A Hot Spot of Olive Biodiversity in the Tunisian Oasis of Degache

 ,
,  ,
,  ,
,  , , , and
, , , and 
Abstract
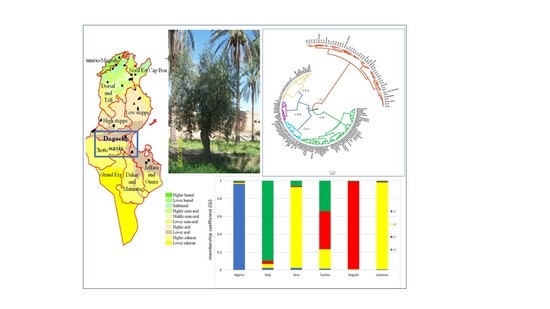
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. DNA Extraction and SSR Assays
2.3. Authentication of Degache genotypes and Relationships with Other Mediterranean germplasm
2.4. Data Analysis
2.4.1. Genetic Diversity
2.4.2. Genetic Relationships and Population Structure
3. Results
3.1. Genetic Diversity of Degache Genotypes
3.2. Genetic Relationships of Degache Genotypes with Other Mediterranean Germplasm
3.3. Genetic Structure
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tunisian Agriculture Ministry-General Direction of Agricultural Production. Available online: http://www.agriculture.tn (accessed on 20 February 2020).
- Wolff, S. Oleoculture and Olive Oil Presses in Phoenician North Africa. In Olive Oil in Antiquity; Eitam, D., Heltzer, M., Eds.; Sargon: Padova, Italy, 1996; pp. 129–136. [Google Scholar]
- Ben Mohamed, M.; Zelasco, S.; Ben Ali, S.; Guasmi, F.; Triki, T.; Conforti, F.L.; Kamoun Naziha, G. Exploring olive trees genetic variability in the South East of Tunisia. Genet. Mol. Res. 2017, 16, gmr16039850. [Google Scholar] [CrossRef]
- Trigui, A.; Msallem, M. Oliviers de Tunisie: Catalogue des Variétés Autochtones and Types Locaux: Identification Variétale and Caractérisation Morpho-Pomologique des Ressources Génétiques Oléicoles de Tunisie; Institution de la Recherche et de l’Enseignement Supérieur Agricoles: Tunis, Tunisia, 2002; Volume 1, p. 159. [Google Scholar]
- Ben Mohamed, M.; Ben Ali, S.; Boussora, F.; Guasmi, F.; Triki, T. Polymorphism of microsatellite (SSR) Markers in tunisian olive (Olea Europaea L.) cultivars. J. Multidisc. Eng. Sci. Stud. 2017, 3, 1247–1252. [Google Scholar]
- Ben Abdelaali, S.; Saddoud Debbabi, O.; Ben Abdelaali, N.; Hajlaoui, M.R.; Mars, M. Fingerprinting of on-farm conserved local Tunisian orange cultivars (Citrus sinensis (L.) osbeck) using microsatellite markers. Acta Biol. Cracov. Bot. 2018, 60, 83–93. [Google Scholar]
- Saddoud Debbabi, O.M.M.; Miazzi, O.; Elloumi, M.; Fendri, F.; Ben Amar, M.; Savoia, S.; Sion, H.; Souabni, S.; Rahmani Mnasri, S. Recovery, assessment, and molecular characterization of minor olive genotypes in Tunisia. Plants 2020, 9, 382. [Google Scholar] [CrossRef] [PubMed]
- Boucheffa, S.; Tamendjari, A.; Sanchez-Gimeno, A.C.; Rovellini, P.; Venturini, S.; di Rienzo, V.; Miazzi, M.M.; Montemurro, C. Diversity Assessment of Algerian Wild and cultivated Olives (Olea europaea L.) by molecular; morphological and chemical traits. Eur. J. Lipid Sci. Technol. 2019, 121, 1800302. [Google Scholar] [CrossRef]
- Di Rienzo, V.; Miazzi, M.M.; Fanelli, V.; Sabetta, W.; Montemurro, C. The preservation and characterization of Apulian olive germplasm biodiversity. Acta Hort. 2018, 1199, 1–6. [Google Scholar] [CrossRef]
- Sion, S.; Taranto, F.; Montemurro, C.; Mangini, G.; Camposeo, S.; Falco, V.; Gallo, A.; Mita, G.; Saddoud Debbabi, O.; Ben Amar, F.; et al. Genetic characterization of apulian olive germplasm as potential source in new breeding programs. Plants 2019, 8, 268. [Google Scholar] [CrossRef]
- Fendri, M.; Trujillo, I.; Trigui, A.; Rodríguez-García, M.I.; Alché Ramírez, J.D. Simple sequence repeat identification and endocarp characterization of olive tree accessions in a Tunisian germplasm collection. Hort. Sci. 2010, 45, 1429–1436. [Google Scholar] [CrossRef]
- Miazzi, M.M.; D’Agostino, N.; Gadaleta, S.; Montemurro, C.; Taranto, F. Genotyping-by-sequencing-derived single-nucleotide polymorphism catalog from a grapevine (Vitis vinifera L.) germplasm collection that includes the most representative Apulian autochthonous cultivars. Acta Hortic. 2019, 1248, 69–75. [Google Scholar] [CrossRef]
- Mnasri, S.; Saddoud Debbabi, O.; Ben Salah, M.; Ferchichi, A. Morphological and molecular characterization of the main varieties cultivated in the region of Hbebsa (North West of Tunisia). Int. J. Agron. Agric. Res. 2014, 5, 87–93. [Google Scholar]
- Laabidi, I.; Gouta, H.; Mezghani, A.; Ayachi, M.; Labidi, F.; Mars, M. Combination of morphological and molecular markers for the characterization of ancient native olive accessions in Central-Eastern Tunisia. Comptes Rendus Biol. 2017, 340, 287–297. [Google Scholar]
- Taamalli, W.; Geuna, F.; Banfi, R.; Bassi, D.; Daoud, D.; Zarrouk, M. Agronomic and molecular analyses for the characterisation of accessions in Tunisian olive germplasm collections. Electron. J. Biotechnol. 2006, 9, 467–481. [Google Scholar] [CrossRef]
- Ben Maachia, S.; Ben Amar, F. Prospection, morphological and Pomological identification of olive ecotypes (Olea europaea L.) in the Degache oasis (Tozeur, Tunisia). J. New Sci. Agric. Biotechnol. 2016, 33, 1892–1900. [Google Scholar]
- Muzzalupo, I.; Vendramin, G.G.; Chiappetta, A. Genetic biodiversity of Italian olives (Olea europaea) germplasm analyzed by SSR markers. Sci. World J. 2014, 296590. [Google Scholar] [CrossRef] [PubMed]
- Pasqualone, A.; di Rienzo, V.D.; Miazzi, M.M.; Fanelli, V.; Caponio, F.; Montemurro, C. High resolution melting analysis of DNA microsatellites in olive pastes and virgin olive oils obtained by talc addition. Eur. J. Lip. Sci. Technol. 2015, 117, 2044–2048. [Google Scholar] [CrossRef]
- Di Rienzo, V.; Fanelli, V.; Miazzi, M.M.; Sabetta, W.; Montemurro, C. A reliable analytical procedure to discover table grape DNA adulteration in industrial wines and musts. Acta Hortic. 2017, 1188, 365–370. [Google Scholar] [CrossRef]
- Baldoni, L.; Cultrera, N.G.; Mariotti, R.; Ricciolini, C.; Arcioni, S.; Vendramin, G.G. A consensus list of microsatellite markers for olive genotyping. Mol. Breed. 2009, 24, 213–231. [Google Scholar] [CrossRef]
- Salimonti, A.; Simone, V.; Cesari, G.; Lamaj, F.; Cattivelli, L.; Perri, E.; Desiderio, F.; Fanizzi, F.P.; Del Coco, L.; Zelasco, S. A first molecular investigation of monumental olive trees in Apulia region. Sci. Hort. 2013, 162, 204–212. [Google Scholar] [CrossRef]
- Sabetta, W.; Miazzi, M.M.; di Rienzo, V.; Fanelli, V.; Pasqualone, A.; Montemurro, C. Development and application of protocols to certify the authenticity and the traceability of Apulian typical products in olive sector. Riv. Ital. Delle Sostanze Grasse 2017, 94, 37–43. [Google Scholar]
- Spadoni, A.; Sion, S.; Gadaleta, S.; Savoia, M.; Piarulli, L.; Fanelli, V.; di Rienzo, V.; Taranto, D.; Miazzi, M.; Montemurro, C.; et al. Simple and rapid method for genomic DNA extraction and microsatellite analysis in tree plants. J. Agric. Sci. Technol. 2019, 21, 1215–1226. [Google Scholar]
- Carriero, F.; Fontanazza, G.; Cellini, F.; Giorio, G. Identification of simple sequence repeats (SSRs) in olive (Olea europaea L.). Theor. Appl. Genet. 2002, 104, 301–307. [Google Scholar] [CrossRef]
- Cipriani, G.; Marrazzo, M.T.; Marconi, R.; Cimato, A.; Testolin, R. Microsatellite markers isolated in olive (Olea europaea L.) are suitable for individual finger-printing and reveal polymorphism within ancient cultivars. Theor. Appl. Genet. 2002, 104, 223–228. [Google Scholar] [CrossRef]
- De la Rosa, R.; James, C.M.; Tobutt, K.R. Isolation and characterization of polymorphic microsatellites in olive (Olea europaea L.) and their transferability to other genera in the Oleaceae. Mol. Ecol. Notes 2002, 2, 265–267. [Google Scholar] [CrossRef]
- Sefc, K.M.; Lopes, M.S.; Mendonica, D.; Rodrigues Dos Santos, M.; Laimer Da Camara Machado, M.; Da Camara Machado, A. Identification of microsatellite loci in olive (Olea europaea L.) and their characterization in Italian and Iberian olive trees. Mol. Ecol. Notes 2000, 9, 1171–1173. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenALEx 6.5: Genetic analysis in excel. Population genetic software for teaching and research an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Wright, S. The genetical structure of populations. Ann. Eugen. 1949, 15, 323–354. [Google Scholar] [CrossRef]
- Kalinowski, S.T. Counting alleles with rarefaction: Private alleles and hierarchical sampling designs. Conserv. Genet. 2004, 5, 539–543. [Google Scholar] [CrossRef]
- Waits, L.P.; Luikart, G.; Taberlet, P. Estimating the probability of identity among genotypes in natural populations: Cautions and guidelines. Mol. Ecol. 2001, 10, 249–256. [Google Scholar] [CrossRef]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Gen. 1980, 32, 314–331. [Google Scholar]
- Kalinowski, S.T.; Taper, M.L.; Marshall, T.C. Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment. Mol. Ecol. 2007, 16, 1099–1106. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar]
- Earl, D.A.; Von Holdt, B.M. Structure Harvester: A website and program for visualizing Structure output and implementing the Evanno method. Conserv. Gen. Res. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software Structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Faostat. 2019. Available online: http://www.fao.org/faostat/en/?#data/QC (accessed on 20 December 2019).
- Dridi, J.; Fendri, M.; Breton, C.M.; Msallem, M. Characterization of olive progenies derived from a Tunisian breeding program by morphological traits and SSR markers. Sci. Hort. 2018, 236, 127–136. [Google Scholar] [CrossRef]
- Taranto, F.; D’Agostino, N.; Fanelli, V.; di Rienzo, V.; Miazzi, M.M.; Pavan, S.; Zelasco, S.; Perri, E.; Montemurro, C. SNP diversity in an olive germplasm collection. Acta Hortic. 2018, 1199, 27–32. [Google Scholar] [CrossRef]
- Miazzi, M.M.; di Rienzo, V.; Mascio, I.; Montemurro, C.; Sion, S.; Sabetta, W.; Vivaldi, G.A.; Camposeo, S.; Caponio, F.; Squeo, G.; et al. Re.Ger.O.P.: An Integrated Project for the Recovery of Ancient and Rare Olive Germplasm. Front. Plant Sci. 2019, 11. [Google Scholar] [CrossRef]
- Tanasijevica, M.; Todorovica, L.S.; Pereira, C.; Pizzigalli, P.; Lionello, S. Impacts of climate change on olive crop evapotranspiration and irrigation requirements in the Mediterranean region. Agric. Water Manag. 2014, 144, 54–68. [Google Scholar] [CrossRef]
- Fraga, H.; Pinto, J.G.; Viola, F.; Santos, J.A. Climate change projections for olive yields in the Mediterranean Basin. Int. J. Climatol. 2019, 40, 769–781. [Google Scholar] [CrossRef]
- Cramer, W.; Guiot, J.; Fader, M.; Garrabou, J.; Gattuso, J.-P.; Iglesias, A.; Lange, M.A.; Lionello, P.; Llasat, M.C.; Paz, S.; et al. Climate change and interconnected risks to sustainable development in the Mediterranean. Nat. Clim. Chang. 2018, 8, 972–980. [Google Scholar] [CrossRef]
- Balloux, F.; Lehmann, L.; de Meeus, T. The population genetics of clonal and partially clonal diploids. Genetics 2003, 164, 1635–1644. [Google Scholar]
- Stoeckel, S.; Grange, J.; Fernández-Manjarres, J.F.; Bilger, I.; Frascaria-Lacoste, N.; Mariette, S. Heterozygote excess in a self-incompatible and partially clonal forest tree species—Prunus avium L. Mol. Ecol. 2006. [Google Scholar] [CrossRef]
- Díez, C.M.; Barrio, I.; Belaj, A.; Barranco, D.; Rallo, L. Centennial olive trees as a reservoir of genetic diversity. Ann. Bot. 2011, 108, 797–807. [Google Scholar] [CrossRef]
- Martí i Fernández, A.; Forcada i Font, C.; Rubio-Cabetas, M.J.; Socias i Company, R. Genetic relationships and population structure of local olive tree accessions from Northeastern Spain revealed by SSR markers. Acta Physiol. Plant 2015, 37, 1726–1732. [Google Scholar] [CrossRef]
- Abdelhamid, S.; Gouta, H.; Gharsallaoui, M.; Ghrab, M. A Review on Current Status of Olive and Olive Oil Production in Tunisia. J. Korean Soc. Int. Agric. 2013, 25, 351–357. [Google Scholar] [CrossRef]
- Chalak, L.; Haouane, H.; Essalouh, L.; Santoni, S.; Besnard, G.; Khadari, B. Extent of the genetic diversity in Lebanese olive (Olea europaea L.) trees: A mixture of an ancient germplasm with recently introduced varieties. Gen. Res. Crop Evol. 2015, 62, 621–633. [Google Scholar] [CrossRef]
- Boucheffa, S.; Miazzi, M.M.; di Rienzo, V.; Mangini, G.; Fanelli, V.; Tamendjari, A.; Pignone, D.; Montemurro, C. The coexistence of oleaster and traditional varieties affects genetic diversity and population structure in Algerian olive (Olea europaea) germplasm. Gen. Res. Crop Evol. 2017, 64, 379–390. [Google Scholar] [CrossRef]
- Di Rienzo, V.; Sion, S.; Taranto, F.; Zammit-Mangion, M.; Miazzi, M. Genetic flow among olive populations within the Mediterranean basin. PeerJ 2018, 18, e5260. [Google Scholar] [CrossRef]
- Hannachi, H.; Breton, C.; Msallem, M.; Ben El Hadj, S.; El Gazzzah, M.; Bervillé, A. Genetic relationships between cultivated and wild olive trees (Olea europaea L. var. europaea and var. sylvestris) based on nuclear and Chloroplast SSR markers. Nat. Res. 2010, 1, 95–103. [Google Scholar] [CrossRef]
- Haddad, B.; Gristina, A.S.; Mercati, F.; Saadi, A.E.; Aiter, N.; Martorana, A.; Sharaf, A.; Carimi, F. Molecular analysis of the official algerian olive collection highlighted a hotspot of biodiversity in the central mediterranean basin. Genes 2020, 11, 303. [Google Scholar] [CrossRef] [PubMed]
- Anestiadou, K.; Nikoloudakis, N. Monumental olive trees of Cyprus contributed to the establishment of the contemporary olive germplasm. PLoS ONE 2017, 12, e0187697. [Google Scholar] [CrossRef]
- Ben Amar, F.; Ben Maachia, S.; Yengui, A. Catalogue des Ressources Génétiques de 431 L’Olivier Dans L’Oasis de Degache (Tunisie); FAO and Ministry of Agriculture of Tunisia: Rome, Italy, 2017. [Google Scholar]
- Grati-Kamoun, N.; Lamy Mahmoud, F.; Rebai, A.; Gargouri, A.; Panaud, O.; Saar, A. Genetic diversity of Tunisian olive tree (Olea europaea L.) cultivars assessed by AFLP markers. Gen. Res. Crop Evol. 2006, 53, 265–275. [Google Scholar] [CrossRef]
- Trujillo, I.; Ojeda, M.A.; Urdiroz, N.M.; Potter, D.; Barranco, D.; Rallo, L. Identification of the Worldwide Olive Germplasm Bank of Córdoba (Spain) using SSR and morphological markers. Tree Genet Genomes 2014, 10, 141–155. [Google Scholar] [CrossRef]
- Diez, C.M.; Trujillo, I.; Martinez-Urdiroz, N.; Barranco, D.; Rallo, L.; Marfil, P. Olive domestication and diversification in the Mediterranean Basin. New Phytol. 2015, 206, 436–447. [Google Scholar] [CrossRef]
- Abdelhamid, S.; Omri, A.; Grati-Kamoun, N.; Marra, F.P.; Caruso, T. Molecular characterization and genetic relationships of cultivated Tunisian olive varieties (Olea europaea L.) using SSR markers. J. New Sci. Agric. Biotechnol. 2017, 40, 2175–2185. [Google Scholar]
- Haouane, H.; El Bakkali, A.; Moukhli, A.; Tollon, C.; Santoni, S.; Oukabli, A. Genetic structure and core collection of the World Olive Germplasm Bank of Marrakech: Towards the optimized management and use of Mediterranean olive genetic resources. Genetica 2011, 139, 1083–1094. [Google Scholar] [CrossRef]
- Rugini, E.; Baldoni, L.; Muleo, R.; Sebastiani, L. The Olive Tree Genome. In Compendium of Plant Genomes; Rugini, E., Baldoni, L., Muleo, R., Sebastiani, L., Eds.; Springer: Basel, Switzerland, 2017; pp. 27–54. [Google Scholar] [CrossRef]
- Besnard, G.; Terral, J.F.; Cornille, A. On the origins and domestication of the olive: A review and perspectives. Ann. Bot. 2018, 121, 385–403. [Google Scholar] [CrossRef]
- Besnard, G.; El Bakkali, A.; Haouane, H.; Baali-Cherif, D.; Moukhli, A.; Khadari, B. Population genetics of Mediterranean and Saharan olives: Geographic patterns of differentiation and evidence for early-generations of admixture. Ann. Bot. 2013, 112, 1293–1302. [Google Scholar] [CrossRef]
- Newton, C.; Lorre, C.; Sauvage, C.; Ivorra, S.; Terral, J.-F. On the origins and spread of Olea europaea L. (olive) domestication: Evidence for shape variation of olive stones at Ugarit, Late Bronze Age, Syria—A window on the Mediterranean Basin and on the westward diffusion of olive varieties. Veg. Hist. Archaeobotany 2013, 23, 567–575. [Google Scholar] [CrossRef]
- D’Agostino, N.; Taranto, F.; Camposeo, S.; Mangini, G.; Fanelli, V.; Gadaleta, S.; Miazzi, M.M.; Pavan, S.; Di Rienzo, V.; Sabetta, W.; et al. GBS-derived SNP catalogue unveiled wide genetic variability and geographical relationships of Italian olive cultivars. Sci. Rep. 2018, 8, 15877. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | Alleles Length (bp) | Na | Ne | I | Ho | He | F | PIC | F Null |
|---|---|---|---|---|---|---|---|---|---|
| DCA03 | 231-243-249 | 3 | 2.82 | 1.06 | 0.88 | 0.64 | −0.37 | 0.57 | −0.155 |
| DCA05 | 194-206 | 2 | 2.00 | 0.69 | 1.00 | 0.50 | −1.00 | 0.37 | −0.333 |
| DCA09 | 162-166-172-194-204-206 | 6 | 4.11 | 1.52 | 0.91 | 0.75 | −0.21 | 0.71 | −0.102 |
| DCA15 | 246-266-270 | 3 | 1.74 | 0.71 | 0.56 | 0.42 | −0.33 | 0.36 | −0.157 |
| DCA16 | 122-124-136-144-148-172-174-176 | 8 | 4.03 | 1.64 | 0.94 | 0.75 | −0.25 | 0.72 | −0.132 |
| DCA17 | 113-115-141-143 | 4 | 2.24 | 0.91 | 0.37 | 0.55 | 0.31 | 0.46 | 0.201 |
| DCA18 | 173-177-179-183 | 4 | 2.64 | 1.08 | 0.82 | 0.62 | −0.33 | 0.54 | −0.147 |
| UDO43 | 172-174-184-210-212 | 5 | 3.29 | 1.35 | 0.89 | 0.69 | −0.28 | 0.65 | −0.131 |
| GAPU101 | 182-192-200-206 | 4 | 2.58 | 1.06 | 0.89 | 0.61 | −0.45 | 0.54 | −0.209 |
| EMOL | 190-192-198 | 3 | 1.87 | 0.77 | 0.43 | 0.46 | 0.07 | 0.39 | 0.023 |
| Total | 42 | ||||||||
| Mean | 4.2 | 2.73 | 1.08 | 0.77 | 0.60 | −0.28 | 0.536 | −0.115 |
| Collection | Na | Ne | I | Ho | He | F | Private Alleles | |
|---|---|---|---|---|---|---|---|---|
| Degache (37) | Total | 42 | 27.3 | |||||
| Mean | 4.2 | 2.74 | 1.085 | 0.774 | 0.604 | −0.285 | 2 | |
| Tunisia (37) | Total | 91 | 48.9 | |||||
| Mean | 9.1 | 4.88 | 1.688 | 0.700 | 0.738 | 0.074 | 15 | |
| Algeria (20) | Total | 76.0 | 41.0 | |||||
| Mean | 7.6 | 4.10 | 1.574 | 0.661 | 0.718 | 0.122 | 13 | |
| Italia (43) | Total | 105.0 | 51.5 | |||||
| Mean | 10.5 | 5.15 | 1.826 | 0.740 | 0.775 | 0.043 | 16 | |
| Syria (17) | Total | 64.0 | 37.7 | |||||
| Mean | 6.4 | 3.77 | 1.407 | 0.720 | 0.653 | −0.095 | 11 | |
| Lebanon (23) | Total | 44 | 24.0 | |||||
| Mean | 4.4 | 2.40 | 0.979 | 0.721 | 0.522 | −0.321 | 4 | |
| Mean | 7.03 | 6.30 | 1.41 | 0.71 | 0.66 | −0.07 | 10 |
| Source | df | SS | MS | Est. Var. | % | p Values |
|---|---|---|---|---|---|---|
| Among groups | 5 | 187.98 | 37.59 | 0.59 | 14% | <0.001 |
| Within groups | 177 | 633.50 | 3.57 | 3.57 | 86% | |
| Total | 353 | 1403.28 | 4.17 | 100% |
| Geographic Origin | Algeria | Italy | Syria | Tunisia | Degache | Lebanon |
|---|---|---|---|---|---|---|
| Algeria | 0 | 0.142 | 0.165 | 0.143 | 0.251 | 0.247 |
| Italy | 0.000 | 0.117 | 0.055 | 0.136 | 0.165 | |
| Syria | 0.000 | 0.118 | 0.223 | 0.212 | ||
| Tunisia | 0.000 | 0.062 | 0.137 | |||
| Degache | 0.000 | 0.228 | ||||
| Lebanon | 0.000 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saddoud Deddabi, O.; Montemurro, C.; Ben Maachia, S.; Ben Amar, F.; Fanelli, V.; Gadaleta, S.; El Riachy, M.; Chehade, A.; Siblini, M.; Boucheffa, S.; et al. A Hot Spot of Olive Biodiversity in the Tunisian Oasis of Degache. Diversity 2020, 12, 358. https://doi.org/10.3390/d12090358
Saddoud Deddabi O, Montemurro C, Ben Maachia S, Ben Amar F, Fanelli V, Gadaleta S, El Riachy M, Chehade A, Siblini M, Boucheffa S, et al. A Hot Spot of Olive Biodiversity in the Tunisian Oasis of Degache. Diversity. 2020; 12(9):358. https://doi.org/10.3390/d12090358
Chicago/Turabian StyleSaddoud Deddabi, Olfa, Cinzia Montemurro, Sihem Ben Maachia, Fathi Ben Amar, Valentina Fanelli, Susanna Gadaleta, Milad El Riachy, Ali Chehade, Mona Siblini, Saliha Boucheffa, and et al. 2020. "A Hot Spot of Olive Biodiversity in the Tunisian Oasis of Degache" Diversity 12, no. 9: 358. https://doi.org/10.3390/d12090358
APA StyleSaddoud Deddabi, O., Montemurro, C., Ben Maachia, S., Ben Amar, F., Fanelli, V., Gadaleta, S., El Riachy, M., Chehade, A., Siblini, M., Boucheffa, S., & Miazzi, M. M. (2020). A Hot Spot of Olive Biodiversity in the Tunisian Oasis of Degache. Diversity, 12(9), 358. https://doi.org/10.3390/d12090358