
A Hotspot of Arid Zone Subterranean Biodiversity: The Robe Valley in Western Australia


Abstract

1. Introduction
2. Materials and Methods
Determining Species Numbers and Subterranean Affinity
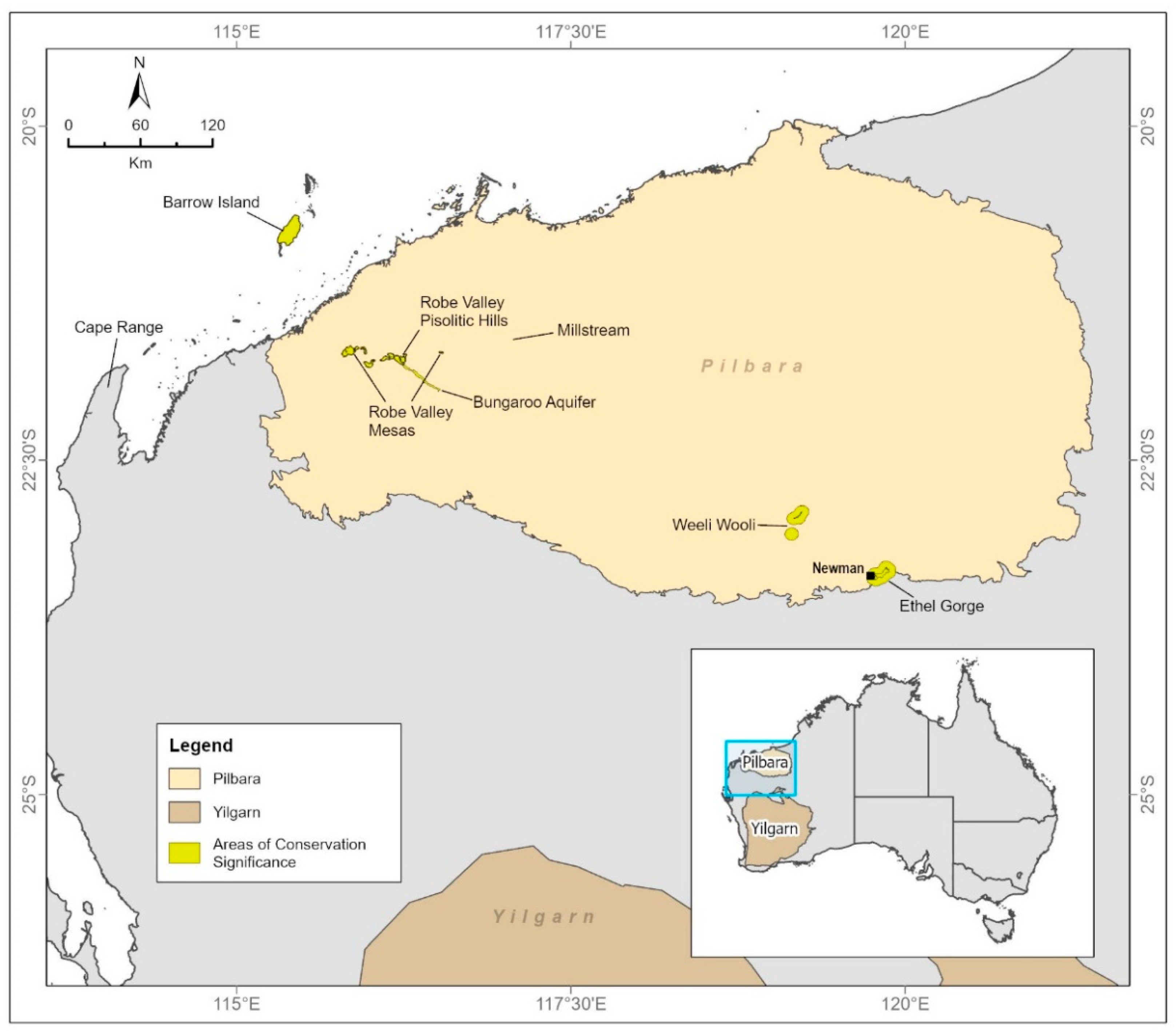
3. Environment and Geology
4. Results
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moldovan, O.T.; Kovác, L.; Halse, S. Cave Ecology; Springer Nature: Cham, Switzerland, 2018; p. 545. [Google Scholar]
- Culver, D.C.; Deharveng, L.; Bedos, A.; Lewis, J.J.; Madden, M.; Reddell, J.R.; Sket, B.; Trontelj, P.; White, D. The mid-latitude biodiversity ridge in terrestrial cave fauna. Ecography 2006, 29, 120–128. [Google Scholar] [CrossRef]
- Schneider, K.; Culver, D.C. Estimating subterranean species richness using intensive sampling and rarefaction curves in a high density cave region in West Virginia. J. Cave Karst Stud. 2004, 66, 39–45. [Google Scholar]
- Hahn, H.J.; Fuchs, A. Distribution patterns of groundwater communities across aquifer types in south-western Germany. Freshwater Biol. 2009, 54, 848–860. [Google Scholar] [CrossRef]
- Halse, S.A. Subterranean fauna of the arid zone. In On the Ecology of Australia’s Arid Zone; Lambers, H., Ed.; Springer Nature: Cham, Switzerland, 2018; p. 388. [Google Scholar]
- Pipan, T.; López, H.; Oromí, P.; Polak, S.; Culver, D.C. Temperature variation and the presence of troglobionts in terrestrial shallow subterranean habitats. J. Nat. Hist. 2010, 45, 253–273. [Google Scholar] [CrossRef]
- Pesce, G.L.; de Laurentiis, P.; Humphreys, W.F. Copepods from ground waters of Western Australia. II. The genus Halicyclops (Crustacea, Copepoda, Cyclopidae). Rec. West. Aust. Mus. 1996, 18, 77–85. [Google Scholar]
- Humphreys, W.F. Relict stygofauna’s living in sea salt, karst and calcrete habitats in arid northwestern Australia contain many ancient lineages. In The Other 99%: The Conservation and Biodiversity of Invertebrates; Ponder, W., Lunney, D., Eds.; Royal Zoological Society of New South Wales: Sydney, Australia, 1999; pp. 219–227. [Google Scholar]
- Eberhard, S.M.; Halse, S.A.; Williams, M.R.; Scanlon, M.D.; Cocking, J.S.; Barron, H.J. Exploring the relationship between sampling efficiency and short range endemism for groundwater fauna in the Pilbara region, Western Australia. Freshwater Biol. 2009, 54, 885–901. [Google Scholar] [CrossRef]
- Halse, S.A.; Scanlon, M.D.; Cocking, J.S.; Barron, H.J.; Richardson, J.B.; Eberhard, S.M. Pilbara stygofauna: Deep groundwater of an arid landscape contains globally significant radiation of biodiversity. Rec. West. Aust. Mus. Suppl. 2014, 78, 443–483. [Google Scholar] [CrossRef]
- Humphreys, G.; Alexander, J.; Harvey, M.S.; Humphreys, W.F. The subterranean fauna of Barrow Island, north-western Australia: 10 years on. Rec. West. Aust. Mus. Suppl. 2013, 83, 145–158. [Google Scholar] [CrossRef][Green Version]
- Mokany, K.; Harwood, T.D.; Halse, S.A.; Ferrier, S. Riddles in the dark: Assessing diversity patterns for cryptic subterranean fauna of the Pilbara. Divers. Distrib. 2019, 25, 240–254. [Google Scholar] [CrossRef]
- Biota. Mesa A and Robe Valley Mesas Troglobitic Fauna Survey; Biota Environmental Sciences: Leederville, Australia, 2006. Available online: https://www.epa.wa.gov.au/sites/default/files/PER_documentation/Vegetation%20and%20Flora%20and%20Fauna_Biota%202006f%20Subterranean%20Fauna%20Assessment.pdf (accessed on 9 September 2021).
- Harvey, M.S.; Berry, O.; Edward, K.L.; Humphreys, G. Molecular and morphological systematics of hypogean schizomids (Schizomida:Hubbardiidae) in semiarid Australia. Invertebr. Syst. 2008, 22, 167–194. [Google Scholar] [CrossRef]
- Guzik, M.T.; Austin, A.D.; Cooper, S.J.B.; Harvey, M.S.; Humphreys, W.F.; Bradford, T.; Eberhard, S.M.; King, R.A.; Leys, R.; Muirhead, K.A.; et al. Is the Australian subterranean fauna uniquely diverse? Invertebr. Syst. 2010, 24, 407–418. [Google Scholar] [CrossRef]
- Byrne, M.; Yeates, D.K.; Joseph, L.; Kearney, M.; Bowler, J.; Williams, A.J.; Cooper, S.; Donnellan, S.C.; Keogh, S.; Leys, R.; et al. Birth of a biome: Insights into the assembly and maintenance of the Australian arid zone biota. Mol. Ecol. 2008, 17, 4398–4417. [Google Scholar] [CrossRef] [PubMed]
- Abrams, K.M.; Huey, J.A.; Hillyer, M.J.; Humphreys, W.F.; Didham, R.K.; Harvey, M.S. Too hot to handle: Cenozoic aridification drives multiple independent incursions of Schizomida (Hubbardiidae) into hypogean environments. Mol. Phylogenet. Evol. 2019, 139, 106532. [Google Scholar] [CrossRef]
- Harms, D.; Curran, M.K.; Klesser, R.; Finston, T.L.; Halse, S.A. Speciation patterns in complex subterranean environments: A case study using short-tailed whipscorpions (Schizomida: Hubbardiidae). Biol. J. Linn. Soc. 2018, 125, 355–367. [Google Scholar] [CrossRef]
- Karanovic, I. Candoninae (Ostracoda) from the Pilbara region in Western Australia. Crustaceana Monogr. 2007, 7, 1–432. [Google Scholar]
- Matthews, E.F.; Abrams, K.M.; Cooper, S.J.B.; Huey, J.A.; Hillyer, M.J.; Humphreys, W.F.; Austin, A.D.; Guzik, M.T. Scratching the surface of subterranean biodiversity: Molecular analysis reveals a diverse and previously unknown fauna of Parabathynellidae (Crustacea: Bathynellacea) from the Pilbara, Western Australia. Mol. Phylogenet. Evol. 2020, 142, 106643. [Google Scholar] [CrossRef]
- Finston, T.L.; Johnson, M.S.; Humphreys, W.F.; Eberhard, S.M.; Halse, S.A. Cryptic speciation in two widespread subterranean amphipod genera reflects historical drainage patterns in an ancient landscape. Mol. Ecol. 2007, 16, 355–365. [Google Scholar] [CrossRef]
- King, R.A.; Fagan-Jefferies, E.; Bradford, T.M.; Stringer, D.M.; Finston, T.L.; Halse, S.A.; Eberhard, S.M.; Humphreys, G.; Humphreys, W.F.; Austin, A.D.; et al. Cryptic diversity Down Under: Defining species in the subterranean amphipod genus Nedsia Barnard and Williams (Hadzioidea: Eriopisidae) from the Pilbara, Western Australia. Invertebr. Syst. 2021, in press. [Google Scholar]
- Pinder, A.M.; Halse, S.A.; Shiel, R.J.; McRae, J.M. An arid zone awash with diversity: Patterns in the distribution of aquatic invertebrates in the Pilbara region of Western Australia. Rec. West. Aust. Mus. Suppl. 2010, 78, 205–246. [Google Scholar] [CrossRef]
- Eberhard, S.M.; Howarth, F.G. Undara lava cave fauna in tropical Queensland with an annotated list of Australian subterranean biodiversity hotspots. Diversity 2021, 13, 326. [Google Scholar] [CrossRef]
- Deharveng, L.; Bedos, A. Biodiversity in the tropics. In Encyclopedia of Caves, 3rd ed.; White, W.B., Culver, D.C., Pipan, T., Eds.; Academic Press: New York, NY, USA, 2019; pp. 146–162. [Google Scholar]
- Halse, S.A.; Pearson, G.B. Troglofauna in the vadose zone: Comparison of scraping and trapping results and sampling adequacy. Subterr. Biol. 2014, 13, 17–34. [Google Scholar] [CrossRef]
- Buick, R.; Thornett, J.R.; McNaughton, N.J.; Smith, J.B.; Barley, M.E.; Savage, M. Record of emergent continental crust ~3.5 billion years ago in the Pilbara Craton of Australia. Nature 1995, 375, 574–577. [Google Scholar] [CrossRef]
- Johnson, D.P. The Geology of Australia; Cambridge University Press: London, UK, 2004. [Google Scholar]
- Johnson, S.L.; Wright, A.H. Central Pilbara Groundwater Study; Hydrogeological Record Series; Report HG 8; Water and Rivers Commission Resource Science Division: East Perth, WA, Australia, 2001; p. 124. [Google Scholar]
- Skrzypek, G.; Dogramaci, S.; Page, G.F.M.; Rouillard, A.; Grierson, P.F. Unique stable isotope signatures of large cyclonic events as a tracer of soil moisture dynamics in the semiarid subtropics. J. Hydrol. 2019, 578, 124124. [Google Scholar] [CrossRef]
- Howarth, F.G.; Moldovan, O.T. The ecological classifications of cave animals and their adaptations. In Cave Ecology; Moldovan, O.T., Kovac, L., Halse, S., Eds.; Springer Nature: Cham, Switzerland, 2018; pp. 41–67. [Google Scholar]
- Ferreira, R.L.; de Oliveira, M.P.A.; Silva, M.S. Subterranean biodiversity in ferruginous landscapes. In Cave Ecology; Moldovan, O.T., Kovac, L., Halse, S., Eds.; Springer Nature: Cham, Switzerland, 2018; pp. 435–447. [Google Scholar]
- Morris, R.C.; Ramanaidou, E.R. Genesis of the channel iron deposits (CID) of the Pilbara region, Western Australia. Australian Aust. J. Earth Sci. 2007, 54, 733–756. [Google Scholar] [CrossRef]
- Danišík, M.; Evans, N.J.; Ramanaidou, E.R.; McDonald, B.J.; Mayers, C.; McInnes, B.I.A. (U–Th)/He chronology of the Robe River channel iron deposits, Hamersley Province, Western Australia. Chem. Geol. 2013, 354, 150–162. [Google Scholar] [CrossRef]
- Mammola, S.; Giachino, P.M.; Piano, E.J.A.; Barberis, M.; Badino, G.; Isaia, M. Ecology and sampling techniques of an understudied subterranean habitat: The Milieu Souterrain Superficiel (MSS). Sci. Nat. 2016, 103, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Department of Water. Ecological Water Requirements of the Lower Robe River; Environmental Water Report Series 22; Department of Water: Perth, Australia, 2012; p. 38. [Google Scholar]
- Edward, K.L.; Harvey, M.S. Short-range endemism in hypogean environments: The pseudoscorpion genera Tyrannochthonius and Lagynochthonius (Pseudoscorpiones: Chthoniidae) in the semiarid zone of Western Australia. Invertebr. Syst. 2008, 22, 259–293. [Google Scholar] [CrossRef]
- Car, C.A.; Megan, S.; Huynh, C.; Harvey, M.S. The millipedes of Barrow Island, Western Australia (Diplopoda). Rec. West. Aust. Mus. Suppl. 2013, 83, 209–219. [Google Scholar] [CrossRef]
- Harvey, M.S.; Leng, M.C. Further observations on Ideoblothrus (Pseudoscorpiones: Syarinidae) from subterranean environments in Australia. Rec. West. Aust. Mus. 2008, 24, 381–386. [Google Scholar] [CrossRef][Green Version]
- Sket, B. Can we agree on an ecological classification of subterranean animals? J. Nat. Hist. 2008, 42, 1549–1563. [Google Scholar] [CrossRef]
- McKenzie, N.L.; van Leeuwen, S.; Pinder, A.M. Introduction to the Pilbara biodiversity survey, 2002–2007. Rec. West. Aust. Mus. Supppl. 2009, 78, 3–89. [Google Scholar] [CrossRef]
- Page, T.J.; Hughes, J.M.; Real, K.M.; Stevens, M.I.; King, R.A.; Humphreys, W.F. Allegory of a cave crustacean: Systematic and biogeographic reality of Halosbaena (Peracarida: Thermosbaenacea) sought with molecular data at multiple scales. Mar. Biodivers. 2016, 48, 1185–1202. [Google Scholar] [CrossRef]
- Des Châtelliers, M.C.; Juget, J.; Lafont, M.; Martin, P. Subterranean aquatic Oligochaeta. Freshwater Biol. 2009, 54, 678–690. [Google Scholar] [CrossRef]
- Sorensen, J.P.R.; Maurice, L.; Edwards, F.K.; Lapworth, D.J.; Read, D.S.; Allen, D.; Butcher, A.S.; Newbold, L.K.; Townsend, B.R.; Williams, P.J. Using boreholes as windows into groundwater ecosystems. PLoS ONE 2013, 8, e70264. [Google Scholar] [CrossRef] [PubMed]
- Halse, S.A.; Scanlon, M.D.; Cocking, J.S. Do Springs Provide a Window to the Groundwater Fauna of the Australian Arid Zone? In Balancing the Groundwater Budget, Proceedings of an International Groundwater Conference, Darwin, Australia, 12–17 May 2002; Yinfoo, D., Ed.; International Association of Hydrologists: Wallingford, UK, 2002; p. 12. [Google Scholar]
- Sket, B.; Paragamian, K.; Trontelj, P. A census of the obligate subterranean fauna of the Balkan Peninsula. In Balkan Biodiversity; Pattern and Process in the European Hotspot; Griffiths, H.I., Kryštufek, B., Reed, J.M., Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 309–332. [Google Scholar]
- Deharveng, L.; Gibert, J.; Culver, D.C. Biodiversity in Europe. In Encyclopedia of Caves, 3rd ed.; White, W.B., Culver, D.C., Pipan, T., Eds.; Academic Press: New York, NY, USA, 2019; pp. 136–145. [Google Scholar]
- Zagmajster, M.; Polak, S.; Fišer, C. Postojna-Planina cave system in Slovenia, a hotspot of subterranean biodiversity and a cradle of speleobiology. Diversity 2021, 13, 271. [Google Scholar] [CrossRef]
- Pipan, T.; Deharveng, L.; Culver, D.C. Hotspots of subterranean biodiversity. Diversity 2020, 12, 209. [Google Scholar] [CrossRef]
- Karanovic, T. Subterranean copepods (Crustacea, Copepoda) from the Pilbara region in Western Australia. Rec. West. Aust. Mus. Suppl. 2006, 70, 1–239. [Google Scholar] [CrossRef]
- Karanovic, T.; Djurakic, M.; Eberhard, S.M. Cryptic species or inadequate taxonomy? Implementation of 2D geometric morphometrics based on integumental organs as landmarks for delimitation and description of copepod taxa. Syst. Biol. 2016, 65, 304–327. [Google Scholar] [CrossRef]
- Trotter, A.J.; McRae, J.M.; Main, D.C.; Finston, T.L. Speciation in fractured rock landforms: Towards understanding the diversity of subterranean cockroaches (Dictyoptera: Nocticolidae: Nocticola) in Western Australia. Zootaxa 2017, 4232, 361–375. [Google Scholar] [CrossRef]
- Moore, G.I.; Humphreys, W.F.; Foster, R. New populations of the rare subterranean blind cave eel Ophisternon candidum (Synbranchidae) reveal recent historical connections throughout north-western Australia. Mar. Freshwater Res. 2018, 69, 1517–1524. [Google Scholar] [CrossRef]
- Perina, G.; Camacho, A.I.; Huey, J.; Horwitz, P.; Koenders, A. Understanding subterranean variability: The first genus of Bathynellidae (Bathynellacea, Crustacea) from Western Australia described through a morphological and multigene approach. Invertebr. Syst. 2018, 32, 423–447. [Google Scholar] [CrossRef]
- Coates, D.J.; Byrne, M.; Moritz, C. Genetic diversity and conservation units: Dealing with the species-population continuum in the age of genomics. Front. Ecol. Evol. 2018, 6. [Google Scholar] [CrossRef]
- Wesener, T.; Voigtländer, K.; Decker, P.; Oeyen, J.P.; Spelda, J. Barcoding of Central European Cryptops centipedes reveals large interspecific distances with ghost lineages and new species records from Germany and Austria (Chilopoda, Scolopendromorpha). ZooKeys 2016, 564, 21–46. [Google Scholar] [CrossRef] [PubMed]
- Halse, S. Conservation and impact assessment of subterranean fauna in Australia. In Cave Ecology; Moldovan, O.T., Kovacz, L., Halse, S., Eds.; Springer Nature: Gland, Switzerland, 2018; pp. 479–493. [Google Scholar]
- Environmental Protection Authority. Report and Recommendations of the Environmental Protection Authority: Mesa A Hub Revised Proposal. Report 1640. Environmental Protection Authority, Perth, Australia. 2019. Available online: https://www.epa.wa.gov.au/sites/default/files/EPA_Report/Mesa%20A%20Hub%20Revised%20Proposal%20-%20EPA%20Report.pdf (accessed on 9 September 2021).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxonomic Group | Family | Species | A | B | C | Comment |
|---|---|---|---|---|---|---|
| Chilopoda | Cryptopidae | Cryptopidae ‘Helix-CHI023’ | ✓ * | ✓ | CO1 | |
| Cryptopidae ‘Helix-CHI026’ | ✓ | CO1 | ||||
| Scolopendridae | Cormocephalus sp. | ✓ * | ||||
| Geophilidae | Geophilidae sp. | ✓ * | ||||
| Diplopoda | Haplodesmidae | Haplodesmidae ‘Helix-DIHA001’ | ✓ * | ✓ | ✓ | CO1 |
| Haplodesmidae ‘Helix-DIHA005’ | ✓ | CO1 | ||||
| Lophoproctidae | Lophoturus madecassus (Marquet & Conde, 1950)# | ✓ | ✓ | |||
| Scolopendrellidae ’Helix-SYM026’ | ✓ | ✓ * | ||||
| Pseudoscorpiones | Chthoniidae | Lagynochthonius asemaEdward & Harvey, 2008 | ✓ | |||
| Tyrannochthonius aridus Edward & Harvey, 2008# | ✓ | ✓ | ||||
| Tyrannochthonius basme Edward & Harvey, 2008 | ✓ | |||||
| Tyrannochthonius‘MesaA’ | ✓ | |||||
| Chthoniidae ‘Helix_PCH047’ | ✓ | CO1, >20% | ||||
| Chthoniidae ‘Helix_PCH049’ | ✓ | CO1, 10% | ||||
| Chthoniidae ‘Helix_PCH050’ | ✓ | CO1 | ||||
| Chthoniidae ‘Helix_PCH058’ | ✓ | CO1 | ||||
| Olpiidae | Olpiidae ‘POL013’ | ✓ | ||||
| Hyidae | Hyidae ‘Helix-PH001’ | ✓ | CO1, 5% | |||
| Hyidae ‘Helix-PH008’ | ✓ | CO1, 5% | ||||
| Syarinidae | Ideoblothrus linnaei Harvey & Leng, 2008 | ✓ | ||||
| Ideoblothrus pisolitus Harvey & Edward, 2007 | ✓ | CO1, 6% | ||||
| Ideoblothrus‘MesaA’ | ✓ | |||||
| Ideoblothrus ‘MesaA2’ | ✓ | CO1,>9% | ||||
| Ideoblothrus ‘MesaA3’ | ✓ | CO1, >9% | ||||
| Ideoblothrus ‘MesaA4’ | ✓ | CO1, >18% | ||||
| Araneae | Gnaphosidae | Gnaphosidae ‘Helix-AG001’ | ✓ | |||
| Oonopidae | Prethopalpus scanloni Baehr et al., 2012 | ✓ | ||||
| Prethopalpus ‘ARA051’ | ✓ | WAM | ||||
| Pholcidae | Trichocyclus sp. ‘MesaA’ | ✓ | ||||
| Theridiidae | Theridiidae ‘Helix-AT001’ | ✓ | ✓ | CO1 | ||
| Schizomida | Draculoides anachoretus(Harvey et al., 2008) | ✓ | ||||
| Draculoides bythius(Harvey et al., 2008) | ✓ | ✓ | ||||
| Hubbardiidae gen. nov. ‘Helix-SCH052’ | ✓ | ✓ | CO1, WAM | |||
| Isopoda | Armadillidae | Troglarmadillo ‘Helix-ISA005’ | ✓ | CO1 | ||
| Troglarmadillo ‘Helix-ISA008’ | ✓ | CO1 | ||||
| Armadillidae ‘Helix—ISA054’ | ✓ | CO1 | ||||
| Armadillidae ‘Helix—ISA056’ | ✓ | CO1 | ||||
| Armadillidae ‘MesaAOES19’ | ✓ | |||||
| Oniscidae | Hanoniscus ‘MesaAOES22’ | ✓ | ||||
| Philosciidae ‘Helix-ISP052’ | ✓ | CO1 | ||||
| Philosciidae ‘Helix-ISP053’ | ✓ | CO1 | ||||
| Diplura | Campodeidae | Campodeidae ‘Helix-DCA001’ | ✓ | ✓ * | CO1, 6% | |
| Campodeidae ‘Helix-DCA002’ | ✓ | CO1 | ||||
| Campodeidae sp. sp. B6 | ✓ | |||||
| Japygidae | Japygidae sp. | ✓ * | ✓ * | ✓ * | ||
| Parajapygidae | Parajapygidae ‘Helix-DPA003’ | ✓ | ✓ | |||
| Parajapygidae ‘Helix-DPA004’ | ✓ | CO1, 12–19% | ||||
| Parajapygidae ‘Helix-DPA006’ | ✓ | CO1 | ||||
| Parajapygidae ‘Helix-DPA008’ | ✓ | CO1 | ||||
| Projapygidae | Projapygidae sp. | ✓ * | ||||
| Zygentoma | Nicoletiidae | Atelurinae ‘Helix-TA009’ | ✓ | CO1 | ||
| Hemitrinemura sp. B4 | ✓ | |||||
| Trinemura ‘MesaA1’ | ✓ | |||||
| Trinemura ‘MesaA2’ | ✓ | |||||
| Nicoletiinae ‘Helix-TN010’ | ✓ | CO1 | ||||
| Nicoletiinae ‘Helix-TN012 | ✓ | CO1 | ||||
| Dictyoptera | Nocticolidae | Nocticola ‘OES11’ | ✓ | |||
| Hemiptera | Cixiidae sp. B02 | Cixiidae sp. B02# | ✓ | ✓ | ||
| Coleoptera | Carabidae | Gracilanillus hirsutus Giachino et al., 2021 | ✓ | ✓ | ||
| Angustanillus striatipennis Giachino et al., 2021 | ✓ | |||||
| Angustanillus armatus Giachino et al., 2021 | ✓ | |||||
| Ptiliidae | Ptinella sp. B01 | ✓ | ||||
| Staphylinidae | ?Staphylinidae ‘MesaKOES2’ | ✓ | ||||
| Curculionidae | Cryptorhynchinae ‘CCU004’ | ✓ | ✓ | ✓ | WAM | |
| Diptera | Sciaridae | Allopnyxia sp. B01# | ✓ |
| Taxonomic group | Family | Species | 2A | 1A |
|---|---|---|---|---|
| Rotifera | - | Bdelloidea sp. ‡ | ✓ * | |
| Philodinidae | Dissotrocha sp. ‡ | ✓ * | ||
| Nematoda | - | Nematoda sp. 02 (PSS) ‡ | ✓ | |
| Nematoda sp. 03 (PSS) ‡ | ✓ | |||
| Nematoda sp. 11 (PSS) ‡ | ✓ | |||
| Annelida | - | Hirudinea sp. ‡ | ✓ * | |
| Enchytraeidae | Enchytraeus sp. AP PSS1 ‡ | ✓ | ||
| Enchytraeus sp. AP PSS2 ‡ | ✓ | |||
| Phreodrilidae | Phreodrilidae sp. AP DVC | ✓ | ✓ | |
| Phreodrilidae sp. AP SVC | ✓ | |||
| Tubificidae | Tubificidae ‘stygo type 2A’ # | ✓ | ||
| Nereididae | Namanereis pilbarensis Glasby et al., 2014 | ✓ | ||
| Acarina | Arrenuridae | Arrenurus sp. nov. 2 (PSS) | ✓ | |
| Mideopsidae | Guineaxonopsis sp. S01 group | ✓ | ✓ | |
| Pezidae | Peza sp. | ✓ * | ||
| Unionicolidae | Unionicolidae sp. B02 | ✓ | ||
| Ostracoda | Limnocytheridae | Gomphodella hirsuta Karanovic, 2006 | ✓ | ✓ |
| Candonidae | Deminutiocandona aenigma Karanovic, 2007 | ✓ | ||
| Humphreyscandona fovea Karanovic & Marmonier, 2003 | ✓ | ✓ | ||
| Humphreyscandona imperfecta Karanovic, 2005 | ✓ | ✓ | ||
| Humphreyscandona pilbarae Karanovic & Marmonier, 2003 | ✓ | |||
| Humphreyscandona woutersi Karanovic & Marmonier, 2003 | ✓ | ✓ | ||
| Pierrecandona posteriorrecta Karanovic, 2007 | ✓ | |||
| Pilbaracandona rosa Karanovic, 2007 | ✓ | ✓ | ||
| Areacandona astrepte Karanovic, 2007 | ✓ | ✓ | ||
| Areacandona cylindrata Karanovic, 2007 | ✓ | |||
| Areacandona ‘4’ (PSS) | ✓ | |||
| Areacandona fortescueiensis Karanovic, 2007 | ✓ | |||
| Kencandona verrucosa Karanovic, 2007 | ✓ | |||
| Candonidae ‘2’ (PSS) | ✓ | |||
| Candonidae gen. 4 ‘1’ | ✓ | |||
| Copepoda | Ridgewayiidae | Stygoridgewayia trispinosa Tang et al., 2008 | ✓ | ✓ |
| Cyclopidae | Apocyclops dengizicus (Lepeshkin, 1900) # | ✓ | ||
| Halicyclops rochai Karanovic, 2006 | ✓ | ✓ | ||
| Orbuscyclops westaustraliensis Karanovic, 2006 | ✓ | |||
| Diacyclops einslei De Laurentiis et al., 1999 | ✓ | |||
| Diacyclops h. humphreysi x unispinosus (see Karanovic, 2006) | ✓ | |||
| Diacyclops humphreysi Pesce & De Laurentiis, 1996 | ✓ | |||
| Diacyclops h. unispinosus Karanovic, 2006 | ✓ | |||
| Diacyclops sobeprolatus Karanovic, 2006 | ✓ | |||
| Ectinosomatidae | Pseudectinosoma galassiae Karanovic, 2006 | ✓ | ✓ | |
| Diosaccidae | Schizopera roberiverensis Karanovic, 2006 | ✓ | ✓ | |
| Ameiridae | Megastygonitocrella unispinosa Karanovic, 2006 | ✓ | ✓ | |
| Megastygonitocrella trispinosa Karanovic, 2006 | ✓ | |||
| Parastenocarididae | Parastenocaris jane Karanovic, 2006 | ✓ | ||
| Syncarida | Bathynellidae | Bathynellidae sp. | ✓ * | |
| Parabathynellidae | Atopobathynella ‘A’ | ✓ | ✓ | |
| Thermosbaenacea | Halosbaenidae | Halosbaena sp. PL (see Page et al., 2016) | ✓ | ✓ |
| Amphipoda | Bogidiellidae | Bogidiellidae sp. | ✓ * | ✓ |
| Eriopisidae | Nedsia ‘mcraei’ King & Cooper, in press | ✓ | ✓ | |
| Nedsia ‘robensis’ King & Cooper, in press | ✓ | ✓ | ||
| Eriopisidae gen. nov. ‘Helix-AMM006’ | ✓ | |||
| Neoniphargidae | Wesniphargus ‘Helix-AMN004’ | ✓ | ||
| Paramelitidae | Pilbarus sp. | ✓ * | ✓ * | |
| Isopoda | Cirolanidae | Haptolana yarraloola Bruce, 2008 | ✓ | |
| Kagalana tonde Bruce, 2008 | ✓ | |||
| Olibrinidae | Adoniscus sp. | ✓ * | ||
| Microcerberidae | Microcerberidae sp. | ✓ * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clark, H.L.; Buzatto, B.A.; Halse, S.A. A Hotspot of Arid Zone Subterranean Biodiversity: The Robe Valley in Western Australia. Diversity 2021, 13, 482. https://doi.org/10.3390/d13100482
Clark HL, Buzatto BA, Halse SA. A Hotspot of Arid Zone Subterranean Biodiversity: The Robe Valley in Western Australia. Diversity. 2021; 13(10):482. https://doi.org/10.3390/d13100482
Chicago/Turabian StyleClark, Huon L., Bruno A. Buzatto, and Stuart A. Halse. 2021. "A Hotspot of Arid Zone Subterranean Biodiversity: The Robe Valley in Western Australia" Diversity 13, no. 10: 482. https://doi.org/10.3390/d13100482
APA StyleClark, H. L., Buzatto, B. A., & Halse, S. A. (2021). A Hotspot of Arid Zone Subterranean Biodiversity: The Robe Valley in Western Australia. Diversity, 13(10), 482. https://doi.org/10.3390/d13100482