Urbanization and Human Population Favor Species Richness of Alien Birds

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Avian Data
2.2. Land-Use and Human-Population Data
2.3. Statistical Analysis
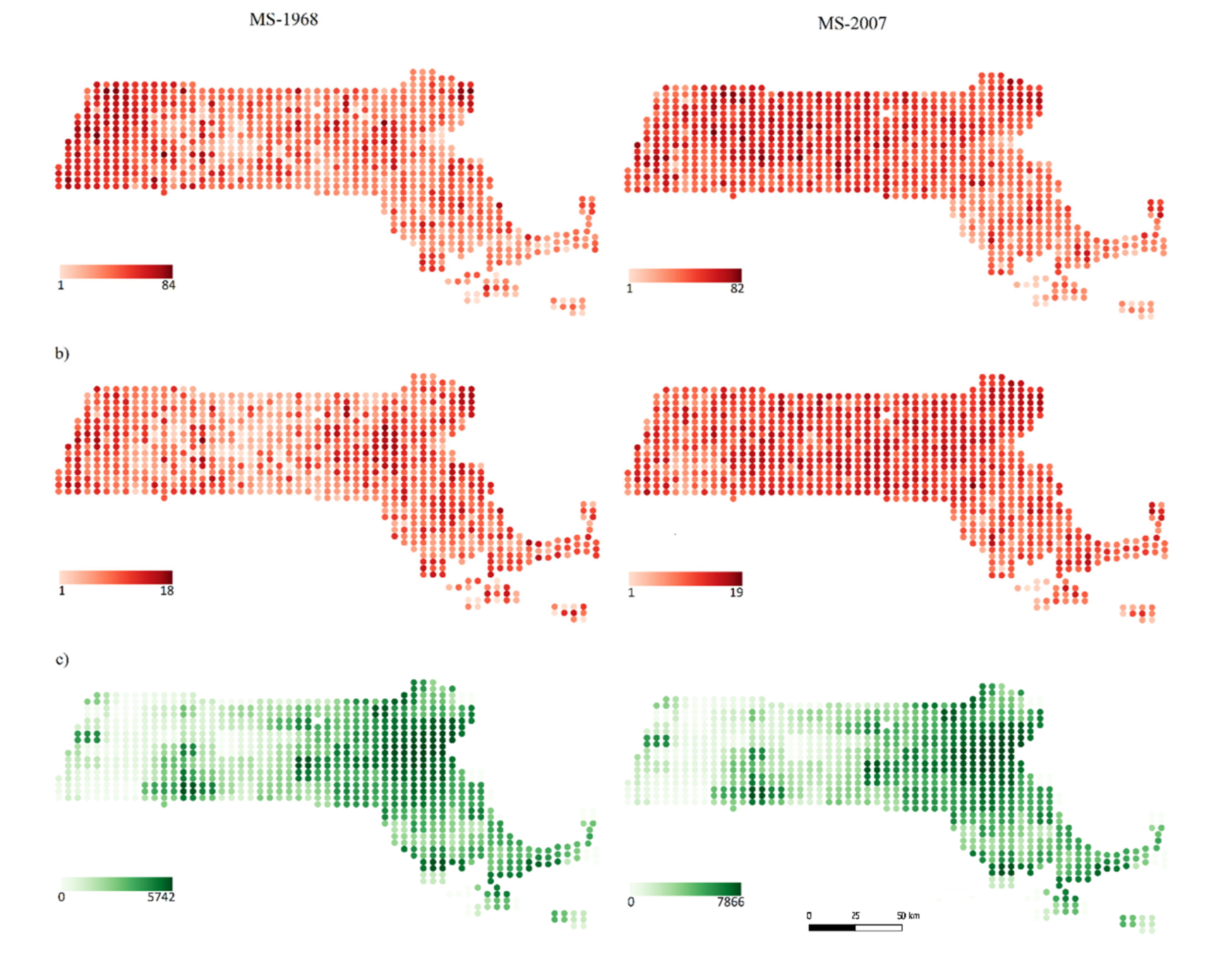
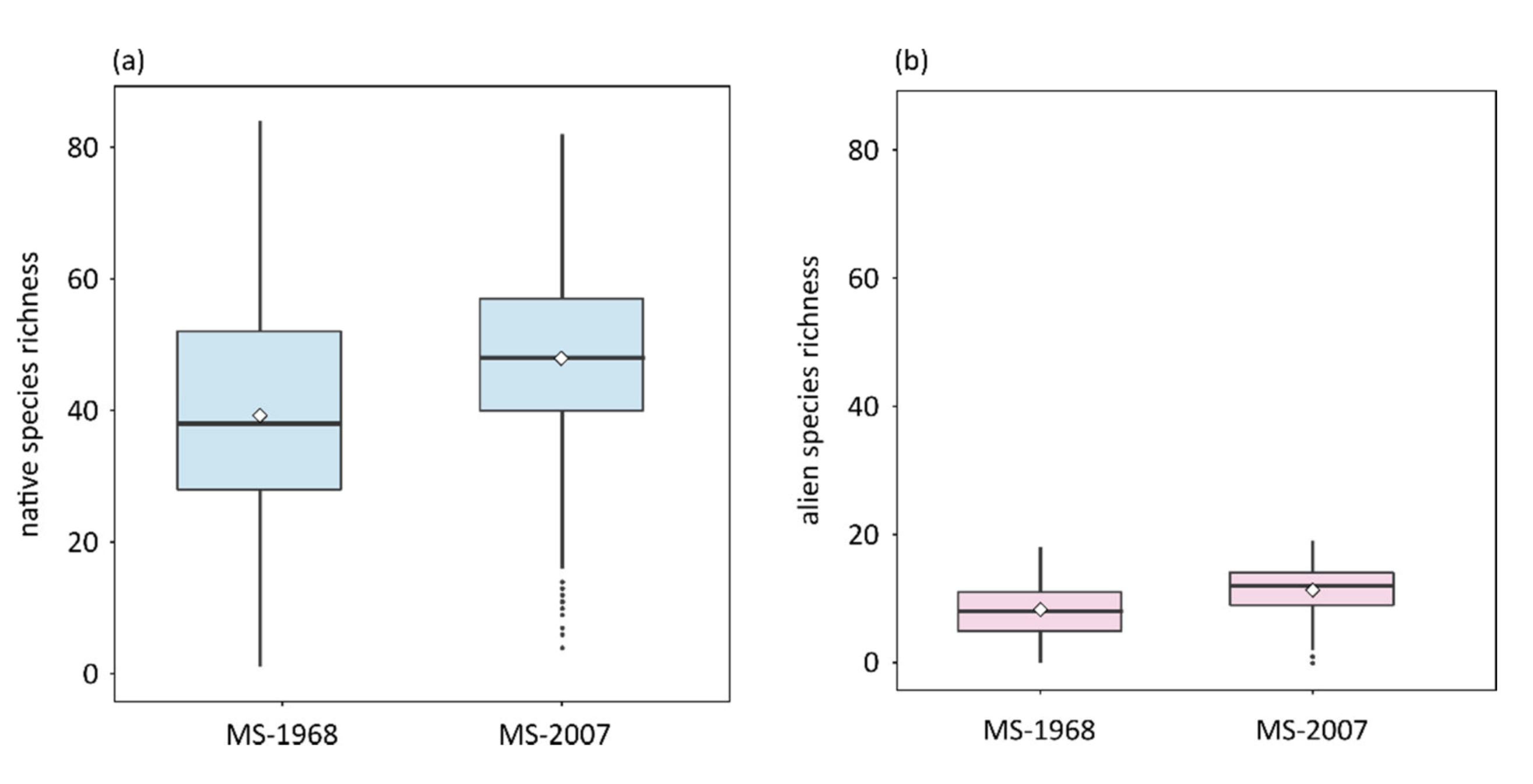
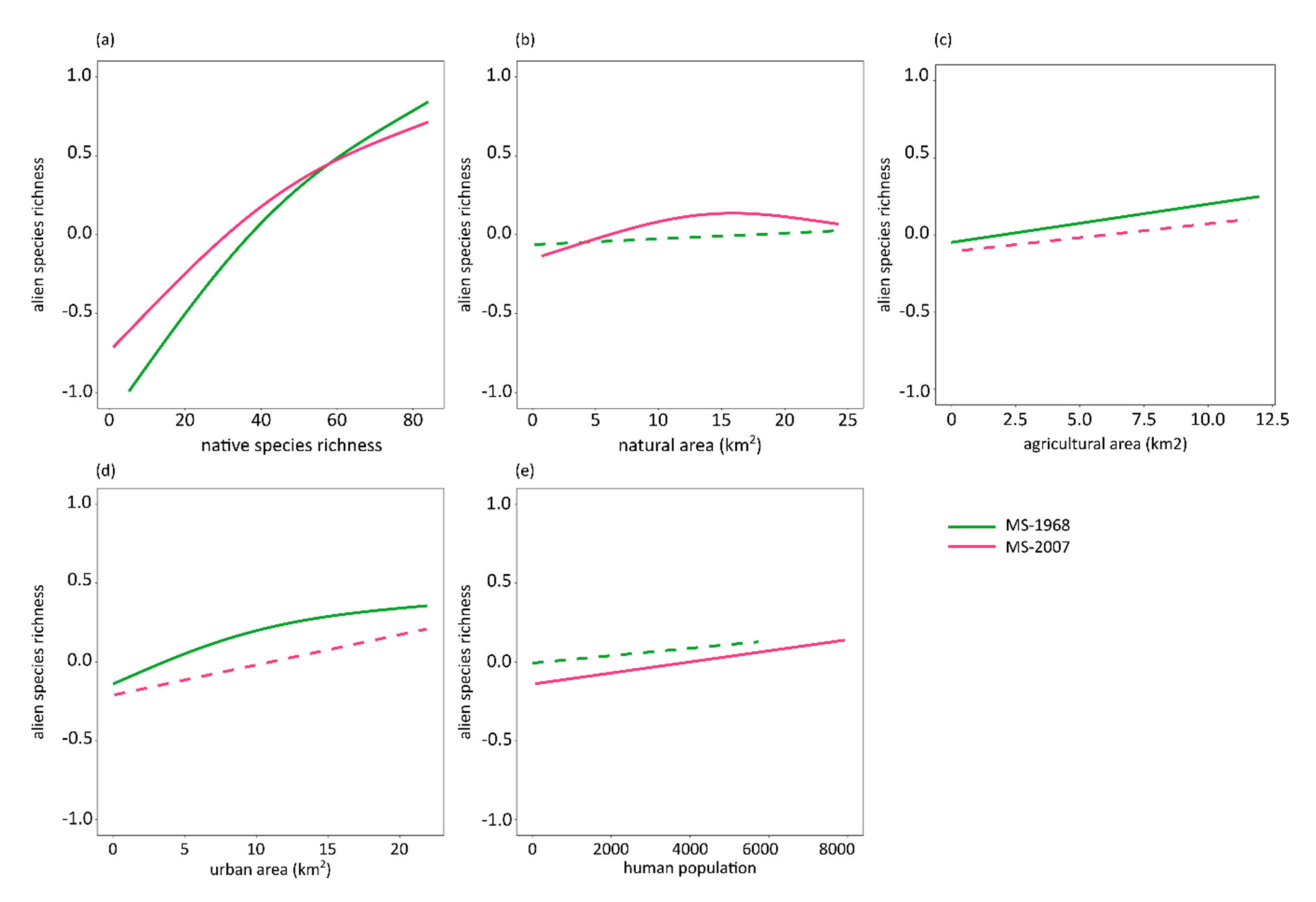
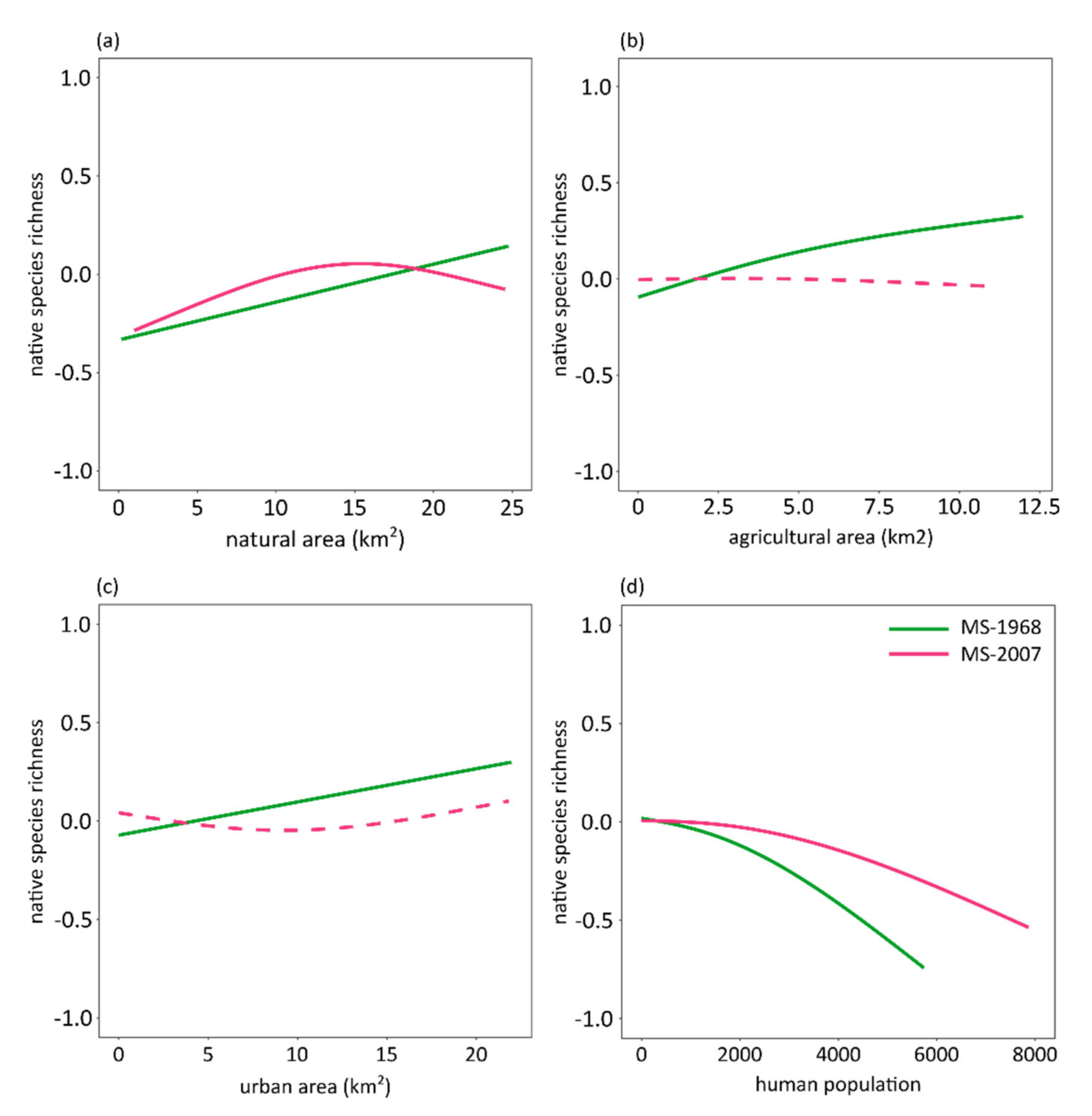
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lewis, S.L.; Maslin, M.A. Defining the anthropocene. Nature 2015, 519, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Hulme, P.E. Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. Appl. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Banks, N.C.; Paini, D.R.; Bayliss, K.L.; Hodda, M. The role of global trade and transport network topology in the human-mediated dispersal of alien species. Ecol. Lett. 2015, 18, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; van Kleunen, M.; Winter, M. Global rise in emerging alien species results from increased accessibility of new source pools. Proc. Natl. Acad. Sci. USA 2018, 115, E2264–E2273. [Google Scholar] [CrossRef]
- Blackburn, T.M.; Gaston, K.J. Abundance, biomass and energy use of native and alien breeding birds in Britain. Biol. Invasions 2018, 20, 3563–3573. [Google Scholar] [CrossRef]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; Winter, M.; Arianoutsou, M. No saturation in the accumulation of alien species worldwide. Nat. Commun. 2017, 8, 14435. [Google Scholar] [CrossRef]
- Dyer, E.E.; Cassey, P.; Redding, D.W.; Collen, B.; Franks, V.; Gaston, K.J.; Jones, K.E.; Kark, S.; Orme, C.D.L.; Blackburn, T.M. The global distribution and drivers of alien bird species richness. PLoS Biol. 2017, 15, e2000942. [Google Scholar] [CrossRef]
- Vilà, M.; Basnou, C.; Pyšek, P.; Josefsson, M.; Genovesi, P.; Gollasch, S.; Nentwig, W.; Olenin, S.; Roques, A.; Roy, D. How well do we understand the impacts of alien species on ecosystem services? A pan-European, cross-taxa assessment. Front. Ecol. Environ. 2010, 8, 135–144. [Google Scholar] [CrossRef]
- Vilà, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarošík, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pyšek, P. Ecological impacts of invasive alien plants: A meta-analysis of their effects on species, communities and ecosystems. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [CrossRef]
- Simberloff, D.; Martin, J.-L.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; García-Berthou, E.; Pascal, M. Impacts of biological invasions: What’s what and the way forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef]
- Duncan, R.P.; Cassey, P.; Pigot, A.L.; Blackburn, T.M. A general model for alien species richness. Biol. Invasions 2019, 21, 2665–2677. [Google Scholar] [CrossRef]
- Davies, R.G.; Orme, C.D.L.; Olson, V.; Thomas, G.H.; Ross, S.G.; Ding, T.-S.; Rasmussen, P.C.; Stattersfield, A.J.; Bennett, P.M.; Blackburn, T.M. Human impacts and the global distribution of extinction risk. Proc. R. Soc. B Biol. Sci. 2006, 273, 2127–2133. [Google Scholar] [CrossRef]
- Lockwood, J.L.; Hoopes, M.F.; Marchetti, M.P. Invasion Ecology; John Wiley and Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Leprieur, F.; Beauchard, O.; Blanchet, S.; Oberdorff, T.; Brosse, S. Fish invasions in the world’s river systems: When natural processes are blurred by human activities. PLoS Biol. 2008, 6, e28. [Google Scholar]
- Pyšek, P.; Bacher, S.; Chytrý, M.; Jarošík, V.; Wild, J.; Celesti-Grapow, L.; Gassó, N.; Kenis, M.; Lambdon, P.W.; Nentwig, W. Contrasting patterns in the invasions of European terrestrial and freshwater habitats by alien plants, insects and vertebrates. Glob. Ecol. Biogeogr. 2010, 19, 317–331. [Google Scholar] [CrossRef]
- Chiron, F.; Shirley, S.; Kark, S. Human-related processes drive the richness of exotic birds in Europe. Proc. R. Soc. B Biol. Sci. 2008, 276, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Aronson, M.F.; Handel, S.N.; La Puma, I.P.; Clemants, S.E. Urbanization promotes non-native woody species and diverse plant assemblages in the New York metropolitan region. Urban Ecosyst. 2015, 18, 31–45. [Google Scholar] [CrossRef]
- Kühn, I.; Klotz, S. Urbanization and homogenization–comparing the floras of urban and rural areas in Germany. Biol. Conserv. 2006, 127, 292–300. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Yasui, S.L.E.; Livingstone, S.; MacIvor, J.S. Are urban systems beneficial, detrimental, or indifferent for biological invasion? Biol. Invasions 2017, 19, 3489–3503. [Google Scholar] [CrossRef]
- Maestas, J.D.; Knight, R.L.; Gilgert, W.C. Biodiversity across a rural land-use gradient. Conserv. Biol. 2003, 17, 1425–1434. [Google Scholar] [CrossRef]
- Clergeau, P.; Savard, J.-P.L.; Mennechez, G.; Falardeau, G. Bird abundance and diversity along an urban-rural gradient: A comparative study between two cities on different continents. Condor 1998, 100, 413–425. [Google Scholar] [CrossRef]
- Fernandez-Juricic, E. Avian spatial segregation at edges and interiors of urban parks in Madrid, Spain. Biodivers. Conserv. 2001, 10, 1303–1316. [Google Scholar] [CrossRef]
- McKinney, M.L. Urbanization as a major cause of biotic homogenization. Biol. Conserv. 2006, 127, 247–260. [Google Scholar] [CrossRef]
- Kark, S.; Allnutt, T.F.; Levin, N.; Manne, L.L.; Williams, P.H. The role of transitional areas as avian biodiversity centres. Glob. Ecol. Biogeogr. 2007, 16, 187–196. [Google Scholar] [CrossRef]
- Shea, K.; Chesson, P. Community ecology theory as a framework for biological invasions. Trends Ecol. Evol. 2002, 17, 170–176. [Google Scholar] [CrossRef]
- Sol, D.; Bartomeus, I.; Griffin, A.S. The paradox of invasion in birds: Competitive superiority or ecological opportunism? Oecologia 2012, 169, 553–564. [Google Scholar] [CrossRef]
- McKinney, M.L. Urbanization, Biodiversity, and ConservationThe impacts of urbanization on native species are poorly studied, but educating a highly urbanized human population about these impacts can greatly improve species conservation in all ecosystems. Bioscience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- Sax, D.F.; Brown, J.H. The paradox of invasion. Glob. Ecol. Biol. 2000, 9, 363–371. [Google Scholar] [CrossRef]
- McKinney, M.L. Effects of urbanization on species richness: A review of plants and animals. Urban Ecosyst. 2008, 11, 161–176. [Google Scholar] [CrossRef]
- Traveset, A.; Richardson, D.M. Mutualistic interactions and biological invasions. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 89–113. [Google Scholar] [CrossRef]
- Jauni, M.; Ramula, S. Meta-analysis on the effects of exotic plants on the fitness of native plants. Perspect. Plant Ecol. Evol. Syst. 2015, 17, 412–420. [Google Scholar] [CrossRef]
- Sugiura, S. Impacts of introduced species on the biota of an oceanic archipelago: The relative importance of competitive and trophic interactions. Ecol. Res. 2016, 31, 155–164. [Google Scholar] [CrossRef]
- Fridley, J.D.; Stachowicz, J.; Naeem, S.; Sax, D.; Seabloom, E.; Smith, M.; Stohlgren, T.; Tilman, D.; Holle, B.V. The invasion paradox: Reconciling pattern and process in species invasions. Ecology 2007, 88, 3–17. [Google Scholar] [CrossRef]
- Tomasetto, F.; Duncan, R.P.; Hulme, P.E. Resolving the invasion paradox: Pervasive scale and study dependence in the native-alien species richness relationship. Ecol. Lett. 2019, 22, 1038–1046. [Google Scholar] [CrossRef] [PubMed]
- Elton, C.S. The Ecology of Invasions by Plants and Animals; Methuen: London, UK, 1958. [Google Scholar]
- Levine, J.M.; Adler, P.B.; Yelenik, S.G. A meta-analysis of biotic resistance to exotic plant invasions. Ecol. Lett. 2004, 7, 975–989. [Google Scholar] [CrossRef]
- Stohlgren, T.J.; Barnett, D.T.; Kartesz, J.T. The rich get richer: Patterns of plant invasions in the United States. Front. Ecol. Environ. 2003, 1, 11–14. [Google Scholar] [CrossRef]
- Poessel, S.A.; Beard, K.H.; Callahan, C.M.; Ferreira, R.B.; Stevenson, E.T. Biotic acceptance in introduced amphibians and reptiles in Europe and North America. Glob. Ecol. Biogeogr. 2013, 22, 192–201. [Google Scholar] [CrossRef]
- Petersen, W.R.; Meservey, W.R. Massachusetts Breeding Bird Atlas; University of Massachusetts Press: Amherst, MA, USA, 2003. [Google Scholar]
- Kamm, M. The Massachusetts Breeding Bird Atlas 2; Mass Audubon: Topsfield, MA, USA, 2013. [Google Scholar]
- Chesser, R.; Burns, K.; Cicero, C.; Dunn, J.; Kratter, A.; Lovette, I.; Rasmussen, P.; Remsen, J., Jr.; Stotz, D.; Winger, B. Check-List of North American Birds (Online); American Ornithological Society: Chicago, IL, USA, 2018. [Google Scholar]
- Dyer, E.E.; Redding, D.W.; Blackburn, T.M. The global avian invasions atlas, a database of alien bird distributions worldwide. Sci. Data 2017, 4, 170041. [Google Scholar] [CrossRef]
- Price, C.V.; Nakagaki, N.; Hitt, K.J.; Clawges, R. Enhanced Historical Land-Use and Land-Cover Data Sets of the US Geological Survey; US Department of the Interior, US Geological Survey. U.S. Geological Survey digital data series 240. 2007. Available online: http://pubs.usgs.gov/ds/2006/240/ (accessed on 28 September 2019).
- Fry, J.A.; Xian, G.; Jin, S.; Dewitz, J.A.; Homer, C.G.; Yang, L.; Barnes, C.A.; Herold, N.D.; Wickham, J.D. Completion of the 2006 national land cover database for the conterminous United States. PE RS Photogramm. Eng. Remote Sens. 2011, 77, 858–864. [Google Scholar]
- Klein Goldewijk, K.; Beusen, A.; Janssen, P. Long-term dynamic modeling of global population and built-up area in a spatially explicit way: HYDE 3.1. Holocene 2010, 20, 565–573. [Google Scholar] [CrossRef]
- Klein Goldewijk, K.; Beusen, A.; Van Drecht, G.; De Vos, M. The HYDE 3.1 spatially explicit database of human-induced global land-use change over the past 12,000 years. Glob. Ecol. Biogeogr. 2011, 20, 73–86. [Google Scholar] [CrossRef]
- Wheeler, B.; Torchiano, M. lmPerm: Permutation Tests for Linear Models. R Package Version 1.1-2. Available online: http://CRAN.R-project.org/package=lmPerm (accessed on 25 August 2013).
- Hastie, T.; Tibshirani, R. Exploring the nature of covariate effects in the proportional hazards model. Biometrics 1990, 46, 1005–1016. [Google Scholar] [CrossRef]
- Wood, S.N. mgcv: GAMs and generalized ridge regression for R. R News 2001, 1, 20–25. [Google Scholar]
- Chace, J.F.; Walsh, J.J. Urban effects on native avifauna: A review. Landsc. Urban Plan. 2006, 74, 46–69. [Google Scholar] [CrossRef]
- Ibáñez-Álamo, J.D.; Rubio, E.; Benedetti, Y.; Morelli, F. Global loss of avian evolutionary uniqueness in urban areas. Glob. Chang. Biol. 2017, 23, 2990–2998. [Google Scholar] [CrossRef] [PubMed]
- Jeschke, J.M. General hypotheses in invasion ecology. Divers. Distrib. 2014, 20, 1229–1234. [Google Scholar] [CrossRef]
- Siriwardena, G.M.; Stevens, D.K.; Anderson, G.Q.; Vickery, J.A.; Calbrade, N.A.; Dodd, S. The effect of supplementary winter seed food on breeding populations of farmland birds: Evidence from two large-scale experiments. J. Appl. Ecol. 2007, 44, 920–932. [Google Scholar] [CrossRef]
- Siriwardena, G.M.; Baillie, S.R.; Wilson, J.D. Temporal variation in the annual survival rates of six granivorous birds with contrasting population trends. Ibis 1999, 141, 621–636. [Google Scholar] [CrossRef]
- Jones, D. The Birds at My Table: Why We Feed wild Birds and Why it Matters; Cornell University Press: Ithaca, MY, USA, 2018. [Google Scholar]
- Oro, D.; Genovart, M.; Tavecchia, G.; Fowler, M.S.; Martínez-Abraín, A. Ecological and evolutionary implications of food subsidies from humans. Ecol. Lett. 2013, 16, 1501–1514. [Google Scholar] [CrossRef]
- Robb, G.N.; McDonald, R.A.; Chamberlain, D.E.; Bearhop, S. Food for thought: Supplementary feeding as a driver of ecological change in avian populations. Front. Ecol. Environ. 2008, 6, 476–484. [Google Scholar] [CrossRef]
- Galbraith, J.A.; Beggs, J.R.; Jones, D.N.; Stanley, M.C. Supplementary feeding restructures urban bird communities. Proc. Natl. Acad. Sci. USA 2015, 112, E2648–E2657. [Google Scholar] [CrossRef]
- Ciach, M.; Fröhlich, A. Habitat type, food resources, noise and light pollution explain the species composition, abundance and stability of a winter bird assemblage in an urban environment. Urban Ecosyst. 2017, 20, 547–559. [Google Scholar] [CrossRef]
- Plummer, K.E.; Risely, K.; Toms, M.P.; Siriwardena, G.M. The composition of British bird communities is associated with long-term garden bird feeding. Nat. Commun. 2019, 10, 1–8. [Google Scholar] [CrossRef]
- Peng, S.; Kinlock, N.L.; Gurevitch, J.; Peng, S. Correlation of native and exotic species richness: A global meta-analysis finds no invasion paradox across scales. Ecology 2019, 100, e02552. [Google Scholar] [CrossRef]
- Wania, A.; Kühn, I.; Klotz, S. Plant richness patterns in agricultural and urban landscapes in Central Germany—Spatial gradients of species richness. Landsc. Urban Plan. 2006, 75, 97–110. [Google Scholar] [CrossRef]
- Oliveira Hagen, E.; Hagen, O.; Ibáñez-Álamo, J.D.; Petchey, O.L.; Evans, K.L. Impacts of urban areas and their characteristics on avian functional diversity. Front. Ecol. Evol. 2017, 5, 84. [Google Scholar] [CrossRef]
- Faeth, S.H.; Bang, C.; Saari, S. Urban biodiversity: Patterns and mechanisms. Ann. N. Y. Acad. Sci. 2011, 1223, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Daniel Kissling, W.; Böhning-Gaese, K.; Jetz, W. The global distribution of frugivory in birds. Glob. Ecol. Biogeogr. 2009, 18, 150–162. [Google Scholar] [CrossRef]
- Kissling, W.D.; Sekercioglu, C.H.; Jetz, W. Bird dietary guild richness across latitudes, environments and biogeographic regions. Glob. Ecol. Biogeogr. 2012, 21, 328–340. [Google Scholar] [CrossRef]
- Eötvös, C.B.; Magura, T.; Lövei, G.L. A meta-analysis indicates reduced predation pressure with increasing urbanization. Landsc. Urban Plan. 2018, 180, 54–59. [Google Scholar] [CrossRef]
- Kauhala, K.; Talvitie, K.; Vuorisalo, T. Free-ranging house cats in urban and rural areas in the north: Useful rodent killers or harmful bird predators? Folia Zool. 2015, 64, 45–56. [Google Scholar] [CrossRef]
- Geue, D.; Partecke, J. Reduced parasite infestation in urban Eurasian blackbirds (Turdus merula): A factor favoring urbanization? Can. J. Zool. 2008, 86, 1419–1425. [Google Scholar] [CrossRef]
- Bobby Fokidis, H.; Greiner, E.C.; Deviche, P. Interspecific variation in avian blood parasites and haematology associated with urbanization in a desert habitat. J. Avian Biol. 2008, 39, 300–310. [Google Scholar] [CrossRef]
- Calegaro-Marques, C.; Amato, S.B. Urbanization breaks up host-parasite interactions: A case study on parasite community ecology of Rufous-bellied Thrushes (Turdus rufiventris) along a rural-urban gradient. PLoS ONE 2014, 9, e103144. [Google Scholar] [CrossRef] [PubMed]
- Colautti, R.I.; Ricciardi, A.; Grigorovich, I.A.; MacIsaac, H.J. Is invasion success explained by the enemy release hypothesis? Ecol. Lett. 2004, 7, 721–733. [Google Scholar] [CrossRef]
- Heger, T.; Jeschke, J.M. The enemy release hypothesis as a hierarchy of hypotheses. Oikos 2014, 123, 741–750. [Google Scholar] [CrossRef]
- Strohbach, M.W.; Haase, D.; Kabisch, N. Birds and the city: Urban biodiversity, land use, and socioeconomics. Ecol. Soc. 2009, 14, 2. [Google Scholar] [CrossRef]
- Leong, M.; Dunn, R.R.; Trautwein, M.D. Biodiversity and socioeconomics in the city: A review of the luxury effect. Biol. Lett. 2018, 14, 20180082. [Google Scholar] [CrossRef]
- Barnagaud, J.-Y.; Barbaro, L.; Papaix, J.; Deconchat, M.; Brockerhoff, E.G. Habitat filtering by landscape and local forest composition in native and exotic New Zealand birds. Ecology 2014, 95, 78–87. [Google Scholar] [CrossRef]
- Spear, D.; Foxcroft, L.C.; Bezuidenhout, H.; McGeoch, M.A. Human population density explains alien species richness in protected areas. Biol. Conserv. 2013, 159, 137–147. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lazarina, M.; Tsianou, M.A.; Boutsis, G.; Andrikou-Charitidou, A.; Karadimou, E.; Kallimanis, A.S. Urbanization and Human Population Favor Species Richness of Alien Birds. Diversity 2020, 12, 72. https://doi.org/10.3390/d12020072
Lazarina M, Tsianou MA, Boutsis G, Andrikou-Charitidou A, Karadimou E, Kallimanis AS. Urbanization and Human Population Favor Species Richness of Alien Birds. Diversity. 2020; 12(2):72. https://doi.org/10.3390/d12020072
Chicago/Turabian StyleLazarina, Maria, Mariana A. Tsianou, Georgios Boutsis, Aristi Andrikou-Charitidou, Elpida Karadimou, and Athanasios S. Kallimanis. 2020. "Urbanization and Human Population Favor Species Richness of Alien Birds" Diversity 12, no. 2: 72. https://doi.org/10.3390/d12020072
APA StyleLazarina, M., Tsianou, M. A., Boutsis, G., Andrikou-Charitidou, A., Karadimou, E., & Kallimanis, A. S. (2020). Urbanization and Human Population Favor Species Richness of Alien Birds. Diversity, 12(2), 72. https://doi.org/10.3390/d12020072