A Comparative Study of Carabid Beetles in Green Spaces and Former Natural Habitats


Abstract
:1. Introduction
2. Methods
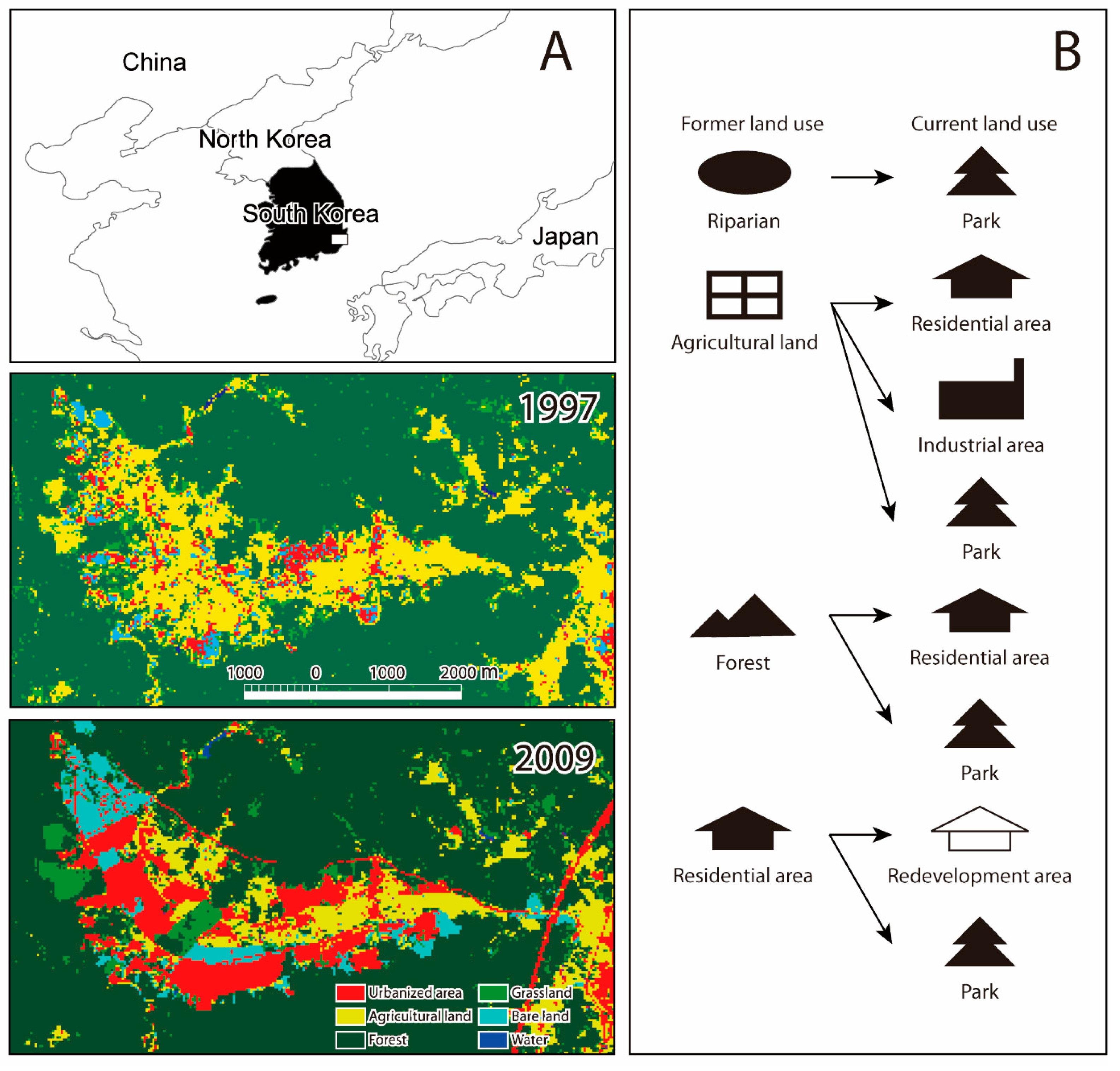
2.1. Former and Current Habitats
2.2. Carabid Sampling
2.3. Data Analysis
3. Results
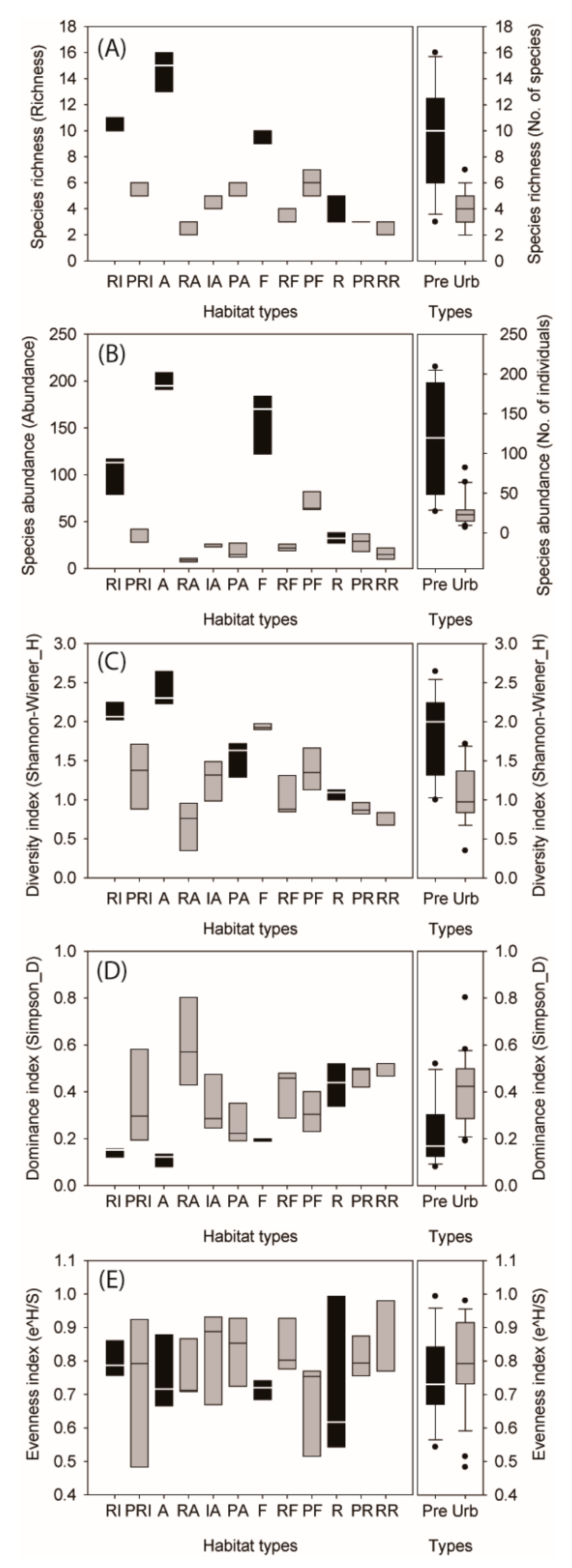
3.1. Carabid Diversity among Urban Habitats
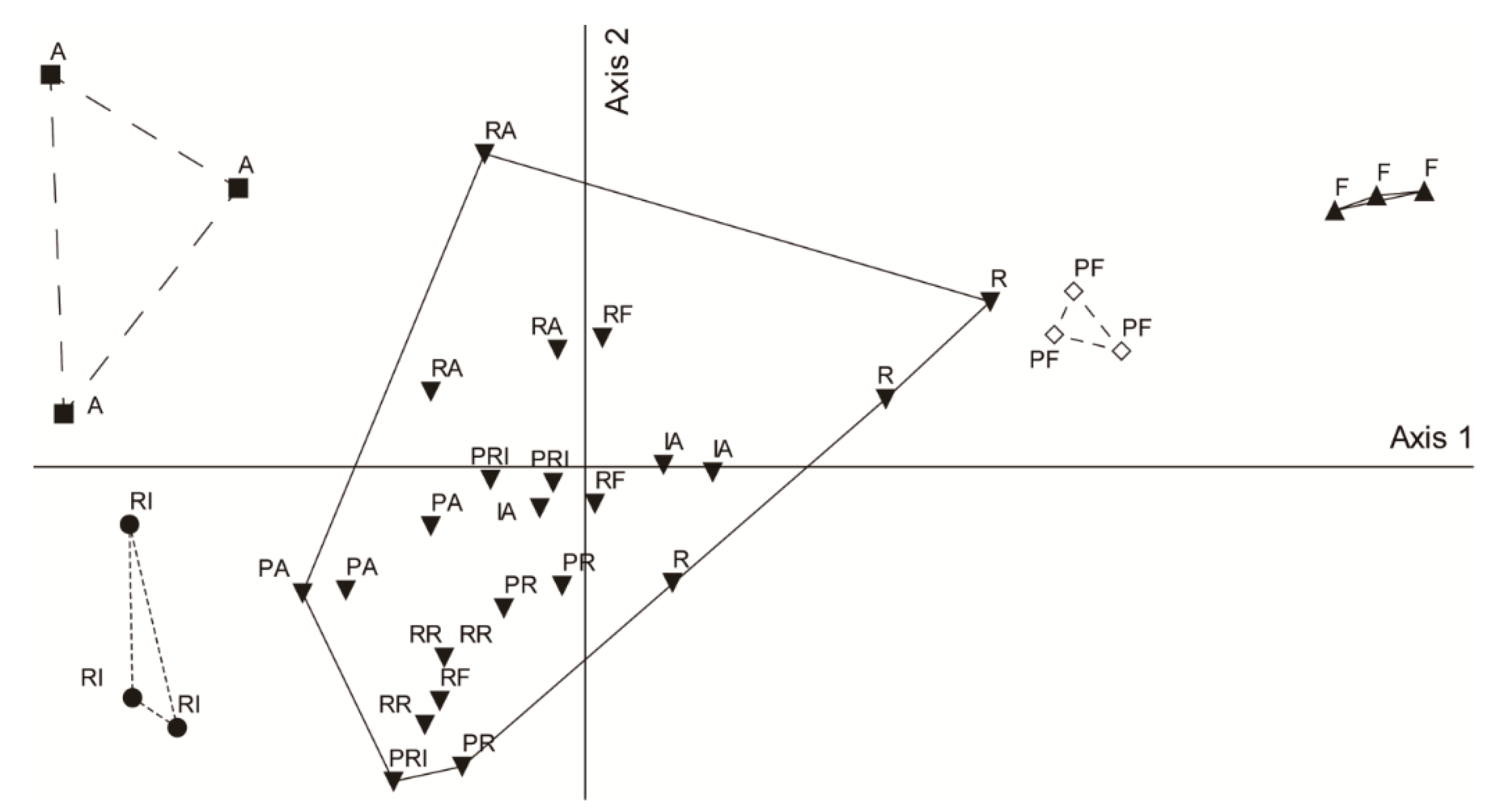
3.2. Change of Carabid Assemblage in Urban Habitats
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- McKinney, M.L. Urbanization as a major cause of biotic homogenization. Biol. Conserv. 2006, 127, 247–260. [Google Scholar] [CrossRef]
- Tzoulas, K.; Korpela, K.; Venn, S.; Yli-Pelkonen, V.; Kaźmierczak, A.; Niemelä, J.; James, P. Promoting ecosystem and human health in urban areas using Green Infrastructure: A literature review. Landsc. Urban Plan. 2007, 81, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Hostetler, M.E.; Allen, W.; Meurk, C. Conserving urban biodiversity? Creating green infrastructure is only the first step. Landsc. Urban Plan. 2011, 100, 369–371. [Google Scholar] [CrossRef]
- Dearborn, D.C.; Kark, S. Motivations for Conserving Urban Biodiversity. Conserv. Biol. 2010, 24, 432–440. [Google Scholar] [CrossRef]
- Carmona, M. Public Places, Urban Spaces: The Dimensions of Urban Design; Routledge: Oxford, UK, 2010. [Google Scholar]
- Gardiner, M.M.; Burkman, C.E.; Prajzner, S.P. The Value of Urban Vacant Land to Support Arthropod Biodiversity and Ecosystem Services. Environ. Èntomol. 2013, 42, 1123–1136. [Google Scholar] [CrossRef]
- Rudd, H.; Vala, J.; Schaefer, V. Importance of Backyard Habitat in a Comprehensive Biodiversity Conservation Strategy: A Connectivity Analysis of Urban Green Spaces. Restor. Ecol. 2002, 10, 368–375. [Google Scholar] [CrossRef] [Green Version]
- Dramstad, W.E. Landscape Ecology Principles in Landscape Architecture and Land-Use Planning; Island Press: Washington, DC, USA, 1996. [Google Scholar]
- Wolch, J.R.; Byrne, J.; Newell, J.P. Urban green space, public health, and environmental justice: The challenge of making cities ‘just green enough’. Landsc. Urban Plan. 2014, 125, 234–244. [Google Scholar] [CrossRef] [Green Version]
- Jim, C. Green-space preservation and allocation for sustainable greening of compact cities. Cities 2004, 21, 311–320. [Google Scholar] [CrossRef]
- Campbell, S. Green Cities, Growing Cities, Just Cities?: Urban Planning and the Contradictions of Sustainable Development. JAPA 1996, 62, 296–312. [Google Scholar] [CrossRef]
- Alvey, A.A. Promoting and preserving biodiversity in the urban forest. Urban For. Urban Green. 2006, 5, 195–201. [Google Scholar] [CrossRef]
- Hahs, A.K.; McDonnell, M.J.; McCarthy, M.A.; Vesk, P.A.; Corlett, R.T.; Norton, B.A.; Clemants, S.E.; Duncan, R.P.; Thompson, K.; Schwartz, M.W. A global synthesis of plant extinction rates in urban areas. Ecol. Lett. 2009, 12, 1165–1173. [Google Scholar] [CrossRef] [PubMed]
- Ramalho, C.E.; Hobbs, R.J. Time for a change: Dynamic urban ecology. Trends Ecol. Evol. 2012, 27, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Davies, K.F.; Margules, C.R. Effects of habitat fragmentation on carabid beetles: Experimental evidence. J. Anim. Ecol. 1998, 67, 460–471. [Google Scholar] [CrossRef]
- Niemelä, J.; Kotze, D.J.; Venn, S.; Penev, L.; Stoyanov, I.; Spence, J.; Hartley, D.; De Oca, E.M. Carabid beetle assemblages (Coleoptera, Carabidae) across urban-rural gradients: An international comparison. Landsc. Ecol. 2002, 17, 387–401. [Google Scholar] [CrossRef]
- Do, Y.; Lineman, M.; Joo, G.-J. Carabid beetles in green infrastructures: The importance of management practices for improving the biodiversity in a metropolitan city. Urban Ecosyst. 2014, 1–13. [Google Scholar] [CrossRef]
- Small, E.C.; Sadler, J.P.; Telfer, M.G. Carabid beetle assemblages on urban derelict sites in Birmingham, UK. J. Insect Conserv. 2002, 6, 233–246. [Google Scholar] [CrossRef]
- Venn, S.J.; Kotze, D.J.; Lassila, T.; Niemelä, J.K. Urban dry meadows provide valuable habitat for granivorous and xerophylic carabid beetles. J. Insect Conserv. 2013, 17, 747–764. [Google Scholar] [CrossRef]
- Simpson, E. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press Urbana: Urbana, IL, USA, 1963. [Google Scholar]
- Hammer, Ø.; Harper, D.A. Paleontological Data Analysis; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Jongman, R.H.; Ter Braak, C.J.; Van Tongeren, O.F. Data Analysis in Community and Landscape Ecology; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Elek, Z.; Lövei, G.L. Patterns in ground beetle (Coleoptera: Carabidae) assemblages along an urbanisation gradient in Denmark. Acta Oecologica 2007, 32, 104–111. [Google Scholar] [CrossRef]
- Gray, J.S. Effects of environmental stress on species rich assemblages. Biol. J. Linn. Soc. 1989, 37, 19–32. [Google Scholar] [CrossRef]
- Strauss, B.; Biedermann, R. Urban brownfields as temporary habitats: Driving forces for the diversity of phytophagous insects. Ecography 2006, 29, 928–940. [Google Scholar]
- Eversham, B.C.; Roy, D.B.; Telfer, M.G. Urban, industrial and other manmade sites as analogues of natural habitats for Carabidae. Ann. Zool. Fenn. 1996, 33, 149–156. [Google Scholar]
- Hartley, D.; Koivula, M.; Spence, J.; Pelletier, R.; Ball, G. Effects of urbanization on ground beetle assemblages (Coleoptera, Carabidae) of grassland habitats in western Canada. Ecography 2007, 30, 673–684. [Google Scholar] [CrossRef]
- Lövei, G.L.; Sunderland, K.D. Ecology and Behavior of Ground Beetles (Coleoptera: Carabidae). Annu. Rev. Èntomol. 1996, 41, 231–256. [Google Scholar] [CrossRef] [PubMed]
- Niemelä, J.; Kotze, D.J. Carabid beetle assemblages along urban to rural gradients: A review. Landsc. Urban Plan. 2009, 92, 65–71. [Google Scholar] [CrossRef]
- Magura, T.; Lövei, G.L. The Permeability of Natural versus Anthropogenic Forest Edges Modulates the Abundance of Ground Beetles of Different Dispersal Power and Habitat Affinity. Diversity 2020, 12, 320. [Google Scholar] [CrossRef]
- Magura, T.; Ferrante, M.; Lövei, G.L. Only habitat specialists become smaller with advancing urbanization. Glob. Ecol. Biogeogr. 2020, 29, 1978–1987. [Google Scholar] [CrossRef]
- Vergnes, A.; Chantepie, S.; Robert, A.; Clergeau, P. Are urban green spaces suitable for woodland carabids? First insights from a short-term experiment. J. Insect Conserv. 2013, 17, 671–679. [Google Scholar] [CrossRef]
- Ishitani, M.; Kotze, D.; Niemelä, J. Changes in carabid beetle assemblages across an urban-rural gradient in Japan. Ecography 2003, 26, 481–489. [Google Scholar] [CrossRef] [Green Version]
- Magura, T.; Tóthmérész, B.; Molnar, T. Changes in carabid beetle assemblages along an urbanisation gradient in the city of Debrecen, Hungary. Landsc. Ecol. 2004, 19, 747–759. [Google Scholar] [CrossRef]
- Magura, T.; Tóthmérész, B.; Lövei, G.L. Body size inequality of carabids along an urbanisation gradient. Basic Appl. Ecol. 2006, 7, 472–482. [Google Scholar] [CrossRef] [Green Version]
- Ribera, I.; Dolédec, S.; Downie, I.S.; Foster, G.N. Effect of land disturbance and stress on species traits of ground beetle assemblages. Ecology 2001, 82, 1112–1129. [Google Scholar] [CrossRef]
- Schmid-Hempel, P. Evolutionary ecology of insect immune defenses. Annu. Rev. Èntomol. 2005, 50, 529–551. [Google Scholar] [CrossRef]
- Magura, T.; Lövei, G.L. Consequences of Urban Living: Urbanization and Ground Beetles. Curr. Landsc. Ecol. Rep. 2020, 2020, 1–13. [Google Scholar] [CrossRef]
- Do, Y.; Joo, G. The effect of fragmentation and intensive management on carabid beetles in coniferous forest. Appl. Ecol. Environ. Res. 2013, 11, 451–461. [Google Scholar] [CrossRef]
- Fahrig, L. Effects of Habitat Fragmentation on Biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef] [Green Version]
- Fahrig, L. Relative Effects of Habitat Loss and Fragmentation on Population Extinction. J. Wildl. Manag. 1997, 61, 603–610. [Google Scholar] [CrossRef]
- Hanski, I.; Moilanen, A.; Gyllenberg, M. Minimum Viable Metapopulation Size. Am. Nat. 1996, 147, 527–541. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Abbrev | HP | FA | F | A | RI | R | RA | RF | PF | PR | PA | PRI | IA | RR |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Amara chalcites | Ach | O | A | 34 | 42 | 26 | 16 | 18 | 31 | 8 | 42 | 33 | 41 | 20 | 24 |
| Amara congrua | Aco | O | A | 16 | 33 | 18 | 8 | 22 | 33 | 16 | 23 | 3 | 22 | ||
| Amara macronota | Ama | E | A | 26 | 15 | 32 | 82 | 2 | 14 | ||||||
| Anisodactylus punctatipennis | Apu | O | A | 55 | 10 | 8 | 54 | 9 | 5 | 63 | 9 | 4 | 4 | 1 | |
| Anisodactylus signatus | Asi | O | A | 27 | 32 | 2 | |||||||||
| Carabus sternbergi | Cst | F | N | 44 | 2 | ||||||||||
| Chlaenius micans | Cmi | F | A | 34 | 9 | 6 | 4 | 12 | |||||||
| Chlaenius naeviger | Can | E | A | 14 | 9 | ||||||||||
| Chlaenius pallipes | Cpa | O | A | 12 | 43 | 3 | 4 | 9 | 27 | ||||||
| Chlaenius virgulifer | Cvi | E | A | 21 | |||||||||||
| Damaster jankowskii | Dja | F | N | 25 | |||||||||||
| Dolichus halensis | Dha | O | N | 91 | 65 | 7 | |||||||||
| Harpalus capito | Hca | O | A | 4 | 57 | 16 | 8 | 7 | |||||||
| Harpalus sinicus | His | O | A | 42 | 3 | ||||||||||
| Nebria chinensis | Nch | O | A | 8 | 28 | ||||||||||
| Pheropsophus javanus | Pja | O | N | 92 | 24 | ||||||||||
| Pheropsophus jessoensis | Pje | O | N | 71 | 36 | ||||||||||
| Synuchus cycloderus | Scy | F | A | 158 | 23 | ||||||||||
| Synuchus melantho | Sme | F | A | 86 | 4 | ||||||||||
| Synuchus nitidus | Sni | F | A | 32 | 12 | ||||||||||
| No. of species | 10 | 16 | 11 | 5 | 7 | 4 | 7 | 3 | 5 | 7 | 3 | 3 | |||
| No. of individuals | 476 | 595 | 309 | 97 | 54 | 67 | 209 | 84 | 72 | 98 | 27 | 47 | |||
| Parameter | Source of Variation | df | MS | F | P |
|---|---|---|---|---|---|
| Richness | Types | 1 | 256.889 | 12.506 | <0.001 |
| Habitat types (Types) | 10 | 20.542 | 33.614 | <0.001 | |
| Error | 24 | 0.611 | |||
| Abundance | Types | 1 | 73,216.889 | 13.571 | 0.004 |
| Habitat types (Types) | 10 | 5395.208 | 32.334 | <0.001 | |
| Error | 24 | 166.861 | |||
| Diversity index | Types | 1 | 4.794 | 9.479 | 0.012 |
| Habitat types (Types) | 10 | 0.506 | 9.933 | <0.001 | |
| Error | 24 | 0.051 | |||
| Dominance index | Types | 1 | 0.278 | 6.003 | 0.034 |
| Habitat types (Types) | 10 | 0.046 | 4.296 | 0.002 | |
| Error | 24 | 0.011 | |||
| Evenness index | Types | 1 | 0.022 | 1.771 | 0.213 |
| Habitat types (Types) | 10 | 0.012 | 0.702 | 0.712 | |
| Error | 24 | 0.017 |
| df | Deviance | Ratio | P | |
|---|---|---|---|---|
| Richness | ||||
| Habitat preference | 2 | 0.980 | 0.490 | 0.613 |
| Flight ability | 1 | 0.045 | 0.045 | 0.832 |
| Types | 1 | 13.423 | 13.423 | <0.001 |
| Habitat preference × Types | 2 | 1.433 | 0.717 | 0.488 |
| Flight ability × Types | 1 | 4.110 | 4.110 | 0.043 |
| Residual | 52 | 51.342 | ||
| Total | 7 | 93.459 | ||
| Abundance | ||||
| Habitat preference | 2 | 2.912 | 1.456 | 0.243 |
| Flight ability | 1 | 2.684 | 2.684 | 0.107 |
| Types | 1 | 21.576 | 21.576 | <0.001 |
| Habitat preference × Types | 2 | 3.333 | 1.666 | 0.199 |
| Flight ability × Types | 1 | 7.314 | 7.314 | 0.009 |
| Residual | 52 | 2022.295 | ||
| Total | 7 | 73.478 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, Y.; Kim, C.-J.; Choi, M.-B.; Do, Y. A Comparative Study of Carabid Beetles in Green Spaces and Former Natural Habitats. Diversity 2020, 12, 479. https://doi.org/10.3390/d12120479
Park Y, Kim C-J, Choi M-B, Do Y. A Comparative Study of Carabid Beetles in Green Spaces and Former Natural Habitats. Diversity. 2020; 12(12):479. https://doi.org/10.3390/d12120479
Chicago/Turabian StylePark, Yonghwan, Chang-Jun Kim, Moon-Bo Choi, and Yuno Do. 2020. "A Comparative Study of Carabid Beetles in Green Spaces and Former Natural Habitats" Diversity 12, no. 12: 479. https://doi.org/10.3390/d12120479
APA StylePark, Y., Kim, C.-J., Choi, M.-B., & Do, Y. (2020). A Comparative Study of Carabid Beetles in Green Spaces and Former Natural Habitats. Diversity, 12(12), 479. https://doi.org/10.3390/d12120479