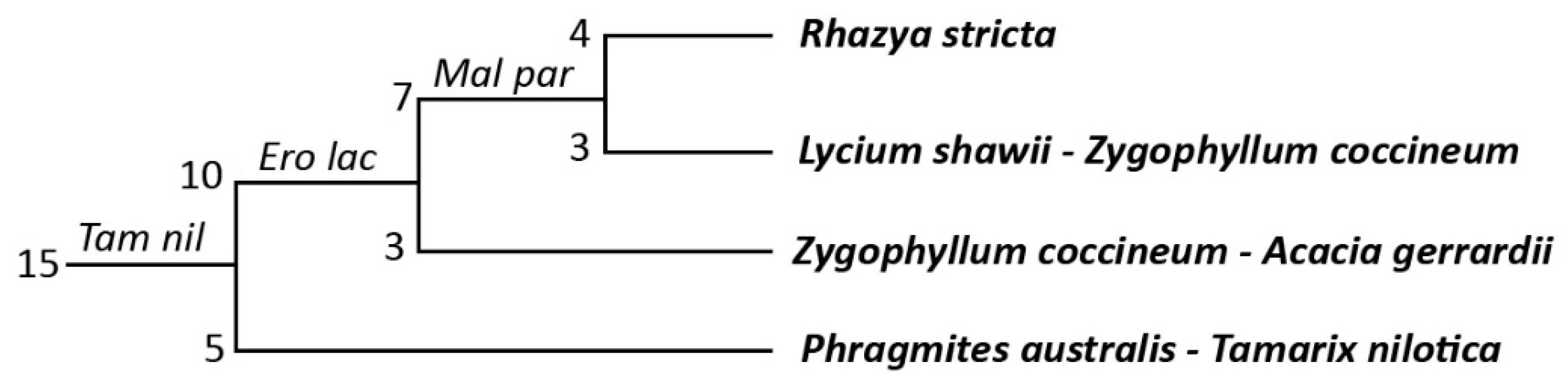
3.1. Vegetation Composition
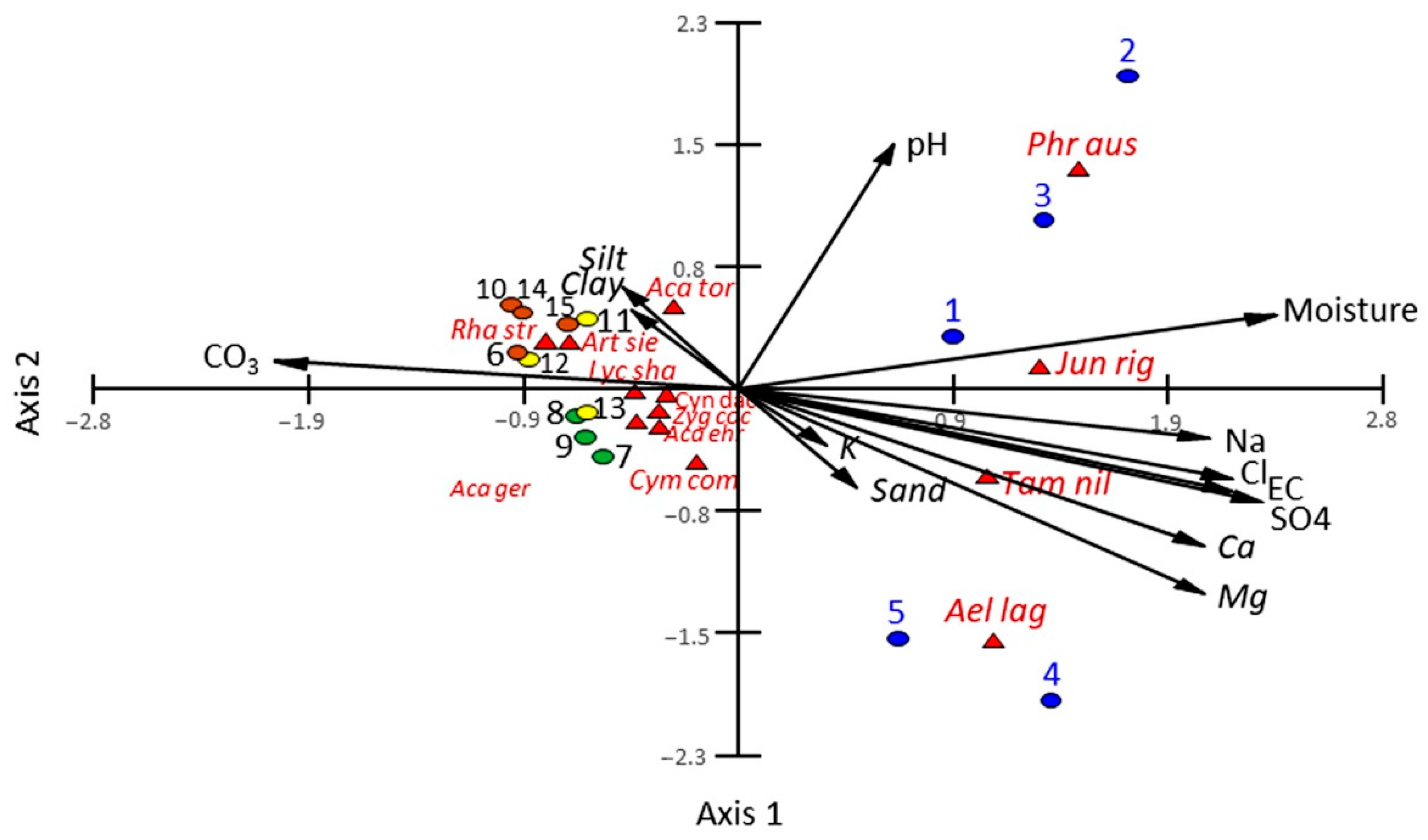
3.2. Vegetation-Soil Relationship
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Al-Obaid, S.; Samraoui, B.; Thomas, J.; El-Serehy, H.A.; Alfarhan, A.H.; Schneider, W.; O’connell, M. An overview of wetlands of Saudi Arabia: Values, threats, and perspectives. Ambio 2017, 46, 98–108. [Google Scholar] [CrossRef]
- Hunter, M.L., Jr.; Acuña, V.; Bauer, D.M.; Bell, K.P.; Calhoun, A.J.; Felipe-Lucia, M.R.; Fitzsimons, J.A.; González, E.; Kinnison, M.; Lindenmayer, D. Conserving small natural features with large ecological roles: A synthetic overview. Biol. Conserv. 2017, 211, 88–95. [Google Scholar] [CrossRef]
- Springer, K.B.; Manker, C.R.; Pigati, J.S. Dynamic response of desert wetlands to abrupt climate change. Proc. Natl. Acad. Sci. USA 2015, 112, 14522–14526. [Google Scholar] [CrossRef] [PubMed]
- Patten, D.T.; Rouse, L.; Stromberg, J.C. Isolated spring wetlands in the Great Basin and Mojave Deserts, USA: Potential response of vegetation to groundwater withdrawal. Environ. Manag. 2008, 41, 398–413. [Google Scholar] [CrossRef]
- Villa, J.A.; Bernal, B. Carbon sequestration in wetlands, from science to practice: An overview of the biogeochemical process, measurement methods, and policy framework. Ecol. Eng. 2018, 114, 115–128. [Google Scholar] [CrossRef]
- Sutton-Grier, A.E.; Sandifer, P.A. Conservation of wetlands and other coastal ecosystems: A commentary on their value to protect biodiversity, reduce disaster impacts, and promote human health and well-being. Wetlands 2019, 39, 1295–1302. [Google Scholar] [CrossRef]
- Orimoloye, I.R.; Kalumba, A.M.; Mazinyo, S.P.; Nel, W. Geospatial analysis of wetland dynamics: Wetland depletion and biodiversity conservation of Isimangaliso Wetland, South Africa. J. King Saud Univ. Sci. 2020, 32, 90–96. [Google Scholar] [CrossRef]
- McKinstry, M.C.; Hubert, W.A.; Anderson, S.H. Wetland and Riparian Areas of the Intermountain West: Ecology and Management; University of Texas Press: Austin, TX, USA, 2004; Volume 4. [Google Scholar]
- Williams, W.D. Biodiversity in temporary wetlands of dryland regions. Int. Ver. Für Theor. Und Angew. Limnol. Verh. 2000, 27, 141–144. [Google Scholar] [CrossRef]
- Noby, K.; Kingma, S.; Heitkönig, I.M.A.; Bulte, E.H.; Naguib, M. Smelly wetlands in the Sahara: Role of sewage ponds in bird migration across Egypt. In Proceedings of Wias Annual Conference; Wageningen University: Lunteren, The Netherlands, 2020; p. 70. [Google Scholar]
- Tinley, K. Survey of Saudi Arabian Wetlands; IUCN/NCWCD Report; General Intelligence Presidency: Riyadh, Saudi Arabia, 1994. [Google Scholar]
- Schenk, H.J.; Holzapfel, C.; Hamilton, J.G.; Mahall, B.E. Spatial ecology of a small desert shrub on adjacent geological substrates. J. Ecol. 2003, 91, 383–395. [Google Scholar] [CrossRef]
- Zhang, D.-M.; Zhao, W.-Z.; Zhang, G.-F. Soil moisture and salt ionic composition effects on species distribution and diversity in semiarid inland saline habitats, northwestern China. Ecol. Res. 2018, 33, 505–515. [Google Scholar] [CrossRef]
- Peters, D.P.; Yao, J.; Sala, O.E.; Anderson, J.P. Directional climate change and potential reversal of desertification in arid and semiarid ecosystems. Glob. Chang. Biol. 2012, 18, 151–163. [Google Scholar] [CrossRef]
- Deng, L.; Wang, K.; Li, J.; Zhao, G.; Shangguan, Z. Effect of soil moisture and atmospheric humidity on both plant productivity and diversity of native grasslands across the Loess Plateau, China. Ecol. Eng. 2016, 94, 525–531. [Google Scholar] [CrossRef]
- Li, W.-Q.; Xiao-Jing, L.; Khan, M.A.; Gul, B. Relationship between soil characteristics and halophytic vegetation in coastal region of North China. Pak. J. Bot. 2008, 40, 1081–1090. [Google Scholar]
- Xi, H.; Feng, Q.; Zhang, L.; Si, J.; Chang, Z.; Yu, T.; Guo, R. Effects of water and salinity on plant species composition and community succession in Ejina Desert Oasis, northwest China. Environ. Earth Sci. 2016, 75, 138. [Google Scholar] [CrossRef]
- Mętrak, M.; Chachulski, Ł.; Navruzshoev, D.; Pawlikowski, P.; Rojan, E.; Sulwiński, M.; Suska-Malawska, M. Nature's patchwork: How water sources and soil salinity determine the distribution and structure of halophytic plant communities in arid environments of the Eastern Pamir. PLoS ONE 2017, 12, e0174496. [Google Scholar] [CrossRef] [PubMed]
- Chaieb, G.; Abdelly, C.; Michalet, R. Interactive effects of climate and topography on soil salinity and vegetation zonation in North-African continental saline depressions. J. Veg. Sci. 2019, 30, 312–321. [Google Scholar] [CrossRef]
- Pringle, C.M. Threats to US public lands from cumulative hydrologic alterations outside of their boundaries. Ecol. Appl. 2000, 10, 971–989. [Google Scholar] [CrossRef]
- Pringle, C.M. Hydrologic connectivity and the management of biological reserves: A global perspective. Ecol. Appl. 2001, 11, 981–998. [Google Scholar] [CrossRef]
- Al-Amro, A.; El-Sheikh, M.; El-Sheikh, A. Vegetation analysis of some wetland habitats in central region of Saudi Arabia. Appl. Ecol. Environ. Res. 2018, 16, 3255–3269. [Google Scholar] [CrossRef]
- Galal, T.M.; Al-Yasi, H.M.; Fadl, M.A. Vegetation zonation along the desert-wetland ecosystem of Taif Highland, Saudi Arabia. Saudi J. Biol. Sci. 2021, 28, 3374–3383. [Google Scholar] [CrossRef] [PubMed]
- Vincent, P. Saudi Arabia: An Environmental Overview; Taylor & Francis: Leiden, The Netherlands, 2008. [Google Scholar]
- Bonham, C.D. Measurements for Terrestrial Vegetation; John Wiley & Sons: New York, NY, USA, 2013. [Google Scholar]
- Mueller-Dombois, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology; Wiley and Sons: New York, NY, USA, 1974. [Google Scholar]
- Collenette, S. Wildflowers of Saudi Arabia; National Commission for Wildlife Conservation and Development (NCWCD): Riyadh, Saudi Arabia, 1999. [Google Scholar]
- Chaudhary, S.A. Flora of the Kingdom of Saudi Arabia; Ministry of Agriculture and Water: Riyadh, Saudi Arabia, 1999. [Google Scholar]
- Raunkiaer, C. Plant Life Forms; Clarendon Press: Oxford, UK, 1937. [Google Scholar]
- Zohary, M. Geobotanical Foundations of the Middle East; Gustav Fischer Verlag: Stuttgart, Germany, 1973. [Google Scholar]
- Bouyoucos, G.J. Hydrometer method improved for making particle size analyses of soils. Agron. J. 1962, 54, 464–465. [Google Scholar] [CrossRef]
- Rowell, D. Soil Science: Methods and Applications; Longman Group: Essex, UK, 1994. [Google Scholar]
- Pierce, W.C.; Haenisch, E.L.; Sawyer, D.T. Quantitative Analysis; Wiley Toppen: Tokyo, Japan, 1958. [Google Scholar]
- Piper, C.S. Soil and Plant Analysis; Interscience Publishers Inc.: New York, NY, USA, 1947. [Google Scholar]
- Allen, S.E.; Grimshaw, H.; Parkinson, J.A.; Quarmby, C. Chemical Analysis of Ecological Materials; Blackwell Scientific Publications: Oxford, UK, 1974. [Google Scholar]
- Hill, M.O. Decorana—A FORTRAN Program for Detrended Correspondence Analysis and Reciprocal Averaging; Ecology and Systematics; Cornell University: Ithaca, NY, USA, 1979; pp. 14850–14852. [Google Scholar]
- Hill, M.O. Twinspan—A FORTRAN Program for Arranging Multivariate Data in an Ordered Two-Way Table by the Classification of the Individuals and Attributes. Ecology and Systematics; Cornell University: Ithaca, NY, USA, 1979; pp. 14850–14890. [Google Scholar]
- Gauch, H.G., Jr.; Whittaker, R.H. Hierarchical classification of community data. J. Ecol. 1981, 69, 537–558. [Google Scholar] [CrossRef]
- Ter Braak, C.J.; Smilauer, P. CANOCO Reference Manual and Canodraw for Windows User's Guide: Software for Canonical Community Ordination (Version 4.5); Biometris: Wageningen, The Netherlands, 2002; Available online: www.canoco.com (accessed on 1 April 2015).
- Zahran, M.A.; Willis, A.J. The Vegetation of Egypt, 2nd ed.; Springer Science & Business Media: Berlin, Germany, 2009. [Google Scholar]
- Abd El-Gawad, A.M. Ecology and allelopathic control of Brassica tournefortii in reclaimed areas of the Nile Delta, Egypt. Turk. J. Bot. 2014, 38, 347–357. [Google Scholar] [CrossRef]
- El-Shabasy, A.; Kasem, W. Systematic composition, species diversity and plant chorology at Wadi Tashar, Jazan, Saudi Arabia. J. Med. Plants Stud. 2018, 6, 83–88. [Google Scholar]
- Osman, A.K.; Al-Ghamdi, F.; Bawadekji, A. Floristic diversity and vegetation analysis of Wadi Arar: A typical desert Wadi of the Northern Border region of Saudi Arabia. Saudi J. Biol. Sci. 2014, 21, 554–565. [Google Scholar] [CrossRef] [PubMed]
- Abdel Khalik, K.; Al-Gohary, I.; Al-Sodany, Y. Floristic composition and vegetation: Environmental relationships of wadi fatimah, mecca, Saudi Arabia. Arid Land Res. Manag. 2017, 31, 316–334. [Google Scholar] [CrossRef]
- Kasem, T.; Marei, A. Floristic Compositions and its affinities to phytogeographical regions in Wadi Khulab of Jazan, Saudi Arabia. Int. J. Plant Soil Sci. 2017, 16, 1–11. [Google Scholar] [CrossRef]
- El Ghazali, G.E.; Al-Soqeer, A.R.A.; El Tayeb, G.E. Floristic and ecological studies on the plant cover of Wadi Al Rummah, Qassim Region, Saudi Arabia. Int. Res. J. Plant Sci. 2013, 4, 310–318. [Google Scholar]
- Khalik, K.A.; El-Sheikh, M.; El-Aidarous, A. Floristic diversity and vegetation analysis of wadi Al-Noman, Mecca, Saudi Arabia. Turk. J. Bot. 2013, 37, 894–907. [Google Scholar] [CrossRef]
- Clayton, W. Poales. In Flowering Plants of the World; Heywood, V.H., Ed.; Oxford University Press: Oxford, UK, 1978; pp. 285–290. [Google Scholar]
- Jeffrey, C. Compositae. In Flowering Plants of the World; Heywood, V.H., Ed.; Oxford University Press: Oxford, UK, 1978; pp. 263–268. [Google Scholar]
- Al-Hashim, M.H.; Taha, M.M.N.; El-Asmar, H.M. Physiographic characteristics along the Ibex Protectorate: Remote sensing application, Hotet Bani Tamim, Central Saudi Arabia. J. Environ. Earth Sci. 2019, 9, 28–42. [Google Scholar]
- Alshammari, A.; Sharawy, S. Wild plants diversity of the Hema Faid Region (Ha'il Province, Saudi Arabia). Asian J. Plant Sci. 2010, 9, 447–454. [Google Scholar] [CrossRef][Green Version]
- Sharawy, S.M.; Alshammari, A.M. Checklist of poisonous plants and animals in Aja Mountain, Ha’il Region, Saudi Arabia. Aust. J. Basic Appl. Sci. 2009, 3, 2217–2225. [Google Scholar]
- Habibi, K. Group dynamics of the Nubian ibex (Capra ibex nubiana) in the Tuwayiq Canyons, Saudi Arabia. J. Zool. 1997, 241, 791–801. [Google Scholar] [CrossRef]
- Li, S.; Su, P.; Zhang, H.; Zhou, Z.; Xie, T.; Shi, R.; Gou, W. Distribution patterns of desert plant diversity and relationship to soil properties in the Heihe River Basin, China. Ecosphere 2018, 9, e02355. [Google Scholar] [CrossRef]
- Gong, Y.; Lv, G.; Guo, Z.; Chen, Y.; Cao, J. Influence of aridity and salinity on plant nutrients scales up from species to community level in a desert ecosystem. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Zhou, W.-L.; Yang, X.-Q.; Hao, P.; Liu, Q.-W.; Cao, D.-C.; Baribault, T.; Li, J.-W. Plant diversity and its maintenance in Populus euphratica riparian forests in the Ejina Oasis, China. Forest. Stud. China 2010, 12, 55–61. [Google Scholar] [CrossRef]
- Wang, Z. Research on desert water management and desert control. Eur. J. Remote Sens. 2020, 53, 1–13. [Google Scholar] [CrossRef]
- Rohit Katuri, J.; Trifonov, P.; Arye, G. Spatial distribution of salinity and sodicity in arid climate following long term brackish water drip irrigated olive orchard. Water 2019, 11, 2556. [Google Scholar] [CrossRef]
- Arora, S.; Dagar, J.C. Salinity tolerance indicators. In Research Developments in Saline Agriculture; Dagar, J.C., Yadav, R.K., Sharma, P.C., Eds.; Springer: Singapore, 2019; pp. 155–201. [Google Scholar]
- Aref, I.M.; El-Juhany, L.; Hegazy, S.S. Comparison of the growth and biomass production of six acacia species in Riyadh, Saudi Arabia after 4 years of irrigated cultivation. J. Arid Environ. 2003, 54, 783–792. [Google Scholar] [CrossRef]
- El-Sheikh, M.A.; Thomas, J.; Alatar, A.A.; Hegazy, A.K.; Abbady, G.A.; Alfarhan, A.H.; Okla, M.I. Vegetation of Thumamah Nature Park: A managed arid land site in Saudi Arabia. Rend. Lincei. 2013, 24, 349–367. [Google Scholar] [CrossRef]
- Shaltout, K.; Mady, M. Analysis of raudhas vegetation in central Saudi Arabia. J. Arid Environ. 1996, 34, 441–454. [Google Scholar] [CrossRef]
- Sardans, J.; Bartrons, M.; Margalef, O.; Gargallo-Garriga, A.; Janssens, I.A.; Ciais, P.; Obersteiner, M.; Sigurdsson, B.D.; Chen, H.Y.; Peñuelas, J. Plant invasion is associated with higher plant–soil nutrient concentrations in nutrient-poor environments. Glob. Chang. Biol. 2017, 23, 1282–1291. [Google Scholar] [CrossRef] [PubMed]
- Incerti, G.; Cartenì, F.; Cesarano, G.; Sarker, T.C.; El-Gawad, A.; Ahmed, M.; D'Ascoli, R.; Bonanomi, G.; Giannino, F. Faster N release, but not C loss, from leaf litter of invasives compared to native species in Mediterranean ecosystems. Front. Plant Sci. 2018, 9, 534. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Plant Community | ||||
|---|---|---|---|---|---|
| A | B | C | D | ||
| Stand no. | 5 | 3 | 3 | 4 | |
| Species no. | 40 | 21 | 21 | 20 | |
| Simpson index | 0.95 | 0.91 | 0.94 | 0.92 | |
| Shannon-evenness | 0.81 | 0.76 | 0.86 | 0.76 | |
| No. | Plant species | Average of importance value | |||
| 1 | Acacia ehrenbergiana Hayne | 7.20 ± 1.38 * | 21.25 ± 5.88 | 8.16 ± 3.82 | 17.27 ± 3.05 |
| 2 | Acacia gerrardii Benth. | 4.99 ± 1.90 | 30.67 ± 10.25 | 16.49 ± 3.13 | 5.09 ± 2.24 |
| 3 | Acacia tortilis (Forssk.) Hayne | 5.88 ± 2.55 | --- | 12.88 ± 6.02 | 18.65 ± 3.45 |
| 4 | Adiantum capillus-veneris L. | 4.42 ± 2.72 | --- | --- | --- |
| 5 | Aeluropus lagopoides (L.) Thwaites | 22.60 ± 7.78 | --- | --- | --- |
| 6 | Alhagi graecorum Boiss. | 1.34 ± 0.82 | 1.29 ± 0.67 | --- | 1.55 ± 0.68 |
| 7 | Artemisia monosperma Delile | --- | 5.90 ± 1.54 | --- | |
| 8 | Artemisia sieberi Besser | 1.17 ± 0.72 | --- | 6.98 ± 1.73 | 22.50 ± 5.54 |
| 9 | Artemisia pycnocephala (Less.) DC. | 0.69 ± 0.43 | --- | --- | --- |
| 10 | Astragalus spinosus (Forssk.) Muschl. | --- | --- | 6.07 ± 1.69 | 10.05 ± 2.77 |
| 11 | Atriplex leucoclada Boiss. | 1.90 ± 0.81 | --- | --- | --- |
| 12 | Blepharis ciliaris (L.) B.L.Burtt | 0.69 ± 0.43 | --- | --- | 3.26 ± 0.84 |
| 13 | Cenchrus ciliaris L. | 3.97 ± 1.02 | --- | 5.00 ± 2.34 | --- |
| 14 | Cucumis prophetarum L. | 0.56 ± 0.35 | --- | --- | --- |
| 15 | Citrullus colocynthis (L.) Schrad. | --- | --- | 1.90 ± 0.89 | --- |
| 16 | Cynodon dactylon (L.) Pers. | 3.14 ± 1.34 | 12.98 ± 3.368 | 11.00 ± 2.59 | --- |
| 17 | Cymbopogon commutatus (Steud.) Stapf | 2.27 ± 1.40 | 9.80 ± 2.93 | --- | 3.97 ± 1.74 |
| 18 | Ephedra foliata Boiss. ex C.A.Mey. | 0.56 ± 0.35 | --- | --- | --- |
| 19 | Ephedra ciliata Fisch. & C.A.Mey. | --- | 3.44 ± 0.96 | 7.22 ± 3.37 | --- |
| 20 | Fagonia bruguieri DC. | 1.92 ± 0.49 | 1.70 ± 0.89 | --- | 2.19 ± 0.97 |
| 21 | Farsetia aegyptia Turra | 1.26 ± 0.48 | --- | 1.90 ± 0.89 | 1.55 ± 0.68 |
| 22 | Ficus salicifolia Vahl | 2.05 ± 1.26 | --- | --- | --- |
| 23 | Gymnocarpos decandrus Forssk. | 1.84 ± 0.47 | --- | --- | 1.55 ± 0.68 |
| 24 | Haloxylon salicornicum (Moq.) Bunge ex Boiss. | 2.67 ± 1.64 | --- | --- | --- |
| 25 | Helianthemum lippii (L.) Dum. Cours. | 1.28 ± 0.48 | 2.91 ± 0.76 | 7.27 ± 0.30 | 1.81 ± |
| 26 | Hyparrhenia hirta (L.) Stapf | 3.16 ± 1.37 | 3.61 ± 0.97 | 5.61 ± 1.31 | --- |
| 27 | Juncus rigidus Desf. | 22.31 ± 3.60 | --- | --- | --- |
| 28 | Lycium shawii Roem. & Schult. | 5.77 ± 0.97 | 3.91 ± 2.03 | 31.22 ± 8.19 | 22.46 ± 9.88 |
| 29 | Moricandia sinaica (Boiss.) Boiss. | --- | 11.37 ± 3.76 | 1.84 ± 0.86 | 1.45 ± 0.64 |
| 30 | Ochradenus baccatus Delile | 4.82 ± 0.81 | 1.35 ± 0.70 | 11.62 ± 1.49 | 6.00 ± 1.52 |
| 31 | Panicum turgidum Forssk. | 1.32 ± 0.81 | 3.12 ± 1.63 | --- | --- |
| 32 | Pennisetum divisum (Forssk. ex J.F.Gmel.) Henrard | --- | --- | --- | 9.38 ± 4.12 |
| 33 | Pergularia tomentosa L. | 0.56 ± 0.35 | --- | 1.90 ± 0.89 | --- |
| 34 | Phoenix dactylifera L. | 3.25 ± 1.23 | --- | --- | --- |
| 35 | Phragmites australis (Cav.) Trin. ex Steud. | 29.97 ± 9.78 | --- | --- | --- |
| 36 | Polygonum aviculare L. | 0.56 ± 0.35 | --- | --- | --- |
| 37 | Prosopis farcta (Banks & Sol.) J.F.Macbr. | 1.32 ± 0.81 | --- | --- | --- |
| 38 | Pulicaria undulata (L.) Kostel. | 0.91 ± 0.56 | 3.26 ± 0.85 | 2.83 ± 1.32 | 1.36 ± 0.60 |
| 39 | Reseda muricata C.Presl | --- | --- | 4.23 ± 1.01 | --- |
| 40 | Rhanterium epapposum Oliv. | 2.69 ± 1.10 | 2.09 ± 1.09 | --- | --- |
| 41 | Rhazya stricta Decne. | --- | 14.81 ± 3.89 | 21.84 ± 7.19 | 47.34 ± 5.06 |
| 42 | Scirpoides holoschoenus (L.) Soják | 1.18 ± 0.72 | --- | --- | --- |
| 43 | Scrophularia deserti Delile | --- | 2.09 ± 1.09 | --- | --- |
| 44 | Stipagrostis plumosa Munro ex T.Anderson | --- | 3.30 ± 1.72 | --- | --- |
| 45 | Tamarix nilotica (Ehrenb.) Bunge | 27.71 ± 4.93 | --- | --- | --- |
| 46 | Teucrium polium L. | 0.59 ± 0.36 | --- | --- | --- |
| 47 | Typha domingensis Pers. | 5.49 ± 3.37 | --- | --- | --- |
| 48 | Zilla spinosa (L.) Prantl | 0.56 ± 0.35 | 1.70 ± 0.89 | 4.62 ± 1.15 | 4.22 ± 1.09 |
| 49 | Zygophyllum coccineum L. | 15.40 ± 3.46 | 59.41 ± 13.27 | 29.41 ± 0.71 | 18.36 ± 5.63 |
| Parameters | Plant Community | F Value | p Value | |||
|---|---|---|---|---|---|---|
| A | B | C | D | |||
| Clay (%) | 7.01 ± 1.46 *AB | 5.34 ± 0.44 B | 11.39 ± 1.93 A | 6.54 ± 1.14 AB | 3.788 | 0.0213 * |
| Silt (%) | 6.86 ± 1.25 A | 5.66 ± 0.66 A | 9.51 ± 2.17 A | 7.31 ± 1.19 A | 1.273 | 0.3028 |
| Sand (%) | 86.13 ± 2.60 AB | 89.00 ± 0.57 A | 79.10 ± 3.95 B | 86.15 ± 1.86 AB | 2.721 | 0.0633 |
| pH | 8.30 ± 0.10 A | 8.16 ± 0.15 A | 8.45 ± 0.08 A | 8.22 ± 0.12 A | 1.093 | 0.3683 |
| EC (dS m−1) | 2.50 ± 0.15 A | 0.85 ± 0.23 B | 0.42 ± 0.02 BC | 0.30 ± 0.05 C | 52.000 | <0.0001 *** |
| CO3 (%) | 0.69 ± 0.08 B | 1.35 ± 0.18 A | 1.20 ± 0.08 A | 1.43 ± 0.15 A | 6.366 | 0.0020 ** |
| Cl (meq/L) | 4.54 ± 0.74 A | 2.15 ± 0.38 B | 1.38 ± 0.44 B | 0.60 ± 0.07 B | 13.034 | <0.0001 *** |
| SO4 (meq/L) | 23.91 ± 2.65 A | 2.49 ± 0.51 B | 0.69 ± 0.09 B | 1.06 ± 0.21 B | 69.373 | <0.0001 *** |
| Ca (meq/L) | 16.90 ± 2.43 A | 2.79 ± 0.53 B | 1.44 ± 0.20 B | 2.61 ± 0.34 B | 33.846 | <0.0001 *** |
| Mg (meq/L) | 8.15 ± 1.39 A | 2.28 ± 0.73 B | 0.69 ± 0.09 B | 1.51 ± 0.43 B | 17.343 | <0.0001 *** |
| Na (meq/L) | 4.59 ± 0.99 A | 1.13 ± 0.54 B | 0.42 ± 0.16 B | 0.31 ± 0.07 B | 12.500 | <0.0001 *** |
| K (meq/L) | 0.38 ± 0.08 A | 0.40 ± 0.13 A | 0.20 ± 0.03 A | 0.35 ± 0.03 A | 1.288 | 0.2978 |
| Moisture (%) | 18.76 ± 2.70 A | 0.97 ± 0.12 B | 0.65 ± 0.09 B | 0.94 ± 0.14 B | 43.734 | <0.0001 *** |
| Variable | Axis 1 | Axis 2 | Axis 3 |
|---|---|---|---|
| Eigenvalues | 0.631 | 0.369 | 0.288 |
| Percentage | 28.513 | 16.684 | 13.016 |
| Cum. Percentage | 28.513 | 45.197 | 58.213 |
| Species-environmental correlations | 0.999 | 0.987 | 0.994 |
| Clay | −0.190 | 0.200 | 0.111 |
| Silt | −0.203 | 0.257 | −0.153 |
| Sand | 0.207 | −0.241 | 0.028 |
| pH | 0.271 | 0.610 | 0.083 |
| EC | 0.867 | −0.252 | 0.301 |
| CO3 | −0.813 | 0.070 | 0.185 |
| Cl | 0.865 | −0.220 | 0.121 |
| SO4 | 0.917 | −0.277 | −0.002 |
| Ca | 0.814 | −0.384 | −0.099 |
| Mg | 0.815 | −0.505 | 0.019 |
| Na | 0.826 | −0.120 | 0.053 |
| K | 0.154 | −0.137 | 0.354 |
| Moisture | 0.940 | 0.183 | −0.036 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd-ElGawad, A.M.; Assaeed, A.M.; Al-Rowaily, S.L.; Dar, B.M.; Malik, J.A. Moisture and Salinity Drive the Vegetation Composition of Wadi Hargan, Riyadh, Saudi Arabia. Diversity 2021, 13, 587. https://doi.org/10.3390/d13110587
Abd-ElGawad AM, Assaeed AM, Al-Rowaily SL, Dar BM, Malik JA. Moisture and Salinity Drive the Vegetation Composition of Wadi Hargan, Riyadh, Saudi Arabia. Diversity. 2021; 13(11):587. https://doi.org/10.3390/d13110587
Chicago/Turabian StyleAbd-ElGawad, Ahmed M., Abdulaziz M. Assaeed, Saud L. Al-Rowaily, Basharat M. Dar, and Jahangir A. Malik. 2021. "Moisture and Salinity Drive the Vegetation Composition of Wadi Hargan, Riyadh, Saudi Arabia" Diversity 13, no. 11: 587. https://doi.org/10.3390/d13110587
APA StyleAbd-ElGawad, A. M., Assaeed, A. M., Al-Rowaily, S. L., Dar, B. M., & Malik, J. A. (2021). Moisture and Salinity Drive the Vegetation Composition of Wadi Hargan, Riyadh, Saudi Arabia. Diversity, 13(11), 587. https://doi.org/10.3390/d13110587