Sea Slugs—“Rare in Space and Time”—But Not Always



Abstract
1. Introduction
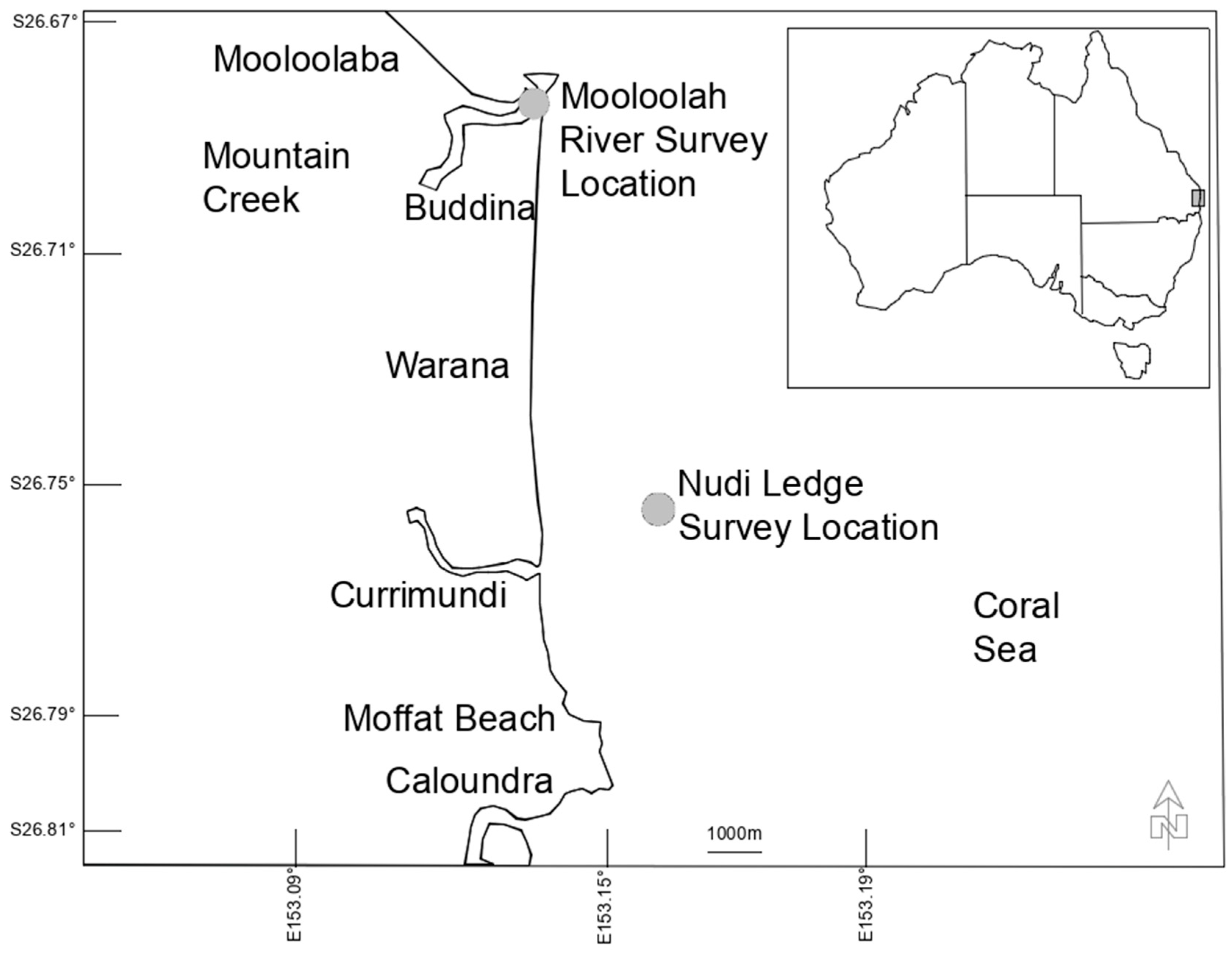
2. Materials and Methods
3. Results
3.1. Numerical Rarity
3.2. Temporal Rarity
3.3. Frequently Observed Species
3.3.1. Trends for Frequently Observed Species—Mooloolah River
3.3.2. Trends of Frequently Observed Species—Nudi Ledge
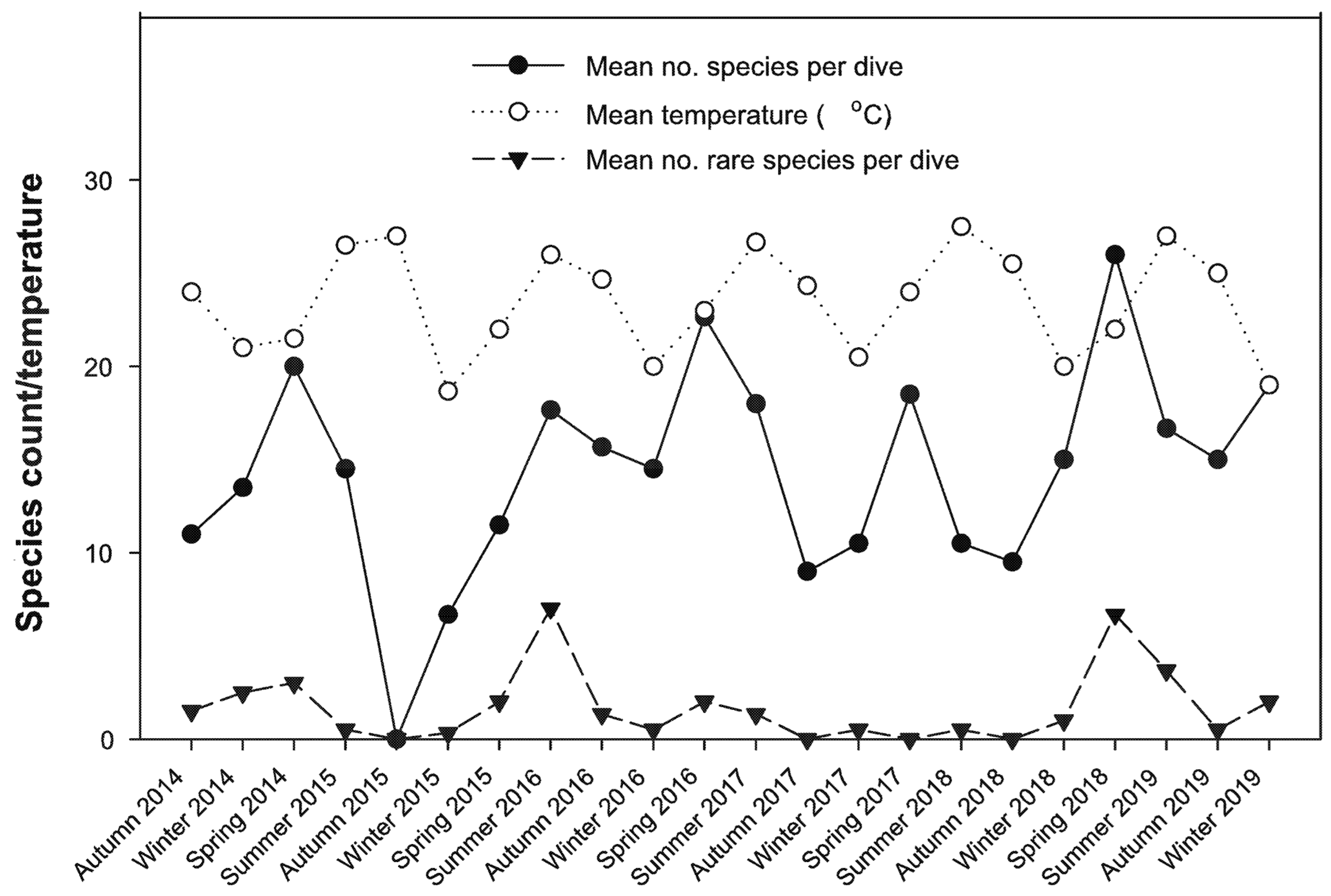
3.4. Correlation between Biotic Variables and Water Temperature
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Preston, F.W. The Commonness, And Rarity, of Species. Ecology. 1948, 29, 254–283. [Google Scholar] [CrossRef]
- Benkendorff, K.; Przeslawski, R. Multiple measures are necessary to assess rarity in macro-molluscs: A case study from southeastern Australia. Biodivers. Conserv. 2008, 17, 2455–2478. [Google Scholar] [CrossRef]
- Rabinowitz, D. Seven forms of rarity. In The Biological Aspects of Rare Plant Conservation; Synge, H., Ed.; John Wiley & Sons: Somerset, NJ, USA, 1981. [Google Scholar]
- Gaston, K.J. Rarity; Chapman & Hall: London, UK, 1994. [Google Scholar]
- Leroy, B.; Gallon, R.; Feunteun, E.; Robuchon, M.; Frédéric, Y. Cross-taxon congruence in the rarity of subtidal rocky marine assemblages: No taxonomic shortcut for conservation monitoring. Ecol. Indic. 2017, 77, 239–249. [Google Scholar] [CrossRef]
- Eisenbarth, J.H.; Undap, N.; Papu, A.; Schillo, D.; Dialao, J.; Reumschüssel, S.; Kaligis, F.; Bara, R.; Schäberle, F.T.; König, M.G.; et al. Marine Heterobranchia (Gastropoda, Mollusca) in Bunaken National Park, North Sulawesi, Indonesia—A Follow-Up Diversity Study. Diversity 2018, 10, 127. [Google Scholar] [CrossRef]
- Larkin, M.F.; Smith, S.D.A.; Willan, R.C.; Davis, T.R. Diel and seasonal variation in heterobranch sea slug assemblages within an embayment in temperate eastern Australia. Mar. Biodivers. 2018, 48, 1541–1550. [Google Scholar] [CrossRef]
- Marshall, J.G.; Willan, R.C. Nudibranchs of Heron Island, Great Barrier Reef: A survey of the Opisthobranchia (sea slugs) of Heron and Wistari Reefs; Backhuys Publishers: Leiden, The Netherlands, 1999. [Google Scholar]
- Willan, R.; Morton, J. Marine Molluscs Part 2 Opisthobranchia. Leigh Laboratory Bulletin 13; University of Auckland, Leigh Marine Laboratory: Auckland, New Zealand, 1984. [Google Scholar]
- Tanamura, D.; Hirose, E. Seasonal Occurrence of Gastropterids (Gastropoda: Cephalaspidea) and Their Habitat Selection in a Subtropical Back-reef on Okinawajima Island (Ryukyu Archipelago, Japan). Zool. Stud. 2017, 56, 1–8. [Google Scholar] [CrossRef]
- Siqueira, T.; Bini, L.M.; Roque, F.O.; Marques Couceiro, S.R.; Trivinho-Strixino, S.; Cottenie, K. Common and rare species respond to similar niche processes in macroinvertebrate metacommunities. Ecography 2012, 35, 183–192. [Google Scholar] [CrossRef]
- Burlakova, L.; Karatayev, A.; Karatayev, V.; May, M.; Bennett, D.; Cook, M. Endemic species: Contribution to community uniqueness, effect of habitat alteration, and conservation priorities. Biol. Conserv. 2011, 144, 155–165. [Google Scholar] [CrossRef]
- Smith, S.D.A.; Peregrin, L.S. The shelled gastropods of the Solitary Islands Marine Park, northern New South Wales, Australia. Molluscan Res. 2020, 40, 142–149. [Google Scholar] [CrossRef]
- Flather, C.H.; Sieg, C.H. Species Rarity: Definition, Causes and Classifications; Island Press: Washington, DC, USA, 2007. [Google Scholar]
- Gray, J.S.; Bjørgesæter, A.; Karl, I.U. On Plotting Species Abundance Distributions. J. Anim. Ecol. 2006, 75, 752–756. [Google Scholar] [CrossRef]
- Bouchet, P.; Lozouet, P.; Maestrati, P.; Heros, V. Assessing the magnitude of species richness in tropical marine environments: Exceptionally high numbers of molluscs at a New Caledonia site. Biol. J. Linn. Soc. 2002, 75, 421–436. [Google Scholar] [CrossRef]
- Betti, F.; Bava, S.; Cattaneo-Vietti, R. Composition and seasonality of a heterobranch assemblage in a sublittoral, unconsolidated, wave-disturbed community in the Mediterranean Sea. J. Molluscan Stud. 2017, 83, 325–332. [Google Scholar] [CrossRef]
- Gaston, K.J. Global patterns in biodiversity. Nature 2000, 405, 220. [Google Scholar] [CrossRef]
- Behrens, D.W. Nudibranch Behavior; New World Publications Inc.: Jacksonville, FL, USA, 2007. [Google Scholar]
- Crocetta, F.; Macali, A.; Furfaro, G.; Cooke, S.; Villani, G.; Valdes, A. Alien molluscan species established along the Italian shores: An update, with discussions on some Mediterranean “alien species” categories. ZooKeys 2013, 277, 91–108. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ekimova, I.A.; Antokhina, T.I.; Schepetov, D.M. “Invasion” in the Russian Arctic: Is global Climate Change a real driver? A remarkable case of two nudibranch species. Ruthenica 2019, 29, 103–113. [Google Scholar]
- Evertsen, J.; Bakken, T. Diversity of Norwegian sea slugs (Nudibranchia): New species to Norwegian coastal waters and new data on distribution of rare species. Fauna Nor. 2013, 32, 45–52. [Google Scholar] [CrossRef][Green Version]
- Rudman, W. Population Fluctuations. Sea Slug Forum. Available online: http://www.seaslugforum.net/factsheet/popfluc (accessed on 27 September 2019).
- Debelius, H.; Kuiter, R.H. Nudibranchs of the World; IAKN-Unterwasserarchiv: Frankfurt, Germany, 2007. [Google Scholar]
- Kaligis, F.; Eisenbarth, J.-H.; Schillo, D.; Dialao, J.; Schäberle, T.F.; Böhringer, N.; Bara, R.; Reumschüssel, S.; König, G.M.; Wägele, H.; et al. Second survey of heterobranch sea slugs (Mollusca, Gastropoda, Heterobranchia) from Bunaken National Park, North Sulawesi, Indonesia — how much do we know after 12 years? Mar. Biodivers. Rec. 2018, 11, 2. [Google Scholar] [CrossRef]
- Davis, T.R.; Harasti, D.; Kelaher, B.; Smith, S.D.A. Spatial and temporal variation in subtidal molluscan diversity amongst temperate estuarine habitats. Mar. Ecol. 2017, 38. [Google Scholar] [CrossRef]
- Arias, A.; Crocetta, F. Umbraculum umbraculum (Gastropoda: Heterobranchia) spreading northwards: Additional evidence to the “tropicalization” of the Bay of Biscay. Cah. Biol. Mar. 2016, 57, 285–286. [Google Scholar]
- Nimbs, M.; Smith, S.D.A. Beyond Capricornia: Tropical Sea Slugs (Gastropoda, Heterobranchia) Extend Their Distributions into the Tasman Sea. Diversity 2018, 10, 99. [Google Scholar] [CrossRef]
- Schultz, S.T.; Goddard, J.H.; Gosliner, T.M.; Mason, D.E.; Pence, W.E.; McDonald, G.R.; Pearse, V.B.; Pearse, J.S. Climate-index response profiling indicates larval transport is driving population fluctuations in nudibranch gastropods from the northeast Pacific Ocean. Limnol. Oceanogr. 2011, 56, 749. [Google Scholar] [CrossRef]
- O’Hara, T. Endemism, rarity and vulnerability of marine species along a temperate coastline. Invertebr. Syst. 2002, 16, 671–684. [Google Scholar] [CrossRef]
- Tanamura, D.; Hirose, E. Population Dynamics of the Sea Slug Plakobranchus ocellatus (Opisthobranch: Sacoglossa: Elysioidea) on a Subtropical Coral Reef off Okinawa-jima Island, Ryukyu Archipelago, Japan. Zool. Stud. 2016, 55, 42. [Google Scholar] [CrossRef]
- Smith, S.D.A.; Davis, T.R. Slugging it out for science: Volunteers provide valuable data on the diversity and distribution of heterobranch sea slugs. Molluscan Res. 2019, 39, 214–223. [Google Scholar] [CrossRef]
- Gosliner, T.M.; Valdes, A.; Behrens, D.W. Nudibranch & Sea Slug Identification Indo-Pacific, 2nd ed.; New World Publications Inc.: Jacksonville, FL, USA, 2018. [Google Scholar]
- Nimbs, M.J.; Smith, S.D.A. An illustrated inventory of the sea slugs of New South Wales, Australia (Gastropoda: Heterobranchia). R. Soc. Vic. 2017, 128, 44–113. [Google Scholar] [CrossRef]
- Nimbs, M.J.; Hutton, I.; Davis, T.R.; Larkin, M.F.; Smith, S.D.A. The heterobranch sea slugs of Lord Howe Island, NSW, Australia (Mollusca: Gastropoda). Proc. R. Soc. Vic. 2020, 132, 12–41. [Google Scholar] [CrossRef]
- Maritime Safety Queensland. Tidal Planes. Available online: https://www.msq.qld.gov.au/Tides/Tidal-planes (accessed on 26 June 2019).
- WoRMS Editorial Board. World Register of Marine Species. Available online: http://www.marinespecies.org (accessed on 27 March 2019).
- Korshunova, T.; Martynov, A.; Bakken, T.; Evertsen, J.; Fletcher, K.; Mudianta, I.; Saito, H.; Lundin, K.; Schrödl, M.; Picton, B. Polyphyly of the traditional family Flabellinidae affects a major group of Nudibranchia: Aeolidacean taxonomic reassessment with descriptions of several new families, genera, and species (Mollusca, Gastropoda). ZooKeys 2017, 717, 1–139. [Google Scholar] [CrossRef]
- Burn, R. Nudibranchs and Related Molluscs; Museum Victoria: Melbourne, Australia, 2015. [Google Scholar]
- Coleman, N. Nudibranchs Encyclopedia, 2nd ed.; Masalai Press and Underwater Australasia: Oakland, Australia; Byron Bay, CA, USA, 2015. [Google Scholar]
- Crocetta, F.; Poursanidis, D.; Tringali, L.P. Biodiversity of sea slugs and shelled relatives (Mollusca: Gastropoda) of the Cretan Archipelago (Greece), with taxonomic remarks on selected species. Quat. Int. 2015, 390, 56–68. [Google Scholar] [CrossRef]
- Borges, P.A.V.; Ugland, K.I.; Dinis, F.; Gaspar, C. Insect and spider rarity in an oceanic island (Terceira, Azores): True rare and pseudo-rare species. In Insect Ecology and Conservation; Fattorini, S., Ed.; Research Signpost: Kerala, India, 2008; pp. 47–70. [Google Scholar]
- Harnik, P.G.; Simpson, C.; Payne, J.L. Long-term differences in extinction risk among the seven forms of rarity. Proc. R. Soc. B. 2012, 279. [Google Scholar] [CrossRef]
- Murphy, M.J. A survey of the marine bivalves and gastropods of Shelly Beach, Port Macquarie, on the north coast of New South Wales, Australia. Vic. Nat. 2015, 132, 73–85. [Google Scholar]
- Poloczanska, E.S.; Smith, S.; Fauconnet, L.; Healy, J.; Tibbetts, I.R.; Burrows, M.T.; Richardson, A.J. Little change in the distribution of rocky shore faunal communities on the Australian east coast after 50 years of rapid warming. J. Exp. Mar. Biol. Ecol. 2011, 400, 145–154. [Google Scholar] [CrossRef]
- DeVantier, L.; Williamson, D.; Willan, R. Nearshore Marine Biodiversity of the Sunshine Coast, South-East Queensland: Inventory of molluscs, corals and fishes July 2010; Baseline Survey Report to the Noosa Integrated Catchment Association: Tewantin, Australia, 2010. [Google Scholar] [CrossRef]
- Short, A.D.; Woodroffe, C.D. The Coast of Australia; Cambridge University Press: New York, NY, USA, 2009. [Google Scholar]
- Almada, F.; Levy, A.; Robalo, J. Not so sluggish: The success of the Felimare picta complex (Gastropoda, Nudibranchia) crossing Atlantic biogeographic barriers. PeerJ 2016, 4, e1561. [Google Scholar] [CrossRef] [PubMed]
- Lim, G.; Balke, M.; Meier, R. Determining Species Boundaries in a World Full of Rarity: Singletons, Species Delimitation Methods. Syst. Biol. 2011, 61, 165–169. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Quartile | Mooloolah River a | Nudi Ledge a |
|---|---|---|
| A | ||
| First quartile (rare) | 1, 85 (34%) | 1, 61 (32%) |
| Second quartile | 2–3, 52 (21%) | 2–3, 40 (21%) |
| Third quartile | 4–14, 52 (21%) | 4–13, 41(22%) |
| Upper quartile | >14, 62 (25%) | >13, 46 (24%) |
| B | ||
| First quartile (rare) | 1–7, 165 (66%) | 1–7, 124 (66%) |
| Second quartile | 8–63, 65 (26%) | 8–63, 48 (26%) |
| Third quartile | 64–511, 15 (6%) | 64–511, 15(8%) |
| Upper quartile | >511, 6 (2%) | >511, 1 (1%) |
| Quartile | Mooloolah River a | Nudi Ledge a |
|---|---|---|
| First quartile (rare) | 1, 96 (38%) | 2, 71 (38%) |
| Second quartile | 2, 52 (21%) | 3–5, 27 (14%) |
| Third quartile | 3–5, 42 (17%) | 6–20, 44 (23%) |
| Upper quartile | >5, 61 (24%) | >20, 46 (24%) |
| Cut-Off Value and Classification | Mooloolah River | Nudi Ledge |
|---|---|---|
| 25% of surveys (rare) | 238 (95%) | 147 (78%) |
| 50% of surveys (uncommon) | 7 (39%) | 25 (13%) |
| 75% of surveys (common) | 2 (1%) | 6 (3%) |
| >75% of surveys (abundant) | 4 (2%) | 10 (5%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schubert, J.; Smith, S.D.A. Sea Slugs—“Rare in Space and Time”—But Not Always. Diversity 2020, 12, 423. https://doi.org/10.3390/d12110423
Schubert J, Smith SDA. Sea Slugs—“Rare in Space and Time”—But Not Always. Diversity. 2020; 12(11):423. https://doi.org/10.3390/d12110423
Chicago/Turabian StyleSchubert, Julie, and Stephen D. A. Smith. 2020. "Sea Slugs—“Rare in Space and Time”—But Not Always" Diversity 12, no. 11: 423. https://doi.org/10.3390/d12110423
APA StyleSchubert, J., & Smith, S. D. A. (2020). Sea Slugs—“Rare in Space and Time”—But Not Always. Diversity, 12(11), 423. https://doi.org/10.3390/d12110423