Limnology and Aquatic Microbial Ecology of Byers Peninsula: A Main Freshwater Biodiversity Hotspot in Maritime Antarctica


Abstract
1. Introduction
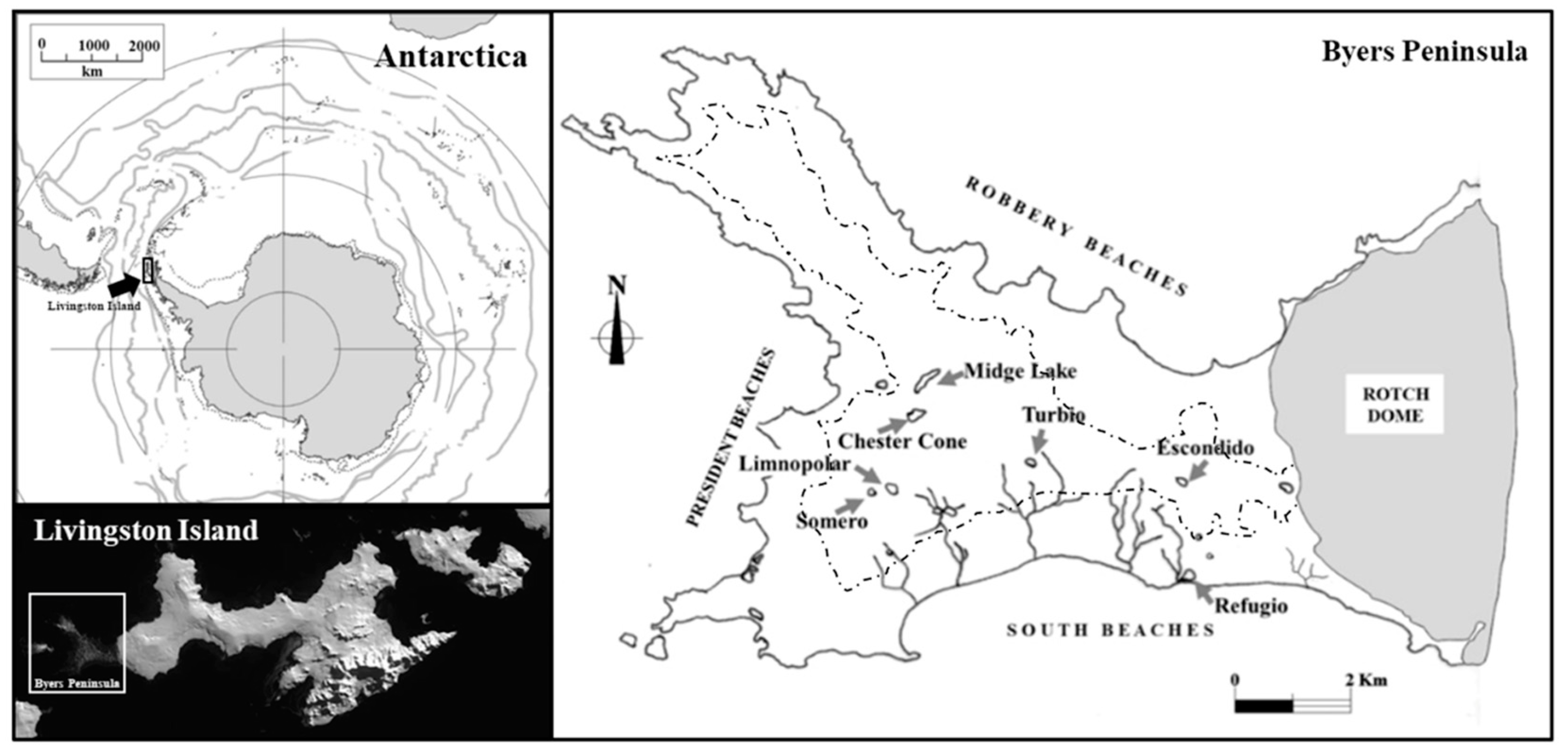
2. Landscape and Weather
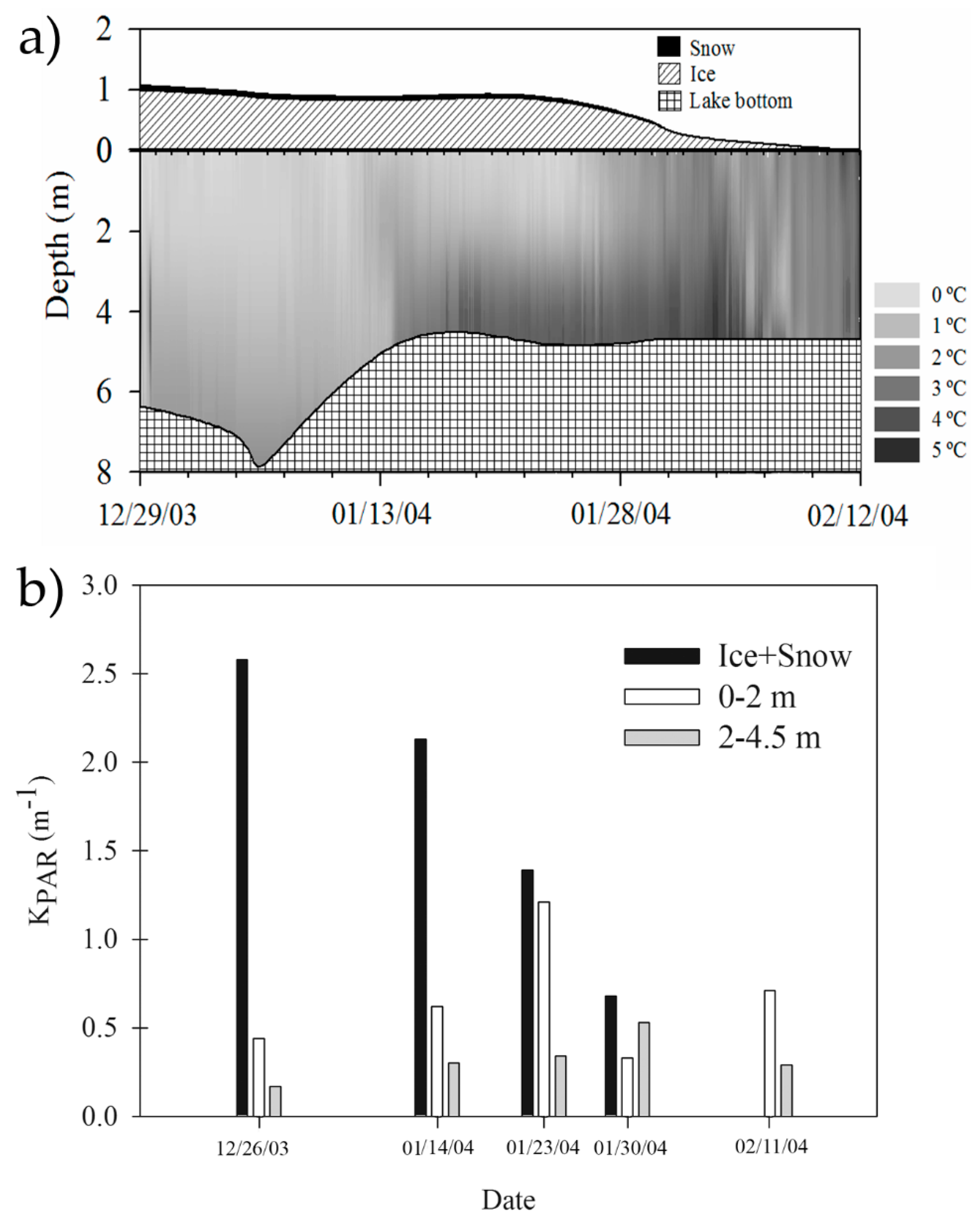
3. Hydrology, Nutrients Sources, and Environmental Gradients
4. Virioplankton
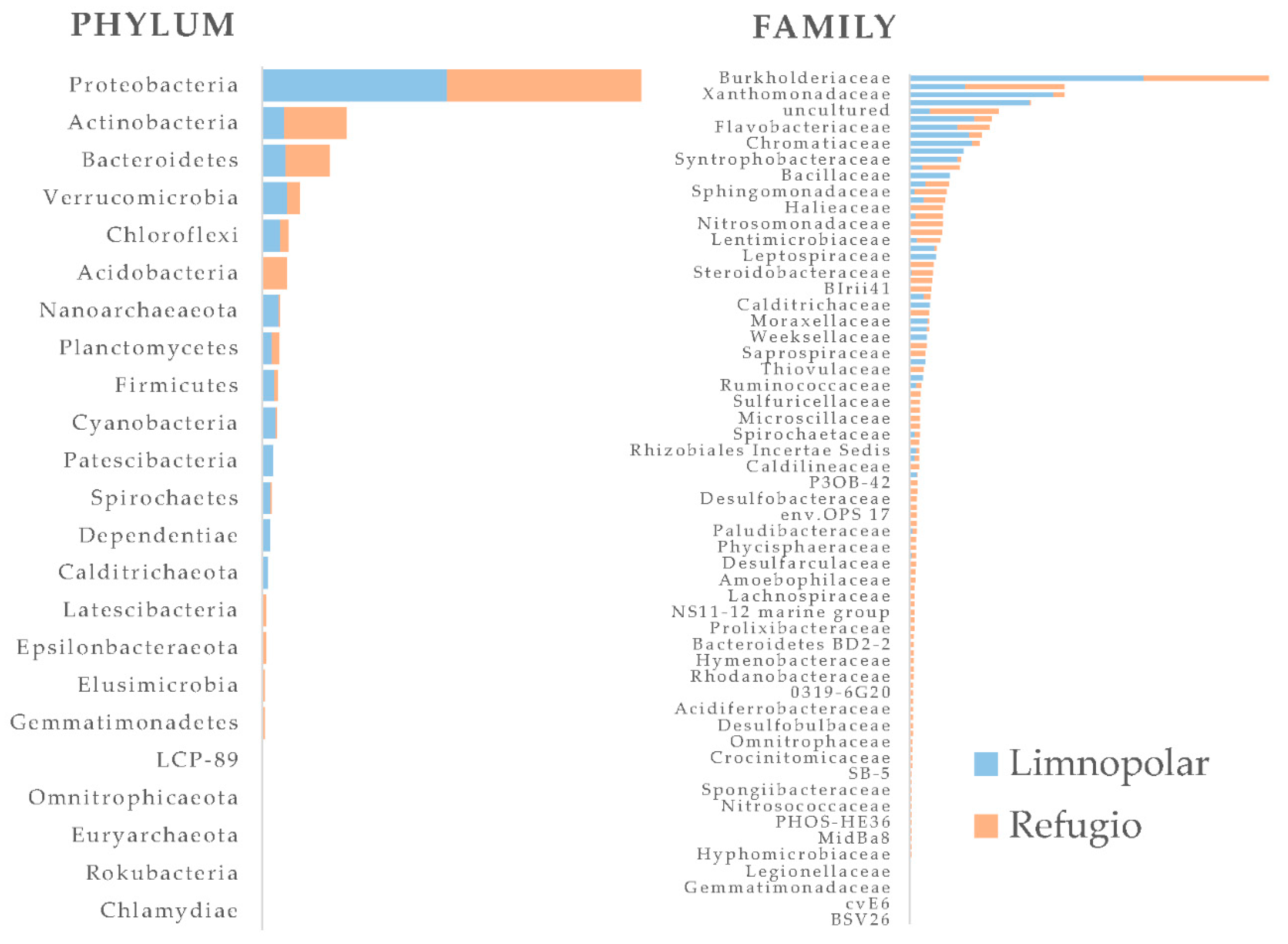
5. Prokaryotes in Plankton and Lacustrine Sediments
6. Phytoplankton
7. Protozoans and Metazoans
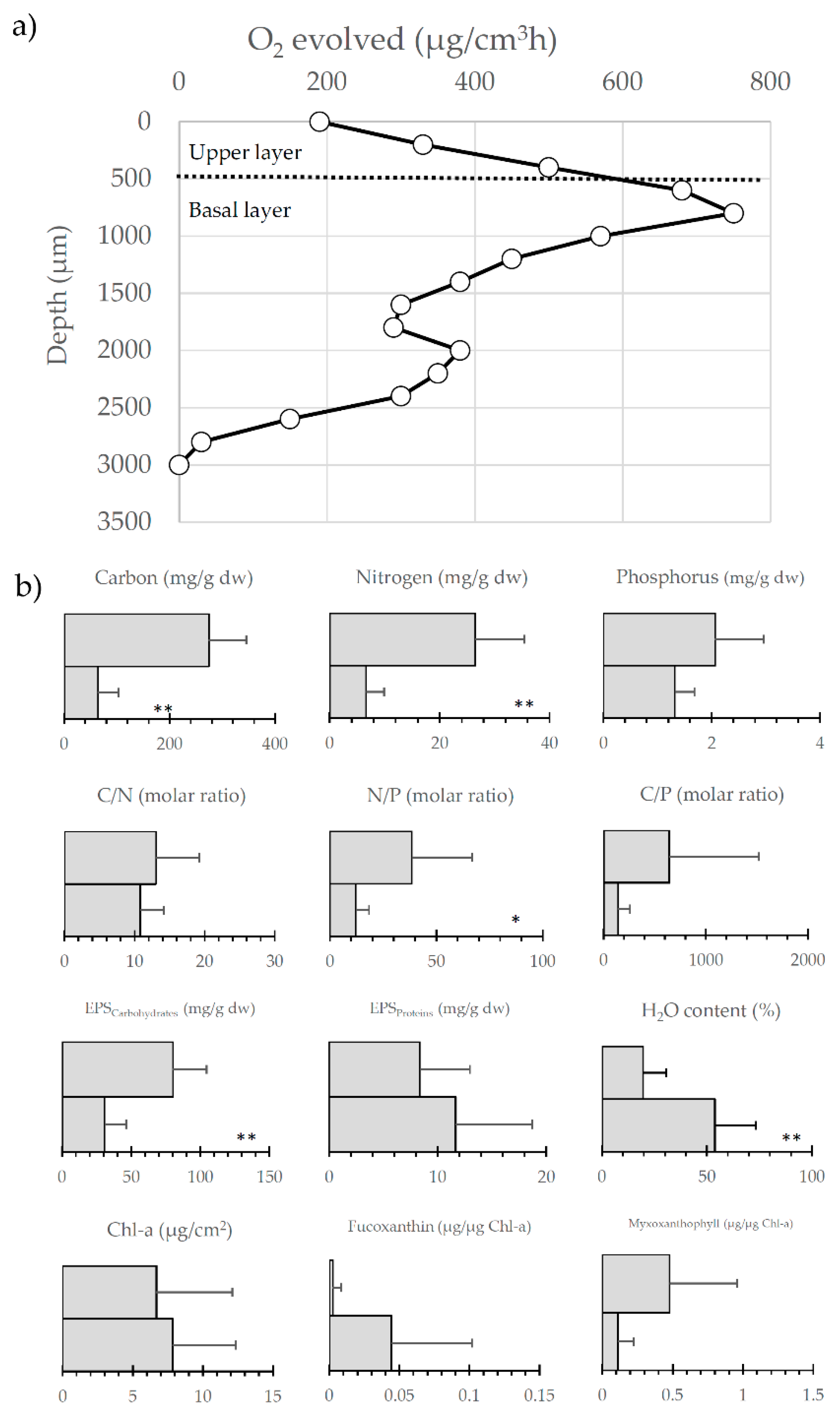
8. Microbial Mats and Autotrophic Biofilms: Function and Diversity
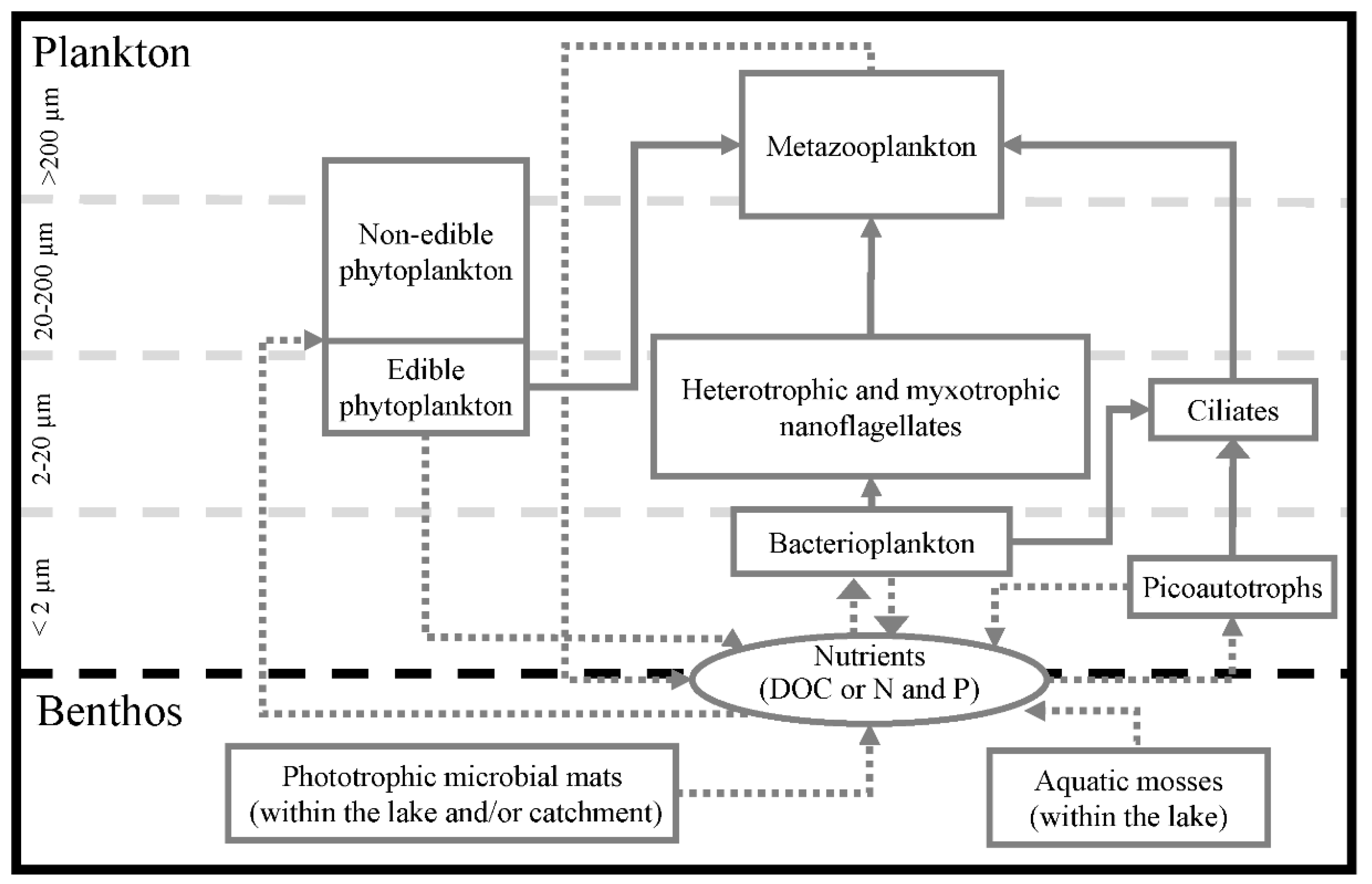
9. Aquatic Food-Web Assembly
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hughes, K.A.; Cowan, D.A.; Wilmotte, A. Protection of Antarctic microbial communities—‘Out of sight, out of mind’. Front. Microbiol. 2015, 6, 151. [Google Scholar] [CrossRef] [PubMed]
- Camacho, A.; Rochera, C.; Villaescusa, J.A.; Velázquez, D.; Toro, M.; Rico, E.; Fernández-Valiente, E.; Justel, A.; Bañón, M.; Quesada, A. Maritime Antarctic lakes as sentinels of climate change. Int. J. Des. Nat. Ecodyn. 2012, 7, 239–250. [Google Scholar] [CrossRef]
- Camacho, A. Planktonic microbial assemblages and the potential effects of metazooplankton predation on the food web of lakes from the maritime Antarctica and sub-Antarctic islands. Rev. Environ. Sci. Biotechnol. 2006, 5, 167–185. [Google Scholar] [CrossRef]
- Mink, S.; López-Martínez, J.; Maestro, A.; Garrote, J.; Ortega, J.A.; Serrano, E.; Durán, J.J.; Schmid, T. Insights into deglaciation of the largest ice-free area in the South Shetland Islands (Antarctica) from quantitative analysis of the drainage system. Geomorphology 2014, 225, 4–24. [Google Scholar] [CrossRef]
- Priscu, J.C.; Fritsen, C.H.; Adams, E.E.; Giovannoni, S.J.; Paerl, H.W.; McKay, C.P.; Doran, P.T.; Gordon, D.A.; Lanoil, B.D.; Pinckney, J.L. Perennial Antarctic Lake Ice: An Oasis for Life in a Polar Desert. Science 1998, 208, 2095–2098. [Google Scholar] [CrossRef] [PubMed]
- Roberts, E.C.; Laybourn-Parry, J. Mixotrophic cryptophytes and their predators in the Dry Valley lakes of Antarctica. Freshw. Biol. 1999, 41, 737–746. [Google Scholar] [CrossRef]
- Karr, E.A.; Sattley, W.M.; Jung, D.O.; Madigan, M.T.; Achenbach, L.A. Remarkable diversity of phototrophic purple bacteria in a permanently frozen Antarctic lake. Appl. Environ. Microbiol. 2003, 69, 4910–4914. [Google Scholar] [CrossRef]
- Bowman, J.P.; McCammon, S.A.; Rea, S.M.; McMeekin, T.A. The microbial composition of three limnologically disparate hypersaline Antarctic lakes. FEMS Microbiol. Lett. 2006, 183, 81–88. [Google Scholar] [CrossRef]
- Yau, S.; Lauro, F.M.; DeMaere, M.Z.; Brown, M.V.; Thomas, T.; Raftery, M.J.; Andrews-Pfannkoch, C.; Lewis, M.; Hoffman, J.M.; Gibson, J.A.; et al. Virophage control of antarctic algal host–virus dynamics. Proc. Natl. Acad. Sci. USA 2011, 108, 6163–6168. [Google Scholar] [CrossRef]
- Sabbe, K.; Verleyen, E.; Hodgson, D.A.; Vanhoutte, K.; Vyverman, W. Benthic diatom flora of freshwater and saline lakes in the Larsemann Hills and Rauer Islands, East-Antarctica. Antarct. Sci. 2003, 15, 227–248. [Google Scholar] [CrossRef]
- Taton, A.; Grubisic, S.; Balthasart, P.; Hodgson, D.A.; Laybourn-Parry, J.; Wilmotte, A. Biogeographical distribution and ecological ranges of benthic cyanobacteria in East Antarctic lakes. FEMS Microbiol. Ecol. 2006, 57, 272–289. [Google Scholar] [CrossRef] [PubMed]
- Laybourn-Parry, J.; Quayle, W.C.; Henshaw, T.; Ruddell, A.; Marchant, H.J. Life on the edge: The plankton and chemistry of Beaver Lake, an ultraoligotrophic epishelf lake, Antarctica. Freshw. Biol. 2001, 46, 1205–1217. [Google Scholar] [CrossRef]
- Cremer, H.; Gore, D.; Hultzsch, N.; Melles, M.; Wagner, B. The diatom flora and limnology of lakes in the Amery Oasis, East Antarctica. Polar Biol. 2004, 27, 513–531. [Google Scholar] [CrossRef]
- Pearce, D.A. The structure and stability of the bacterioplankton community in Antarctic freshwater lakes, subject to extremely rapid environmental change. FEMS Microbiol. Ecol. 2005, 53, 61–72. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Vinocur, A.; Maidana, N.I. Spatial and temporal variations in moss-inhabiting summer diatom communities from Potter Peninsula (King George Island, Antarctica). Polar Biol. 2010, 33, 443–455. [Google Scholar] [CrossRef]
- Callejas, C.; Gill, P.R.; Catalan, A.I.; Azziz, G.; Castro-Sowinski, S.; Batista, S. Phylotype diversity in a benthic cyanobacterial mat community on King George Island, maritime Antarctica. World J. Microbiol. Biotechnol. 2011, 27, 1507–1512. [Google Scholar] [CrossRef]
- Quesada, A.; Camacho, A.; Rochera, C.; Velàzquez, D. Byers Peninsula: A reference site for coastal, terrestrial and limnetic ecosystem studies in maritime Antarctica. Polar Sci. 2009, 3, 181–187. [Google Scholar] [CrossRef]
- Toro, M.; Camacho, A.; Rochera, C.; Rico, E.; Bañón, M.; Fernández-Valiente, E.; Marco, E.; Justel, A.; Avendaño, M.C.; Ariosa, Y.; et al. Limnological characteristics of the freshwater ecosystems of Byers Peninsula, Livingston Island, in maritime Antarctica. Polar Biol. 2007, 30, 635–649. [Google Scholar] [CrossRef]
- Fernández-Valiente, E.; Camacho, A.; Rochera, C.; Rico, E.; Vincent, W.F.; Quesada, A. Community structure and physiological characterization of microbial mats in Byers Peninsula, Livingston Island (SouthShetland Islands, Antarctica). FEMS Microbiol. Ecol. 2007, 59, 377–385. [Google Scholar] [CrossRef]
- Benayas, L.; Pertierra, L.; Tejedo, P.; Lara, F.; Bermudez, O.; Hughes, K.A.; Quesada, A. A review of scientific research trends within ASPA No. 126 Byers Peninsula, South Shetland Islands, Antarctica. Antarct. Sci. 2013, 25, 128–145. [Google Scholar] [CrossRef]
- Hathway, B.; Lomas, S.A. The Upper Jurassic-Lower Cretaceous Byers Group, South Shetland Islands, Antarctica: Revised stratigraphy and regional correlations. Cretac. Res. 1998, 19, 43–67. [Google Scholar] [CrossRef]
- Navas, A.; Soto, J.; López-Martínez, J. Radionuclides in soils of Byers Peninsula, South Shetland Islands, Western Antarctica. Appl. Radiat. Isot. 2005, 62, 809–816. [Google Scholar] [CrossRef] [PubMed]
- López-Martínez, J.; Serrano, E.; Schmid, T.; Mink, S.; Linés, C. Periglacial processes and landforms in the South Shetland Islands (northern Antarctic Peninsula region). Geomorphology 2012, 155–156, 62–79. [Google Scholar] [CrossRef]
- Moura, P.A.; Francelino, M.R.; Schaefer, C.E.G.R.; Simas, F.N.B.; de Mendoça, B.A.F. Distribution and characterization of soils and landform relationships in Byers Peninsula, Livingston Island, Maritime Antarctica. Geomorphology 2012, 155–156, 45–54. [Google Scholar] [CrossRef]
- Santamans, A.C.; Boluda, R.; Picazo, A.; Gil, C.; Ramos-Miras, J.; Tejedo, P.; Pertierra, L.R.; Benayas, J.; Camacho, A. Soil features in rookeries of Antarctic penguins reveal sea to land biotransport of chemical pollutants. PLoS ONE 2017, 12, e0181901. [Google Scholar] [CrossRef] [PubMed]
- Emslie, S.D.; Baumann, K.; van Tuinen, M. Late Holocene occupation of Gentoo Penguins (Pygoscelis papua) on Byers Peninsula, Livingston Island, Antarctica. Polar Biol. 2010, 34, 283–290. [Google Scholar] [CrossRef]
- Bañón, M.; Justel, A.; Velázquez, D.; Quesada, A. Regional weather survey on Byers Peninsula, Livingston Island, South Shetland Islands, Antarctica. Antarct. Sci. 2013, 25, 146–156. [Google Scholar] [CrossRef]
- Van Lipzig, N.P.M.; King, J.C.; Lachlan-Cope, T.A.; Van den Broeke, M.R. Precipitation, sublimation and snow drift in the Antarctic Peninsula regionfrom a regional atmospheric model. J. Geophys. Res. 2004, 109, D24106. [Google Scholar] [CrossRef]
- Björck, S.; Olsson, S.; Ellis-Evans, C.; Håkansson, H.; Humlum, O.; De Lirio, J.M. Late Holocene palaeoclimatic records from lake sediments on James Ross Island, Antarctica. Palaeogeogr. Palaeocl. 1996, 121, 195–220. [Google Scholar] [CrossRef]
- Oliva, M.; Hrbacek, F.; Ruiz-Fernández, J.; de Pablo, M.A.; Vieira, G.; Ramos, M.; Antoniades, D. Active layer dynamics in three topographically distinct lake catchments in Byers Peninsula (Livingston Island, Antarctica). Catena 2017, 149, 548–559. [Google Scholar] [CrossRef]
- Rochera, C.; Toro, M.; Rico, E.; Fernández-Valiente, E.; Villaescusa, J.A.; Picazo, A.; Quesada, A.; Camacho, A. Structure of planktonic microbial communities along a trophic gradient in lakes of Byers Peninsula. Antarct. Sci. 2013, 25, 277–287. [Google Scholar] [CrossRef]
- Villaescusa, J.A.; Casamayor, E.O.; Rochera, C.; Velázquez, D.; Chicote, A.; Quesada, A.; Camacho, A. A close link between bacterial community composition and environmental heterogeneity in maritime Antarctic lakes. Int. Microbiol. 2010, 13, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Picazo, A.; Rochera, C.; Villaescusa, J.A.; Miralles-Lorenzo, J.; Velázquez, D.; Quesada, A.; Camacho, A. Bacterioplankton community composition along environmental gradients in lakes from Byers Peninsula (Maritime Antarctica) as determined by Next-Generation Sequencing. Front. Microbiol. 2019, 10, 908. [Google Scholar] [CrossRef] [PubMed]
- Navas, A.; López-Martínez, J.; Casas, J.; Machín, J.; Durán, J.J.; Serrano, E.; Cuchi, J.-A.; Mink, S. Soil characteristics on varying lithological substrates in the South Shetland Islands, maritime Antarctica. Geoderma 2008, 144, 123–139. [Google Scholar] [CrossRef]
- Lyons, W.B.; Welch, K.A.; Welch, S.A.; Camacho, A.; Rochera, C.; Michaud, L.; de Wit, R.; Carey, A.E. Geochemistry of streams from Byers Peninsula, Livingston Island. Antarct. Sci. 2013, 25, 181–190. [Google Scholar] [CrossRef]
- Cabrerizo, A.; Dachs, J.; Barceló, D.; Jones, K.C. Influence of organic matter content and human activities on the occurrence of organic pollutants in Antarctic soils, lichens, grass, and mosses. Environ. Sci. Technol. 2012, 46, 1396–1405. [Google Scholar] [CrossRef]
- Otero, X.L.; Fernández, S.; de Pablo Hernandez, M.A.; Nizoli, E.C.; Quesada, A. Plant communities as a key factor in biogeochemical processes involving micronutrients (Fe, Mn, Co, and Cu) in Antarctic soils (Byers Peninsula, maritime Antarctica). Geoderma 2013, 195–196, 145–154. [Google Scholar] [CrossRef]
- Villaescusa, J.A.; Jørgensen, S.E.; Rochera, C.; Velázquez, D.; Quesada, A.; Camacho, A. Carbon dynamics modelization and biological community sensitivity to temperature in an oligotrophic freshwater Antarctic lake. Ecol. Model. 2016, 319, 21–30. [Google Scholar] [CrossRef]
- Fassnacht, S.R.; Toro, M.; Meiman, P.J.; Whitt, Z.C. The effect of aeolian deposition on the surface roughness of melting snow, Byers Peninsula, Antarctica. Hydrol. Process. 2010, 24, 2007–2013. [Google Scholar] [CrossRef]
- Moorhead, D.L. Mesoscale dynamics of ephemeral wetlands in the Antarctic Dry Valleys: Implications to production and distribution of organic matter. Ecosystems 2007, 10, 87–95. [Google Scholar] [CrossRef]
- Rochera, C.; Justel, A.; Fernández-Valiente, E.; Bañón, M.; Rico, E.; Toro, M.; Camacho, A.; Quesada, A. Interannual meteorological variability and its effects on a lake from maritime Antarctica. Polar Biol. 2010, 33, 1615–1628. [Google Scholar] [CrossRef]
- López-Bueno, A.; Tamames, J.; Velázquez, D.; Moya, A.; Quesada, A.; Alcamí, A. High diversity of the viral community from an Antarctic lake. Science 2009, 326, 858–861. [Google Scholar] [CrossRef] [PubMed]
- López-Bueno, A.; Rastrojo, A.; Peiró, R.; Arenas, M.; Alcamí, A. Ecological connectivity shapes quasispecies structure of RNA viruses in an Antarctic lake. Mol. Ecol. 2015, 24, 4812–4825. [Google Scholar] [CrossRef]
- Rochera, C.; Quesada, A.; Toro, M.; Rico, E.; Camacho, A. Plankton assembly in an ultra-oligotrophic Antarctic lake over the summer transition from the ice-cover to ice-free period: A size spectra approach. Polar Sci. 2017, 11, 72–82. [Google Scholar] [CrossRef]
- Madan, N.J.; Marshall, W.A.; Laybourn-Parry, J. Virus and microbial loop dynamics over an annual cycle in three contrasting Antarctic lakes. Freshw. Biol. 2005, 50, 1291–1300. [Google Scholar] [CrossRef]
- Schiaffino, M.R.; Unrein, F.; Gasol, J.M.; Massana, R.; Balague, V.; Izaguirre, I. Bacterial community structure in a latitudinal gradient of lakes: The roles of spatial versus environmental factors. Freshw. Biol. 2011, 56, 1973–1991. [Google Scholar] [CrossRef]
- Villaescusa, J.A.; Rochera, C.; Velázquez, D.; Rico, E.; Quesada, A.; Camacho, A. Bacterioplankton summer dynamics in a maritime Antarctic lake. Limnetica 2013, 32, 253–268. [Google Scholar]
- Papale, M.; Rizzo, C.; Villescusa, J.A.; Rochera, C.; Camacho, A.; Michaud, L.; Lo Giudice, A. Prokaryotic assemblages in the maritime Antarctic Lake Limnopolar (Byers Peninsula, South Shetland Islands). Extremophiles 2017, 21, 947–961. [Google Scholar] [CrossRef]
- Hahn, M.W.; Koll, U.; Jezberová, J.; Camacho, A. Global phylogeography of pelagic Polynucleobacter bacteria: Restricted geographic distribution of subgroups, isolation by distance and influence of climate. Environ. Microbiol. 2015, 17, 829–840. [Google Scholar] [CrossRef]
- Gugliandolo, C.; Michaud, L.; Lo Giudice, A.; Lentini, V.; Rochera, C.; Camacho, A.; Luciana Maugeri, T. Prokaryotic community in lacustrine sediments of Byers Peninsula (Livingston Island, Maritime Antarctica). Microb. Ecol. 2016, 71, 387–400. [Google Scholar] [CrossRef]
- Villaescusa, J.A.; Casamayor, E.O.; Rochera, C.; Quesada, A.; Michaud, L.; Camacho, A. Heterogeneous vertical structure of the bacterioplankton community in a non-stratified Antarctic lake. Antarct. Sci. 2013, 5, 229–238. [Google Scholar] [CrossRef]
- Yilmaz, P.; Parfrey, L.W.; Yarza, P.; Gerken, J.; Pruesse, E.; Quast, C.; Schweer, T.; Peplies, J.; Ludwig, W.; Glöckner, F.O. The SILVA and ‘‘All-species Living Tree Project (LTP)’’ taxonomic frameworks. Nucleic Acids Res. 2014, 42, D643–D648. [Google Scholar] [CrossRef]
- Pearce, D.A.; Galand, P.E. Microbial biodiversity and biogeography. In Polar Lakes and Rivers, Limnology of Arctic and Antarctic Aquatic Ecosystems, 1st ed.; Vincent, W., Laybourn-Parry, J., Eds.; Oxford University Press: New York, NY, USA, 2008; pp. 213–230. [Google Scholar] [CrossRef]
- Valdespino-Castillo, P.M.; Cerqueda-García, D.; Espinosa, A.C.; Batista, S.; Merino-Ibarra, M.; Tas, N.; Alcántara-Hernández, R.J.; Falcón, L.I. Microbial distribution and turnover in Antarctic microbial mats highlight the relevance of heterotrophic bacteria in low-nutrient environments. FEMS Microbiol. Ecol. 2018, 94, fiy129. [Google Scholar] [CrossRef]
- Michaud, L.; Caruso, C.; Mangano, S.; Interdonato, F.; Bruni, V.; Lo Giudice, A. Predominance of Flavobacterium, Pseudomonas, and Polaromonas within the prokaryotic community of freshwater shallow lakes in the northern Victoria Land, East Antarctica. FEMS Microbiol. Ecol. 2012, 82, 391–404. [Google Scholar] [CrossRef]
- Schiaffino, M.R.; Lara, E.; Fernández, L.D.; Balagué, V.; Singer, D.; Seppey, C.C.W.; Massana, R.; Izaguirre, I. Microbial eukaryote communities exhibit robust biogeographical patterns along a gradient of Patagonian and Antarctic lakes. Environ. Microbiol. 2016, 18, 5249–5264. [Google Scholar] [CrossRef]
- Simek, K.; Bobkova, J.; Macek, M.; Nedoma, J.; Psenner, R. Ciliate grazing on picoplankton in a eutrophic reservoir during the summer phytoplankton maximum: A study at the species and community level. Limnol. Oceanogr. 1995, 40, 1077–1090. [Google Scholar] [CrossRef]
- Petz, W.; Valbonesi, A.; Schiftner, U.; Quesada, A.; Ellis-Evans, J.C. Ciliate biogeography in Antarctic and Arctic freshwater ecosystems: Endemism or global distribution of species? FEMS Microb. Ecol. 2007, 59, 396–408. [Google Scholar] [CrossRef]
- Lynn, D.H. The Ciliated Protozoa: Characterization, Classification and Guide to the Literature, 3rd ed.; Springer: Dordrecht, The Netherlands, 2008. [Google Scholar]
- Roberts, E.C.; Priscu, J.C.; Laybourn-Parry, J. Microplankton dynamics in a perennially ice-covered Antarctic lake—Lake Hoare. Freshw. Biol. 2004, 49, 853–869. [Google Scholar] [CrossRef]
- Bayly, I.A.E.; Gibson, J.A.E.; Wagner, B.; Swadling, K.M. Taxonomy, ecology and zoogeography of two East Antarctic freshwater calanoid copepod species: Boeckella poppei and Gladioferens antarcticus. Antarct. Sci. 2003, 15, 439–448. [Google Scholar] [CrossRef]
- Menu-Marque, S.; Morrone, J.J.; De Mitrovich, C.L. Distributional patterns of the South American species of Boeckella (Copepoda: Centropagidae): A track analysis. J. Crustac. Biol. 2000, 20, 262–272. [Google Scholar] [CrossRef][Green Version]
- Gibson, J.A.E.; Bayly, I.A.E. New insights into the origins of crustaceans of Antarctic lakes. Antarct. Sci. 2007, 19, 157–164. [Google Scholar] [CrossRef]
- Pla-Rabes, S.; Toro, M.; Van De Vijver, B.; Rochera, C.; Villaescusa, J.A.; Camacho, A.; Quesada, A. Stability and endemicity of benthic diatom assemblages from different substrates in a maritime stream on Byers Peninsula, Livingston Island, Antarctica: The role of climate variability. Antarct. Sci. 2013, 25, 254–269. [Google Scholar] [CrossRef]
- Rochera, C.; Villaescusa, J.A.; Velázquez, D.; Fernández-Valiente, E.; Quesada, A.; Camacho, A. Vertical structure of bi-layered microbial mats from Byers Peninsula, Maritime Antarctica. Antarct. Sci. 2013, 25, 270–276. [Google Scholar] [CrossRef]
- Rochera, C.; Fernández-Valiente, E.; Van de Vijver, B.; Rico, E.; Toro, M.; Vincent, W.F.; Quesada, A.; Camacho, A. Community structure and photosynthetic activity of benthic biofilms from a waterfall in the maritime Antarctica. Polar Biol. 2013, 36, 1709–1722. [Google Scholar] [CrossRef]
- Camacho, A.; Rochera, C.; Hennebelle, R.; Ferrari, C.; Quesada, A. Total mercury and methyl-mercury contents and accumulation in polar microbial mats. Sci. Total Environ. 2015, 509–510, 145–153. [Google Scholar] [CrossRef]
- Velázquez, D.; Jungblut, A.D.; Rochera, C.; Rico, E.; Camacho, A.; Quesada, A. Trophic interactions in microbial mats on Byers Peninsula, maritime Antarctica. Polar Biol. 2017, 40, 1115–1126. [Google Scholar] [CrossRef]
- Almela, P.; Velázquez, D.; Rico, E.; Justel, A.; Quesada, A. Carbon pathways through the food web of a microbial mat from Byers Peninsula, Antarctica. Front. Microbiol. 2019, 10, 628. [Google Scholar] [CrossRef]
- Kopalová, K.; Van De Vijver, B. Structure and ecology of freshwater benthic diatom communities from Byers Peninsula, Livingston Island, South Shetland Islands. Antarct. Sci. 2013, 25, 239–253. [Google Scholar] [CrossRef]
- Van de Vijver, B.; Beyens, L. Freshwater diatoms from Ile de la Possession (Crozet Archipelago, Sub-Antarctica): An ecological assessment. Polar Biol. 1999, 22, 178–188. [Google Scholar] [CrossRef]
- Lotter, A.F.; Bigler, C. Do diatoms in the Swiss Alps reflect the length of ice-cover? Aquat. Sci. 2000, 62, 125–141. [Google Scholar] [CrossRef]
- Van De Vijver, B.; Agius, J.T.; Gibson, J.A.E.; Quesada, A. An unusual spine-bearing Pinnularia species from the Antarctic Livingston Island (South Shetland Islands). Diatom Res. 2009, 24, 431–441. [Google Scholar] [CrossRef]
- Van De Vijver, B.; Zidarova, R. Five new taxa in the genus Pinnularia sectio Distantes (Bacillariophyta) from Livingston Island (South Shetland Islands). Phytotaxa 2011, 24, 39–50. [Google Scholar] [CrossRef]
- Zidarova, R.; Kopalová, K.; Van De Vijver, B. The genus Pinnularia (Bacillariophyta) excluding the section Distantes on Livingston Island (South Shetland Islands) with the description of twelve new taxa. Phytotaxa 2012, 44, 11–37. [Google Scholar] [CrossRef]
- Velázquez, D.; López-Bueno, A.; Aguirre de Cárcer, D.; de los Ríos, A.; Alcamí, A.; Quesada, A. Ecosystem function decays by fungal outbreaks in Antarctic microbial mats. Sci. Rep. 2016, 6, 22954. [Google Scholar] [CrossRef]
- Velázquez, D.; Rochera, C.; Camacho, A.; Quesada, A. Temperature effects on carbon and nitrogen metabolism in some Maritime Antarctic freshwater phototrophic communities. Polar Biol. 2011, 34, 1045–1055. [Google Scholar] [CrossRef]
- Krebs, C.J. Ecology: The Experimental Analysis of Distribution and Abundance, 5th ed.; Benjamin Cummings: Menlo Park, CA, USA, 2001. [Google Scholar]
- Butler, H.G.; Edworthy, M.G.; Ellis-Evans, J.C. Temporal plankton dynamics in an oligotrophic maritime Antarctic lake. Freshw. Biol. 2000, 43, 215–230. [Google Scholar] [CrossRef]
- Pociecha, A.; Dumont, H.J. Life cycle of Boeckella poppei Mrazek and Branchinecta gaini Daday (King George Island, South Shetlands). Polar Biol. 2008, 31, 245–248. [Google Scholar] [CrossRef]
- Reissig, M.; Trochine, C.; Queimaliños, C.; Balseiro, E.; Modenutti, B. Impact of fish introduction on planktonic food webs in lakes of the Patagonian Plateau. Biol Conserv. 2006, 132, 437–447. [Google Scholar] [CrossRef]
- Butler, H.G.; Atkinson, A.; Gordon, M. Omnivory and predation impact of the calanoid copepod Boeckella poppei in a maritime Antarctic lake. Polar Biol. 2005, 28, 815–821. [Google Scholar] [CrossRef]
- Gasol, J.M.; Pedros-Aliò, C.; Vaqué, D. Regulation of bacterial assemblages in oligotrophic plankton systems: Results from experimental and empirical approaches. Anton. Leeuw. J. Microb. 2002, 81, 435–452. [Google Scholar] [CrossRef]
- Reynolds, C.S. Phytoplankton periodicity: The interactions of form, function and environmental variability. Freshw. Biol. 1984, 14, 111–142. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lake | Chester Cone | Midge Lake | Limnopolar | Escondido | Turbio | Somero | Refugio |
|---|---|---|---|---|---|---|---|
| X-UTM | 597,500 | 597,700 | 597,100 | 599,475 | 598,000 | 596,800 | 602,200 |
| Y-UTM | 3,053,550 | 3,054,150 | 3,052,200 | 3,052,650 | 3,051,800 | 3,052,150 | 3,050,550 |
| Maximum depth (m) | 5.0 | 8.2 | 5.5 | 4.5 | 7.8 | 0.5 | 0.5 |
| Lake Surface (Km2) | 0.039 | 0.054 | 0.023 | 0.022 | 0.021 | 0.011 | 0.016 |
| Catchment size (Km2) | 0.09 | 0.27 | 0.58 | 0.08 | 0.58 | 0.06 | 0.12 |
| Conductivity (µS/cm) | 58 ± 6 | 68 ± 12 | 65 ± 9 | 60 ± 7 | 60 ± 17 | 81 ± 14 | 148 ± 31 |
| Total phosphorus (µM) | 0.28 ± 0.21 | 0.24 ± 0.17 | 0.39 ± 0.18 | 0.28 ± 0.04 | 1.27 ± 0.92 | 1.02 ± 0.49 | 16.84 ± 6.16 |
| Bacterioplankton (cells/mL) × 106 | 0.99 ± 0.39 | 0.98 ± 0.36 | 1.39 ± 0.86 | 1.50 ± 0.41 | 2.26 ± 2.19 | 3.27 ± 1.61 | 8.20 ± 2.68 |
| Chlorophyll a (µg/L) | 0.08 ± 0.01 | 0.14 ± 0.06 | 0.10 ± 0.07 | 0.38 ± 0.17 | 0.37 ± 0.19 | 0.94 ± 0.66 | 23.41 ± 9.98 |
| Group | Dominant Taxa | |
|---|---|---|
| DNA Virioplankton | Phycodnaviridae, Caudovirales, Mimiviridae | |
| RNA Virioplankton | Dicistroviridae, Secoviridae, Marnaviridae, Iflaviridae, Potyviridae, Picornaviridae and Tombusviridae | |
| Heterotrophic bacterioplankton (Families) | Burkholderiaceae, Sporichthyaceae, Flavobacteriaceae, Chromatiaceae, Chitinophagaceae, Xanthomonadaceae, Sphingomonadaceae | |
| Autotrophic picoplankton | Picocyanobacteria, Prasinophytes | |
| Plastidic nanoflagellates (genera) | Ochromonas, Chromulina, Pseudokephyrion. | |
| Heterotrophic nanoflagellates (genera) | Spumella, Oikomonas | |
| Microphytoplankton (genera) | Pennate diatoms | Navicula, Nitzschia, Achnanthes, Fragilaria |
| Chlorophytes | Ankistrodesmus, Ankyra, Chlamydomonas | |
| Cryptophytes | Chroomonas | |
| Nanoplanktonic ciliates | Balanion planctonicum, Cyclidium glaucoma | |
| Rotifers (genera) | Notholca | |
| Copepods | Boeckella poppei | |
| Cladocerans | Macrothrix ciliata | |
| Anostraca | Branchinecta gainii | |
| Chironomids | Parochlus steinenii, Belgica antarctica | |
| Oligochaetes | Lumbricillus healyae | |
| Bryophytes | Drepanocladus longifolius | |
| Group | Dominant Taxa | |
|---|---|---|
| RNA Viruses | Bacillariornavirus, Picornavirales | |
| DNA Viruses | Circoviridae, Nanoviridae, Microviridae, Geminiviridae | |
| Bacteria | Alphaproteobacteria, Bacteroidetes, Cloroflexi, Actinobacteria | |
| Fungi | Ascomycota, Basidiomycta, Chytridiomycota | |
| Phototrophs | Cyanobacteria | Leptolyngbya, Oscillatoria, Phormidium, Porphyrosiphon, Nostoc |
| Diatoms | Fragilaria, Nitzschia, Gomphonema, Planothidium, Navicula | |
| Chlorophytes | Ulothrix, Klebsormidium, Chlamydomonas, Chloromonas | |
| Chrysophytes | Ochromonas | |
| Nematodes | Plectidae, Monhysteridae | |
| Tardigrades | Hypsibiidae, Macrobioitidae, Calohypsibiidae | |
| Rotifers | Philodinidae, Lecanidae, Lindiidae | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rochera, C.; Camacho, A. Limnology and Aquatic Microbial Ecology of Byers Peninsula: A Main Freshwater Biodiversity Hotspot in Maritime Antarctica. Diversity 2019, 11, 201. https://doi.org/10.3390/d11100201
Rochera C, Camacho A. Limnology and Aquatic Microbial Ecology of Byers Peninsula: A Main Freshwater Biodiversity Hotspot in Maritime Antarctica. Diversity. 2019; 11(10):201. https://doi.org/10.3390/d11100201
Chicago/Turabian StyleRochera, Carlos, and Antonio Camacho. 2019. "Limnology and Aquatic Microbial Ecology of Byers Peninsula: A Main Freshwater Biodiversity Hotspot in Maritime Antarctica" Diversity 11, no. 10: 201. https://doi.org/10.3390/d11100201
APA StyleRochera, C., & Camacho, A. (2019). Limnology and Aquatic Microbial Ecology of Byers Peninsula: A Main Freshwater Biodiversity Hotspot in Maritime Antarctica. Diversity, 11(10), 201. https://doi.org/10.3390/d11100201