
Spatial Diversity in Bacterial Communities across Barren and Vegetated, Native and Invasive, Coastal Dune Microhabitats


Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Site
2.2. Soil Sample Collection
2.3. DNA Extraction & Sequencing
2.4. GIS & Statistical Analyses
3. Results
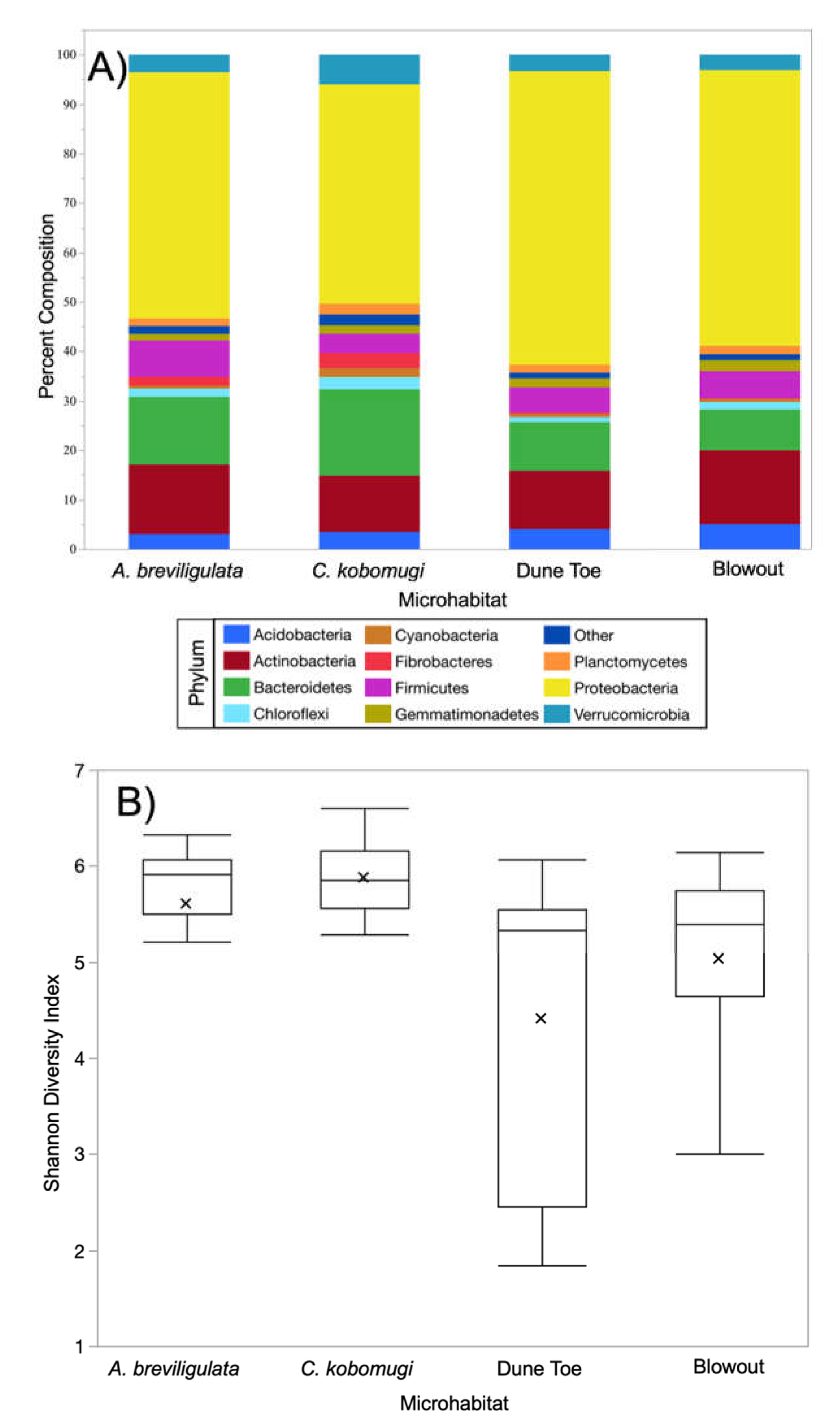
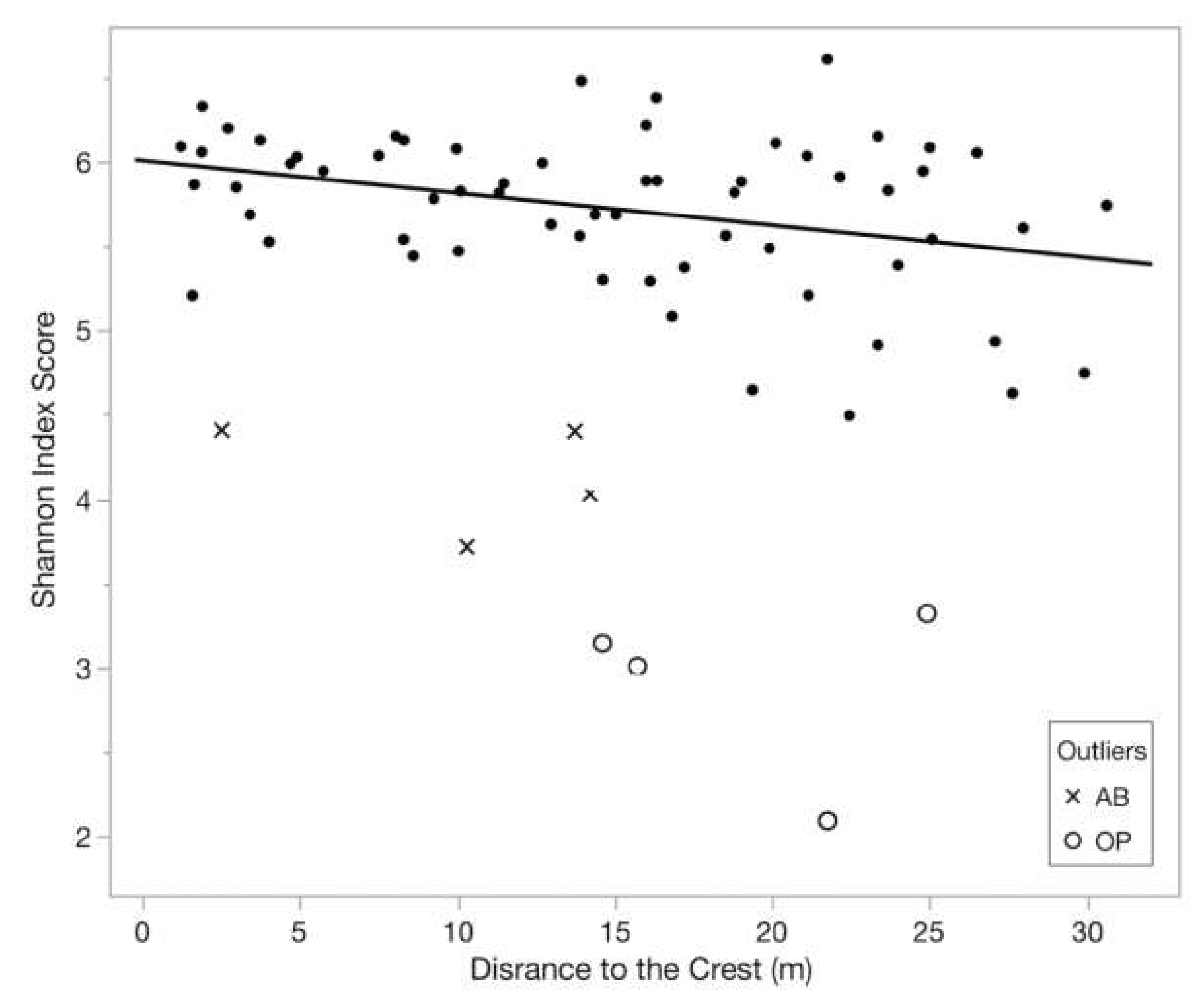
3.1. Bacterial Community Composition & Diversity
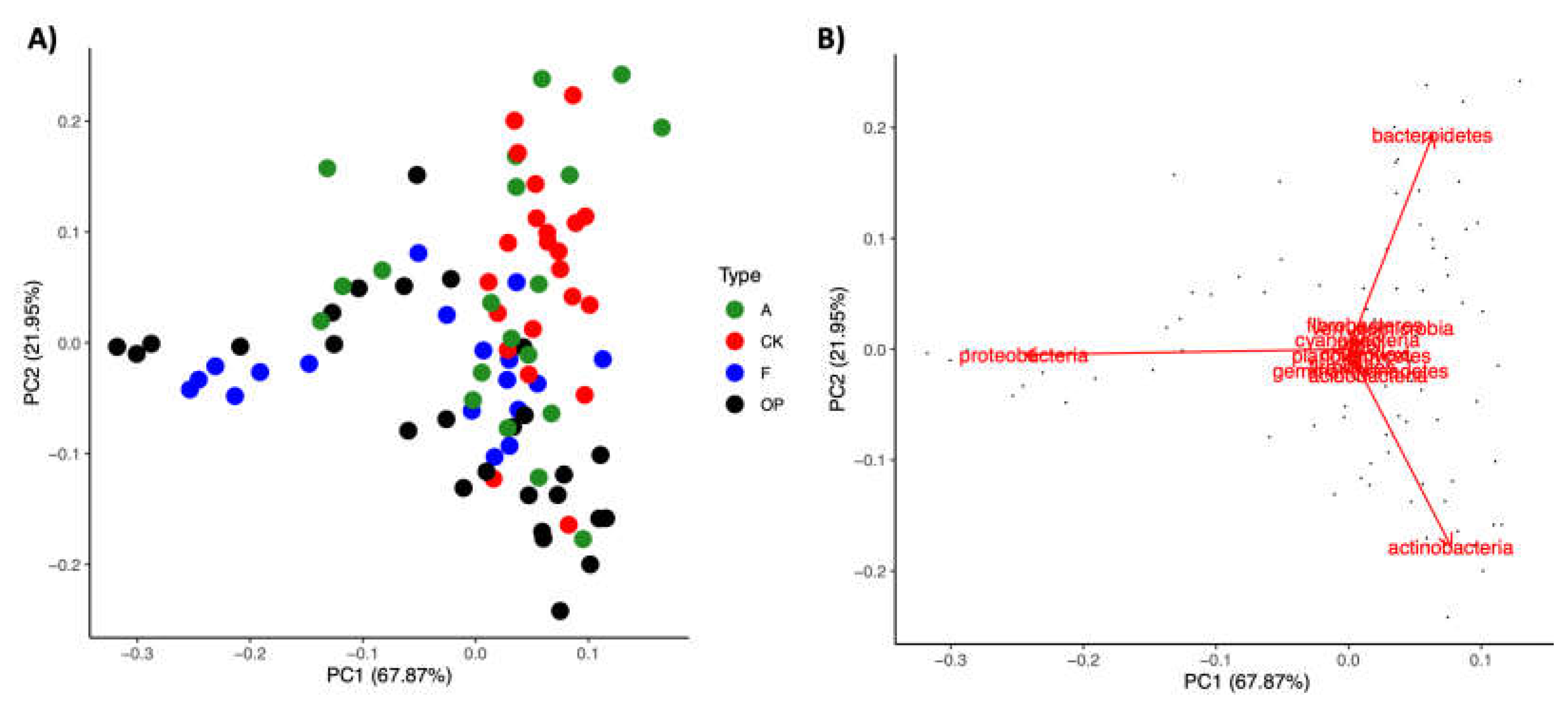
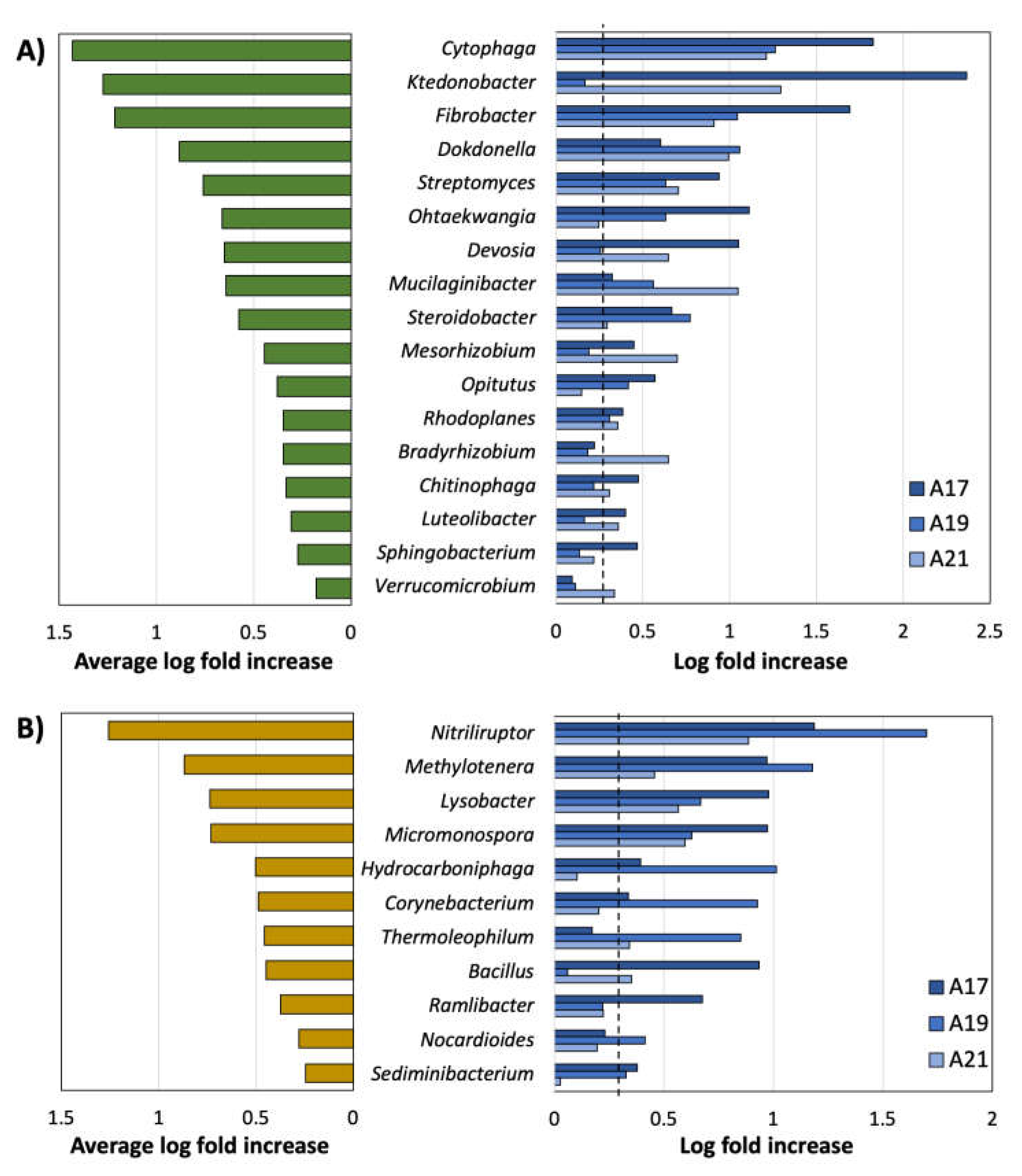
3.2. Phylum-Level Variability in Microbiome Composition
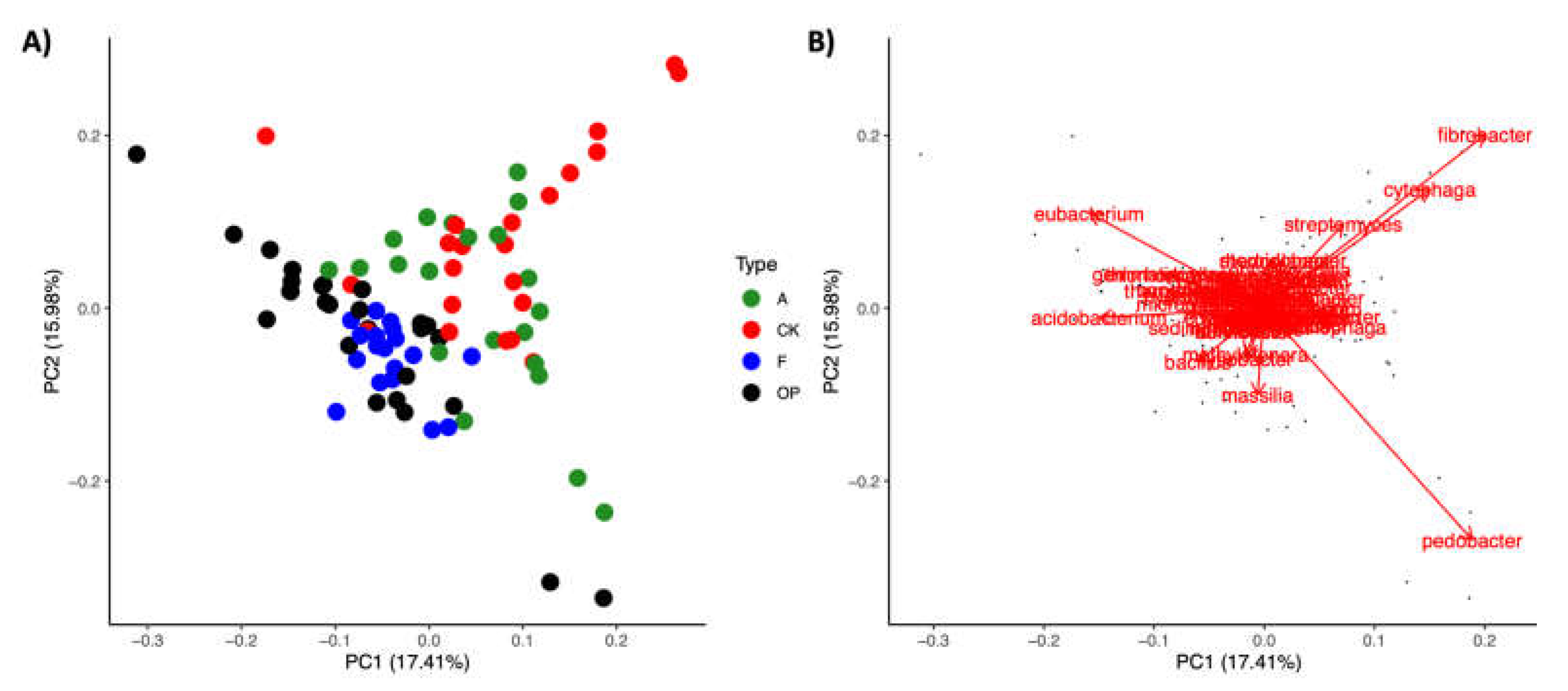
3.3. Genus-Level Variability in Microbiome Composition
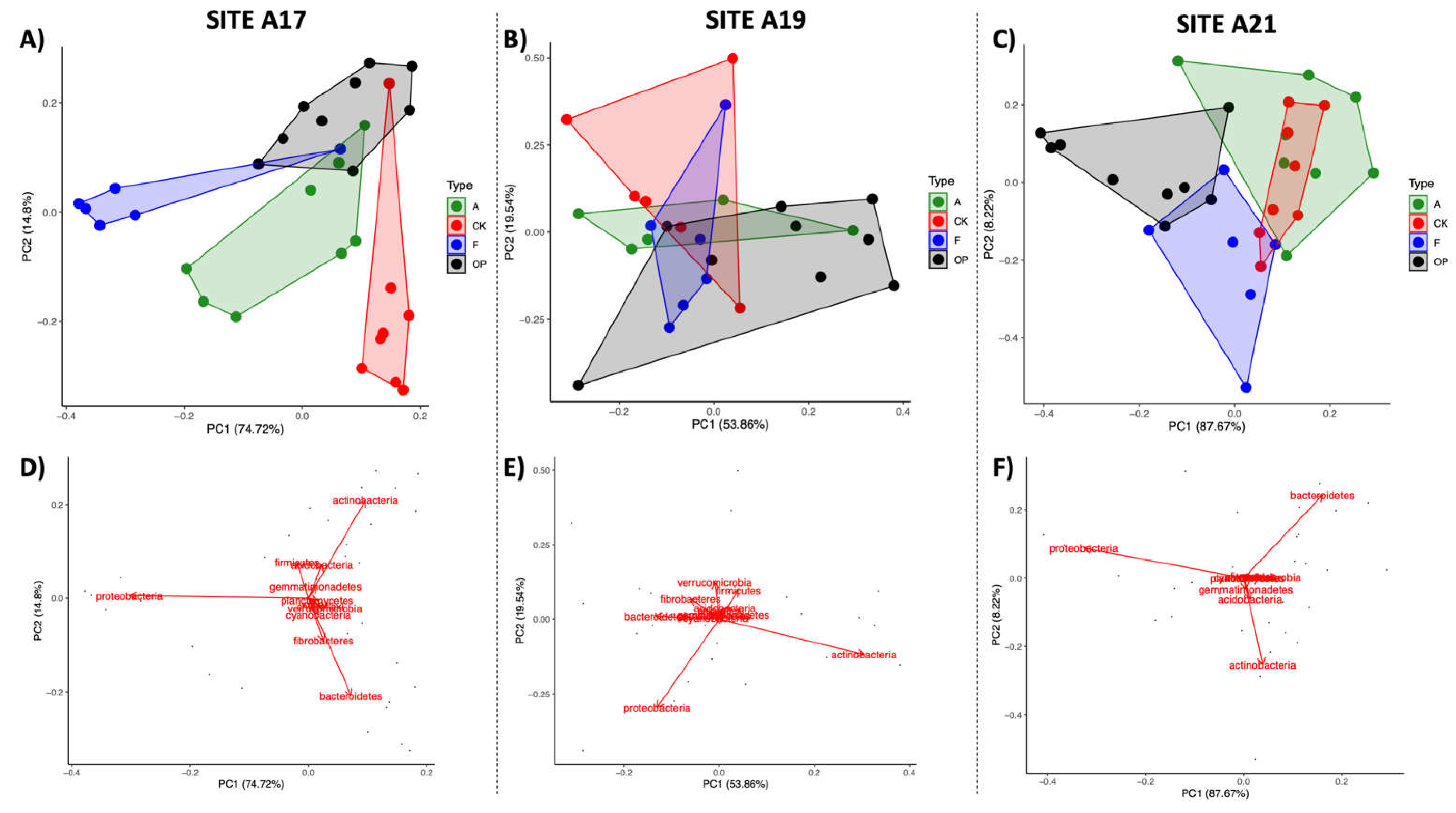
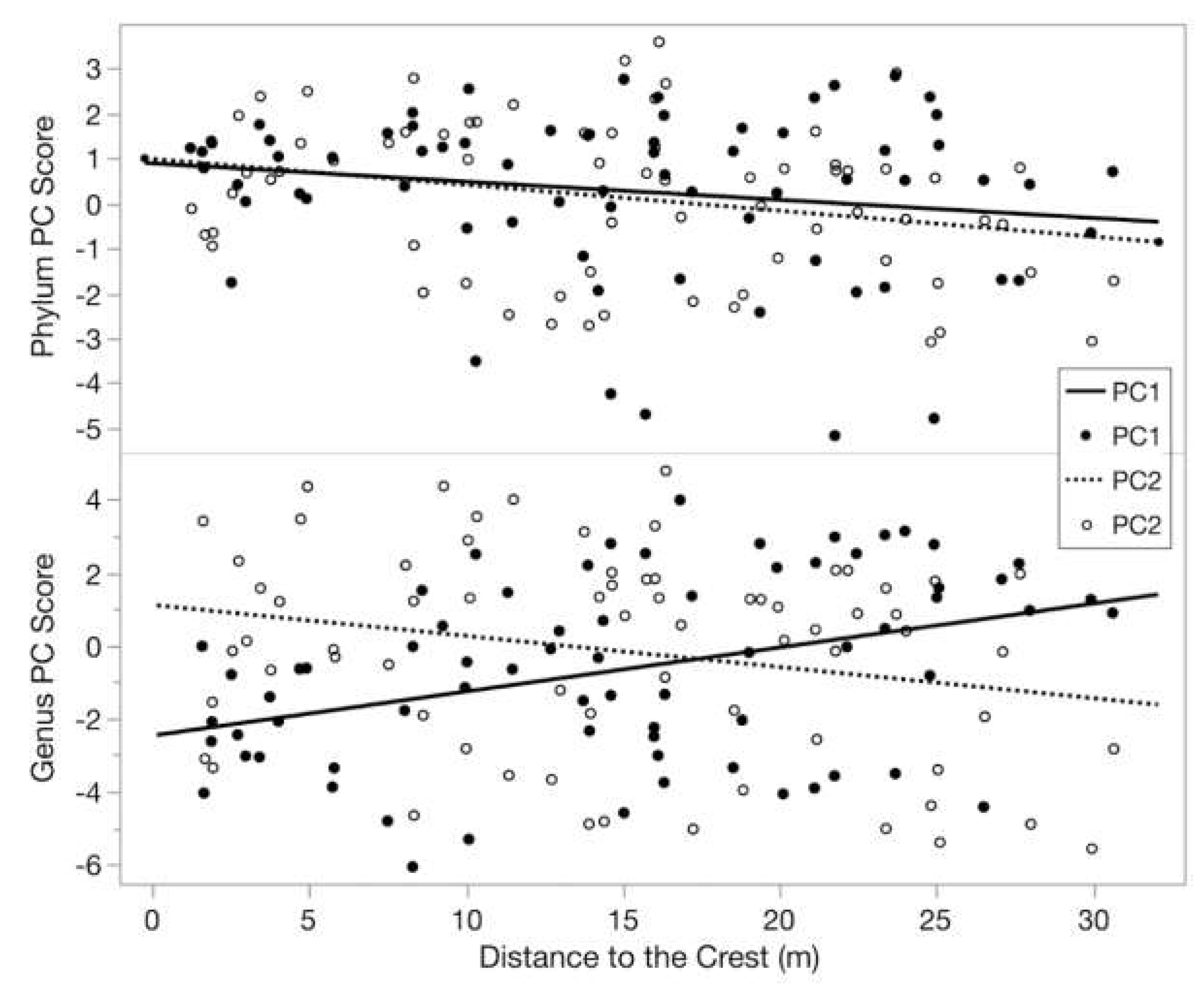
3.4. Spatial Patterns
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meyer, L.; Brinkman, S.; van Kesteren, L.; Leprince-Ringuet, N.; van Boxmeer, F. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change Technical Support. Unit for the Synthesis Report; IPCC: Geneva, Switzerland, 2014; p. 169. [Google Scholar]
- Portner, H.O.; Roberts, D.C.; Masson-Delmotte, V.; Zhai, P.; Tignor, M.; Poloczanska, E.; Mintenbeck, K.; Alegría, A.; Nicolai, M.; Okem, A.; et al. Summary for Policymakers. In IPCC Special Report on the Ocean and Cryosphere in a Changing Climate; IPCC: Geneva, Switzerland, 2019. [Google Scholar]
- Isaac, P.G. Essentials of Nucleic Acid Analysis: A Robust Approach. Ann. Bot. 2009, 104, 5–6. [Google Scholar] [CrossRef] [Green Version]
- Jones, C.; Lawton, J.H.; Shachak, M. Organisms as Ecosystem Engineers. Okios 1994, 69, 373–386. [Google Scholar] [CrossRef]
- Cheplick, G.P. Changes in Plant Abundance on a Coastal Beach Following Two Major Storm Surges. J. Torrey Bot. Soc. 2016, 143, 180–191. [Google Scholar] [CrossRef]
- Charbonneau, B.R.; Wootton, L.S.; Wnek, J.P.; Langley, J.A.; Posner, M.A. A Species Effect on Storm Erosion: Invasive Sedge Stabilized Dunes More than Native Grass during Hurricane Sandy. J. Appl. Ecol. 2017, 54, 1385–1394. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, N.; Nandasena, N.A.K.; Jinadasa, K.B.S.N.; Sasaki, Y.; Tanimoto, K.; Mowjood, M.I.M. Developing Effective Vegetation Bioshield for Tsunami Protection. Civ. Eng. Environ. Syst. 2009, 26, 163–180. [Google Scholar] [CrossRef]
- Zarnetske, P.L.; Hacker, S.D.; Seabloom, E.W.; Ruggiero, P.; Killian, J.R.; Maddux, T.B.; Cox, D. Biophysical Feedback Mediates Effects of Invasive Grasses on Coastal Dune Shape. Ecology 2012, 93, 1439–1450. [Google Scholar] [CrossRef] [PubMed]
- Sigren, J.M.; Figlus, J.; Armitage, A.R. Coastal Sand Dunes and Dune Vegetation: Restoration, Erosion, and Storm Protection. Shore Beach 2014, 82, 5–12. [Google Scholar]
- Forster, S.M.; Nicolson, T.H. Aggregation of Sand from a Maritime Embryo Sand Dune by Microorganisms and Higher Plants. Soil Biol. Biochem. 1981, 13, 199–203. [Google Scholar] [CrossRef]
- Hesp, P.A. A Review of Biological and Geomorphological Processes Involved in the Initiation and Development of Incipient Foredunes. Proc. R. Soc. Edinb. 1989, 96. [Google Scholar] [CrossRef]
- Hacker, S.D.; Jay, K.R.; Cohn, N.; Goldstein, E.B.; Hovenga, P.A.; Itzkin, M.; Moore, L.J.; Mostow, R.S.; Mullins, E.V.; Ruggiero, P. Species-Specific Functional Morphology of Four US Atlantic Coast Dune Grasses: Biogeographic Implications for Dune Shape and Coastal Protection. Diversity 2019, 11, 82. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.T.; Whitman, W.B.; Coleman, D.C.; Chen, T.H.; Chiu, C.Y. Composition of Bacterial Communities in Sand Dunes of Subtropical Coastal Forests. Biol. Fertil. Soils 2014, 50, 809–814. [Google Scholar] [CrossRef]
- Liu, R.; Li, K.; Zhang, H.; Zhu, J.; Joshi, D. Spatial Distribution of Microbial Communities Associated with Dune Landform in the Gurbantunggut Desert, China. J. Microbiol. 2014, 52, 898–907. [Google Scholar] [CrossRef]
- Abdul Majid, S.; Graw, M.F.; Chatziefthimiou, A.D.; Nguyen, H.; Richer, R.; Louge, M.; Sultan, A.A.; Schloss, P.; Hay, A.G. Microbial Characterization of Qatari Barchan Sand Dunes. PLoS ONE 2016, 11, e0161836. [Google Scholar] [CrossRef] [Green Version]
- Shet, S.A.; Garg, S. Prokaryotic Diversity of Tropical Coastal Sand Dunes Ecosystem Using Metagenomics. 3 Biotech. 2021, 11, 252. [Google Scholar] [CrossRef]
- Howes, A. Environmental Factors, Not. Plants, Contribute to Functional Diversity of Soil Bacteria in the Dunes of Lake Michigan; University of Louisville: Louisville, KY, USA, 2017. [Google Scholar]
- Nayak, S.; Behera, S.; Dash, P.K. Potential of Microbial Diversity of Coastal Sand Dunes: Need for Exploration in Odisha Coast of India. Sci. World J. 2019, 2019, 2758501. [Google Scholar] [CrossRef] [Green Version]
- Viles, H.A. Microbial Geomorphology: A Neglected Link between Life and Landscape. Geomorphology 2012, 157–158, 6–16. [Google Scholar] [CrossRef]
- Batten, K.M.; Scow, K.M.; Espeland, E.K. Soil Microbial Community Associated with an Invasive Grass Differentially Impacts Native Plant Performance. Microb. Ecol. 2008, 55, 220–228. [Google Scholar] [CrossRef]
- Chang, E.H.; Chen, C.T.; Chen, T.H.; Chiu, C.Y. Soil Microbial Communities and Activities in Sand Dunes of Subtropical Coastal Forests. Appl. Soil Ecol. 2011, 49, 256–262. [Google Scholar] [CrossRef]
- Bertin, C.; Yang, X.; Weston, L.A. The Role of Root Exudates and Allelochemicals in the Rhizosphere. Plant. Soil 2003, 256, 67–83. [Google Scholar] [CrossRef]
- Huang, X.-F.; Chaparro, J.M.; Reardon, K.F.; Zhang, R.; Shen, Q.; Vivanco, J.M. Rhizosphere Interactions: Root Exudates, Microbes, and Microbial Communities. Botany 2014, 92, 267–275. [Google Scholar] [CrossRef]
- Godinho, A.L.; Bhosle, S. Sand Aggregation by Exopolysaccharide-Producing Microbacterium arborescens––AGSB. Curr. Microbiol. 2009, 58, 616–621. [Google Scholar] [CrossRef]
- Wasserstrom, H.; Kublik, S.; Wasserstrom, R.; Schulz, S.; Schloter, M.; Steinberger, Y. Bacterial Community Composition in Costal Dunes of the Mediterranean along a Gradient from the Sea Shore to the Inland. Sci. Rep. 2017, 7, 40266. [Google Scholar] [CrossRef] [Green Version]
- Soka, G.; Ritchie, M. Arbuscular Mycorrhizal Symbiosis and Ecosystem Processes: Prospects for Future Research in Tropical Soils. Open J. Ecol. 2014, 4, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Kloepper, J.W.; Ryu, C.-M. Rhizosphere Bacteria Help Plants Tolerate Abiotic Stress. Trends Plant. Sci. 2009, 14, 1–4. [Google Scholar] [CrossRef]
- Li, W.; Sheng, H.; Ekawati, D.; Jiang, Y.; Yang, H. Variations in the Compositions of Soil Bacterial and Fungal Communities Due to Microhabitat Effects Induced by Simulated Nitrogen Deposition of a Bamboo Forest in Wetland. Forests 2019, 10, 1098. [Google Scholar] [CrossRef] [Green Version]
- David, A.S.; Seabloom, E.W.; May, G. Plant Host Species and Geographic Distance Affect the Structure of Aboveground Fungal Symbiont Communities, and Environmental Filtering Affects Belowground Communities in a Coastal Dune Ecosystem. Microb. Ecol. 2016, 71, 912–926. [Google Scholar] [CrossRef] [PubMed]
- Titus, J.H.; del Moral, R. The Role of Mycorrhizal Fungi and Microsites in Primary Succession on Mount St. Helens. Am. J. Bot. 1998, 85, 370–375. [Google Scholar] [CrossRef]
- Rajaniemi, T.K.; Allison, V.J. Abiotic Conditions and Plant Cover Differentially Affect Microbial Biomass and Community Composition on Dune Gradients. Soil Biol. Biochem. 2009, 41, 102–109. [Google Scholar] [CrossRef]
- Muñoz Vallés, S.; Cambrollé, J.; Gallego-Fernández, J.B. Effect of Soil Characteristics on Plant Distribution in Coastal Ecosystems of SW Iberian Peninsula Sand Spits. Plant. Ecol. 2015, 216, 1551–1570. [Google Scholar] [CrossRef]
- Gallego-Fernández, J.B.; Martínez, M.L. Environmental Filtering and Plant Functional Types on Mexican Foredunes along the Gulf of Mexico. Écoscience 2011, 18, 52–62. [Google Scholar] [CrossRef]
- Ley, R.E.; Lipson, D.A.; Schmidt, S.K. Microbial Biomass Levels in Barren and Vegetated High Altitude Talus Soils. Soil Sci. Soc. Am. J. 2001, 65, 111–117. [Google Scholar] [CrossRef]
- Li, Z.; Zu, C.; Wang, C.; Yang, J.; Yu, H.; Wu, H. Different Responses of Rhizosphere and Non-Rhizosphere Soil Microbial Communities to Consecutive Piper nigrum L. Monoculture. Sci. Rep. 2016, 6, 35825. [Google Scholar] [CrossRef]
- Poosakkannu, A.; Nissinen, R.; Männistö, M.; Kytöviita, M.-M. Microbial Community Composition but Not Diversity Changes along Succession in Arctic Sand Dunes: Deschampsia flexuosa Associated Microbiomes. Environ. Microbiol. 2017, 19, 698–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, J.; Hesp, P.A. Salt Spray Distribution and Its Impact on Vegetation Zonation on Coastal Dunes: A Review. Estuaries Coasts 2020, 43, 1885–1907. [Google Scholar] [CrossRef]
- Wichern, J.; Wichern, F.; Joergensen, R.G. Impact of Salinity on Soil Microbial Communities and the Decomposition of Maize in Acidic Soils. Geoderma 2006, 137, 100–108. [Google Scholar] [CrossRef]
- Yang, J.; Ma, L.; Jiang, H.; Wu, G.; Dong, H. Salinity Shapes Microbial Diversity and Community Structure in Surface Sediments of the Qinghai-Tibetan Lakes. Sci. Rep. 2016, 6, 25078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elbl, J.; Zahora, J. The Comparison of Microbial Activity in Rhizosphere and Non-Rhizosphere Soil Stressed by Drought. Mendel. Net. 2014, 7, 234–240. [Google Scholar]
- Charbonneau, B.R.; Nicoletta, R.; Wootton, L.S. A Decade of Expansion of the Invasive Plant Carex kobomugi in a Coastal Foredune System. Biol. Invasions 2020, 22, 2099–2112. [Google Scholar] [CrossRef]
- Fei, S.; Phillips, J.; Shouse, M. Biogeomorphic Impacts of Invasive Species. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 69–87. [Google Scholar] [CrossRef] [Green Version]
- Gallego-Fernández, J.B.; Martínez, M.L.; García-Franco, J.G.; Zunzunegui, M. Multiple Seed Dispersal Modes of an Invasive Plant Species on Coastal Dunes. Biol. Invasions 2021, 23, 111–127. [Google Scholar] [CrossRef]
- Kourtev, P.S.; Ehrenfeld, J.G.; Haggblom, M. Exotic Plant Species Alter the Microbial Community Structure and Function in the Soil. Ecology 2002, 83, 3152–3166. [Google Scholar] [CrossRef]
- Batten, K.M.; Scow, K.M.; Davies, K.F.; Harrison, S.P. Two Invasive Plants Alter Soil Microbial Community Composition in Serpentine Grasslands. Biol. Invasions 2006, 8, 217–230. [Google Scholar] [CrossRef]
- Leege, L.M.; Murphy, P.G. Ecological Effects of the Non-Native Pinus nigra on Sand Dune Communities. Can. J. Bot. 2001, 79, 429–437. [Google Scholar] [CrossRef]
- Stefanowicz, A.M.; Zubek, S.; Stanek, M.; Grześ, I.M.; Rożej-Pabijan, E.; Błaszkowski, J.; Woch, M.W. Invasion of Rosa rugosa Induced Changes in Soil Nutrients and Microbial Communities of Coastal Sand Dunes. Sci. Total Environ. 2019, 677, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Jurkšienė, G.; Janušauskaitė, D.; Baliuckas, V. Microbial Community Analysis of Native Pinus sylvestris L. and Alien Pinus mugo L. on Dune Sands as Determined by Ecoplates. Forests 2020, 11, 1202. [Google Scholar] [CrossRef]
- Wu, B.; Wang, S.; Wei, M.; Zhou, J.; Jiang, K.; Du, D.; Wang, C. The Invasive Tree Staghorn sumac Affects Soil N2-fixing Bacterial Communities in North China. Plant. Biol. 2019, 21, 951–960. [Google Scholar] [CrossRef] [PubMed]
- Day, N.J.; Antunes, P.M.; Dunfield, K.E. Changes in Arbuscular Mycorrhizal Fungal Communities during Invasion by an Exotic Invasive Plant. Acta Oecologica 2015, 67, 66–74. [Google Scholar] [CrossRef]
- Elgersma, K.J.; Ehrenfeld, J.G.; Yu, S.; Vor, T. Legacy Effects Overwhelm the Short-Term Effects of Exotic Plant Invasion and Restoration on Soil Microbial Community Structure, Enzyme Activities, and Nitrogen Cycling. Oecologia 2011, 167, 733–745. [Google Scholar] [CrossRef] [PubMed]
- Tanner, R.A.; Gange, A.C. The Impact of Two Non-Native Plant Species on Native Flora Performance: Potential Implications for Habitat Restoration. Plant. Ecol. 2013, 214, 423–432. [Google Scholar] [CrossRef]
- Malacrinò, A.; Sadowski, V.A.; Martin, T.K.; Cavichiolli de Oliveira, N.; Brackett, I.J.; Feller, J.D.; Harris, K.J.; Combita Heredia, O.; Vescio, R.; Bennett, A.E. Biological Invasions Alter Environmental Microbiomes: A Meta-Analysis. PLoS ONE 2020, 15, e0240996. [Google Scholar] [CrossRef]
- Corbin, J.D.; D’Antonio, C.M. Gone but Not Forgotten? Invasive Plants’ Legacies on Community and Ecosystem Properties. Invasive Plant Sci. Manag. 2012, 5, 117–124. [Google Scholar] [CrossRef]
- Durán Vinent, O.; Moore, L.J. Barrier Island Bistability Induced by Biophysical Interactions. Nat. Clim. Chang. 2015, 5, 158–162. [Google Scholar] [CrossRef]
- Small, J.A. Carex kobomugi at Island Beach, New Jersey. Ecology 1954, 35, 289–291. [Google Scholar] [CrossRef]
- Wootton, L.S.; Halsey, S.D.; Bevaart, K.; McGough, A.; Ondreicka, J.; Patel, P. When Invasive Species Have Benefits as Well as Costs: Managing Carex kobomugi (Asiatic Sand Sedge) in New Jersey’s Coastal Dunes. Biol. Invasions 2005, 7, 1017–1027. [Google Scholar] [CrossRef]
- Charbonneau, B.R. From The Sand They Rise: Post-Storm Foredune Plant Recolonization And Its Biogeomorphic Implications; University of Pennsylvania: Philadelphia, PA, USA, 2019; pp. 2–19. [Google Scholar]
- Hesp, P. Foredunes and Blowouts: Initiation, Geomorphology and Dynamics. Geomorphology 2002, 48, 245–268. [Google Scholar] [CrossRef]
- Sallenger, A.H.J. Storm Impact Scale for Barrier Islands. J. Coast. Res. 2000, 16, 890–895. [Google Scholar]
- Friese, C.F.; Koske, R.E. The Spatial Dispersion of Spores of Vesicular-Arbuscular Mycorrhizal Fungi in a Sand Dune: Microscale Patterns Associated with the Root Architecture of American Beachgrass. Mycol. Res. 1991, 95, 952–957. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. Vienna, Austria, 2016. Available online: Https://Www.R-Project.Org/ (accessed on 1 August 2021).
- Janssen, P.H. Identifying the Dominant Soil Bacterial Taxa in Libraries of 16S RRNA and 16S RRNA Genes. Appl. Environ. Microbiol. 2006, 72, 1719–1728. [Google Scholar] [CrossRef] [Green Version]
- Thomas, F.; Hehemann, J.-H.; Rebuffet, E.; Czjzek, M.; Michel, G. Environmental and Gut Bacteroidetes: The Food Connection. Front. Microbiol. 2011, 2, 93. [Google Scholar] [CrossRef] [Green Version]
- Larsbrink, J.; McKee, L.S. Bacteroidetes bacteria in the soil: Glycan acquisition, enzyme secretion, and gliding motility. In Advances in Applied Microbiology; Elsevier: Amsterdam, The Netherlands, 2020; Volume 110, pp. 63–98. ISBN 978-0-12-820703-1. [Google Scholar]
- Zhang, B.; Hong, J.; Zhang, Q.; Jin, D.; Gao, C. Contrast in Soil Microbial Metabolic Functional Diversity to Fertilization and Crop Rotation under Rhizosphere and Non-Rhizosphere in the Coal Gangue Landfill Reclamation Area of Loess Hills. PLoS ONE 2020, 15, e0229341. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Acuña, J.J.; Inostroza, N.G.; Mora, M.L.; Radic, S.; Sadowsky, M.J.; Jorquera, M.A. Endophytic Bacterial Communities Associated with Roots and Leaves of Plants Growing in Chilean Extreme Environments. Sci. Rep. 2019, 9, 4950. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Acuña, J.J.; Inostroza, N.G.; Duran, P.; Mora, M.L.; Sadowsky, M.J.; Jorquera, M.A. Niche Differentiation in the Composition, Predicted Function, and Co-Occurrence Networks in Bacterial Communities Associated With Antarctic Vascular Plants. Front. Microbiol. 2020, 11, 1036. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, G.T.; Bates, S.T.; Eilers, K.G.; Lauber, C.L.; Caporaso, J.G.; Walters, W.A.; Knight, R.; Fierer, N. The Under-Recognized Dominance of Verrucomicrobia in Soil Bacterial Communities. Soil Biol. Biochem. 2011, 43, 1450–1455. [Google Scholar] [CrossRef] [Green Version]
- Abdul Rahman, N.; Parks, D.H.; Vanwonterghem, I.; Morrison, M.; Tyson, G.W.; Hugenholtz, P. A Phylogenomic Analysis of the Bacterial Phylum Fibrobacteres. Front. Microbiol. 2016, 6, 1469. [Google Scholar] [CrossRef] [Green Version]
- Ransom-Jones, E.; Jones, D.L.; McCarthy, A.J.; McDonald, J.E. The Fibrobacteres: An Important Phylum of Cellulose-Degrading Bacteria. Microb. Ecol. 2012, 63, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.P.; Bastos, M.S.; Xavier, V.B.; Cassel, E.; Astarita, L.V.; Santarém, E.R. Plant Growth and Resistance Promoted by Streptomyces spp. in Tomato. Plant. Physiol. Biochem. 2017, 118, 479–493. [Google Scholar] [CrossRef]
- Mello, L.D.P.S.; dos Santos, A.C.; dos Santos, R.M.; Kandasamy, S.; Lazarovits, G.; Rigobelo, E.C. Application of the Bacterial Strains Ruminobacter amylophilus, Fibrobacter succinogenes and Enterococcus faecium for Growth Promotion in Maize and Soybean Plants. Aust. J. Crop. Sci. 2020, 14, 2020–2027. [Google Scholar] [CrossRef]
- Xu, L.; Zhang, H.; Xing, Y.-T.; Li, N.; Wang, S.; Sun, J.-Q. Complete Genome Sequence of Sphingobacterium psychroaquaticum Strain SJ-25, an Aerobic Bacterium Capable of Suppressing Fungal Pathogens. Curr. Microbiol. 2020, 77, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Wani, P.A.; Khan, M.S.; Zaidi, A. Chromium-Reducing and Plant Growth-Promoting Mesorhizobium Improves Chickpea Growth in Chromium-Amended Soil. Biotechnol. Lett. 2007, 30, 159–163. [Google Scholar] [CrossRef]
- Rivas, R.; Velázquez, E.; Willems, A.; Vizcaíno, N.; Subba-Rao, N.S.; Mateos, P.F.; Gillis, M.; Dazzo, F.B.; Martínez-Molina, E. A New Species of Devosia That Forms a Unique Nitrogen-Fixing Root-Nodule Symbiosis with the Aquatic Legume Neptunia natans (L.f.) Druce. Appl. Environ. Microbiol. 2002, 68, 5217–5222. [Google Scholar] [CrossRef] [Green Version]
- Madhaiyan, M.; Poonguzhali, S.; Lee, J.-S.; Senthilkumar, M.; Lee, K.C.; Sundaram, S. Mucilaginibacter gossypii sp. nov. and Mucilaginibacter gossypiicola sp. nov., Plant-Growth-Promoting Bacteria Isolated from Cotton Rhizosphere Soils. Int. J. Syst. Evol. Microbiol. 2010, 60, 2451–2457. [Google Scholar] [CrossRef]
- Kang, S.J.; Jung, Y.T.; Oh, K.H.; Oh, T.K.; Yoon, J.H. Mucilaginibacter boryungensis sp. nov., Isolated from Soil. Int. J. Syst. Evol. Microbiol. 2011, 61, 1549–1553. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.-H.; Kang, S.-J.; Park, S.; Oh, T.-K. Mucilaginibacter litoreus sp. nov., Isolated from Marine Sand. Int. J. Syst. Evol. Microbiol. 2012, 62, 2822–2827. [Google Scholar] [CrossRef]
- Kämpfer, P.; Busse, H.J.; McInroy, J.A.; Glaeser, S.P. Mucilaginibacter auburnensis sp. nov., Isolated from a Plant Stem. Int. J. Syst. Evol. Microbiol. 2014, 64, 1736–1742. [Google Scholar] [CrossRef] [PubMed]
- Bautista, V.V.; Monsalud, R.G.; Yokota, A. Devosia yakushimensis sp. nov., Isolated from Root Nodules of Pueraria Lobata (Willd.) Ohwi. Int. J. Syst. Evol. Microbiol. 2020, 60, 627–632. [Google Scholar] [CrossRef]
- Mayrberger, J.M. Studies of Genera Cytophaga-Flavobacterium in Context of the Soil Carbon Cycle; Michigan State University: East Lansing, MI, USA, 2011. [Google Scholar]
- Arntzen, M.Ø.; Várnai, A.; Mackie, R.I.; Eijsink, V.G.H.; Pope, P.B. Outer Membrane Vesicles from Fibrobacter succinogenes S85 Contain an Array of Carbohydrate-Active Enzymes with Versatile Polysaccharide-Degrading Capacity: F. succinogenes Outer Membrane Vesicles. Environ. Microbiol. 2017, 19, 2701–2714. [Google Scholar] [CrossRef] [Green Version]
- Tolba, S.T.M.; Ibrahim, M.; Amer, E.A.M.; Ahmed, D.A.M. First Insights into Salt Tolerance Improvement of Stevia by Plant Growth-Promoting Streptomyces Species. Arch. Microbiol. 2019, 201, 1295–1306. [Google Scholar] [CrossRef]
- Xu, L.; Sun, J.-Q.; Wang, L.-J.; Gao, Z.-W.; Sun, L.-Z.; Wu, X.-L. Sphingobacterium alkalisoli Sp. Nov., Isolated from a Saline-Alkaline Soil. Int. J. Syst. Evol. Microbiol. 2017, 67, 1943–1948. [Google Scholar] [CrossRef] [PubMed]
- Vaishnav, A.; Singh, J.; Singh, P.; Rajput, R.S.; Singh, H.B.; Sarma, B.K. Sphingobacterium sp. BHU-AV3 Induces Salt Tolerance in Tomato by Enhancing Antioxidant Activities and Energy Metabolism. Front. Microbiol. 2020, 11, 433. [Google Scholar] [CrossRef] [PubMed]
- Nozari, R.M.; Ortolan, F.; Astarita, L.V.; Santarém, E.R. Streptomyces spp. Enhance Vegetative Growth of Maize Plants under Saline Stress. Braz. J. Microbiol. 2021, 52, 1371–1383. [Google Scholar] [CrossRef] [PubMed]
- Costa, O.Y.A.; Raaijmakers, J.M.; Kuramae, E.E. Microbial Extracellular Polymeric Substances: Ecological Function and Impact on Soil Aggregation. Front. Microbiol. 2018, 9, 1636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedlacek, D.J. Soil Aggregation by Streptomyces Bacteria. Ohio J. Sci. 2018, 118, A45. [Google Scholar]
- Matsuda, Y.; Ida, Y.; Shinogi, T.; Kakutani, K.; Nonomura, T.; Toyoda, H. In Vitro Supression of Mycelial Growth of Fusarium oxysporum by Extracellular Chitosanase of Sphingobacterium multivorum and Cloning of the Chitosanase Gene csnSM1. J. Gen. Plant. Pathol. 2001, 67, 318–324. [Google Scholar] [CrossRef]
- Martin, P.A.W.; Travers, R.S. Worldwide Abundance and Distribution of Bacillus thuringiensis Isolates. Appl. Environ. Microbiol. 1989, 55, 2437–2442. [Google Scholar] [CrossRef] [Green Version]
- González, A.; Díaz, R.; Díaz, M.; Borrero, Y.; Bruzón, Y.R.; Carreras, B.; Gato, R. Characterization of Bacillus thuringiensis Soil Isolates from Cuba, with Insecticidal Activity against Mosquitoes. Rev. Biol. Trop. 2011, 59, 1007–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmisano, M.M.; Nakamura, L.K.; Duncan, K.E.; Istock, C.A.; Cohan, F.M. Bacillus sonorensis sp. nov., a Close Relative of Bacillus licheniformis, Isolated from Soil in the Sonoran Desert, Arizona. Int. J. Syst. Evol. Microbiol. 2001, 51, 1671–1679. [Google Scholar] [CrossRef]
- Roberts, M.S.; Nakamura, L.K.; Cohan, F.M. Bacillus vallismortis sp. nov., a Close Relative of Bacillus subtilis, Isolated from Soil in Death Valley, California. Int. J. Syst. Bacteriol. 1996, 46, 470–475. [Google Scholar] [CrossRef] [Green Version]
- Yoon, J.-H.; Kang, S.-J.; Oh, T.-K. Pedobacter terrae sp. nov., Isolated from Soil. Int. J. Syst. Evol. Microbiol. 2007, 57, 2462–2466. [Google Scholar] [CrossRef]
- Dahal, R.H.; Kim, J. Pedobacter humicola sp. nov., a Member of the Genus Pedobacter Isolated from Soil. Int. J. Syst. Evol. Microbiol. 2016, 66, 2205–2211. [Google Scholar] [CrossRef]
- Roh, S.W.; Quan, Z.-X.; Nam, Y.-D.; Chang, H.-W.; Kim, K.-H.; Kim, M.-K.; Im, W.-T.; Jin, L.; Kim, S.-H.; Lee, S.-T.; et al. Pedobacter agri sp. nov., from Soil. Int. J. Syst. Evol. Microbiol. 2008, 58, 1640–1643. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Kong, W.; Wu, N.; Zhang, Y. Bacterial Diversity and Community along the Succession of Biological Soil Crusts in the Gurbantunggut Desert, Northern China: Variation of Bacteria along the Succession of BSCs. J. Basic Microbiol. 2016, 56, 670–679. [Google Scholar] [CrossRef]
- Ofek, M.; Hadar, Y.; Minz, D. Ecology of Root Colonizing Massilia (Oxalobacteraceae). PLoS ONE 2012, 7, e40117. [Google Scholar] [CrossRef]
- Nagy, M.L.; Pérez, A.; Garcia-Pichel, F. The Prokaryotic Diversity of Biological Soil Crusts in the Sonoran Desert (Organ Pipe Cactus National Monument, AZ). FEMS Microbiol. Ecol. 2005, 54, 233–245. [Google Scholar] [CrossRef] [Green Version]
- Ren, M.; Li, X.; Zhang, Y.; Jin, Y.; Li, S.; Huang, H. Massilia armeniaca sp. nov., Isolated from Desert Soil. Int. J. Syst. Evol. Microbiol. 2018, 68, 2319–2324. [Google Scholar] [CrossRef]
- Marchante, E.; Kjøller, A.; Struwe, S.; Freitas, H. Invasive Acacia longifolia Induce Changes in the Microbial Catabolic Diversity of Sand Dunes. Soil Biol. Biochem. 2008, 40, 2563–2568. [Google Scholar] [CrossRef]
- Niu, J.; Rang, Z.; Zhang, C.; Chen, W.; Tian, F.; Yin, H.; Dai, L. The Succession Pattern of Soil Microbial Communities and Its Relationship with Tobacco Bacterial Wilt. BMC Microbiol. 2016, 16, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanikawa, D.; Nakamura, Y.; Tokuzawa, H.; Hirakata, Y.; Hatamoto, M.; Yamaguchi, T. Effluent Treatment in an Aquaponics-Based Closed Aquaculture System with Single-Stage Nitrification–Denitrification Using a down-Flow Hanging Sponge Reactor. Int. Biodeterior. Biodegrad. 2018, 132, 268–273. [Google Scholar] [CrossRef]
- Yi, X.; Yi, K.; Fang, K.; Gao, H.; Dai, W.; Cao, L. Microbial Community Structures and Important Associations Between Soil Nutrients and the Responses of Specific Taxa to Rice-Frog Cultivation. Front. Microbiol. 2019, 10, 1752. [Google Scholar] [CrossRef] [Green Version]
- Navarrete, A.A.; Soares, T.; Rossetto, R.; van Veen, J.A.; Tsai, S.M.; Kuramae, E.E. Verrucomicrobial Community Structure and Abundance as Indicators for Changes in Chemical Factors Linked to Soil Fertility. Antonie Van Leeuwenhoek 2015, 108, 741–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguirre-von-Wobeser, E.; Rocha-Estrada, J.; Shapiro, L.R.; de la Torre, M. Enrichment of Verrucomicrobia, Actinobacteria and Burkholderiales Drives Selection of Bacterial Community from Soil by Maize Roots in a Traditional Milpa Agroecosystem. PLoS ONE 2018, 13. [Google Scholar] [CrossRef]
- Tian, Y.; Gao, L. Bacterial Diversity in the Rhizosphere of Cucumbers Grown in Soils Covering a Wide Range of Cucumber Cropping Histories and Environmental Conditions. Microb. Ecol. 2014, 68, 794–806. [Google Scholar] [CrossRef] [PubMed]
- Calderoli, P.A.; Collavino, M.M.; Behrends Kraemer, F.; Morrás, H.J.M.; Aguilar, O.M. Analysis of Nif H-RNA Reveals Phylotypes Related to Geobacter and Cyanobacteria as Important Functional Components of the N2-Fixing Community Depending on Depth and Agricultural Use of Soil. MicrobiologyOpen 2017, 6, e00502. [Google Scholar] [CrossRef] [PubMed]
- Masuda, Y.; Yamanaka, H.; Xu, Z.-X.; Shiratori, Y.; Aono, T.; Amachi, S.; Senoo, K.; Itoh, H. Diazotrophic Anaeromyxobacter Isolates from Soils. Appl. Environ. Microbiol. 2020, 86, e00956-20. [Google Scholar] [CrossRef]
- Bonanomi, G.; Cesarano, G.; Antignani, V.; Di Maio, C.; De Filippis, F.; Scala, F. Conventional Farming Impairs Rhizoctonia solani Disease Suppression by Disrupting Soil Food Web. J. Phytopathol. 2018, 166, 663–673. [Google Scholar] [CrossRef]
- Lebreton, L.; Guillerm-Erckelboudt, A.-Y.; Gazengel, K.; Linglin, J.; Ourry, M.; Glory, P.; Sarniguet, A.; Daval, S.; Manzanares-Dauleux, M.J.; Mougel, C. Temporal Dynamics of Bacterial and Fungal Communities during the Infection of Brassica rapa Roots by the Protist Plasmodiophora brassicae. PLoS ONE 2019, 14, e0204195. [Google Scholar] [CrossRef] [Green Version]
- Olanya, O.M.; Lakshman, D.K. Potential of Predatory Bacteria as Biocontrol Agents for Foodborne and Plant Pathogens. J. Plant. Pathol. 2015, 97, 405–417. [Google Scholar]
- Yang, G.; Lin, J.; Zeng, E.Y.; Zhuang, L. Extraction and Characterization of Stratified Extracellular Polymeric Substances in Geobacter Biofilms. Bioresour. Technol. 2019, 276, 119–126. [Google Scholar] [CrossRef]
- Yin, W.; Wang, Y.; Liu, L.; He, J. Biofilms: The Microbial “Protective Clothing” in Extreme Environments. Int. J. Mol. Sci. 2019, 20, 3423. [Google Scholar] [CrossRef] [Green Version]
- Gomes, D.O.; da Chagas, L.G.S.; Ramos, G.B.; Ciuffa, A.Z.; Rezende, L.M.; Cuccato, L.P.; Dos Reis, T.F.M.; Pires, B.C.; Lima, A.M.C. Biofilm Production of Leptospira spp. Strains. Acta Sci. Vet. 2018, 46, 5. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pairwise Comparison | R Value | p Value |
|---|---|---|
| A. breviligulata versus C. kobomugi | 0.08866 | 0.0466 |
| A. breviligulata versus Barren | 0.06821 | 0.0231 |
| C. kobomugi versus Barren | 0.2294 | 0.0005 |
| Microhabitat % Composition | Statistical Result | ||||
|---|---|---|---|---|---|
| Genus | AB | CK | Barren | ANOVA | Tukey HSD |
| Devosia | 0.86 ± 0.14 | 0.90 ± 0.09 | 0.24 ± 0.05 | F2,85 = 35.88, p < 0.0001 * | (AB = CK) > Barren |
| Fibrobacter | 2.24 ± 0.42 | 3.09 ± 0.71 | 0.17 ± 0.03 | F2,85 = 42.64, p < 0.0001 * | |
| Mesorhizobium | 0.76 ± 0.11 | 0.91 ± 0.09 | 0.41 ± 0.07 | F2,85 = 14.01, p < 0.0001 * | |
| Mucilaginibacter | 0.72 ± 0.17 | 0.79 ± 0.19 | 0.13 ± 0.01 | F2,85 = 24.13, p < 0.0001 * | |
| Ohtaekwangia | 0.86 ± 0.26 | 0.60 ± 0.15 | 0.10 ± 0.04 | F2,85 = 19.87 p < 0.0001 * | |
| Sphingobacterium | 0.90 ± 0.10 | 0.86 ± 0.07 | 0.50 ± 0.05 | F2,85 = 13.10, p < 0.0001 * | |
| Bacteriovorax | 0.44 ± 0.11 | 0.24 ± 0.07 | 0.16 ± 0.03 | F2,85 = 4.80, p = 0.01 * | AB > Barren |
| Flavisolibacter | 0.75 ± 0.18 | 0.42 ± 0.08 | 0.22 ± 0.03 | F2,85 = 10.23, p < 0.0001 * | |
| Cytophaga | 1.15 ± 0.24 | 2.86 ± 0.54 | 0.09 ± 0.01 | F2,85 = 74.78, p < 0.0001 * | CK >AB > Barren |
| Streptomyces | 1.10 ± 0.32 | 2.12 ± 0.34 | 0.35 ± 0.04 | F2,85 = 24.04, p < 0.0001 * | |
| Ktedonobacter | 0.01 ± 0.00 | 0.46 ± 0.26 | 0.00 ± 0.00 | F2,85 = 6.65, p = 0.002 * | CK > (AB = Barren) |
| Opitutus | 0.66 ± 0.07 | 1.15 ± 0.13 | 0.50 ± 0.05 | F2,85 = 19.28, p < 0.0001 * | |
| Acidobacterium | 2.63 ± 0.31 | 2.50 ± 0.23 | 3.75 ± 0.30 | F2,85 = 4.40, p = 0.02 * | Barren > CK |
| Bacillus | 1.34 ± 0.20 | 0.79 ± 0.19 | 2.14 ± 0.31 | F2,85 = 6.88, p = 0.002 * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boss, B.L.; Charbonneau, B.R.; Izquierdo, J.A. Spatial Diversity in Bacterial Communities across Barren and Vegetated, Native and Invasive, Coastal Dune Microhabitats. Diversity 2021, 13, 525. https://doi.org/10.3390/d13110525
Boss BL, Charbonneau BR, Izquierdo JA. Spatial Diversity in Bacterial Communities across Barren and Vegetated, Native and Invasive, Coastal Dune Microhabitats. Diversity. 2021; 13(11):525. https://doi.org/10.3390/d13110525
Chicago/Turabian StyleBoss, Brianna L., Bianca R. Charbonneau, and Javier A. Izquierdo. 2021. "Spatial Diversity in Bacterial Communities across Barren and Vegetated, Native and Invasive, Coastal Dune Microhabitats" Diversity 13, no. 11: 525. https://doi.org/10.3390/d13110525
APA StyleBoss, B. L., Charbonneau, B. R., & Izquierdo, J. A. (2021). Spatial Diversity in Bacterial Communities across Barren and Vegetated, Native and Invasive, Coastal Dune Microhabitats. Diversity, 13(11), 525. https://doi.org/10.3390/d13110525