The Fate of Endangered Rock Sedge (Carex rupestris) in the Western Carpathians—The Future Perspective of an Arctic-Alpine Species under Climate Change


 ,
, 
Abstract
1. Introduction
2. Materials and Methods
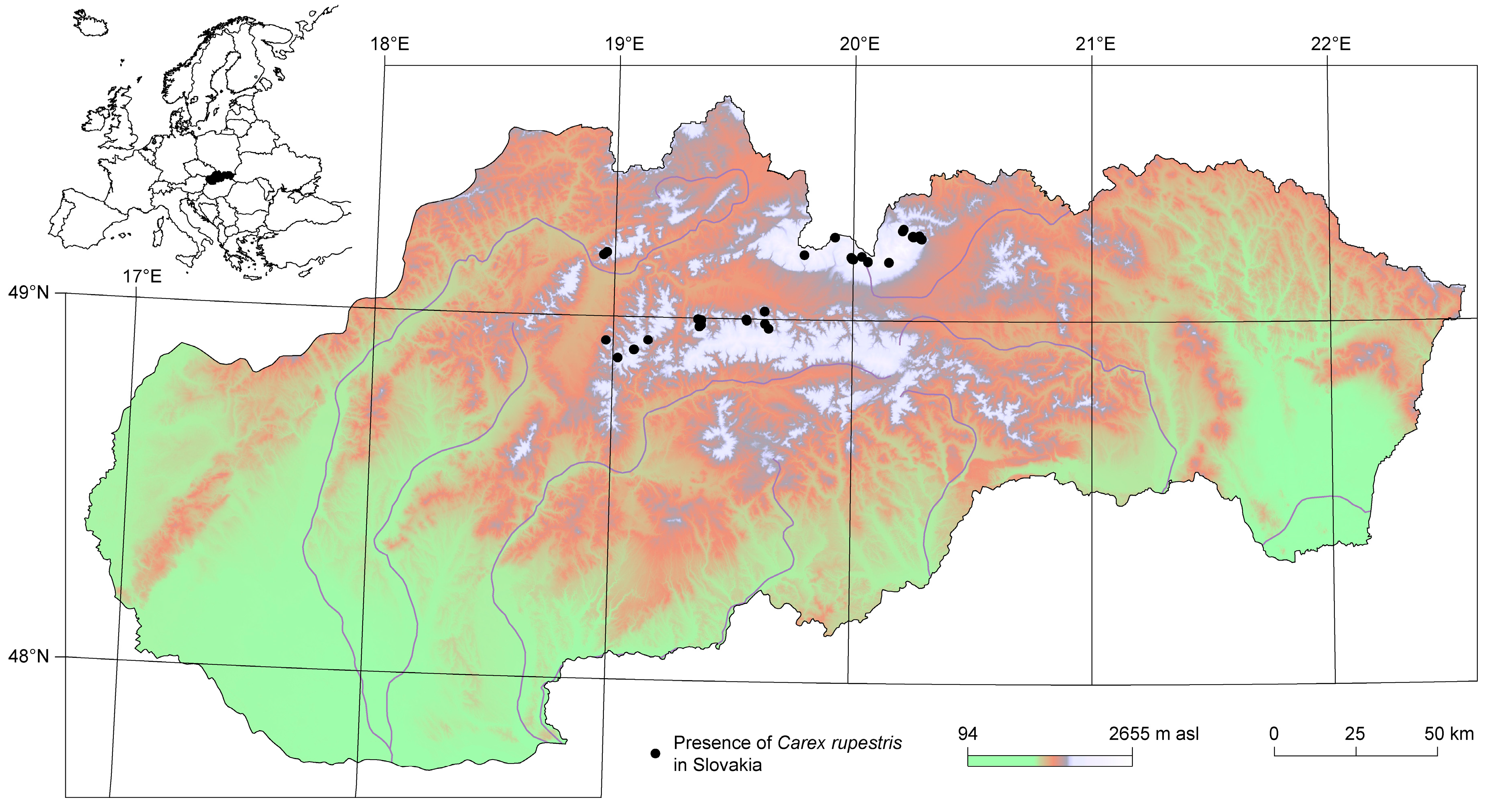
2.1. Vegetation Data
2.2. Environmental Data
2.3. Data Analysis
3. Results
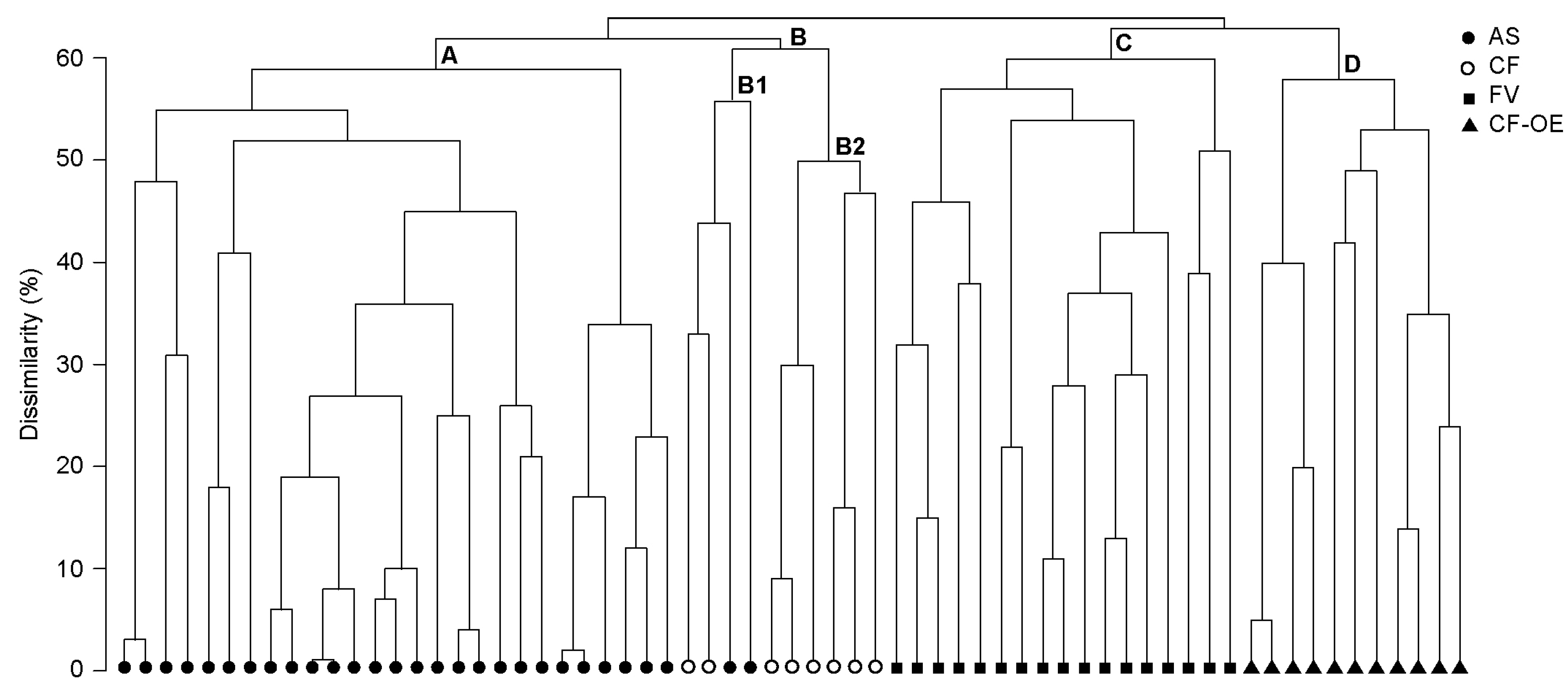
3.1. Vegetation Classification of Plots with Carex rupestris
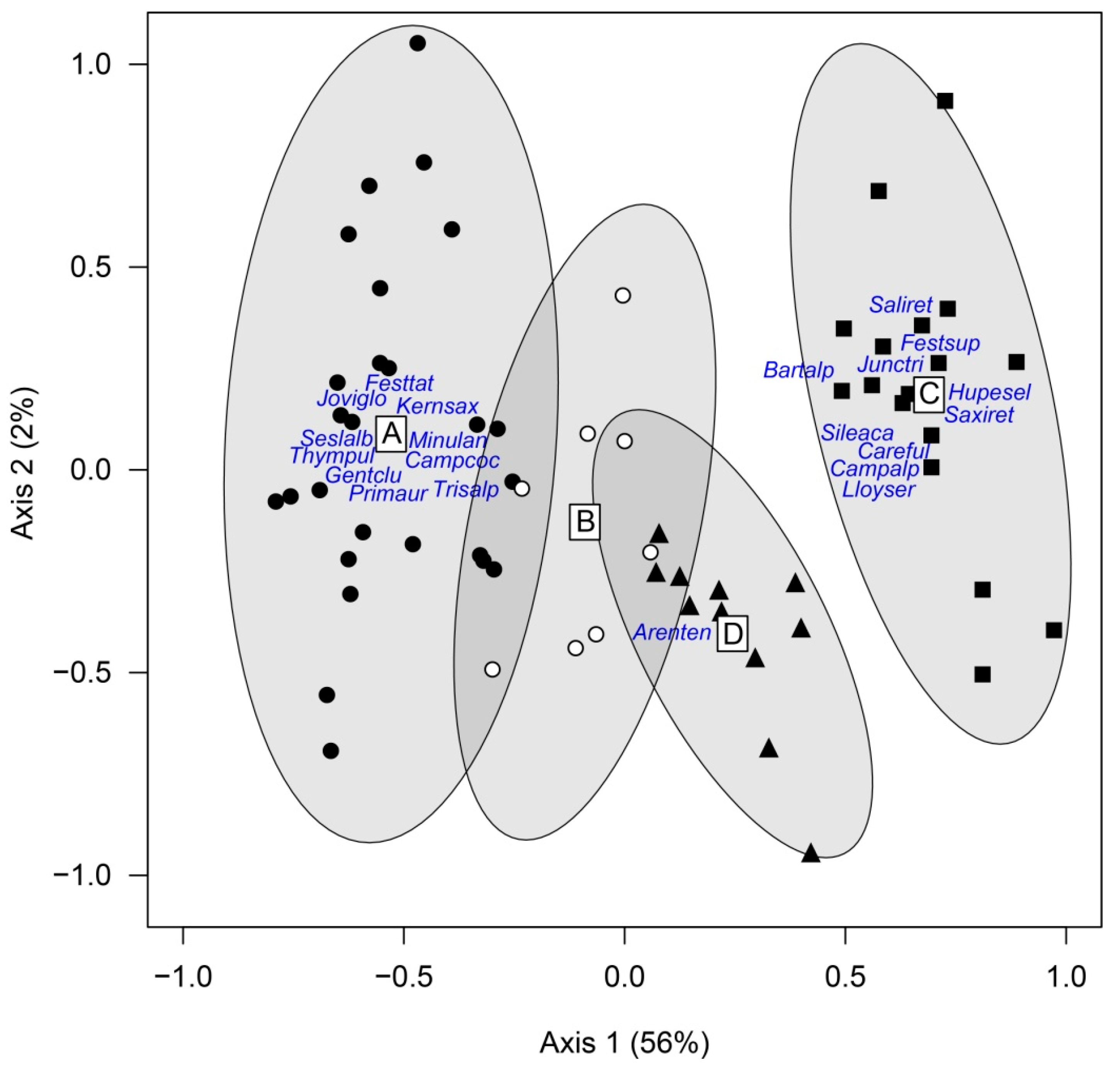
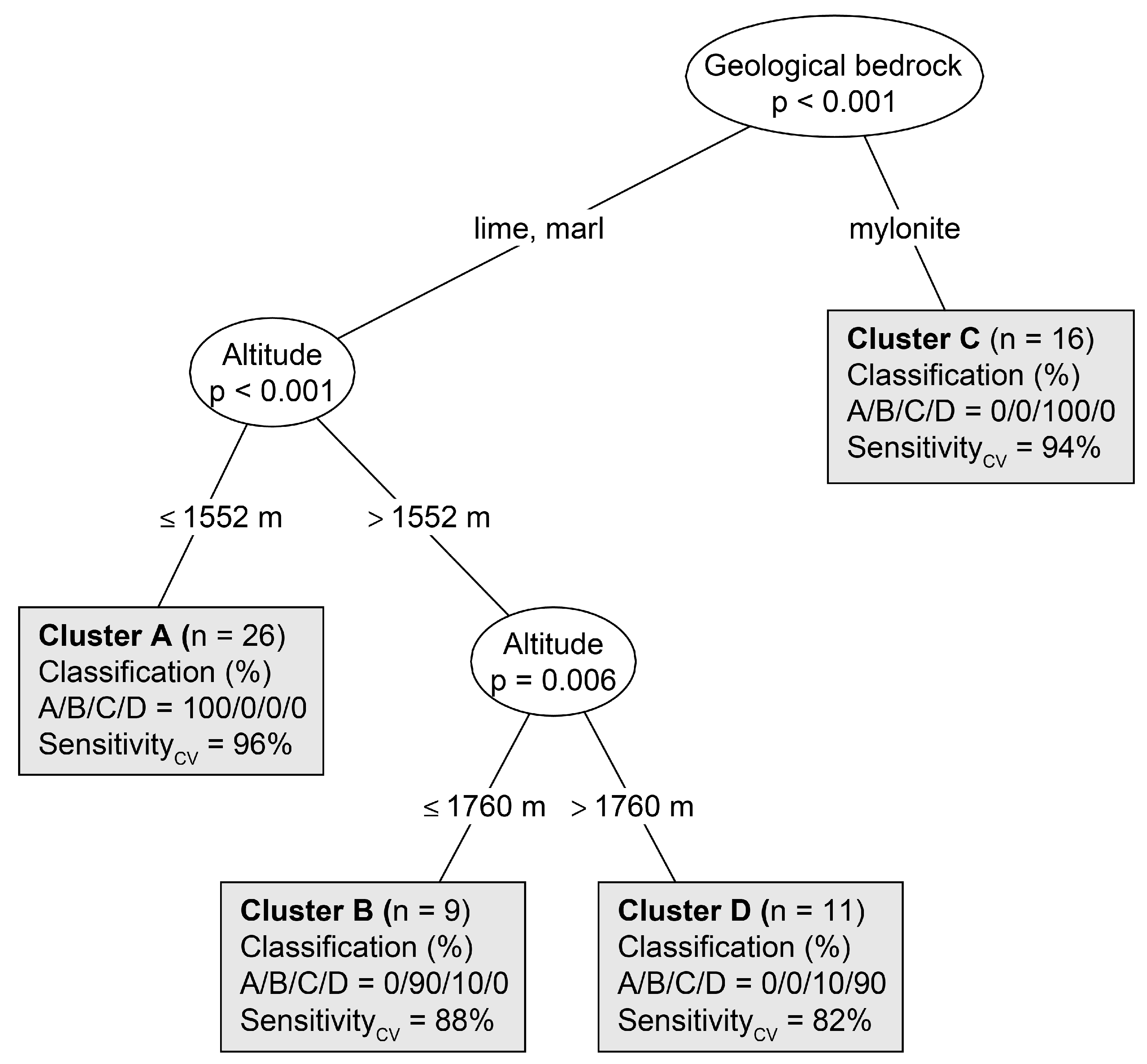
3.2. Environmental Discrimination of Vegetation Groups
3.3. Distribution of Carex rupestris
4. Discussion
4.1. Vegetation Classification and Environmental Differentiation of Plots with Carex rupestris
4.2. The Fate of Carex rupestris in the Western Carpathians
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ozenda, P. La Végétation de la Chaîne Alpine dans l’Espace Montagnard Européen, 1st ed.; Masson: Paris, France, 1985; p. 344. [Google Scholar]
- Oriolo, G. Naked rush swards (Oxytropido-Elynion Br.-Bl. 1949) on the Alps and the Apennines and their syntaxonomical position. Fitosociologia 2001, 38, 91–101. [Google Scholar]
- Petrík, A.; Šibík, J.; Valachovič, M. The class Carici rupestris-Kobresietea bellardii Ohba 1974 also in the Western Carpathians. Hacquetia 2005, 4, 33–51. [Google Scholar]
- Hájková, P.; Hájek, M.; Apostolova, I. Diversity of wetland vegetation in the Bulgarian high mountains, main gradients and context-dependence of the pH role. Plant Ecol. 2006, 184, 111–130. [Google Scholar] [CrossRef]
- Dítě, D.; Peterka, T.; Dítětová, Z.; Hájková, P.; Hájek, M. Arcto-alpine species at their niche margin: The Western Carpathian refugia of Juncus castaneus and J. triglumis in Slovakia. Ann. Bot. Fennici 2017, 54, 67–82. [Google Scholar] [CrossRef]
- Schönswetter, P.; Popp, M.; Brochmann, C. Rare arctic-alpine plants of the European Alps have different immigration histories: The snow bed species Minuartia biflora and Ranunculus pygmaeus. Mol. Ecol. 2006, 15, 709–720. [Google Scholar] [CrossRef] [PubMed]
- Hultén, E.; Fries, M. Atlas of North European Vascular Plants, 1st ed.; Koeltz—Scientific Books: Königstein, Germany, 1986; p. 1172. [Google Scholar]
- Gottfried, M.; Pauli, H.; Futschik, A.; Akhalkatsi, M.; Barančok, P.; Alonso, J.L.B.; Coldea, G.; Dick, J.; Erschbamer, B.; Calzado, M.R.F.; et al. Continent-wide response of mountain vegetation to climate change. Nat. Clim. Chang. 2012, 2, 111–115. [Google Scholar] [CrossRef]
- Pawłowski, B. Dwie ciekawe turzyce z Czarnej Hory. Spraw. Kom. Fizjogr. 1931, 65, 145–152. [Google Scholar]
- Holub, J.; Grulich, V. Carex rupestris All. In Červená kniha ohrozených a vzácnych druhov rastlín a živočíchov SR a ČR 4. Vyššie rastliny, 1st ed.; Čeřovský, J., Feráková, V., Holub, J., Maglocký, Š., Procházka, F., Eds.; Príroda: Bratislava, Slovakia, 1999; p. 81. [Google Scholar]
- Eliáš jun, P.; Dítě, D.; Kliment, J.; Hrivnák, R.; Feráková, V. Red list of ferns and flowering plants of Slovakia, 5th ed. Biologia 2015, 70, 218–228. [Google Scholar]
- Mitka, J.; Michalik, S. Turzyca skalna. In Czerwona Księga Karpat Polskich Rośliny naczyniowe; Mirek, Z., Piękoś-Mirkowa, H., Eds.; Instytut Botaniki im. W. Szafera Polskiej Akademii Nauk: Kraków, Poland, 2008; pp. 526–527. [Google Scholar]
- Didukh, Y.P. (Ed.) Chervona Knyha Ukrayiny: Roslynnyi svit; Globalconsulting: Kyiv, Ukraine, 2009; p. 589. [Google Scholar]
- Dítě, D.; Hájek, M.; Svitková, I.; Košuthová, A.; Šoltés, R.; Kliment, J. Glacial-relict symptoms in the Western Carpathian flora. Folia Geobot. 2018, 53, 277–300. [Google Scholar] [CrossRef]
- Petrík, A.; Dúbravcová, Z.; Jarolímek, I.; Kliment, J.; Šibík, J.; Valachovič, M. Syntaxonomy and ecology of plant communities of the Carici rupestris-Kobresietea bellardii in the Western Carpathians. Biologia 2006, 61, 393–412. [Google Scholar] [CrossRef]
- Bernátová, D.; Petrík, A. Ďalšie lokality Carex rupestris All. na Slovensku. Biologia 1983, 38, 491–494. [Google Scholar]
- Bernátová, D.; Uhlířová, J. Doplnok k rozšíreniu ostrice skalnej (Carex rupestris All.) vo Veľkej Fatre. Biologia 1986, 41, 75–77. [Google Scholar]
- Bernátová, D.; Uhlířová, J. Globulario cordifoliae-Caricetum humilis ass. nova in the rocky part of the Veľká Fatra Mts. Biologia 1994, 49, 1–11. [Google Scholar]
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, 1st ed.; Cambridge University Press: New York, NY, USA, 2013; p. 1535. [Google Scholar]
- Li, D.; Wu, S.; Liu, L.; Zhang, Y.; Li, S. Vulnerability of the global terrestrial ecosystems to climate change. Glob. Chang. Biol. 2018, 24, 4095–4106. [Google Scholar] [CrossRef] [PubMed]
- Pederson, N.; Dyer, J.M.; McEwan, R.W.; Hessl, A.E.; Mock, C.J.; Orwig, D.A.; Rieder, H.E.; Cook, B.I. The legacy of episodic climatic events in shaping temperate, broadleaf forests. Ecol. Monogr. 2014, 84, 599–620. [Google Scholar] [CrossRef]
- Spinoni, J.; Antofie, T.; Barbosa, P.; Bihari, Z.; Lakatos, M.; Szalai, S.; Szentimrey, T.; Vogt, J. An overview of drought events in the Carpathian Region in 1961–2010. Adv. Sci. Res. 2013, 10, 21–32. [Google Scholar] [CrossRef]
- Bartholy, J.; Pongrácz, R.; Torma, C.; Pieczka, I.; Kardos, P.; Hunyady, A. Analysis of regional climate change modelling experiments for the Carpathian basin. Int. J. Glob. Warm. 2009, 1, 238–252. [Google Scholar] [CrossRef]
- Palamarchuk, L.; Shpyg, V.; Krakovskaia, S. Floods in the Carpathians: Synoptic analysis and numerical modeling. Geophys. Res. Abstr. 2005, 7, 00967. [Google Scholar]
- Navarro, L.; Morin, H.; Bergeron, Y.; Girona, M.M. Changes in spatiotemporal patterns of 20th century spruce budworm outbreaks in eastern Canadian boreal forests. Front. Plant Sci. 2018. [Google Scholar] [CrossRef]
- Seidl, R.; Schelhaas, M.J.; Rammer, W.; Verkerk, P.J. Increasing forest disturbances in Europe and their impact on carbon storage. Nat. Clim. Chang. 2014, 4, 806–810. [Google Scholar] [CrossRef]
- Ernakovich, J.G.; Hopping, K.A.; Berdanier, A.B.; Simpson, R.T.; Kachergis, E.J.; Steltzer, H.; Wallenstein, M.D. Predicted responses of arctic and alpine ecosystems to altered seasonality under climate change. Glob. Chang. Biol. 2014, 20, 3256–3269. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.C.; Hill, J.K.; Ohlemuller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef] [PubMed]
- Niskanen, A.K.J.; Niittynen, J.; Aalto, J.; Väre, H.; Luoto, M. Lost at high latitudes: Arctic and endemic plants under threat as climate warms. Divers. Distrib. 2019, 25, 809–821. [Google Scholar] [CrossRef]
- Kobiv, Y. Response of rare alpine plant species to climate change in the Ukrainian Carpathians. Folia Geobot. 2017, 52, 217–226. [Google Scholar] [CrossRef]
- Czortek, P.; Kapfer, J.; Delimat, A.; Eycott, A.; Grytnes, J.A.; Orczewska, A.; Ratyńska, H.; Zięba, A.; Jaroszewicz, B. Plant species composition shifts in the Tatra Mts as a response to environmental change: A resurvey study after 90 years. Folia Geobot. 2018. [Google Scholar] [CrossRef]
- Evangelista, A.; Frate, L.; Carranza, M.; Attorre, F.; Pelino, G.; Stanisci, A. Changes in composition, ecology and structure of high-mountain vegetation: A re-visitation study over 42 years. AoB Plants 2016, 8, plw004. [Google Scholar] [CrossRef] [PubMed]
- Spinoni, J.; Szalai, S.; Szentimrey, T.; Lakatos, M.; Bihari, Z.; Nagy, A.; Németh, Á.; Kovács, T.; Mihic, D.; Dacic, M.; et al. Climate of the Carpathian Region in the period 1961–2010: Climatologies and trends of 10 variables. Int. J. Clim. 2015, 35, 1322–1341. [Google Scholar] [CrossRef]
- Braun-Blanquet, J. Pflanzensoziologie. Grundzüge der Vegetationskunde, 3rd ed.; Springer: Wien, Austria, 1964; p. 631. [Google Scholar]
- Westhoff, V.; van den Maarel, E. The Braun-Blanquet approach. In Classification of Plant Communities, 1st ed.; Whittaker, R.H., Ed.; W. Junke: The Hague, The Netherlands, 1978; pp. 289–399. [Google Scholar]
- Barkman, J.J.; Doing, H.; Segal, S. Kritische Bemerkungen und Vorschläge zur quantitativen Vegetationsanalyse. Acta Bot. Neerl. 1964, 13, 394–419. [Google Scholar] [CrossRef]
- Šibík, J. Slovak Vegetation Database. In Vegetation Databases for the 21st Century; Biodiversity & Ecology; Dengler, J., Oldeland, J., Jansen, F., Chytrý, M., Ewald, J., Finckh, M., Glöckler, F., Lopez-Gonzalez, G., Peet, R.K., Schaminée, J.H.J., Eds.; BEE Press: Hamburg, Germany, 2012; Volume 4, p. 429. [Google Scholar]
- Hennekens, S.M.; Schaminée, J.H.J. TURBOVEG, a comprehensive data base management system for vegetation data. J. Veg. Sci. 2001, 12, 589–591. [Google Scholar] [CrossRef]
- Marhold, K.; Hindák, F. (Eds.) Zoznam Nižších a Vyšších Rastlín Slovenska, 1st ed.; Veda: Bratislava, Slovakia, 1998; p. 688. [Google Scholar]
- Kliment, J.; Bernátová, D.; Jarolímek, I.; Petrík, A.; Šibík, J.; Uhlířová, J. Elyno-Seslerietea. In Rastlinné Spoločenstvá Slovenska. Vysokohorská Vegetácia, 1st ed.; Kliment, J., Valachovič, M., Eds.; Veda: Bratislava, Slovakia, 2007; Volume 4, pp. 147–208. [Google Scholar]
- Kliment, J.; Šibík, J.; Šibíková, I.; Dúbravcová, Z.; Jarolímek, I.; Uhlířová, J. High-altitude vegetation of the Western Carpathians—A syntaxonomical review. Biologia 2010, 65, 965–989. [Google Scholar] [CrossRef]
- Jarolímek, I.; Šibík, J. (Eds.) Diagnostic, Constant and Dominant Taxa of the Higher Vegetation Units of Slovakia, 1st ed.; Veda: Bratislava, Slovakia, 2008; p. 332. [Google Scholar]
- GRASS Development Team. Geographic Resources Analysis Support System (GRASS) Software; Version 6.4.0; Open Source Geospatial Foundation: Chicago, IL, USA, 2010. [Google Scholar]
- Hofierka, J.; Šúri, M. The solar radiation model for Open source GIS: Implementation and applications. In Proceedings of the Open Source Free Software GIS-GRASS Users Conference 2002, Trento, Italy, 11–13 September 2002. [Google Scholar]
- Krcho, J. Zovšeobecnenie rovnice izalumklín na topografickej ploche a v jej skalárnom poli. Geogr. Časopis 1967, 19, 107–129. [Google Scholar]
- Scharmer, K.; Greif, J. (Eds.) Database and exploitation software. In The European Solar Radiation Atlas, 1st ed.; Presses des Mines: Paris, France, 2000; Volume 2, p. 296. [Google Scholar]
- Mereďa, P., Jr.; Kučera, J.; Marhold, K.; Senko, D.; Slovák, M.; Svitok, M.; Šingliarová, B.; Hodálová, I. Ecological niche differentiation between tetra- and octoploids of Jacobaea vulgaris. Preslia 2016, 88, 113–136. [Google Scholar]
- Olsen, R.S. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; US Dept. of Agriculture: Washington, DC, USA, 1954; p. 19. [Google Scholar]
- Hijmans, R.J.; Cameron, S.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Clim. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Weyant, J.; Azar, C.; Kainuma, M.; Kejun, J.; Nakicenovic, N.; Shukla, P.R.; La Rovere, E.; Yohe, G. Report of 2.6 versus 2.9 Watts/m2 RCPP Evaluation Panel; Intergovernmental Panel on Climate Change Secretariat: Geneva, Switzerland, 2009; p. 53. [Google Scholar]
- Riahi, K.; Rao, S.; Krey, V.; Cho, C.; Chirkov, V.; Fischer, G.; Kindermann, G.; Nakicenovic, N.; Rafaj, P. RCP8.5—A scenario of comparatively high greenhouse gas emissions. Clim. Chang. 2011, 109, 33–57. [Google Scholar] [CrossRef]
- Tichý, L. JUICE, software for vegetation classification. J. Veg. Sci. 2002, 13, 451–453. [Google Scholar] [CrossRef]
- Podani, J. Syn-tax 2000. In Computer Program for Data Analysis in Ecology and Systematics for Windows 95, 98 & NT. User’s Manual, 1st ed.; Scientia Publication: Budapest, Hungary, 2001; p. 53. [Google Scholar]
- Anderson, M.J.; Willis, T.J. Canonical analysis of principal coordinates: A useful method of constrained ordination for ecology. Ecology 2003, 84, 511–525. [Google Scholar] [CrossRef]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002; p. 495. [Google Scholar]
- Pinheiro, J.C.; Bates, D.M. Mixed-Effects Models in S and S-PLUS. Springer: New York, NY, USA, 2000; p. 528. [Google Scholar]
- De’ath, G.; Fabricius, K.E. Classification and regression trees: A powerful yet simple technique for ecological data analysis. Ecology 2000, 81, 3178–3192. [Google Scholar] [CrossRef]
- Hothorn, T.; Hornik, K.; Zeileis, A. Unbiased recursive partitioning: A conditional inference framework. J. Comput. Graph. Stat. 2006, 15, 651–674. [Google Scholar] [CrossRef]
- Austin, M.P.; Van Niel, K.P. Improving species distribution models for climate change studies: Variable selection and scale. J. Biogeogr. 2010, 38, 1–8. [Google Scholar] [CrossRef]
- Lomba, A.; Pellissier, L.; Randin, C.; Vicente, J.; Moreira, F.; Honrado, J.; Guisan, A. Overcoming the rare species modelling paradox: A novel hierarchical framework applied to an Iberian endemic plant. Biol. Conserv. 2010, 143, 2647–2657. [Google Scholar] [CrossRef]
- Breiner, F.T.; Guisan, A.; Bergamini, A.; Nobis, M.P. Overcoming limitations of modelling rare species by using ensembles of small models. Methods Ecol. Evol. 2015, 6, 1210–1218. [Google Scholar] [CrossRef]
- Ripley, B.D. Pattern Recognition and Neural Networks; Cambridge University Press: New York, NY, USA, 1996; p. 415. [Google Scholar]
- Elith, J.; Leathwick, J.R.; Hastie, T. A working guide to boosted regression trees. J. Anim. Ecol. 2008, 77, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.N. Generalized Additive Models: An Introduction with R, 2nd ed.; Chapman and Hall/CRC: Boca Raton, FL, USA, 2017; p. 496. [Google Scholar]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Breiner, F.T.; Nobis, M.P.; Bergamini, A.; Guisan, A. Optimizing ensembles of small models for predicting the distribution of species with few occurrences. Methods Ecol. Evol. 2018, 9, 802–808. [Google Scholar] [CrossRef]
- Kuhn, M.; Johnson, K. Applied Predictive Modeling; Springer: New York, NY, USA, 2013; p. 600. [Google Scholar]
- QGIS Development Team. QGIS Geographic Information System, Version 3.6.0.; Open Source Geospatial Foundation Project: Beaverton, OR, USA, 2019. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Broennimann, O.; Di Cola, V.; Guisan, A. Ecospat: Spatial Ecology Miscellaneous Methods; Version 3.0. R Package; 2018; Available online: https://rdrr.io/cran/ecospat/ (accessed on 18 September 2019).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. nlme: Linear and Nonlinear Mixed Effects Models; Version 3.1-141; R Package; 2019; Available online: https://cran.r-project.org/web/packages/nlme/citation.html (accessed on 18 September 2019).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P. vegan: Community Ecology Package; Version 2.3-5; R Package; 2017; Available online: https://rdrr.io/rforge/vegan/ (accessed on 18 September 2019).
- Šibík, J.; Petriík, A.; Kliment, J. Syntaxonomical revision of plant communities with Carex firma and Dryas octopetala (alliance Caricion firmae) in the Western Carpathians. Pol. Bot. J. 2004, 49, 181–202. [Google Scholar]
- Holub, J. Carex rupestris All. Sborn. Národ. Muz. 1972, 27, 25–26. [Google Scholar]
- Fudali, E.; Kučera, J. Bryogeographical elements of moss flora in glacial cirques “Śnieżne Kotły” (Karkonosze Mts) and their threat. Acta Soc. Bot. Pol. 2003, 72, 79–85. [Google Scholar] [CrossRef][Green Version]
- Birks, H.H. The late-quaternary history of arctic and alpine plants. Plant Ecol. Divers. 2008, 1, 135–146. [Google Scholar] [CrossRef]
- Feurdean, A.; Tămaş, T.; Tanţău, I.; Fărcaş, S. Elevational variation in regional vegetation responses to late-glacial climate changes in the Carpathians. J. Biogeogr. 2012, 39, 258–271. [Google Scholar] [CrossRef]
- Kliment, J.; Šibíková, I.; Šibík, J. On the occurrence of the arctic-alpine and endemic species in the high-altitude vegetation of the Western Carpathians. Thaiszia J. Bot. 2011, 21, 45–60. [Google Scholar]
- Alsos, I.G.; Müller, E.; Eidesen, P.B. Germinating seeds or bulbils in 87 of 113 tested Arctic species indicate potential for ex situ seed bank storage. Polar Biol. 2013, 36, 819–830. [Google Scholar] [CrossRef]
- Chlebicki, A. Grzyby mikroskopijne na reliktowych stanowiskach turzycy skalnej Carex rupertris v Karpatach. Rocz. Bieszcz. 2000, 9, 61–68. [Google Scholar]
- Searcy, K.B.; Wilson, B.F.; Fownes, J.H. Influence of bedrock and aspect on soils and plant distribution in the Holyoke Range, Massachusetts. J. Torrey Bot. Soc. 2003, 130, 158–169. [Google Scholar] [CrossRef]
- Vonlanthen, C.M.; Kammer, P.M.; Eugster, W.; Bühler, A.; Veit, H. Alpine vascular plant species richness: The importance of daily maximum temperature and pH. Plant Ecol. 2006, 184, 13–25. [Google Scholar] [CrossRef]
- Lunde, T. An investigation into the pH-amplitude of some mountain plants in the county of Troms. Acta Boreal. 1962, 20, 1–103. [Google Scholar]
- Gensac, P. Plant and soil groups in the alpine grasslands of the Vanoise Massif, French Alps. Arct. Alp. Res. 1990, 22, 195–201. [Google Scholar] [CrossRef]
- Gough, L.; Shaver, G.R.; Carol, J.; Royer, D.L.; Laundre, J.A. Vascular plant species richness in Alaskan arctic tundra: The importance of soil pH. J. Ecol. 2000, 88, 54–66. [Google Scholar] [CrossRef]
- Schmidtlein, S.; Ewald, J. Landscape patterns of indicator plants for soil acidity in the Bavarian Alps. J. Biogeogr. 2003, 30, 1493–1503. [Google Scholar] [CrossRef]
- Darmody, R.G.; Thorn, C.E.; Schlyter, P.; Dixon, J.C. Relationship of vegetation distribution to soil propertiesin Kärkevagge, Swedish Lapland. Arct. Antarct. Alp. Res. 2004, 36, 21–32. [Google Scholar] [CrossRef]
- Thuiller, W.; Albert, C.; Araujo, M.B.; Berry, P.M.; Cabeza, M.; Guisan, A.; Hickler, T.; Midgley, G.F.; Paterson, J.; Schurr, F.M.; et al. Predicting global change impacts on plant species’ distributions: Future challenges. Perspect. Plant Ecol. 2008, 9, 137–152. [Google Scholar] [CrossRef]
- Chardon, N.I.; Wipf, S.; Rixen, C.; Beilstein, A.; Doak, D.F. Local trampling disturbance effects on alpine plant populations and communities: Negative implications for climate change vulnerability. Ecol. Evol. 2018, 8, 7921–7935. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; May, R.M.; Lehman, C.L.; Nowak, M.A. Habitat destruction and the extinction debt. Nature 1994, 371, 65–66. [Google Scholar] [CrossRef]
- Kuussaari, M.; Bommarco, R.; Heikkinen, R.K.; Helm, A.; Krauss, J.; Lindborg, R.; Öckinger, E.; Pärtel, M.; Pino, J.; Rodà, F.; et al. Extinction debt: A challenge for biodiversity conservation. Trends Ecol. Evol. 2009, 24, 564–571. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
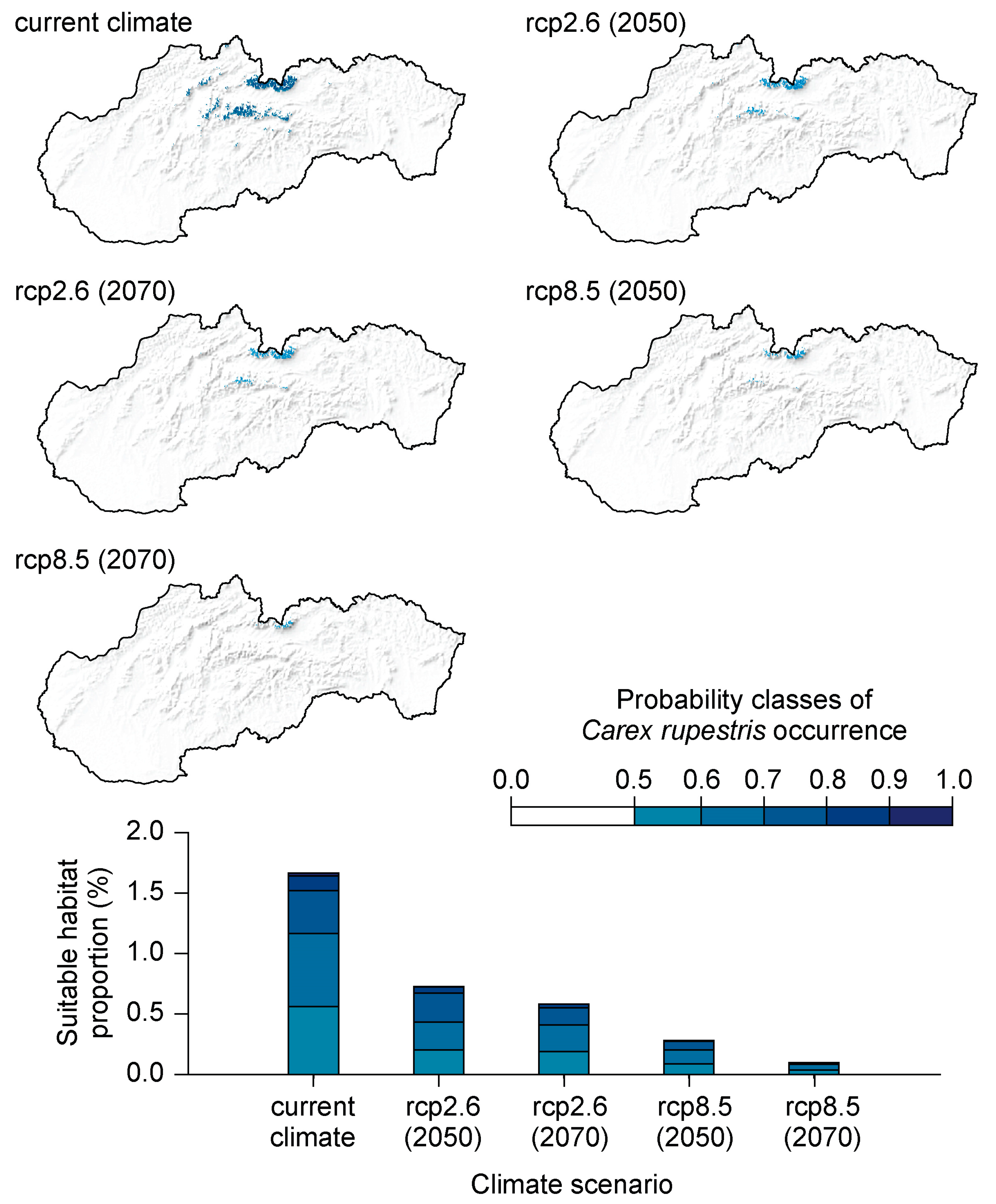{kind=link}
| Variable | Unit | Range | F/χ2 | P | Cluster A | Cluster B | Cluster C | Cluster D |
|---|---|---|---|---|---|---|---|---|
| Vegetation data | ||||||||
| Nr. of relevés | 27 | 10 | 17 | 11 | ||||
| Species richness | 14–50 | 10.43 | <0.0001 | 23.5 (21.3–25.8) a | 27.1 (19.1–35.2) a | 36.1 (32.3–39.8) b | 29.6 (26.6–32.7) ab | |
| Topology | ||||||||
| Altitude | m | 1297–2012 | 89.24 | <0.0001 | 1455 (1425–1485) a | 1652 (1597–1708) b | 1862 (1805–1919) c | 1930 (1862–1997) c |
| Slope | ° | 10–80 | 14.13 | <0.0001 | 47 (43–51) a | 43 (34–51) a | 54 (48–60) a | 25 (18–33) b |
| Aspect | 0.04–3.47 | 0.15 | 0.9319 | 1.99 (1.77–2.22) | 1.39 (0.38–2.40) | 2.09 (1.66–2.52) | 1.22 (0.43–2.01) | |
| Geological bedrock | % lime/marl/mylonite | – | 85.63 | <0.0001 | 100/0/0 a | 62/38/0 b | 0/6/94 c | 45/55/0 b |
| Climate | ||||||||
| Temperature | °C | 0.35–4.07 | 111.6 | <0.0001 | 3.0 (2.8–3.2) a | 2.1 (1.7–2.5) b | 0.5 (0.4–0.7) c | 1.3 (1.2–1.4) d |
| Precipitation | mm | 1283–2123 | 124.4 | <0.0001 | 1431 (1403–1458) a | 1463 (1351–1574) a | 1938 (1899–1977) b | 1809 (1755–1863) c |
| Solar radiation | ||||||||
| Duration | h·m−2·y−1 | 10–140 | 5.59 | 0.0019 | 120 (114–125) a | 96 (84–109) ab | 100 (79–121) ab | 83 (74–92) b |
| Global radiation | MWh·m−2·y−1 | 16–76 | 4.68 | 0.0054 | 67 (62–71) a | 57 (45–69) ab | 50 (40–60) b | 53 (46–61) ab |
| Direct radiation | MWh·m−2·y−1 | 0.2–55 | 3.75 | 0.0156 | 48 (44–52) a | 40 (30–50) ab | 35 (26–43) b | 36 (30–43) ab |
| Diffused radiation | MWh·m−2·y−1 | 0.01–19.5 | 16.85 | <0.0001 | 18.3 (17.7–18.9) a | 16.0 (14.6–17.5) b | 13.8 (11.7–15.8) b | 15.8 (14.8–16.7) b |
| Reflected radiation | MWh·m−2·y−1 | 0.03–16.93 | 3.33 | 0.0256 | 0.98 (0.80–1.16) ab | 1.37 (0.85–1.89) ab | 1.55 (0.36–3.45) a | 1.34 (1.11–1.58) b |
| Soil characteristics | ||||||||
| pHH20 | – | 4.30–7.75 | 41.64 | <0.0001 | 7.39 (7.26–7.53) a | 7.11 (6.77–7.44) ac | 5.54 (5.21–5.86) b | 6.53 (5.95–7.11) c |
| pHKCl | – | 4.03–7.69 | 39.91 | <0.0001 | 7.28 (7.09–7.46) a | 7.11 (6.76–7.45) ac | 5.29 (4.93–5.66) b | 6.24 (5.86–6.62) c |
| Ca | g·kg−1 | 0.6–12.4 | 9.389 | 0.0001 | 6.2 (4.9–7.4) a | 6.4 (4.1–8.7) a | 2.3 (1.4–3.1) b | 6.8 (5.2–8.4) a |
| K | mg·kg−1 | 47–340 | 6.887 | 0.0008 | 189 (157–221) a | 145 (106–183) ab | 103 (76–130) b | 122 (67–176) ab |
| Mg | mg·kg−1 | 14–2148 | 45.44 | <0.0001 | 1210 (956–1463) a | 615 (58–1172) b | 48 (23–72) c | 144 (61–226) b |
| Ctot | % | 2.8–30.7 | 19.35 | <0.0001 | 22.1 (20.2–24.0) a | 20.2 (15.7–24.7) a | 9.3 (6.6–12.0) b | 18.4 (13.6–23.2) a |
| N | % | 0.23–2.65 | 8.665 | 0.0002 | 1.56 (1.29–1.83) a | 1.37 (1.00–1.74) a | 0.70 (0.51–0.89) b | 1.39 (0.98–1.79) ab |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Svitková, I.; Svitok, M.; Petrík, A.; Bernátová, D.; Senko, D.; Šibík, J. The Fate of Endangered Rock Sedge (Carex rupestris) in the Western Carpathians—The Future Perspective of an Arctic-Alpine Species under Climate Change. Diversity 2019, 11, 172. https://doi.org/10.3390/d11090172
Svitková I, Svitok M, Petrík A, Bernátová D, Senko D, Šibík J. The Fate of Endangered Rock Sedge (Carex rupestris) in the Western Carpathians—The Future Perspective of an Arctic-Alpine Species under Climate Change. Diversity. 2019; 11(9):172. https://doi.org/10.3390/d11090172
Chicago/Turabian StyleSvitková, Ivana, Marek Svitok, Anton Petrík, Dana Bernátová, Dušan Senko, and Jozef Šibík. 2019. "The Fate of Endangered Rock Sedge (Carex rupestris) in the Western Carpathians—The Future Perspective of an Arctic-Alpine Species under Climate Change" Diversity 11, no. 9: 172. https://doi.org/10.3390/d11090172
APA StyleSvitková, I., Svitok, M., Petrík, A., Bernátová, D., Senko, D., & Šibík, J. (2019). The Fate of Endangered Rock Sedge (Carex rupestris) in the Western Carpathians—The Future Perspective of an Arctic-Alpine Species under Climate Change. Diversity, 11(9), 172. https://doi.org/10.3390/d11090172