Implications of Spatial Habitat Diversity on Diet Selection of European Bison and Przewalski’s Horses in a Rewilding Area


Abstract
:1. Introduction
2. Materials and Methods
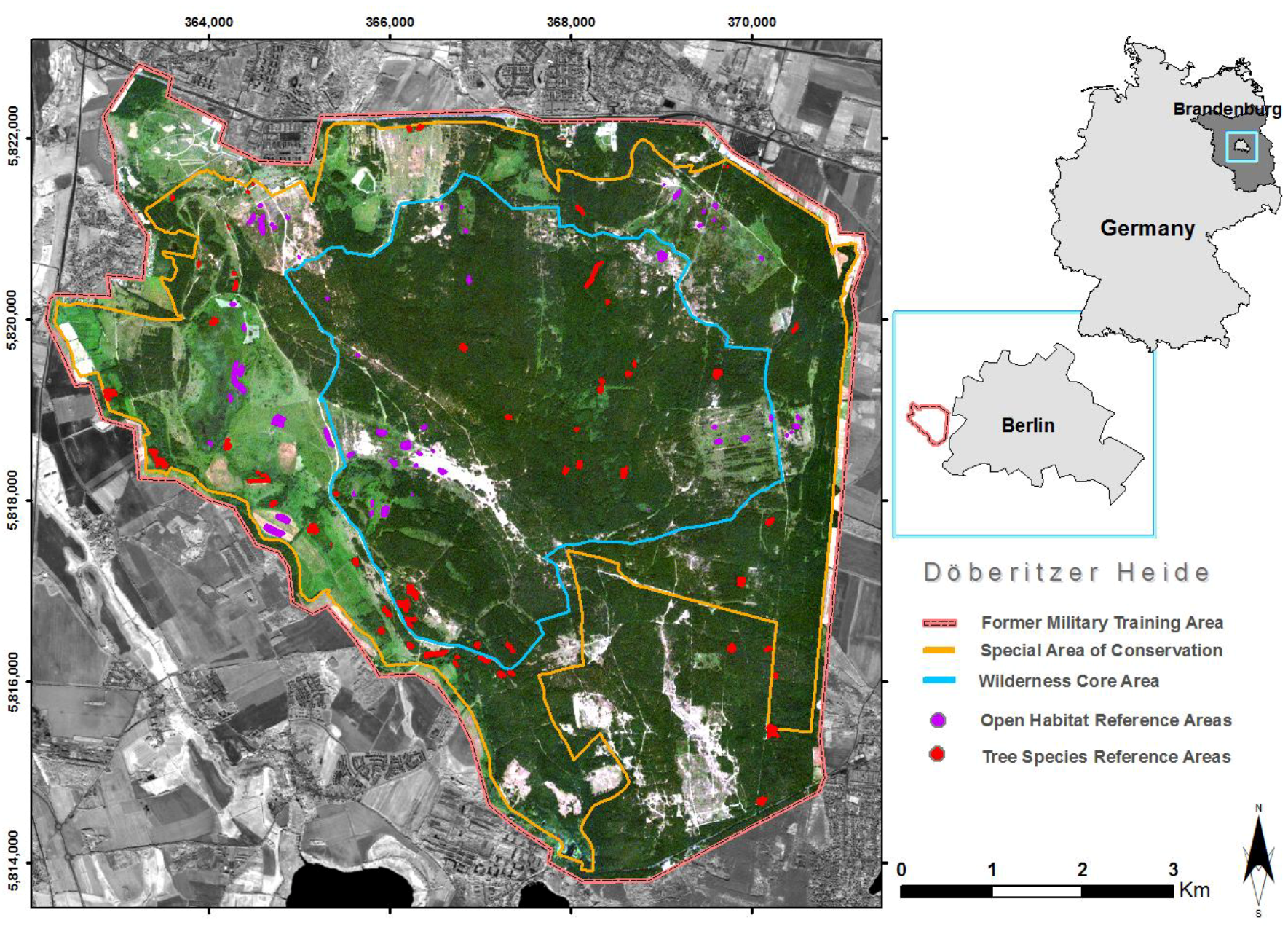
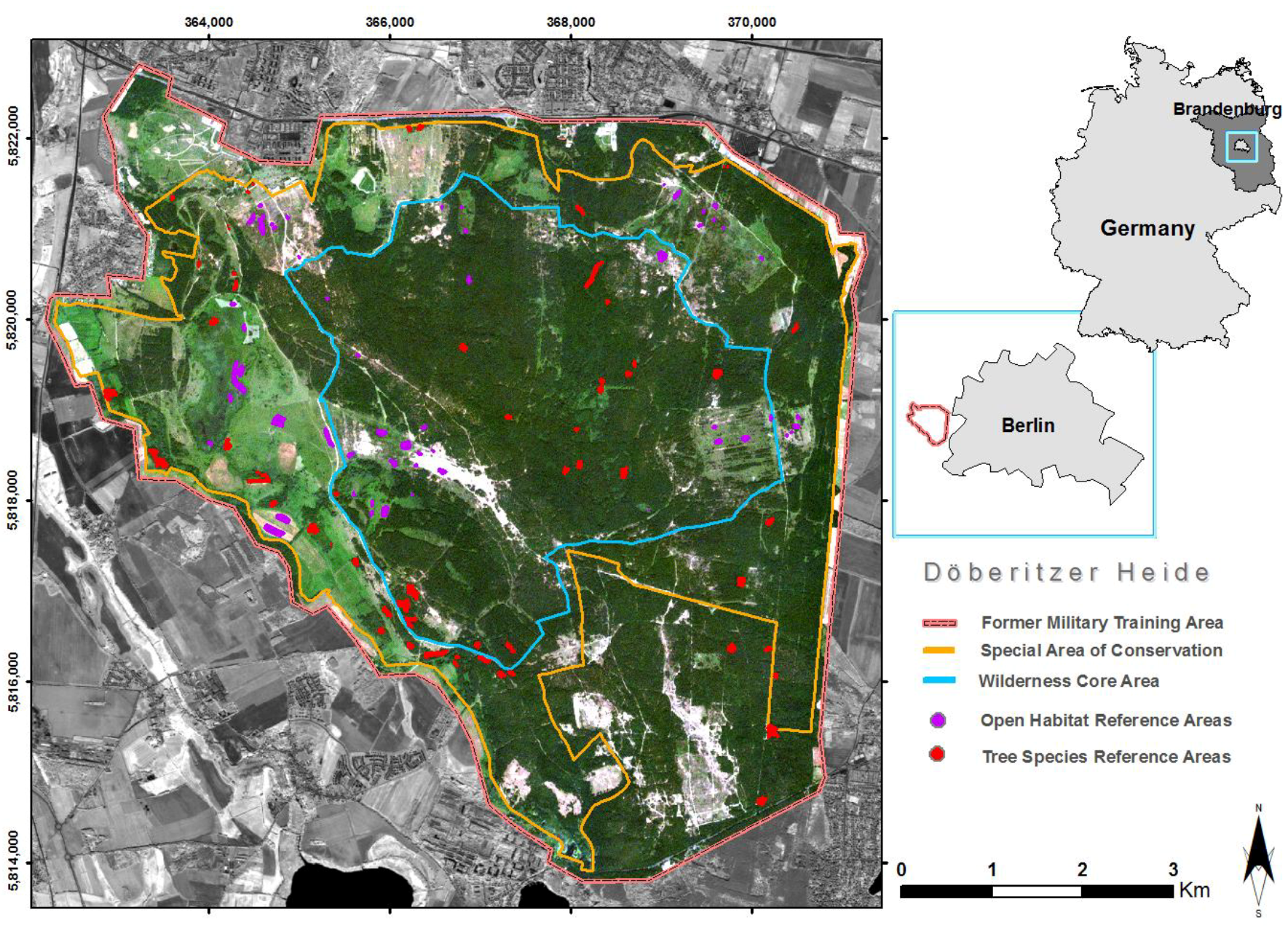
2.1. Study Area
2.2. Diet Selection
2.3. Satellite Imagery
2.4. Habitat Categorization
2.5. Spectral Habitat Modeling
2.6. Mapping of Potential Habitat Preferences
2.7. Diet Data Analyses
3. Results
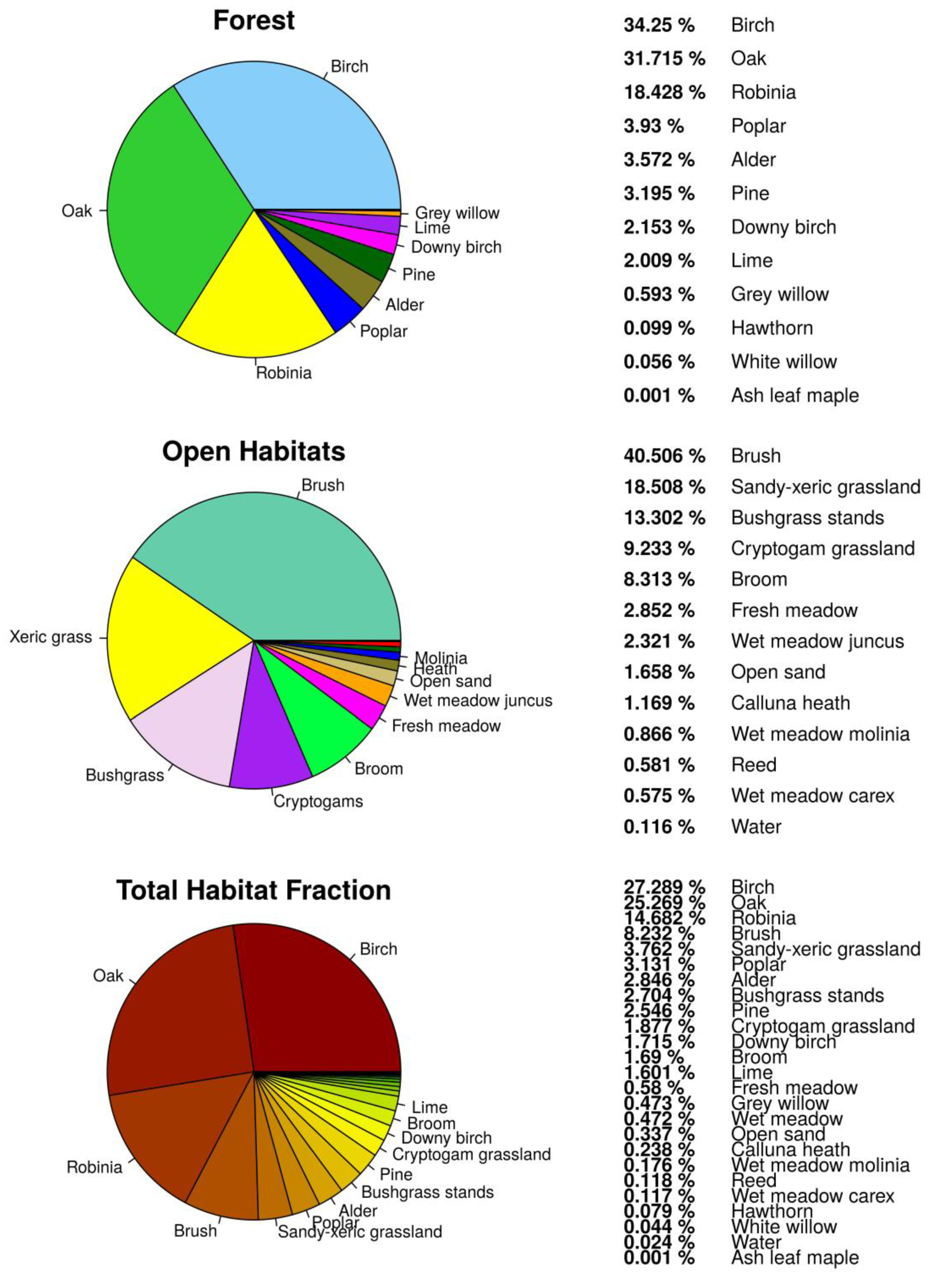
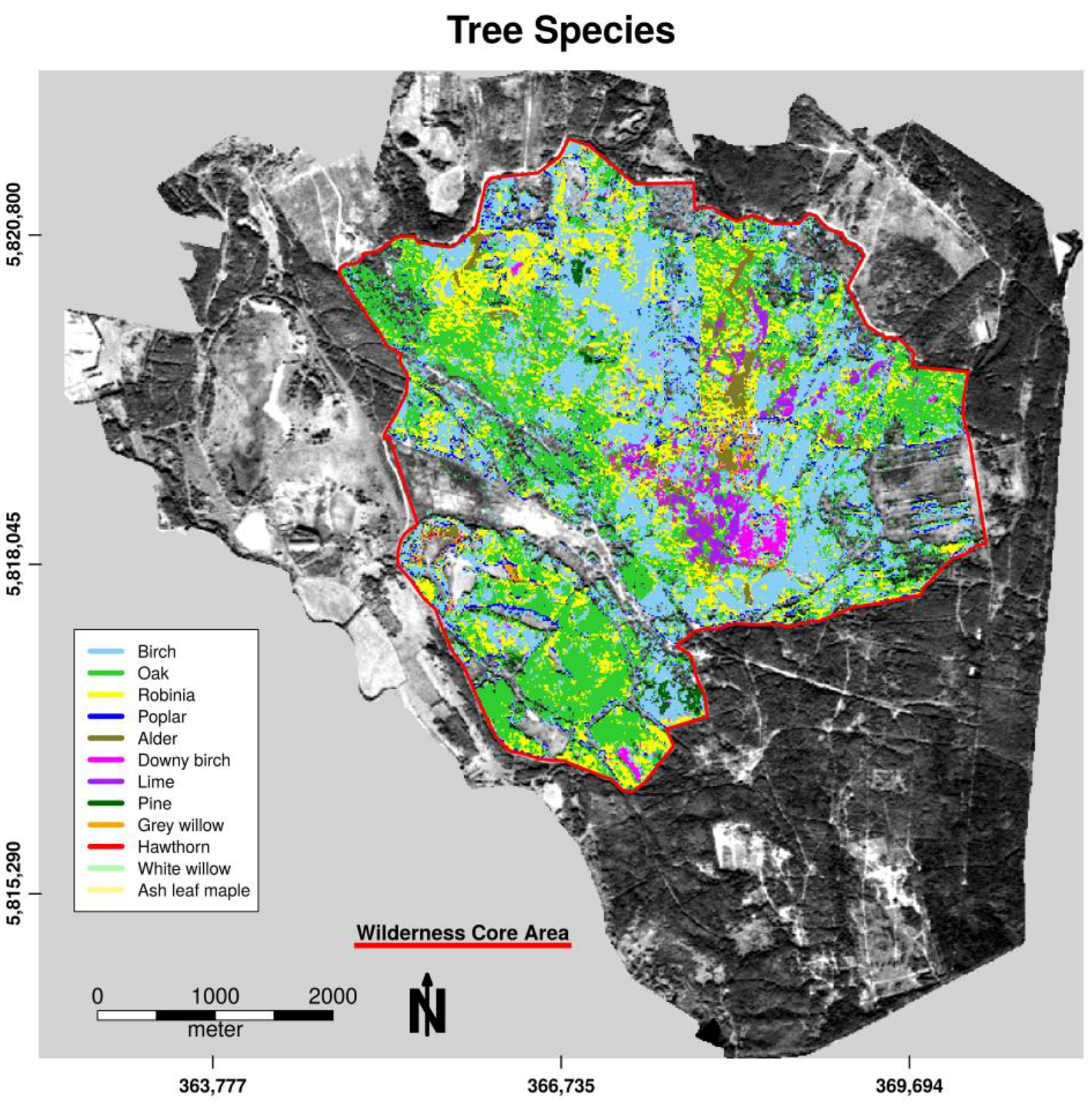
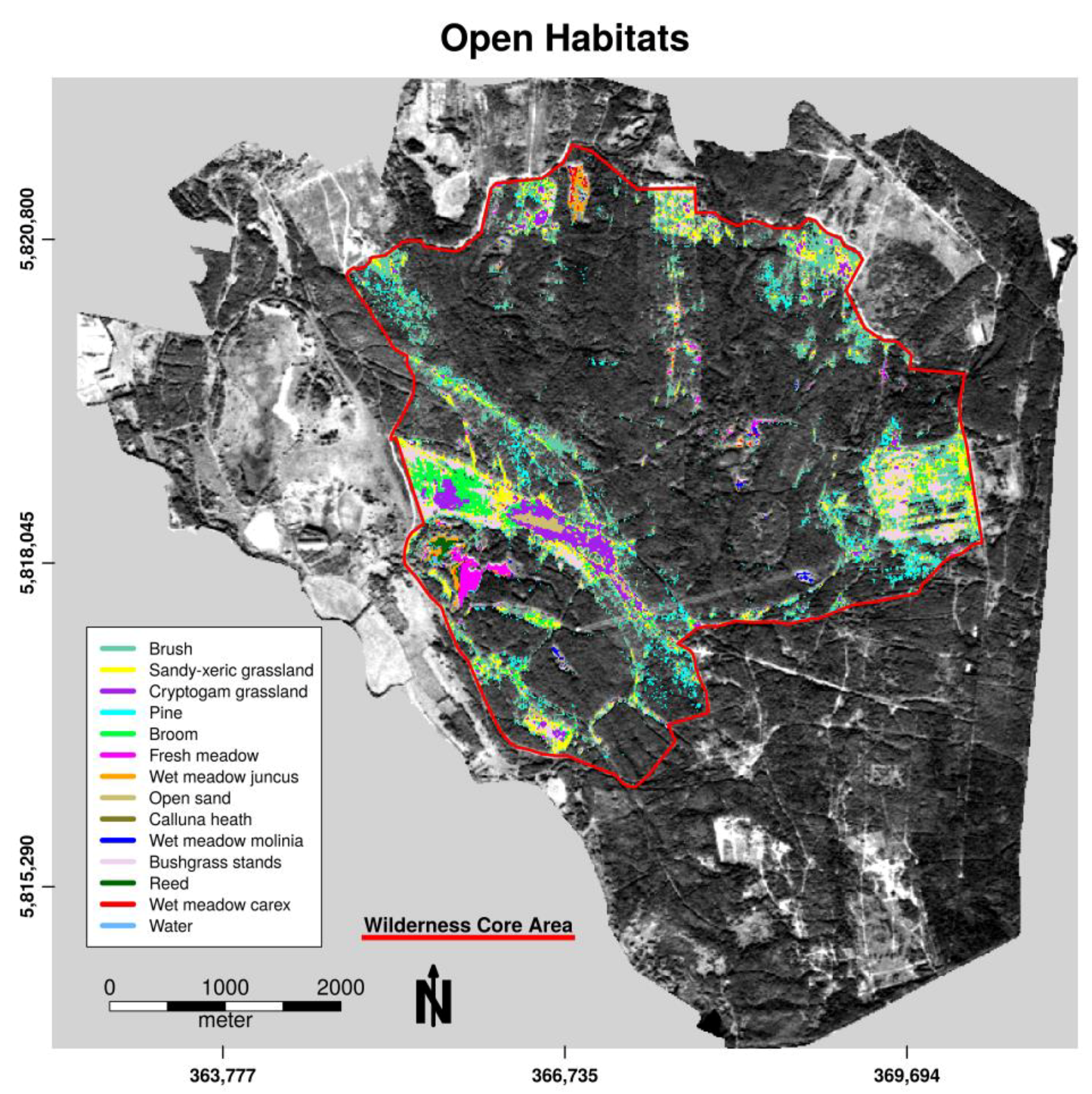
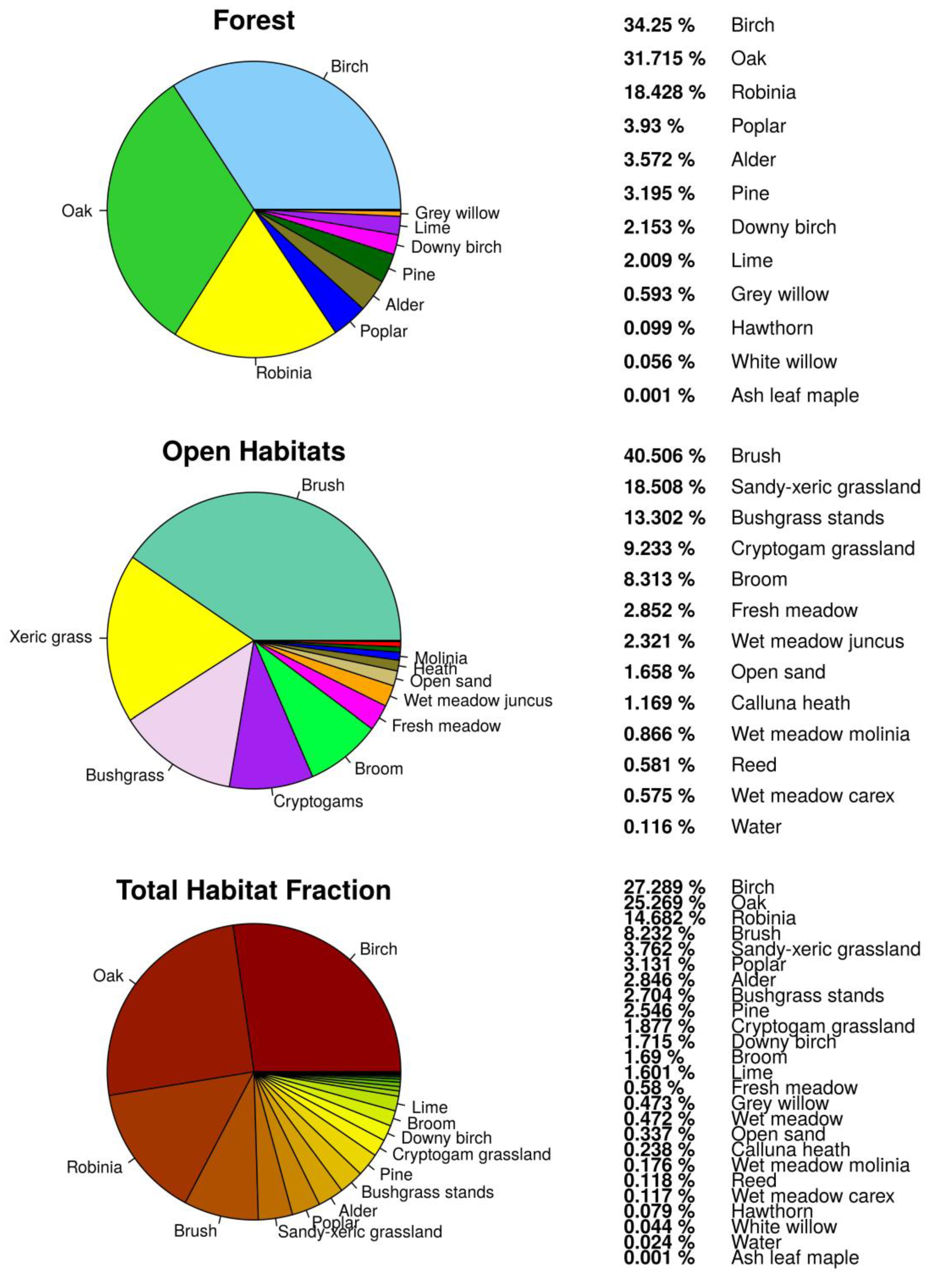
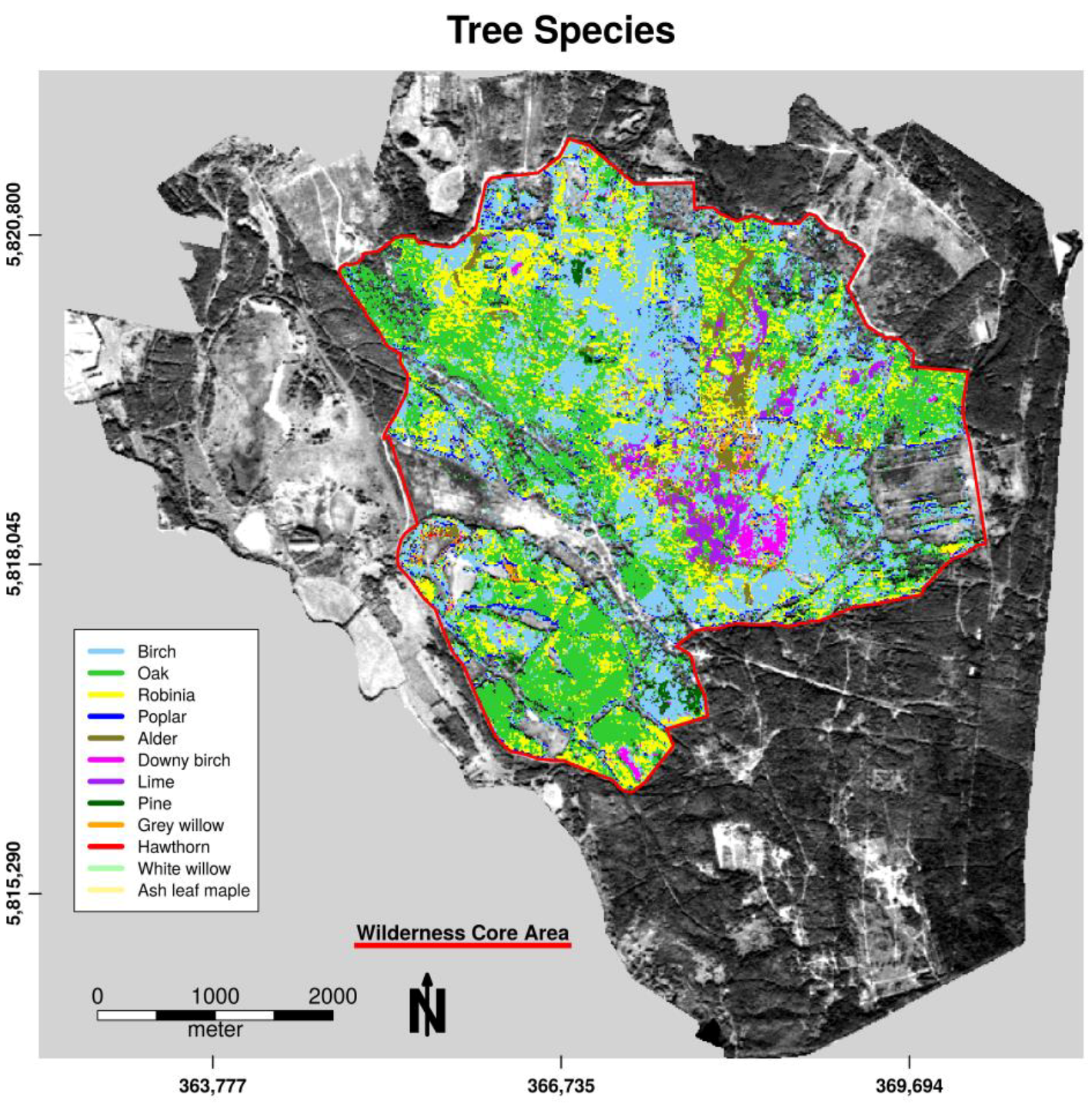
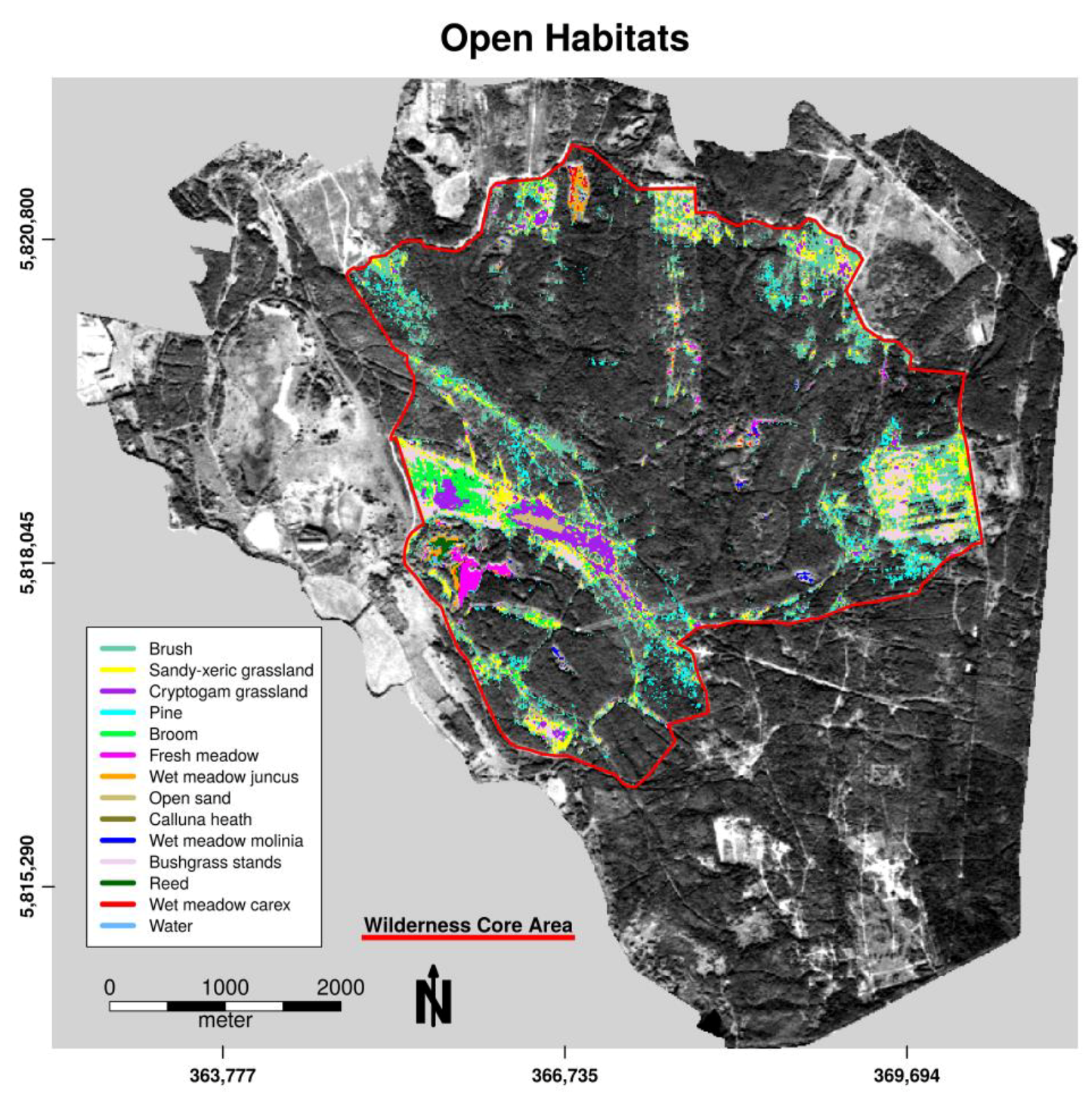
3.1. Habitat Classification
3.2. Habitat Distribution within the Study Area
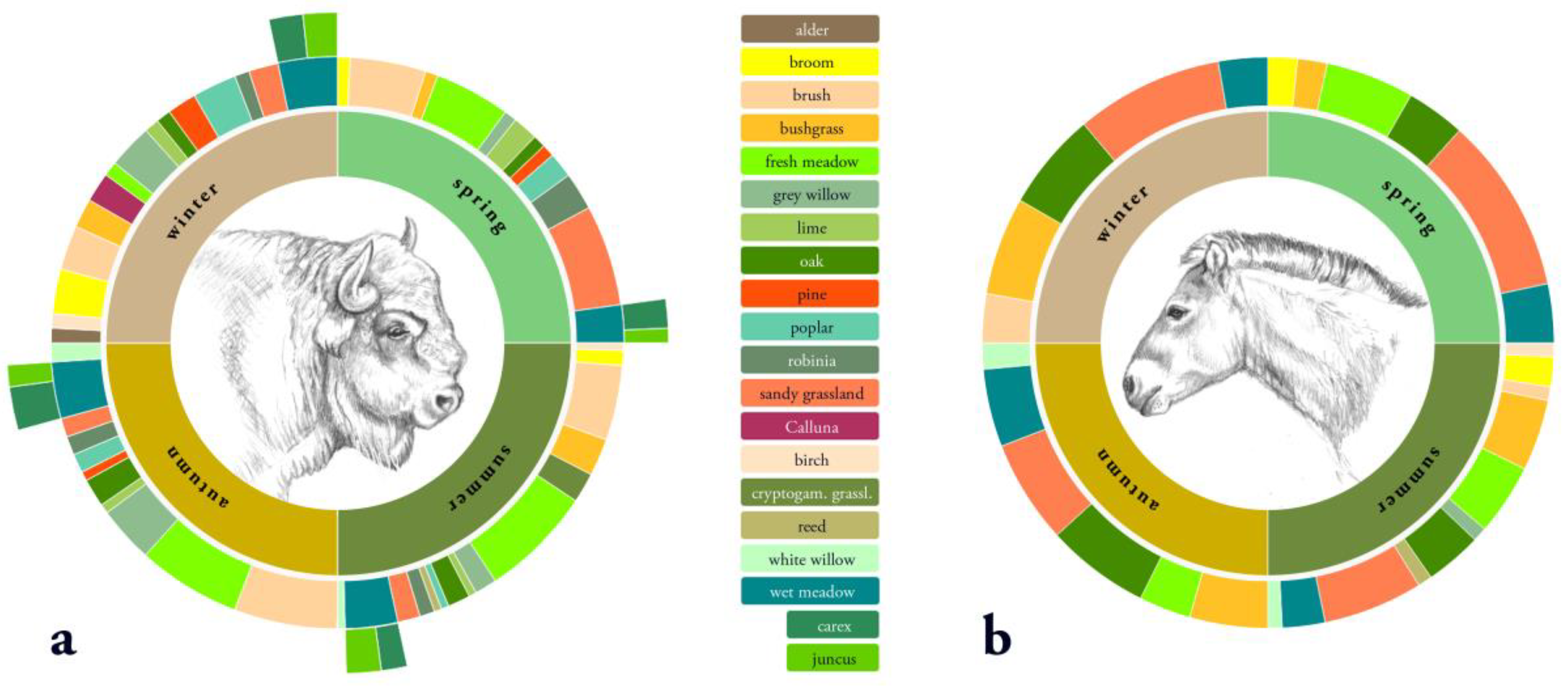
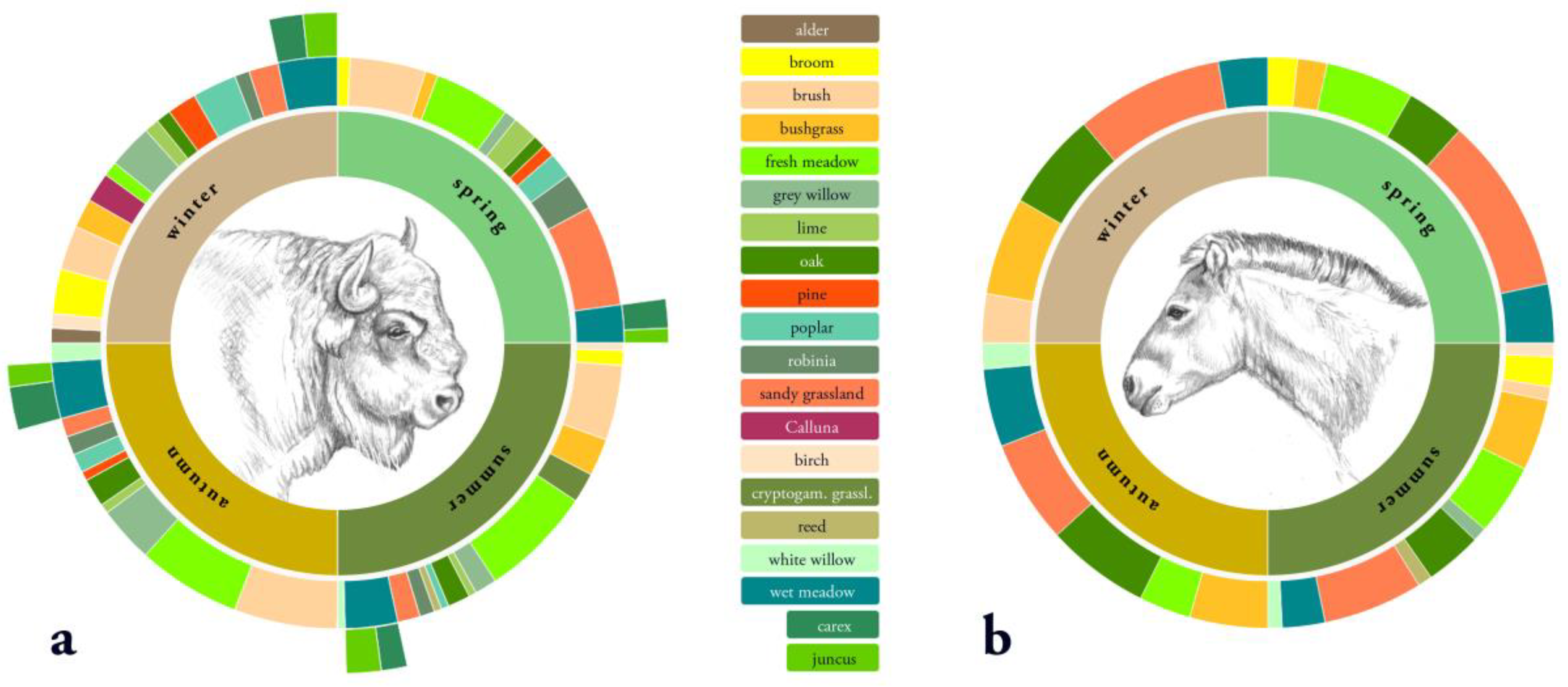
3.3. Diet Selection
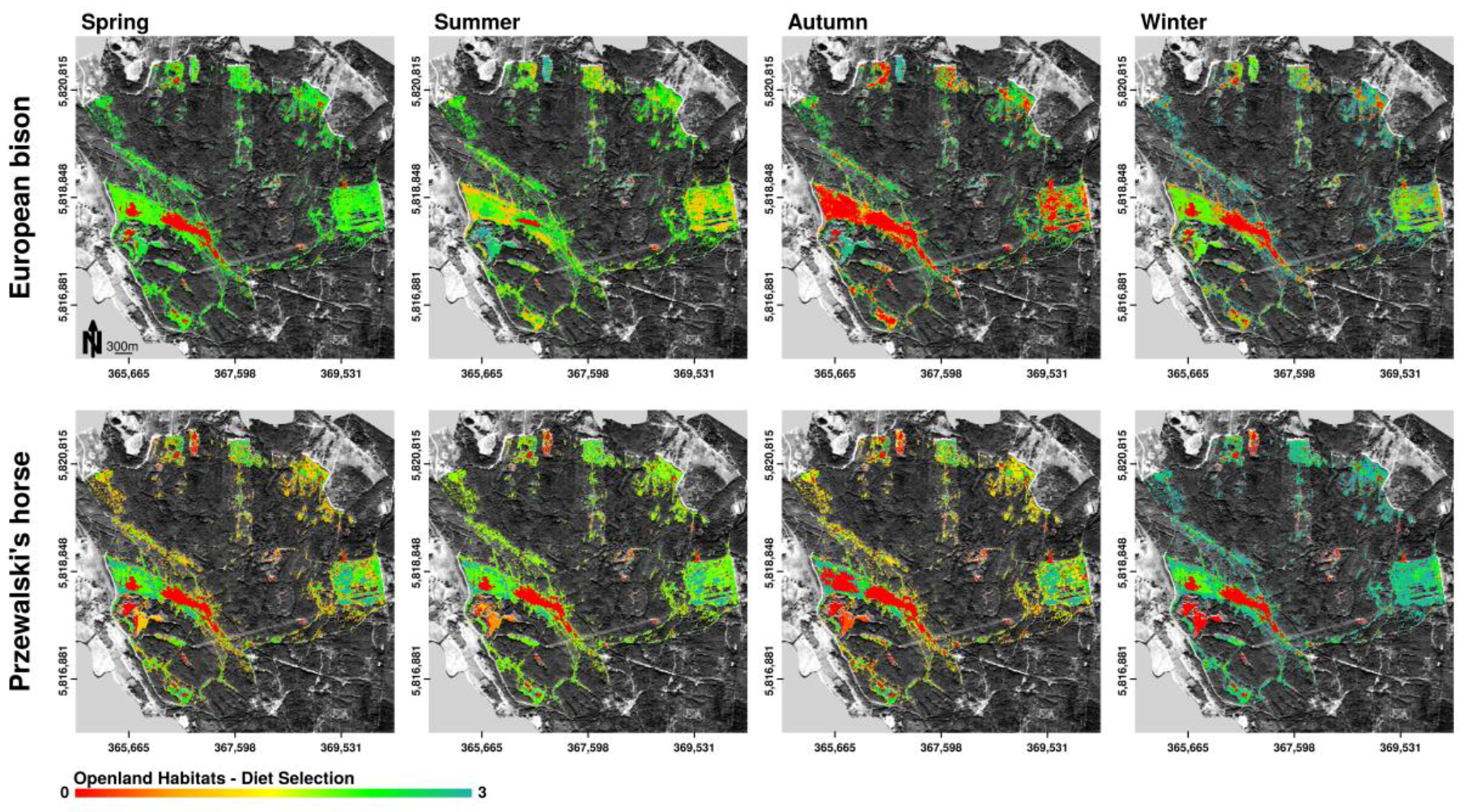
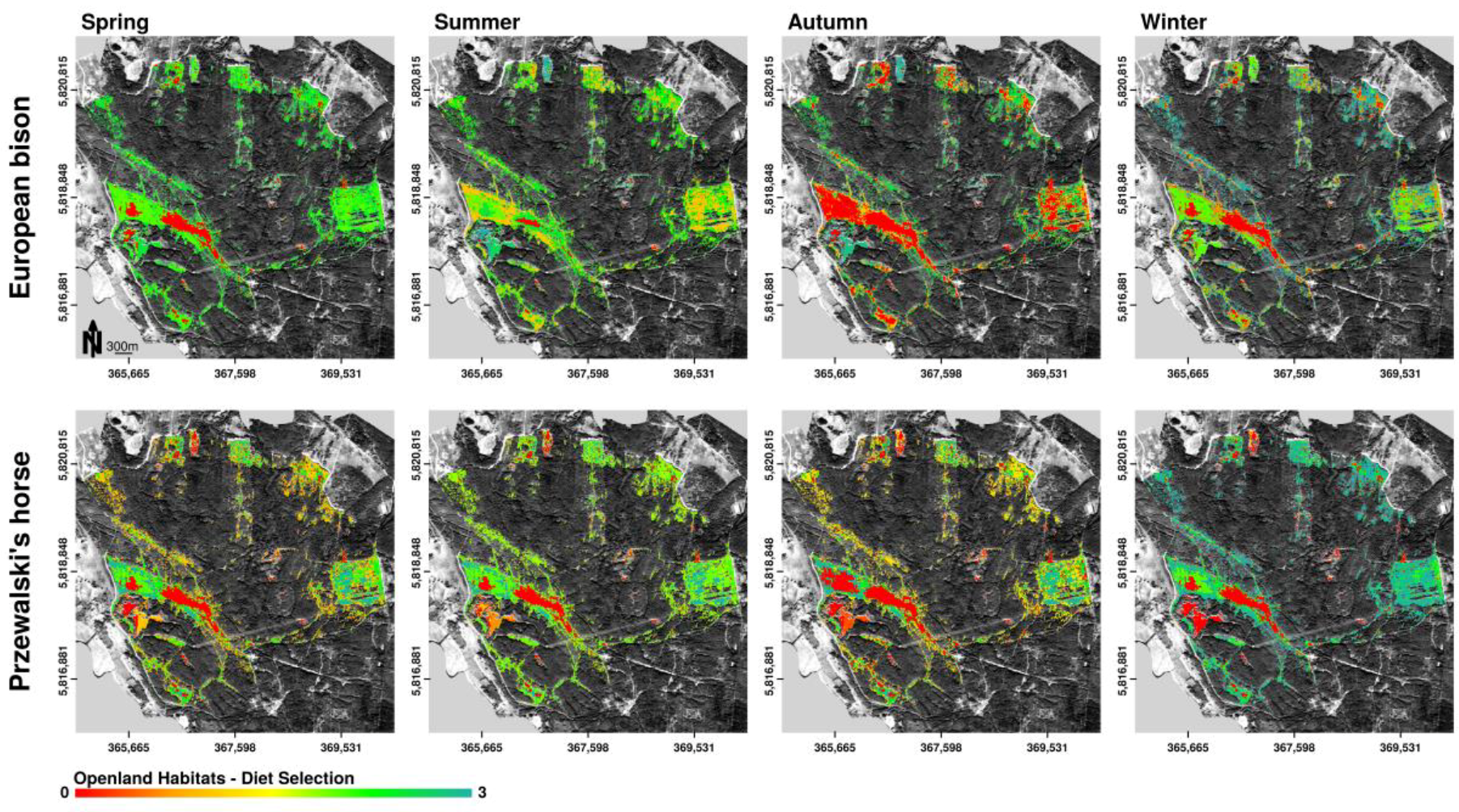
3.3.1. Diet Selection Openland Habitats
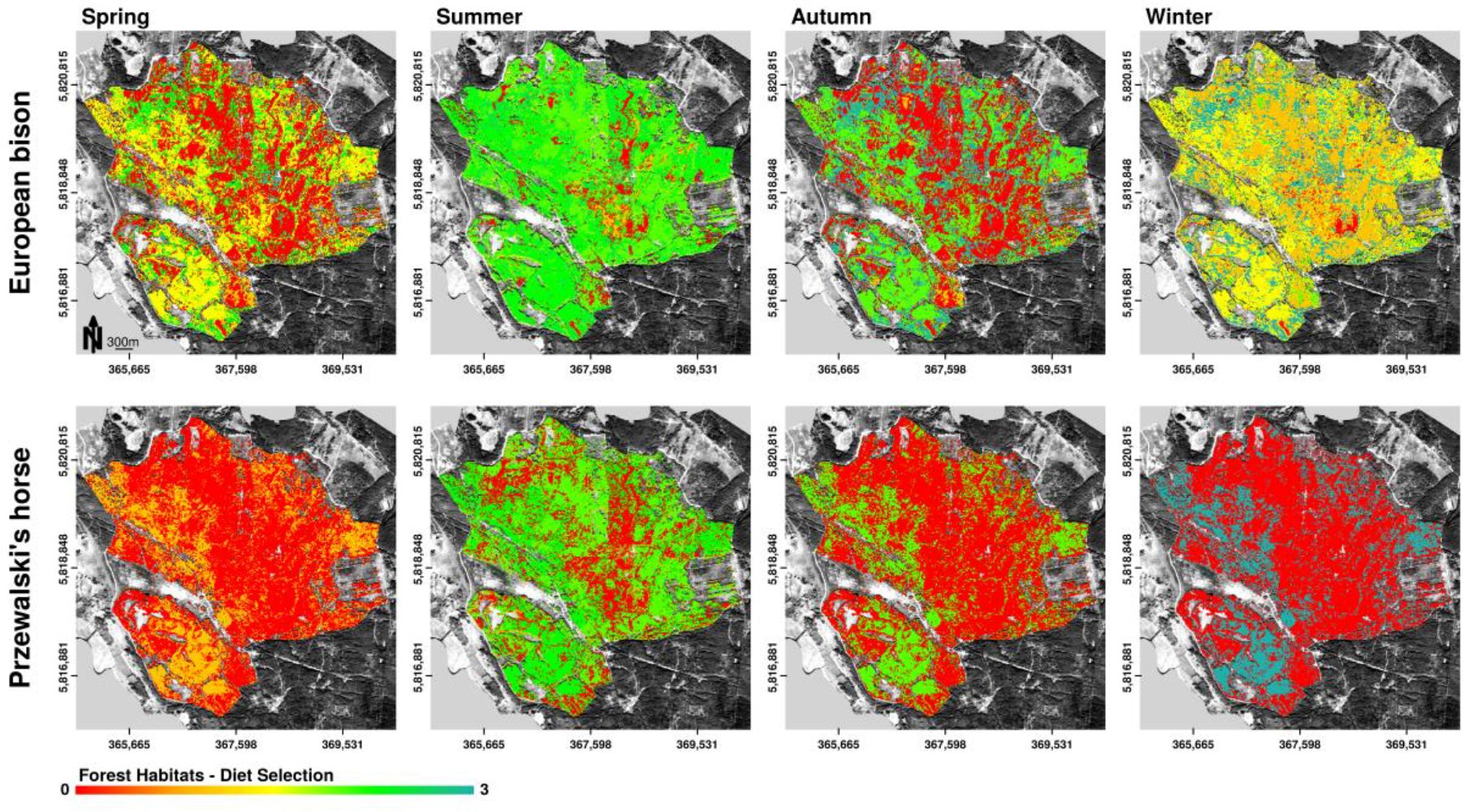
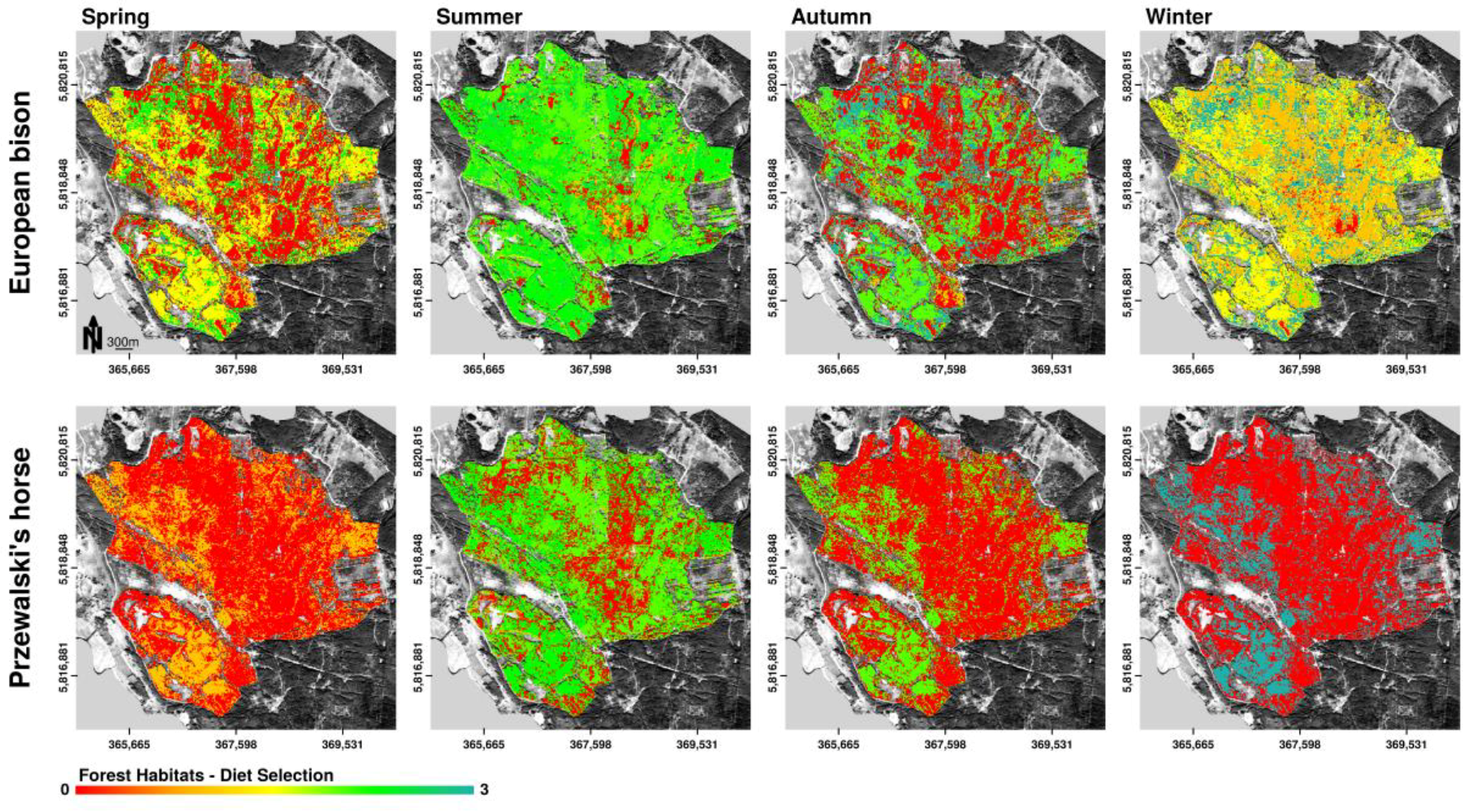
3.3.2. Diet Selection Forest Habitats
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Aaris-Sørensen, K.; Mühldorff, R.; Petersen, E.B. The Scandinavian reindeer (Rangifer tarandus L.) after the last glacial maximum: Time, seasonality and human exploitation. J. Archaeol. Sci. 2007, 34, 914–923. [Google Scholar]
- Sommer, R.S.; Benecke, N.; Lõugas, L.; Nelle, O.; Schmölcke, U. Holocene survival of the wild horse in Europe: A matter of open landscape? J. Quat. Sci. 2011, 26, 805–812. [Google Scholar] [CrossRef]
- Svenning, J.C. A review of natural vegetation openness in north-western Europe. Biol. Conserv. 2002, 104, 133–148. [Google Scholar] [CrossRef]
- Gordon, I.J.; Hester, A.J.; Festa-Bianchet, M. The management of wild large herbivores to meet economic, conservation and environmental objectives. J. Appl. Ecol. 2004, 41, 1021–1031. [Google Scholar] [CrossRef]
- Kuijper, D.P.J.; Cromsigt, J.P.M.G.; Churski, M.; Adams, B.; Jedrzejewska, B.; Jedrzejewski, W. Do ungulates preferentially feed in forest gaps in European temperate forests? For. Ecol. Manag. 2009, 258, 1528–1535. [Google Scholar] [CrossRef]
- Griffiths, C.; Harris, S. Prevention of Secondary Extinctions through Taxon Substitution. Conserv. Biol. 2010, 24, 645–646. [Google Scholar] [CrossRef] [PubMed]
- Svenning, J.C.; Pedersen, P.B.M.; Donlan, C.J.; Ejrnӕs, R.; Faurby, S.; Galetti, M.; Hansen, D.M.; Sandel, B.; Sandom, C.J.; Terborgh, J.W.; et al. Science for a wilder Anthropocene: Synthesis and future directions for trophic rewilding research. Proc. Natl. Acad. Sci. USA 2016, 113, 898–906. [Google Scholar] [CrossRef]
- Owen-Smith, R.N. Megaherbivores: The Influence of Very Large Body Size on Ecology; Cambridge University Press: Cambridge, UK, 1988. [Google Scholar]
- Olff, H.; Ritchie, M.E. Effects of herbivores on grassland plant diversity. Tree 1998, 13, 261–265. [Google Scholar] [CrossRef]
- Beever, E.A.; Tausch, R.J.; Thogmartin, W.E. Multi-scale responses of vegetation to removal of horse grazing from Great Basin (USA) mountain ranges. Plant Ecol. 2008, 196, 163–184. [Google Scholar] [CrossRef]
- Estes, J.A.; Terborgh, J.; Brashares, J.S.; Power, M.E.; Berger, J.; Bond, W.J.; Carpenter, S.R.; Essington, T.S.; Holt, R.D.; Jackson, J.B.C.; et al. Trophic downgrading of planet Earth. Science 2011, 333, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Schwark, L.; Zink, K.; Lechterbeck, J. Reconstruction of postglacial to early Holocene vegetation history in terrestrial central Europe via cuticular lipid biomarkers and pollen records from lake sediments. Geology 2002, 30, 463–466. [Google Scholar] [CrossRef]
- Birks, H.J.B. Mind the gap: How open were European primeval forests? Trends Ecol. Evol. 2005, 20, 154–156. [Google Scholar] [CrossRef]
- Kowalczyk, R.; Taberlet, P.; Coissac, E.; Valentini, A.; Miquel, C.; Kaminski, T.; Wojcik, J.M. Influence of management practices on large herbivore diet-Case of European bison in Białowieża Primeval Forest (Poland). For. Ecol. Manag. 2011, 261, 821–828. [Google Scholar] [CrossRef]
- Naundrup, P.J.; Svenning, J.C. A Geographic Assessment of the Global Scope for Rewilding with Wild-Living Horses (Equus ferus). PLoS ONE 2015, 10, e0132359. [Google Scholar] [CrossRef] [PubMed]
- Soule, M.; Noss, R. Rewilding and biodiversity: Complementary goals for continental conservation. Wild Earth 1998, 8, 1–11. [Google Scholar]
- Schumacher, H.; Finck, P.; Klein, M.; Ssymank, A.; Paulsch, C. Wildnis im Dialog—Wildnis und Natura 2000; BfN-Skripten 452; Bundesamt für NaturschutzKonstantinstr: Bonn, Germany, 2017; p. 126. [Google Scholar]
- Danell, K.; Bergström, R.; Duncan, P.; Pastor, J. Large Herbivore Ecology, Ecosystem Dynamics and Conservation; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Waldram, M.S.; Bond, W.J.; Stock, W.D. Ecological engineering by a mega-grazer: White rhino impacts on a South African savanna. Ecosystems 2008, 11, 101–112. [Google Scholar] [CrossRef]
- Haynes, G. Elephants (and extinct relatives) as earth-movers and ecosystem engineers. Geomorphology 2012, 157–158, 99–107. [Google Scholar] [CrossRef]
- Jaroszewicz, B.; Pirożnikow, E. Diversity of plant species eaten and dispersed by the European bison Bison bonasus in the Białowieża Forest. Eur. Bison Conserv. Newsl. 2008, 1, 14–29. [Google Scholar]
- Finck, P.; Riecken, U.; Schröder, E. Pasture Landscapes and Nature Conservation—New strategies for the preservation of open landscapes in Europe. In Pasture Landscapes and Nature Conservation; Springer: Berlin/Heidelberg, Germany, 2002; pp. 1–13. [Google Scholar]
- Menard, C.; Duncan, P.; Fleurance, G.; Georges, J.Y.; Lila, M. Comparative foraging and nutrition of horses and cattle in European wetlands. J. Appl. Ecol. 2002, 39, 120–133. [Google Scholar] [CrossRef]
- Kerley, G.I.H.; Kowalczyk, R.; Cromsigt, J.P.G.M. Conservation implications of the refugee species concept and the European bison: King of the forest or refugee in a marginal habitat? Ecography 2011, 35, 519–529. [Google Scholar] [CrossRef]
- Smit, C.; Ruifrok, J.L.; van Klink, R.; Olff, H. Rewilding with large herbivores: The importance of grazing refuges for sapling establishment and wood-pasture formation. Biol. Conserv. 2015, 182, 134–142. [Google Scholar] [CrossRef]
- Hofman-Kaminska, E.; Kowalczyk, R. Farm crops depredation by European bison (Bison bonasus) in the vicinity of forest habitats in northeastern Poland. Environ. Manag. 2012, 50, 530–541. [Google Scholar] [CrossRef] [PubMed]
- Van Dierendonck, M.C.; Wallis de Vries, M.F. Ungulate reintroduction: Experiences with the Takhi or Przewalski Horse (Equus ferus przewalskii) in Mongolia. Conserv. Biol. 1996, 10, 728–740. [Google Scholar] [CrossRef]
- Jackson, J. The Red-Cockaded Woodpecker Recovery Program: Professional Obstacles to Co-Operation. In Species Recovery: Finding the Lessons, Improving the Process; Clark, T.W., Reading, R.P., Clarke, A.L., Eds.; Endangered Island Press: Washington, DC, USA, 1994; pp. 157–181. [Google Scholar]
- Boyd, L.; Bandi, N. Reintroduction of takhi, Equus ferus przewalskii, to Hustai National Park, Mongolia: Time budget and synchrony of activity pre- and post release. Appl. Anim. Behav. Sci. 2002, 78, 87–102. [Google Scholar] [CrossRef]
- King, S.R.B. Home range and habitat use of free-ranging Przewalski horses at Hustai National Park, Mongolia. Appl. Anim. Behav. Sci. 2002, 78, 103–113. [Google Scholar] [CrossRef]
- Hughes, F.M.R.; Stroh, P.A.; Adams, W.M.; Kirby, K.J.; Mountford, J.O.; Warrington, S. Monitoring and evaluating large-scale, ′open-ended′ habitat creation projects: A journey rather than a destination. J. Nat. Conserv. 2011, 19, 245–253. [Google Scholar] [CrossRef]
- Ramos, A.; Petit, O.; Longour, P.; Pasquaretta, C.; Sueur, C. Space use and movement patterns in a semi-free-ranging herd of European bison (Bison bonasus). PLoS ONE 2016, 11, e0147404. [Google Scholar] [CrossRef]
- Cromsigt, J.P.G.M.; Kemp, Y.J.M.; Rodriguez, E.; Kivit, H. Rewilding Europe’s large grazer community: How functionally diverse are the diets of European bison, cattle, and horses? Restor. Ecol. 2017, 26, 891–899. [Google Scholar] [CrossRef]
- Deinet, S.; Ieronymidou, C.; McRae, L.; Burfield, I.J.; Foppen, R.P.; Collen, B.; Böhm, M. Wildlife Comeback in Europe: The Recovery of Selected Mammal and Bird Species; Final report to Rewilding Europe by ZSL, BirdLife International and the European Bird Census Council; ZSL: London, UK, 2013. [Google Scholar]
- Bergmann, G.T.; Craine, J.M.; Robeson, M.S., II; Fierer, N. Seasonal Shifts in Diet and Gut Microbiota of the American bison (Bison bison). PLoS ONE 2015, 10, e0142409. [Google Scholar] [CrossRef]
- Corbane, C.; Lang, S.; Pipkins, K.; Alleaume, S.; Deshayes, M.; GarcíaMillán, V.E.; Strasser, T.; Vanden Borre, J.; Toon, S.; Michael, F. Remote sensing for mapping natural habitats and their conservation status—New opportunities and challenges. Int. J. Appl. Earth Obs. 2015, 37, 7–16. [Google Scholar] [CrossRef]
- Gómez, C.; White, J.C.; Wulder, M.A. Optical remotely sensed time series data for land cover classification: A review. ISPRS J. Photogramm. Remote Sens. 2016, 116, 55–72. [Google Scholar] [CrossRef]
- Joshi, N.; Baumann, M.; Ehammer, A.; Fensholt, R.; Grogan, K.; Hostert, P.; Jepsen, M.R.; Kuemmerle, T.; Meyfroidt, P.; Mitchard, E.T.; et al. A review of the application of optical and radar remote sensing data fusion to land use mapping and monitoring. Remote Sens. 2016, 8, 70. [Google Scholar] [CrossRef]
- Turner, W.; Spector, S.; Gardiner, N.; Fladeland, M.; Sterling, E.; Steininger, M. Remote sensing for biodiversity science and conservation. Trends Ecol. Evol. 2003, 18, 306–314. [Google Scholar] [CrossRef]
- Förster, M.; Frick, A.; Walentowski, H.; Kleinschmit, B. Approaches to utilising QuickBird data for the monitoring of NATURA 2000 habitats. Community Ecol. 2008, 9, 155–168. [Google Scholar] [CrossRef]
- Förster, M.; Schmidt, T.; Schuster, C.; Kleinschmit, B. Multi-temporal detection of grassland vegetation with RapidEye imagery and a spectral-temporal library. In Proceedings of the 2012 Geoscience and Remote Sensing Symposium (IGARSS), Munich, Germany, 10 July 2012; pp. 4930–4933. [Google Scholar]
- Schuster, C.; Schmidt, T.; Conrad, C.; Kleinschmit, B.; Förster, M. Grassland habitat mapping by intra-annual time series analysis–Comparison of RapidEye and TerraSAR-X satellite data. Int. J. Appl. Earth Obs. 2015, 34, 25–34. [Google Scholar] [CrossRef]
- Laliberte, A.S.; Rango, A.; Havstad, K.M.; Paris, J.F.; Beck, R.F.; McNeely, R.; Gonzalez, A.L. Object-oriented image analysis for mapping shrub encroachment from 1937 to 2003 in southern New Mexico. Remote Sens Environ. 2004, 93, 198–210. [Google Scholar] [CrossRef]
- Selkowitz, D.J. A comparison of multi-spectral, multi-angular, and multi-temporal remote sensing datasets for fractional shrub canopy mapping in Arctic Alaska. Remote Sens Environ. 2010, 114, 1338–1352. [Google Scholar] [CrossRef]
- Coppin, P.R.; Bauer, M.E. Digital change detection in forest ecosystems with remote sensing imagery. Remote Sens. Rev. 1996, 13, 207–234. [Google Scholar] [CrossRef]
- Key, T.; Warner, T.A.; McGraw, J.B.; Fajvan, M.A. A comparison of multispectral and multitemporal information in high spatial resolution imagery for classification of individual tree species in a temperate hardwood forest. Remote Sens. Environ. 2001, 75, 100–112. [Google Scholar] [CrossRef]
- Kim, M.; Madden, M.; Warner, T.A. Forest type mapping using object-specific texture measures from multispectral Ikonos imagery. Photogramm. Eng. Remote Sens. 2009, 75, 819–829. [Google Scholar] [CrossRef]
- Feilhauer, H.; Thonfeld, F.; Faude, U.; He, K.S.; Rocchini, D.; Schmidtlein, S. Assessing floristic composition with multispectral sensors—A comparison based on monotemporal and multiseasonal field spectra. Int. J. Appl. Earth Obs. 2013, 21, 218–229. [Google Scholar] [CrossRef]
- Feilhauer, H.; Dahlke, C.; Doktor, D.; Lausch, A.; Schmidtlein, S.; Schulz, G.; Stenzel, S. Mapping the local variability of Natura 2000 habitats with remote sensing. Appl. Veg. Sci. 2014, 17, 765–779. [Google Scholar] [CrossRef]
- Kampichler, C.; Wieland, R.; Calmé, S.; Weissenberger, H.; Arriaga-Weiss, S. Classification in conservation biology: A comparison of five machine-learning methods. Ecol. Inf. 2010, 5, 441–450. [Google Scholar] [CrossRef]
- Mountrakis, G.; Im, J.; Ogole, C. Support vector machines in remote sensing: A review. ISPRS J. Photogramm. Remote Sens. 2011, 66, 247–259. [Google Scholar] [CrossRef]
- Belgiu, M.; Drăguţ, L. Random forest in remote sensing: A review of applications and future directions. ISPRS J. Photogramm. Remote Sens. 2016, 114, 24–31. [Google Scholar] [CrossRef]
- Richter, R.; Schläpfer, D. Atmospheric/Topographic Correction for Satellite Imagery; ATCOR-2/3 User Guide, Version 8.3. 1, February 2014; DLR-German Aerospace Center: Wessling, Germany; ReSe Appl. Schläpfer Langeggweg 3: Wil, Switzerland, 2013. [Google Scholar]
- Scheffler, D.; Hollstein, A.; Diedrich, H.; Segl, K.; Hostert, P. AROSICS: An automated and robust open-source image co-registration software for multi-sensor satellite data. Remote Sens. 2017, 9, 676. [Google Scholar] [CrossRef]
- Neumann, C.; Weiss, G.; Schmidtlein, S.; Itzerott, S.; Lausch, A.; Doktor, D.; Brell, M. Gradient-Based Assessment of Habitat Quality for Spectral Ecosystem Monitoring. Remote Sens. 2015, 7, 2871–2898. [Google Scholar] [CrossRef]
- Neumann, C.; Itzerott, S.; Weiss, G.; Kleinschmit, B.; Schmidtlein, S. Mapping multiple plant species abundance patterns—A multiobjective optimization procedure for combining reflectance spectroscopy and species ordination. Ecol. Inf. 2016, 36, 61–76. [Google Scholar] [CrossRef]
- Neumann, C.; Weiss, G.; Itzerott, S.; Kuehling, M.; Fuerstenow, J.; Luft, L.; Nitschke, P. Entwicklung und Erprobung eines innovativen, naturschutzfachlichen Monitoringverfahrens auf der Basis von Fernerkundungsdaten am Beispiel der Doeberitzer Heide, Brandenburg; GFZ German Research Centre: Potsdam, Germany, 2013. [Google Scholar]
- Clements, F.E. Plant Succession: An Analysis of the Development of Vegetation; Carnegie Institution of Washington: Washington, DC, USA, 1916. [Google Scholar]
- Whittaker, R.H. Classification of natural communities. Bot. Rev. 1962, 28, 1–239. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Cutler, D.R.; Edwards, T.C.; Beard, K.H.; Cutler, A.; Hess, K.T.; Gibson, J.; Lawler, J.J. Random forests for classification in ecology. Ecology 2007, 88, 2783–2792. [Google Scholar] [CrossRef]
- Lawrence, R.L.; Wood, S.D.; Sheley, R.L. Mapping invasive plants using hyperspectral imagery and Breiman Cutler classifications (Random Forest). Remote Sens. Environ. 2006, 100, 356–362. [Google Scholar] [CrossRef]
- Immitzer, M.; Atzberger, C.; Koukal, T. Tree species classification with random forest using very high spatial resolution 8-band WorldView-2 satellite data. Remote Sens. 2012, 4, 2661–2693. [Google Scholar] [CrossRef]
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling, R package version 2.6-7; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Liaw, A.; Wiener, M. Classification and Regression by random Forest. R News 2002, 2, 18–22. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Olsen, F.W.; Hansen, R.M. Food relations of wild free-roaming horses to livestock and big game, Red Desert, Wyoming. J. Range Manag. 1977, 30, 17–20. [Google Scholar] [CrossRef]
- Krysl, L.J.; Hubbert, M.E.; Sowell, B.E.; Plumb, G.E.; Jewett, T.K.; Smith, M.A.; Waggoner, J.W. Horses and cattle grazing in the Wyoming Red Desert. I. Food habits and dietary overlap. J. Range Manag. 1984, 37, 72–76. [Google Scholar] [CrossRef]
- Berger, J. Wild Horses at the Great Basin; University of Chicago Press: Chicago, IL, USA, 1986. [Google Scholar]
- Hunt, W.F.; Hay, R.J.M.; Clark, D. Pasture species preferences by horses in New Zealand. In Proceedings of the XVI International Grassland Congress, Nice, France, 4–11 October 1989; p. 797. [Google Scholar]
- Kissell, R.J., Jr. Competitive Interactions among Bighorn Sheep, Feral Horses, and Mule Deer in Bighorn Canyon National Recreation Area and the Pryor Mountain Wild Horse Range. PhD Thesis, Montana State University, Bozeman, MT, USA, 1996. [Google Scholar]
- Fahnestock, J.T.; Detling, J.K. The influence of herbivory on plant cover and species composition in the Pryor Mountain Wild Horse Range, USA. Plant Ecol. 1999, 144, 145–157. [Google Scholar] [CrossRef]
- Feranec, R.S.; Hadly, E.A.; Paytan, A. Stable isotopes reveal seasonal competition for resources between late Pleistocene bison (Bison) and horse (Equus) from Rancho La Brea, southern California. Palaeogeogr. Palaeoecl. 2009, 271, 153–160. [Google Scholar] [CrossRef]
- Scasta, J.D.; Beck, J.L.; Angwin, C.J. Meta-analysis of diet composition and potential conflict of wild horses with livestock and wild ungulates on western rangelands of North America. Rangel. Ecol. Manag. 2016, 69, 310–318. [Google Scholar] [CrossRef]
- Van Dierendonck, M.C.; Bandi, N.; Batdorj, D.; Dugerlham, S.; Munkhtsog, B. Behavioural observations of reintroduced Takhi or Przewalski horses (Equus ferus przewalskii) in Mongolia. Appl. Anim. Behav. Sci. 1996, 50, 95–114. [Google Scholar] [CrossRef]
- Prins, H.H.T. Origins and development of grassland communities in northwestern Europe. In Grazing and Conservation Management; Wallis de Vries, M.F., van de Koppel, J., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1998; pp. 55–105. [Google Scholar]
- Moehlman, P.D. Equids: Zebras, Asses and Horses: Status Survey and Conservation Action Plan; IUCN/SCC Equid Specialist Group; IUCN (The World Conservation Union): Gland, Switzerland; Cambridge, UK, 2002. [Google Scholar]
- Dawson, M.; Lane, C.; Saunders, G. Proceedings of the National Feral Horse Management Workshop; Invasive Animals Cooperative Research Centre: Canberra, Australia, 2006. [Google Scholar]
- Hofmann, R.R. Evolutionary steps of ecophysiological adaptation and diversification of ruminants: A comparative view of their digestive system. Oecologia 1989, 45, 443–457. [Google Scholar] [CrossRef] [PubMed]
- Warmuth, V.; Eriksson, A.; Bower, M.A.; Barker, G.; Barrett, E.; Hanks, B.K.; Shuicheng, L.; Lomitashvili, D.; Ochir-Goryaeva, M.; Sizonov, G.V.; et al. Reconstructing the origin and spread of horse domestication in the Eurasian steppe. Proc. Natl. Acad. Sci. USA 2010, 109, 8202–8206. [Google Scholar] [CrossRef] [PubMed]
- Albon, S.D.; Langvatn, R. Plant phenology and the benefits of migration in a temperate ungulate. Oikos 1992, 65, 502–513. [Google Scholar] [CrossRef]
- Villalba, J.J.; Provenza, F.D.; Bryant, J.P. Consequences of the interaction between nutrients and plant secondary metabolites on herbivore selectivity: Benefits or detriments for plants? Oikos 2002, 97, 282–292. [Google Scholar] [CrossRef]
- Villalba, J.J.; Provenza, F.D.; Han, G.; Provenza, D. Experience influences diet mixing by herbivores: Implications for plant biochemical diversity. Oikos 2004, 107, 100–109. [Google Scholar] [CrossRef]
- Krasińska, M.; Krasiński, Z.A. European Bison—The Nature Monograph; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Waggoner, V.; Hinkes, M. Summer and fall browse utilization by an Alaskan bison herd. J. Wildl. Manag. 1986, 50, 322–324. [Google Scholar] [CrossRef]
- Larter, N.C.; Gates, C.C. Diet and habitat selection of wood bison in relation to seasonal changes in forage quantity and quality. Can. J. Zoo 1991, 69, 2677–2685. [Google Scholar] [CrossRef]
- Bocherens, H.; Hofman-Kamińska, E.; Drucker, D.G.; Schmölcke, U.; Kowalczyk, R. European bison as a refugee species? Evidence from isotopic data on early Holocene bison and other large herbivores in northern Europe. PLoS ONE 2015, 10, e0115090. [Google Scholar] [CrossRef]
- Kowalczyk, R. European bison-king of the forest, or meadows and river valleys? In European Bison Conservation in the Białowieża Forest. Threats and Prospects of the Population Development; Kowalczyk, R., et al., Eds.; Mammal Research Institute, Academy of Science: Bialowieza, Poland, 2010; pp. 123–134. [Google Scholar]
- Kuemmerle, T.; Perzanowski, K.; Chaskovskyy, O.; Ostapowicz, K.; Halada, L.; Bashta, A.T.; Kruhlov, I.; Hostert, P.; Waller, D.M.; Radeloff, V.C. European bison habitat in the Carpathian Mountains. Biol. Conserv. 2010, 143, 908–916. [Google Scholar] [CrossRef]
- Cromsigt, J.P.G.M.; Kerley, G.I.H.; Kowalczyk, R. The difficulty of using species distribution modeling for the conservation of refugee species-the example of European bison. Divers. Distrib. 2012, 18, 1253–1257. [Google Scholar] [CrossRef]
- Zielke, L.; Wrage-Mönnig, N.; Müller, J. Seasonal preferences in diet selection of semi-free ranging European bison (Bison bonasus). Eur. Bison Conserv. Newsl. 2017, 10, 61–70. [Google Scholar]
- Bakker, J.; Van Wieren, S.E. The Impact of Grazing on Plant Communities. In Grazing and Conservation Management; Wallis De Vries, M.F., Bakker, J.P., Van Wieren, S.E., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1998; pp. 137–184. [Google Scholar]
- Janis, C.M. The evolutionary strategy of the equidae and the origins of rumen and cecal digestion. Evolution 1976, 30, 757–774. [Google Scholar] [CrossRef] [PubMed]
- Duncan, P.; Foose, T.J.; Gordon, I.J.; Gakahu, C.G.; Lloyd, M. Comparative nutrient extraction from forages by grazing bovids and equids: A test of the nutritional model of equid/bovid competition and coexistence. Oecologia 1990, 84, 411–418. [Google Scholar] [CrossRef]
- Duncan, P. Horses and Grasses. The Nutritional Ecology of Equids and Their Impact on the Camargue; Springer: New York, NY, USA, 1992. [Google Scholar]
- Gordon, I.J. Vegetation community selection by ungulates on the Isle of Rhum. II. Vegetation community selection. J. Appl. Ecol. 1989, 26, 53–64. [Google Scholar] [CrossRef]
- Kurtén, B. Pleistocene Mammals of Europe; Weidenfeld & Nicolson: London, UK, 1968. [Google Scholar]
- MacFadden, B.J.; Cerling, T.E. Mammalian herbivore communities, ancient feeding ecology, and carbon isotopes: A 10 million-year sequence from the Neogene of Florida. J. Vertebr. Paleontol. 1996, 16, 103–115. [Google Scholar] [CrossRef]
- Koch, P.L.; Hoppe, K.A.; Webb, S.D. The isotopic ecology of late Pleistocene mammals in North America Part 1. Florida. Chem. Geol. 1998, 152, 119–138. [Google Scholar] [CrossRef]
- Hutchinson, G.E. Concluding Remarks. Cold Springs Harbor Symposium. Quantum Biol. 1958, 22, 415–427. [Google Scholar] [CrossRef]
- McKane, R.B.; Johnson, L.C.; Shaver, G.R.; Nadelhoffer, K.J.; Rastetter, E.B.; Fry, B.; Giblin, A.E.; Kielland, K.; Kwiatkowski, B.L.; Laundre, J.A.; et al. Resource-based niches provide a basis for plant species diversity and dominance in arctic tundra. Nature 2002, 415, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J. Diet shifts in moose due to predator avoidance. Oecologia 1983, 60, 185–189. [Google Scholar] [CrossRef]
- Reynolds, H.W.; Peden, D.G. Vegetation, bison diets, and snow cover. In Bison Ecology in Relation to Agricultural Development in the Slave River Lowlands, N.W.T. Introduction; Reynolds, H.W., Hawley, A.W.L., Eds.; Canadian Wildlife Service Occasional Paper; Food and Agriculture Organization: Rome, Italy, 1987; Volume 63, pp. 39–44. [Google Scholar]
- Rominger, E.M.; Oldemeyer, J.L. Early-winter diet of woodland caribou in relation to snow accumulation Selkirk Mountains British-Columbia, Canada. Can. J. Zool. 1990, 68, 2691–2694. [Google Scholar] [CrossRef]
- Pearson, S.M.; Turner, M.G.; Wallace, L.L.; Romme, W.H. Winter habitat use by large ungulates following fire in Northern Yellowstone National Park. Ecol. Appl. 1995, 5, 744–755. [Google Scholar] [CrossRef]
- Bailey, D.W.; Gross, J.E.; Laca, E.A.; Rittenhouse, L.R.; Coughenour, M.B.; Swift, D.M.; Sims, P.L. Mechanisms that result in large herbivore grazing distribution patterns. J. Range Manag. 1996, 49, 386–400. [Google Scholar] [CrossRef]
- Dussault, C.; Ouellet, J.P.; Courtois, R.; Huot, J.; Breton, L.; Jolicoeur, H. Linking moose habitat selection to limiting factors. Ecography 2005, 28, 619–628. [Google Scholar] [CrossRef]
- Fortin, D.; Morales, J.M.; Boyce, M.S. Elk winter foraging at fine scale in Yellowstone National Park. Oecologia 2005, 145, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Hampson, B.A.; Morton, J.M.; Mills, P.C.; Trotter, M.G.; Lamb, D.W.; Pollitt, C.C. Monitoring distances travelled by horses using GPS tracking collars. Aust. Vet. J. 2010, 88, 176–181. [Google Scholar] [CrossRef]
- Hennig, J.D.; Beck, J.L.; Scasta, J.D. Spatial ecology observations from feral horses equipped with global positioning system transmitters. Hum.-Wildl. Interact. 2018, 12, 9. [Google Scholar]
- Leverkus, S.E.; Fuhlendorf, S.D.; Geertsema, M.; Allred, B.W.; Gregory, M.; Bevington, A.R.; Engle, D.M.; Scasta, J.D. Resource selection of free-ranging horses influenced by fire in northern Canada. Hum.-Wildl. Interact. 2018, 12, 10. [Google Scholar]
- King, S.R.B.; Schoenecker, K.A. Comparison of methods to examine diet of feral horses from noninvasively collected fecal samples. Rangel. Ecol. Manag. 2019. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Percentage Occurrence of Selected Plant Species in Observation Events | Description | Category |
|---|---|---|
| 95%–100% | highest food preference | 3 |
| 50%–95% | high to moderate preference | 2 |
| 5%–50% | moderate to rare preference | 1 |
| 0%–5% | rejected | 0 |
| Reference | Predicted Habitat | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Poplar | Alder | White Willow | Oak | Robinia (Black Locust) | Lime | Downy Birch | Birch | Haw-Thorn | Pine | Grey Willow | Ash Maple | PA [%] | ||
| Poplar | 352 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 99.72 | |
| Alder | 0 | 2890 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 99.93 | |
| White willow | 0 | 0 | 19 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 100.00 | |
| Oak | 0 | 0 | 0 | 858 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 99.77 | |
| Robinia (black locust) | 2 | 0 | 0 | 0 | 899 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 99.78 | |
| Lime | 0 | 0 | 0 | 0 | 0 | 333 | 0 | 0 | 0 | 0 | 0 | 0 | 100.00 | |
| Downy birch | 0 | 0 | 0 | 0 | 0 | 0 | 431 | 6 | 0 | 0 | 0 | 0 | 98.63 | |
| Birch | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 623 | 0 | 0 | 0 | 0 | 100.00 | |
| Hawthorn | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 283 | 0 | 0 | 0 | 100.00 | |
| Pine | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 436 | 0 | 0 | 100.00 | |
| Grey willow | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 148 | 0 | 98.67 | |
| Ash maple | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 23 | 100.00 | |
| UA [%] | 99.44 | 100.0 | 100.0 | 99.77 | 99.78 | 100.0 | 100.0 | 98.58 | 100.0 | 100.0 | 100.0 | 100.0 | ||
| Reference | Predicted Habitat | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pine | Heath | Open Sand | Broom | Xeric gr. a | Crypto-Gams | Fresh Mead. b | Wet Molinia | Wet Carex | Reed | Water | Bushgrass c | Wet Juncus | Brush | PA [%] | |
| Pine | 163 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 100.0 |
| Heath | 2 | 89 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 96.74 |
| Open sand | 0 | 0 | 14 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 93.34 |
| Broom | 0 | 0 | 0 | 206 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 100.00 |
| Xeric gr. a | 0 | 0 | 0 | 0 | 489 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 100.00 |
| Cryptogams | 0 | 0 | 0 | 0 | 0 | 128 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 100.00 |
| Fresh mead. b | 0 | 0 | 0 | 0 | 0 | 0 | 775 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 100.00 |
| Wet Molinia | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 450 | 0 | 0 | 0 | 0 | 0 | 0 | 100.00 |
| Wet Carex | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 697 | 0 | 0 | 0 | 0 | 0 | 100.00 |
| Reed | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 84 | 0 | 0 | 0 | 0 | 100.00 |
| Water | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 42 | 0 | 0 | 0 | 100.00 |
| Bushgrass c | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 382 | 0 | 0 | 99.74 |
| Wet Juncus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 206 | 0 | 100.00 |
| Brush | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 525 | 100.00 |
| UA [%] | 98.79 | 100.0 | 100.0 | 99.52 | 99.80 | 99.22 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zielke, L.; Wrage-Mönnig, N.; Müller, J.; Neumann, C. Implications of Spatial Habitat Diversity on Diet Selection of European Bison and Przewalski’s Horses in a Rewilding Area. Diversity 2019, 11, 63. https://doi.org/10.3390/d11040063
Zielke L, Wrage-Mönnig N, Müller J, Neumann C. Implications of Spatial Habitat Diversity on Diet Selection of European Bison and Przewalski’s Horses in a Rewilding Area. Diversity. 2019; 11(4):63. https://doi.org/10.3390/d11040063
Chicago/Turabian StyleZielke, Luisa, Nicole Wrage-Mönnig, Jürgen Müller, and Carsten Neumann. 2019. "Implications of Spatial Habitat Diversity on Diet Selection of European Bison and Przewalski’s Horses in a Rewilding Area" Diversity 11, no. 4: 63. https://doi.org/10.3390/d11040063
APA StyleZielke, L., Wrage-Mönnig, N., Müller, J., & Neumann, C. (2019). Implications of Spatial Habitat Diversity on Diet Selection of European Bison and Przewalski’s Horses in a Rewilding Area. Diversity, 11(4), 63. https://doi.org/10.3390/d11040063